Embed Size (px)
Citation preview

Eric A. Rickart1*, Lawrence R. Heaney2, and Danilo S. Balete2†
1Natural History Museum of Utah, 301 Wakara Way, Salt Lake City, UT 84108 USA2Field Museum of Natural History, 1400 S Lake Shore Drive, Chicago, IL 60605 USA
Mammals of Mt. Pinatubo, Luzon Island, Philippines:Extreme Resilience Following Catastrophic Disturbance
Keywords: bats, biogeography, disturbance tolerance, elevational gradients, endemism, faunal recovery, habitat recovery, rodents, volcanic eruption
The catastrophic eruption of Mt. Pinatubo in 1991 destroyed the forests that covered the peak and impacted the surrounding habitat over a broad area of central Luzon. Information on the mammal fauna of Mt. Pinatubo prior to the eruption is limited but documents a variety of native mammals. In 2011 and 2012, we surveyed mammals at localities along an elevational gradient on the eastern slope of the mountain where vegetation had been devastated by pyroclastic flows and subsequent lahars, and habitat reflected early stages of plant succession. We documented eight species of bats (five fruit bats and three insectivorous bats), seven species of small non-volant mammals (two non-native and five native rodents), and two native large mammals. Additional species of bats and non-volant mammals present in the vicinity of Mt. Pinatubo prior to the eruption or recently documented at other localities in the Zambales Mountains may be present in remnant forest habitat elsewhere on Pinatubo. Across five survey localities where habitat was in the early stages of regeneration, native species of small non-volant mammals were more widespread and much more abundant than were non-natives. The most abundant native species, Apomys sacobianus, may be endemic to Mt. Pinatubo. It is an extreme example of a “disturbance specialist” that thrives in a severely disturbed habitat. Results underscore the disturbance tolerance of many native small mammals of Luzon and reveal a resilience that is remarkable for a highly endemic insular fauna. Mt. Pinatubo presents opportunities for further studies on how natural disturbance has shaped the evolution of the Philippine biota and may influence its future conservation.
*Corresponding Author: [email protected] †Deceased 01 Jul 2017
INTRODUCTION Mt. Pinatubo, a volcanic peak in central Luzon Island, Philippines, was long considered dormant until it erupted explosively on 15 Jun 1991 in the second most powerful terrestrial volcanic eruption of the 20th century. The eruption covered the upper slopes of the mountain with massive amounts of ejecta in the form of falling ash and pyroclastic flows, destroying the summit and creating a
large caldera in its place, thereby reducing the height of the peak by 250 m. Subsequent heavy precipitation from seasonal typhoons and monsoons created a caldera lake and triggered destructive lahars that continued for many years following the eruption (Newhall et al. 1996).
Dormant for about 500 years preceding the 1991 eruption, Mt. Pinatubo and surrounding lowlands had supported old-growth forest. Although the lowlands of central Luzon were largely deforested during the 20th century (Heaney et al. 2016b), remnant mature dipterocarp forest remained
121
Philippine Journal of Science150 (S1): 121-133, Special Issue on BiodiversityISSN 0031 - 7683Date Received: 24 Sep 2020

in portions of Clark Air Base, and the upper portions of Pinatubo were heavily forested prior to the 1991 eruption (Punongbayan et al. 1996).
In 2006, 15 years after the eruption, an assessment of plant succession on Mt. Pinatubo revealed a predominance of non-native plants and the influence of elevation on species richness, diversity, and plant cover (Marler and del Moral 2011). Although there were no comprehensive surveys of the mammals of Mt. Pinatubo prior to the eruption, there are specimens housed at the United States National Museum of Natural History (USNM) from Clark Air Base and nearby localities at low elevations in Tarlac and Pampanga provinces. These include rodents reported from rice croplands (Barbehenn et al. 1973), and bats and rodents obtained by D.H. Johnson and others (records summarized in Heaney et al. 2010). However, there is no direct information on the mammals that may have been present at higher elevations on Pinatubo prior to its eruption.
Several previous studies have dealt with the disturbance ecology of the mammals of Luzon. These have utilized field surveys of small mammals across gradients of habitat disturbance ranging from relatively pristine old-growth forests to heavily disturbed human-modified landscapes. The principal results of these studies can be summarized as follows: although some native species are highly tolerant of disturbance, in general, diversity and abundance of native species decline with increasing levels of disturbance whereas the opposite is true for non-native species (Rickart et al. 2011, 2013; Heaney et al. 2016b; Reginaldo and Ong 2020).
Here, we present the results of a survey of the mammals of Mt. Pinatubo conducted in 2011 and 2012. This work was undertaken to determine the nature of the pioneering mammal community in early successional habitat two decades after the 1991 eruption. Based on disturbance gradient surveys elsewhere on Luzon (Rickart et al. 2011, 2013) we developed the following hypotheses to inform and direct the work on Mt. Pinatubo, which we test in this paper:
• Non-native small mammals are numerically dominant in the most severely disturbed habitat.
• A subset of native species that tolerate severe habitat disturbance coexists with non-native species.
• Native small mammals are resilient, moving into areas of regenerating habitat and ultimately displacing non-native species as native vegetation matures.
Of special interest to us was the status of Apomys sacobianus, a native mouse that was known only from the holotype captured on the lower slopes of Mt. Pinatubo in
1956. As discussed below, the presence of a potentially rare endemic species prior to the eruption raised the question of its possible extinction, given the frequent assumption that rare mammals on oceanic islands are intrinsically susceptible to extinction following significant ecological disturbance (Berglund et al. 2009; Walsh et al. 2012).
STUDY AREA AND METHODS
Location and GeologyMt. Pinatubo (15.13°N, 120.35°E; current elevation 1,486 m), is a dacitic-andesitic stratovolcano and caldera located in the Zambales Mountains of central Luzon on the tripoint boundary of Pampanga, Tarlac, and Zambales provinces (MGB 2010). It is one of several volcanoes along the Bataan Lineament formed by the eastward subduction of the Eurasian Plate beneath the Philippine Mobile Belt along the Manila Trench (Wolfe and Self 1983). The eruptive history of Pinatubo consists of two phases. “Ancestral” Pinatubo was a massive stratovolcano that may have had an elevation approaching 2,300 m. It has been dated at around 1.1 Ma and left no evidence that it erupted explosively. In contrast, “modern” Pinatubo had at least six periods of explosive eruption, each separated by longer periods of inactivity. Its earliest eruption occurred more than 30,000 BP and the most recent (prior to 1991) ca. 500 BP. The 1991 eruption was estimated to be one of the least powerful in the discernable geological record of Pinatubo (Newhall et al. 1996).
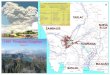
Survey Localities Our survey was conducted in 2011 and 2012 on the eastern slope of Mt. Pinatubo in Pampanga Province, between 300 and 1,100 m elevation (Figure 1). Survey localities were selected in consultation with the indigenous Aeta who manage the area as their ancestral domain. In 2011, we surveyed three localities in portions of the rolling hills (300–700 m) in Mabalacat, Pampanga Province near the boundary of Bamban, Tarlac Province, on the northeastern flank of Mt. Pinatubo north of the Sacobia River (Figure 2A). In 2012, we surveyed the upper eastern slope of Mt. Pinatubo (680–1,100 m), near the northern tributaries of Pasig-Potrero River in the western-most portion of Mabalacat, Pampanga Province. Localities were in areas of early-stage plant succession with sparse tree cover, as described below. Because of the steep slopes and loose lahar substrate, erosion was especially severe along riverbanks and gullies (Figure 2B).
Locality 1: 8.4 km N, 12.3 km E Mt. Pinatubo peak, 365 m elev., 15.20556o N, 120.46278o E; 27 Feb–04
Rickart et al.: Mammals of Mt. PinatuboPhilippine Journal of ScienceVol. 150 No. S1, Special Issue on Biodiversity
122

Mar 2011. This locality, selected as a base camp, was a nursery established by the Aeta of Sitio Burog in an attempt to reforest the surrounding area. Trees planted here included Eucalyptus (ca. 12 m tall), Leucaena, Swietenia, and particularly Gmelina, along with native Pterocarpus. Ground cover was dominated by the exotic shrub Lantana camara, grasses including cogon (Imperata cylindrica) and “talahib” (Saccharum spontaneum), and thick but isolated clumps of wild banana (Musa).
Climbing and erect bamboos (Dinochloa and Bambusa) were common along the steep stream banks. A narrow strip of riparian habitat along a stream included a variety of native palms (Areca, Arenga, Caryota, and Calamus) and other scattered trees up to 20 m tall (Afzelia, Alstonia, Artocarpus, Ficus, Garcinia, Hibiscus, Parkia, Premna, Macaranga, Mangifera, Myristica, Pterocarpus, and Terminalia). Within the remnant riparian vegetation, velvet bean (Mucuna), jade vine (Strongylodon), and
Figure 1. Map of Mt. Pinatubo showing the location of numbered survey localities along the eastern slope of the mountain. Contour intervals 400 m. Inset map shows the location of Mt. Pinatubo on Luzon.
Figure 2. Habitat features in areas surveyed on Mt. Pinatubo in 2011 and 2012. A) View downslope from a point on the northeastern flank of Mt. Pinatubo north of the Sacobia River illustrating the general nature of the regenerating habitat surrounding localities 1–3 in March 2011. B) View upstream along an upper tributary of the Pasig-Potrero River showing the eroding lahar and nature of the surrounding regenerating vegetation in March 2012; the camp at locality 4 was in the left-center of the image (a small tarp is barely visible) and locality 5 was on the steep slope above and beyond the camp.
Rickart et al.: Mammals of Mt. PinatuboPhilippine Journal of ScienceVol. 150 No. S1, Special Issue on Biodiversity
123

chestnut vine (Tetrastigma) were present but uncommon. Epiphytes, mainly ferns, were uncommon, and orchids and mosses were rare. Leaf litter was sparse, and there was no humus layer. Adjacent to this locality the Aeta had active taro farms, and sections of the steeper slopes were regularly burned to clear trails and facilitate hunting of wild pigs.
Locality 2: 7.0 km N, 11.7 km E Mt. Pinatubo peak, 490 m elev., 15.19222o N, 120.45722o E; 05–10 Mar 2011. Vegetation here was similar to that at Locality 1 but with a more extensive cover of wild banana and bamboo. It also had more interspersed trees of similar height (15–20 m) and composition as those at Locality 1, with the addition of Radermachera and Spathodea. Epiphytic ferns and orchids were present but uncommon. Leaf litter was largely absent and there was no humus layer. This locality bordered “kaingins” (slash-and-burn farming areas), some fallow and some newly cultivated by the Aeta, planted with root crops (cassava, sweet potato, taro, and yams), as well as other vegetables and fruit.
Locality 3: 6.2 km N, 10.7 km E Mt. Pinatubo peak, 670 m elev., 15.18611°N, 120.44972o E; 13–18 Mar 2011. Vegetation was similar to that found at Locality 2, characterized by extensive wild banana and bamboo cover but with more extensive riparian vegetation. Scattered trees, including some dipterocarps (Shorea) and nutmegs (Myristica), were 30–40 m tall with diameters at breast height (dbh) of 50–70 cm. Cultivated kaingins, like those Locality 2, also were present. Under the taller trees leaf litter had accumulated up to 3 cm.
Locality 4: 2.0 km N, 5.1 km E Mt. Pinatubo peak, 680 m elev., 15.14944o N, 120.39694o E; 13–16 Mar 2012. This locality, which served as our base camp in 2012, was along an upper tributary of the Pasig-Potrero River east of the Pinatubo caldera. The habitat here was in the early stage of forest development involving mainly two tree species: Parasponia rugosa, a widespread native species, and a common introduced species, Muntingia calabura. Figs, particularly Ficus minahassae, were common. Grasses (including Imperata, Saccharum, and Andropogon) – along with ferns, shrubs (Lantana), and vines (Mucuna and Dolichos) – covered exposed ridges and gaps between the narrow stands of trees. Musa was uncommon. Only limited mist netting for bats was conducted at this locality.
Locality 5: 2.5 km N, 5.4 km E Mt. Pinatubo peak, 960 m elev., 15.15333o N, 120.39972o E; 06–12 March 2012. Vegetation at this site consisted of abundant wild bananas, along with a sparse cover of figs (Ficus minahassae, F. nota, and F. hispida) and clumps of Saccharum. On exposed ridges and steeper slopes, Mallotus, Muntingia, and Parasponia were common, ca. 5–6 m tall (rarely reaching 10 m), and 15–50 cm dbh. Understory species
included saplings of taller trees and a few tree ferns (Cyathea). Grasses, ferns, and scrambling Lantana comprised the ground vegetation. Canopy vines were sparse and consisted mainly of Mikania and Mucuna. There was a thin (ca. 1–3 cm) layer of leaf litter in flat areas but with no humus layer. Abundant rocks were exposed on the ground. Fallen logs were rare but on slopes, erosion had exposed buried tree trunks charred in the 1991 eruption.
Locality 6: 2.8 km N, 5.3 km E Mt. Pinatubo peak, 1,080 m elev., 15.15611o N, 120.39570o E; 12–17 March 2012. The vegetation at this uppermost site resembled that at Locality 5, but Parasponia was less abundant and Muntingia appeared to be absent, and taller trees with greater dbh were more numerous. Ficus and Musa were abundant, as were Mikania and Mucuna vines. The area toward the highest point (ca. 1,100 m) was covered predominantly with Saccharum with scattered Ficus and Musa.
Field MethodsTo facilitate comparability of results, survey methods followed standardized techniques developed during previous surveys of Philippine small mammals (Heaney et al. 2013; Rickart et al. 1991, 2013; Balete et al. 2009; Alviola et al. 2011). Latitude and longitude (decimal degrees, based on the Luzon, Philippines datum), and elevation (m) of each locality were determined using a hand-held GPS (global positioning system) device. Locality coordinates were plotted on 1:50,000 topographic maps and straight-line N-S and E-W distances (in km) measured to the peak of Mt. Pinatubo (Figure 1). At each locality, we sampled within ± 50 m of the central elevation.
We used mist nets set in likely flyways to survey small fruit bats and insectivorous bats. Nets were tended continually during the early evening period of peak bat activity (dusk to 20:00 h), then every 30 min until 22:00h, and thereafter left open until dawn. We did removal trapping to survey small (< 500 g weight) non-volant mammals, using snap traps baited with either roasted coconut coated with peanut butter (“coconut bait”), or live earthworms collected locally. Most traps were set on the ground surface at burrow openings, along runways, beneath the ground cover, and in root tangles. Some coconut-baited traps were placed above the ground surface as “arboreal” traps. At each locality, traplines were set in all accessible microhabitats and operated for three consecutive days before moving them to a new site for another three-day period. This procedure was continued until all local microhabitats were sampled, after which we proceeded to a new locality. Traps were serviced twice daily to remove captured animals and replenish bait: the early morning after sunrise and the late afternoon before sundown (ca. 07:00h and 17:00h, respectively). Specimens recovered
Rickart et al.: Mammals of Mt. PinatuboPhilippine Journal of ScienceVol. 150 No. S1, Special Issue on Biodiversity
124

in the morning were considered nocturnal-crepuscular captures and those in the late afternoon as diurnal captures.
Approximately 40% of the bats, including individuals of all species documented, were prepared as voucher specimens and the remainder was released following identification. Some non-volant mammals damaged by scavengers were discarded after identification; all others (> 90%) were prepared as voucher specimens. Prior to their preservation, standard external measurements were recorded for each specimen, and a sample of muscle tissue was preserved for future genetic analyses. Specimens were preserved whole in fluid or prepared as complete skeletons. All specimens were deposited at the Field Museum of Natural History, Chicago (FMNH) for verification of field identification and further taxonomic studies. After completion of these studies, half of the specimens will be returned to the National Museum of the Philippines, Manila.
Mammal nomenclature follows Heaney et al. (2010, 2016b). Capture, handling, and collection of animals in the field were done under permits from the Philippine Department of Environment and Natural Resources (DENR) and were conducted in accordance with all relevant Philippine laws and regulations.
Data AnalysisIn assessing the elevational range of occurrence across the survey gradient, we inferred the presence of species at a given elevation if it was documented at higher and lower localities (Rickart et al. 1991). Our unit of sampling effort for non-volant small mammals was a trap-night (one trap set for 24 h) and for bats, a net-night (one mist net set for 24 h). To measure the adequacy or completeness of our survey for nonvolant small mammals, we used species-accumulations curves showing the cumulative number of species captured as a function of sampling effort (number of trap-nights). We plotted accumulation curves for each survey locality, and a total curve for all localities combined, starting at the lowest elevation and proceeding to the highest. We do not present any species accumulations curves for bats because of low sampling effort and disrupted netting schedules due to poor weather.
Relative abundance was measured as trapping (or netting) success. For bats, it was expressed as the number captured per net night. For non-volant small mammals, we calculated the number of captures per 100 trap-nights (percent trap success): the number of individuals captured multiplied by 100 and divided by the number of trap-nights. We used these measures to assess patterns of distribution and relative abundance of individual species, and native vs. non-native species groups. Bait preference of non-volant mammals was expressed as bait-specific
captures. Overall ground trapping success, as a proxy for relative abundance, was expressed as weighted (by bait type) and unweighted (equalized) percentages of captures. Weighted trap success was determined by dividing the total number of individuals captured by the total trap-nights at the locality, and the quotient multiplied by 100 to calculate the percent trap success. Unweighted trap success was computed by averaging the trap successes for the two bait types at each locality. In assessing the significance of the diel activity, expected capture frequencies were computed based on the duration of the diurnal (10 h) and nocturnal/crepuscular (14 h) activity periods.
We used the χ2 test to determine the significance of differences in abundance across survey localities and to evaluate patterns of diel activity and bait attractiveness. We conducted χ2 tests only on total samples of 20 or more individuals, with five or more individuals per “treatment” in at least 80% of the treatments (e.g. per elevation or bait type). For sample size less than 20, we determined significance with binomial tests where appropriate. We used Spearman’s coefficient of rank correlation (rs) to assess relationships of species richness and relative abundance with elevation.
RESULTS
BatsSurveys across six localities yielded 308 captures of fruit bats representing five species (Table 1). Cynopterus brachyotis, the most abundant species, accounted for nearly half (49%) of the captures, followed by Macroglossus minimus and Ptenochirus jagori. These three species were documented (or inferred to occur) at all sampling localities. Eonycteris spelaea and Rousettus amplexicaudatus were less abundant; the former was recorded at the three lowest localities, and the latter was captured at 365 m only. We captured 15 insectivorous bats representing three species – Hipposideros bicolor, H. diadema, and Myotis horsfieldii (Table 1). Data were not sufficient for more detailed assessments of distribution and abundance.
Non-volant Small MammalsOur total trapping effort consisted of 4,507 trap nights, 3,838 with coconut bait on the ground, 33 coconut “arboreal” trap-nights, and 636 earthworm trap-nights on the ground. In total, trapping yielded 329 captures representing seven rodent species (Table 2) – including five native (Apomys sacobianus, A. zambalensis, Bullimus luzonicus, Chrotomys mindorensis, and Rattus everetti) and two non-native species (Rattus exulans
Rickart et al.: Mammals of Mt. PinatuboPhilippine Journal of ScienceVol. 150 No. S1, Special Issue on Biodiversity
125

and R. tanezumi). The number of native species reached asymptotes at each of the five survey localities (Figure 3A) but at two localities (490 and 960 m), we infer the presence of an additional species (A. zambalensis) captured at adjacent upper and lower elevations (Table 2). Although further trapping would likely have added
A. zambalensis and B. luzonicus at localities where they were not recorded, it is unlikely that the total number of species would have increased with additional effort, as illustrated by the early asymptote for the entire survey transect (Figure 3B).
Table 1. Captures of bats at six localities on Mt. Pinatubo, Luzon Island, using mist nets. For fruit bats, captures per net night are shown in parentheses at localities with more than 10 net nights. Capture frequencies greater than expected from χ2 tests based on numbers of net nights at each locality are indicated by bold font. Number of species equals documented (plus inferred).
SpeciesLocality/elevation (m)
Total χ2 P1365
2490
3670
4680
5960
61,080
Fruit bats
Cynopterus brachyotis 18 (1.12) 14 (1.00) 48 (3.20) 4 38 (3.45) 29 151 69.22 < 0.001
Eonycteris spelaea 5 (0.31) 1 (0.07) 6 (0.40) 0 0 0 12
Macroglossus minimus 10 (0.62) 22 (1.57) 42 (2.80) 0a 3 (0.27) 12 89
Ptenochirus jagori 6 (0.38) 1 (0.07) 28 (1.87) 4 8 (0.73) 5 52
Rousettus amplexicaudatus 4 (0.25) 0 0 0 0 0 4
Insectivorous bats
Hipposideros bicolor 0 2 0 0 0 0 2
Hipposideros diadema 0 0 1 0 0 0 1
Myotis horsfieldii 12 0 0 0 0 0 12
Net-nights 16 14 15 3 11 4 63
Total fruit bats 43 38 124 8 49 46 308 102.1 < 0.001
Total insectivorous bats 12 2 1 0 0 0 15
No. of species 6 5 5 2 + 1 3 3 8aPresence inferred from occurrence at higher and lower elevations
Table 2. Occurrence of non-volant small mammals at five localities on Mt. Pinatubo, Luzon Island; values indicate the number caught at each locality (percent trap success in parentheses). Captures greater than expected from χ2 tests based on numbers of trap-nights at each locality are in bold. Number of species equals documented (plus inferred).
SpeciesLocality/elevation (m)
Total χ2 P1365
2490
3670
5960
61,080
Apomys sacobianus 46 (4.96) 40 (4.37) 4 (0.45) 54 (6.32) 82 (9.30) 226 (5.05) 71.40 < 0.001
Apomys zambalensis 3 (0.35) 0b 3 (0.34) 0b 2 (0.30) 8 (0.18)
Bullimus luzonicus 1 (0.35) 6 (0.73) 10 (1.12) 0 0 17 (0.38)
Chrotomys mindorensis 4 (0.43) 2 (0.24) 2 (0.26) 3 (0.35) 7 (0.79) 18 (0.40)
Rattus everetti 12 (1.29) 15 (1.64) 18 (2.01) 9 (1.05) 6 (0.68) 60 (1.34) 7.50 0.112
Rattus exulansa 0 2 (0.22) 2 (0.26) 0 0 4 (0.09)
Rattus tanezumia 12 (1.29) 2 (0.22) 0b 0b 1 (0.11) 15 (0.33)
Total captures (native species) 66 63 37 66 97 329 28.93 < 0.001
Total captures (non-native) 12 4 2 0 1 19
Total trap nights (ground) 927 915 895 855 882 4474
Total trap nights (arboreal) 9 6 0 3 15 33
No. of native species 5 4 + 1 5 3 + 1 4 5
No. of non-native species 1 2 1 + 1 0 + 1 1 2aNon-native species bPresence inferred from occurrence at higher and lower elevations
Rickart et al.: Mammals of Mt. PinatuboPhilippine Journal of ScienceVol. 150 No. S1, Special Issue on Biodiversity
126

The species richness of native non-volant mammals was nearly uniform across the survey gradient (Table 2). Four of the five native species either were documented or inferred to occur at all five of the localities. Species richness was not significantly correlated with elevation (rs = –0.62; P = 0.50). Trap success on the ground (Table 3) was not significantly correlated with elevation, either with coconut bait (rs = 0.200; P = 0.75) or earthworms (rs = 0.60; P = 0.28), nor were weighted or unweighted overall trap success (rs = 0.60; P = 0.28 and rs = 0.50; P = 0.39, respectively). Among native species, Apomys sacobianus was the most abundant at four out of five localities (Table 2) and was by far the most abundant overall with 226 captures (nearly 69% of the total native captures). Rattus everetti was the second most abundant, followed by Chrotomys mindorensis, Bullimus luzonicus, and Apomys zambalensis. Of the two non-native species, Rattus tanezumi was moderately common at the lowest elevation but uncommon or not documented elsewhere, and R. exulans was documented at two localities where it was uncommon (Table 2).
Overall trap success was significantly greater with earthworm compared to coconut bait (Table 3). Earthworm bait was significantly more effective than coconut in capturing Apomys sacobianus and Chrotomys mindorensis (Table 4). The remaining species did not exhibit significant bait preferences, although only Rattus everetti was caught in sufficient numbers for this to be biologically meaningful (i.e. revealing no significant difference in bait attractiveness rather than an artifact due to sample size). Capture data revealed that all species were overwhelmingly most active during the nocturnal/crepuscular period (Table 4). Arboreal trapping was very limited due to the general scarcity of suitable arboreal habitat, resulting in the capture of only a single Rattus tanezumi (Tables 3 and 4).
DISCUSSION
Historical Records and Adequacy of SamplingSpecimen records from low elevations around Mt. Pinatubo (summarized in Heaney et al. 2010) provide invaluable background information on the mammal fauna of the study area prior to the 1991 eruption. Several species of bats were obtained by D.H. Johnson in 1956, including four fruit bats – Acerodon jubatus and Pteropus vampyrus from Crow Valley in Tarlac Province, and Eonycteris spelaea and Ptenochirus jagori from Clark Air Base. The first two probably were collected at tree roosts, and the second two possibly from caves. Thirteen species of insectivorous bats obtained by Johnson include Hipposideros ater, H. diadema, H. obscurus, Rhinolophus arcuatus, R. inops, R. macrotus, R. philippinensis, R. subrufus, R. virgo, Miniopteris schreibersi, Myotis horsfieldii, Scotophilus kuhli, and Chaerophon plicata. Some of these (Hipposideros spp. and Rhinolophus spp.) were collected from day roosts in caves and tunnels along the Sacobia River, and others were collected on Clark Air Base, presumably from buildings and other structures (unpublished data from USNM specimen labels).
Two additional fruit bats, Haplonycteris fisheri and Otopteropus cartilagonodus, occur elsewhere within the Zambales region. They are seldom found outside forested habitats where they forage in the understory. Both are widespread on Luzon but they seldom occur together, which suggests segregation due to interspecific competition (Heaney et al. 2016b). Haplonycteris occurs on Mt. Natib where it was present below 1,000 m but uncommon (Rickart et al. 2013). Both species occur on Mt. Tapulao where Otopteropus was found across a broad elevational range and Haplonycteris was less common and found only at low to mid-elevations (Balete et al. 2009; unpublished FMNH specimen records). Given its
Figure 3. Species accumulation curves for native non-volant small mammals on Mt. Pinatubo A) at individual localities and B) for the entire transect, with the beginning of sampling at each locality starting with the lowest in elevation. Each symbol represents one day of sampling.
Rickart et al.: Mammals of Mt. PinatuboPhilippine Journal of ScienceVol. 150 No. S1, Special Issue on Biodiversity
127

occurrence on Mt. Natib and Mt. Tapulao, Haplonycteris was probably present on Mt. Pinatubo prior to the eruption. It might persist in areas of remnant forest nearby and if so may recolonize once adequate forest has developed on the mountain. Because of its limited distribution in the Zambales region, Otopteropus is perhaps less likely to have occurred on Mt. Pinatubo in the past.
All eight species of bats documented in our survey (Table 1) are widespread and common in the Philippines. However, four of these (C. brachyotis, M. minimus, R. amplexicaudatus, and H. obscurus) represent the first records from Pampanga Province (species distribution maps in Heaney et al. 2016b). Differences between the historical records and our survey results can be attributed
Table 3. Summary of overall capture frequencies by bait type and diel period of all native non-volant small mammals along an elevational gradient on Mt. Pinatubo. Captures greater than expected from χ2 tests of ground-traps with the two types of bait are in bold.
Locality/elevation (m)
Total1365
2490
3670
5960
61,080
Coconut bait (ground)
Trap nights 846 823 769 734 666 3838
Captures 53 59 28 39 56 236
Percent trap success 6.26 7.17 3.64 5.31 8.41 6.15
Earthworm bait (ground)
Trap nights 81 92 126 121 216 636
Captures 13 4 9 27 40 93
Percent trap success 16.05 4.35 7.14 22.31 18.52 14.62
Χ2 9.94 0.96 3.21 38.90 14.61 53.27
P 0.001 0.328 0.073 0.001 0.001 0.001
Overall trap success (%)
Weighted 7.12 6.88 4.13 7.72 11.00 7.35
Unweighted 11.16 5.76 5.39 13.81 13.54 10.38
Diel period
Diurnal 0 2 0 2 3 7
Nocturnal/crepuscular 66 61 37 64 94 322
Percent diurnal 0 3.17 0 3.03 3.09 2.13
Coconut bait (arboreal)
Trap nights 9 6 0 3 15 33
Captures 0 0 0 0 1 0
Percent trap success 6.67
Table 4. Summary of captures of native non-volant small mammals on Mt. Pinatubo by bait type, diel period, and trap position. Captures greater than expected are shown in bold.
Species Total trapped
Bait type Diel period Trap position
Coconut Worm Nocturnal/crepuscular Diurnal Ground
surfaceAbove ground
Apomys sacobianus 226 154 72*** 219*** 7 226 0
Apomys zambalensis 8 7 1 8b 0 8 0
Bullimus luzonicus 17 15 2 17b 0 17 0Chrotomys mindorensis 18 11 7b 17b 1 18 0Rattus everetti 60 49 11 60*** 0 60 0Rattus exulansa 4 4 0 4 0 4 0Rattus tanezumia 15 14 1 15b 0 14 1
aNon-native speciesbP < 0.05, from binomial distribution ***P < 0.001
Rickart et al.: Mammals of Mt. PinatuboPhilippine Journal of ScienceVol. 150 No. S1, Special Issue on Biodiversity
128

to methodology, notably our use of mist nets, which are very effective but were not widely used in the late 1950s when the principal methods were collecting by hand at roost sites or through the use of shotguns (Genoways et al. 2020). We also acknowledge that bat netting was limited at some of our survey localities due to logistical problems. Future surveys in areas of remnant forest and exploration of caves and other roosting sites may add many of the bats documented historically along with additional species.
Although we are confident that non-volant mammal trapping was sufficient to document the species present across our survey gradient (Figure 3B), additional species may occur elsewhere on Mt. Pinatubo, particularly in areas where the effects of the 1991 eruption may have been less severe. Specimen records predating the eruption include those for Tryphomys adustus from Clark Air Base and localities to the north in Tarlac Province (Heaney et al. 2016b). This poorly-known murid rodent is endemic to Luzon Island, with records from central and northern Luzon and from near sea level to 2,000 m in non-forested habitat ranging from grassy riparian vegetation to areas of second-growth adjacent to rice fields (Barbehenn et al. 1973; Heaney et al. 2016b). From these facts, it is possible that T. adustus persists in the vicinity of Mt. Pinatubo. Directed surveys for this species are clearly warranted.
Although our survey did not target large mammals (> 500 kg weight), we did gather pertinent information on their status. In 2011, wild pigs (Sus philippensis) were heard moving through thick stands of Saccharum adjacent to Locality 1. Subsequently, a group of Aeta hunters burned a section of this grassland and obtained a young adult female and a piglet. The Aeta reliably reported that deer (Rusa marianna) also were present on the mountain. Other species of large mammals with records from the Zambales Mountains are the arboreal native rodent Phloeomys pallidus, and the non-native carnivores Paradoxurus philippinensis and Viverra tangalunga (Balete et al. 2009; Heaney et al. 2010; Rickart et al 2013). All three species have broad elevational ranges extending into the lowlands and all often occur in disturbed areas with a mix of agriculture and second-growth habitat (Heaney et al. 2016b). Although we did not document them, all three probably persist in remnant habitat elsewhere on the mountain and would likely increase with the regeneration of forest cover.
Biogeography, Elevation, and Community Structure Of the five species of native rodents documented from Mt. Pinatubo, Rattus everetti is a widespread Philippine endemic, Chrotomys mindorensis is endemic to Luzon and Mindoro, and Bullimus luzonicus is restricted to Luzon (Heaney et al. 2010). Apomys zambalensis is a regional endemic widespread in the Zambales Mountains
(Heaney et al. 2016b). In surveys of Mt. Tapulao (Balete et al. 2009) and Mt. Natib (Rickart et al. 2013) it was the most abundant species with the broadest elevational distribution.
Apomys sacobianus was described on the basis of a single specimen captured along the Sacobia River in 1956 (Johnson 1962). After 55 years, its rediscovery during our Pinatubo survey provided new material that confirmed its taxonomic status (Heaney et al. 2014) and placed it within the phylogenetic and biogeographic context (Justiniano et al. 2015). Our surveys of Mt. Natib and Mt. Tapulao (Balete et al. 2009; Rickart et al. 2013) failed to document A. sacobianus even in areas of second-growth similar to habitat in which it was abundant on Mt. Pinatubo, revealing a much narrower geographic distribution for this species. Apomys sacobianus may prove to be endemic to Mt. Pinatubo and its immediate surroundings, but additional field surveys in central Luzon are required to demonstrate this.
Data from surveys of non-volant small mammals on mountains throughout Luzon reveal that the number of native species present on a given mountain is strongly correlated with its maximum elevation (Heaney et al. 2016a). During our survey of Mt. Pinatubo, lack of potable water made it difficult to work at higher elevations beyond a reasonable distance from base camps, and we were unable to establish survey localities above 1,100 m. Although we think it unlikely that we would have recorded additional species in the regenerating habitat at higher elevations, we acknowledge that the following discussion is based, in part, on conjecture. We documented five native species on Mt. Pinatubo (six including historical records of Tryphomys adustus as discussed above). At its current elevation (1,486 m) the elevation-species richness relationship would predict at least five species on Pinatubo, which is the number documented for Mt. Irid with a slightly lower elevation at 1,469 m (Balete et al. 2013). However, before the 1991 eruption, Mt. Pinatubo stood at 1,745 m (Newhall et al. 1996) and its upper slopes were covered with old-growth forest (Punongbayan et al. 1996). From the elevation-species richness relationship, the predicted total number of species for pre-eruption Pinatubo would fall between six (as documented for Mt. Palali at 1,707 m; Alviola et al. 2011) and eight (for Mt. Anacuao at 1,850 m; Heaney et al. 2013). This suggests that several species may have been lost as a result of the eruption.
In identifying other native non-volant mammals that may have been present on Mt. Pinatubo before the 1991 eruption, we must consider those known to occur elsewhere within the Zambales Mountains complex. Mt. Tapulao, the highest peak in the region (2,037 m), has nine species (Balete et al. 2009) of which five have not
Rickart et al.: Mammals of Mt. PinatuboPhilippine Journal of ScienceVol. 150 No. S1, Special Issue on Biodiversity
129

been documented on Mt. Pinatubo. Two of these, Apomys brownorum and Rhynchomys tapulao, are known only from the summit of Mt. Tapulao and may be endemic to that mountain, and it is unlikely that either occurred on Pinatubo. The three remaining species are Crocidura grayi (the only native shrew on Luzon), Apomys microdon, and A. musculus (both are small arboreal mice). All are widespread on Luzon and have elevational ranges that either extend down to near sea level or (in the case of A. musculus) from the highest peaks down to mid-elevation montane forest (Heaney et al. 2016b). We conclude that these species probably occurred on Mt. Pinatubo before the eruption. All require forest cover to maintain mesic conditions that particularly favor C. grayi, and to provide the above-ground foraging space and nest sites utilized by both A. microdon and A. musculus (Heaney et al. 2016b). Although the early-successional habitats of our survey localities lacked these essential features, areas of remnant forest habitat elsewhere on the mountain may support these three species. Based on previous studies cited above, we predict their expansion into regenerating habitat on Mt. Pinatubo once the secondary forest has had time to develop.
Another pattern that has emerged from mammal surveys elsewhere on Luzon is the significant correlation between the local maximum elevation of a mountain range or isolated peak, and the number of non-volant small mammals that are locally endemic (Heaney et al. 2016a). From this elevation-local endemism relationship, we would expect pre-eruption Pinatubo to have at least one local endemic. As discussed above, Apomys sacobianus may be a Pinatubo endemic. Unlike many of the endemic non-volant mammals of Luzon that are restricted to mountaintops, A. sacobianus occurs from the lowlands to mid-elevations (at least 1080 m; Table 2). It is one of several species of Apomys (subgenus Megapomys) that diversified within the lowlands as opposed to the highlands (Justiniano et al. 2015). The remaining question is whether there might have been locally endemic species restricted to high elevations on Mt. Pinatubo prior to the 1991 eruption. This is highly unlikely given the geological history of frequent explosive eruptions of even greater magnitude than the 1991 event (Newhall et al. 1996).
Disturbance Ecology and Conservation In light of our results, we review the three hypotheses regarding the expected distribution and abundance of non-volant small mammals presented in the introduction:
• Non-native small mammals are numerically dominant in the most severely disturbed habitat.
This hypothesis clearly was not supported by our results. The five localities where non-volant mammals were
surveyed all had early second-growth habitat in which non-native species were much less abundant than native species, accounting for only 19 (5.5%) of 348 total animals captured. Non-native rodents were unevenly distributed across the survey gradient (Table 2); most were captured at the three lowest localities adjacent to areas with cultivated kaingin, whereas only one individual was captured at a higher elevation where there was no kaingin. Most previous disturbance gradient studies on Luzon involved highly disturbed second-growth habitat near active agriculture (Rickart et al. 2011; Reginaldo and Ong 2020). The non-native rodents involved here and in our other studies (Rattus exulans and R. tanezumi) are human commensals that achieve the greatest abundance in croplands or in other human-dominated habitats where they have ready access to food crops or stored produce; both are at a severe disadvantage in areas where they compete with native species for naturally occurring foods. In retrospect, this hypothesis should be amended to read: “non-native small mammals are most abundant in severely disturbed habitats where they have access to food crops or other artificial food sources.”
• A subset of native species that tolerate severe habitat disturbance coexists with non-native species.
Results of the Pinatubo survey strongly support this hypothesis. Bullimus luzonicus, Chrotomys mindorensi, and Rattus everetti are among the subset of native small mammals found to be highly tolerant of disturbance in surveys elsewhere on Luzon (Rickart et al. 2011). Apomys sacobianus and A. zambalensis are local or regional endemics that share this ability to tolerate disturbance. In the case of A. sacobianus, “tolerate” is an understatement in that it was by far the most abundant small mammal on the mountain. This species is discussed in further detail below.
• Native small mammals are resilient, moving into areas of regenerating habitat and ultimately displacing non-native species as native vegetation matures.
This hypothesis also is supported by our results but to a much greater extent than we anticipated. The native rodents on Mt. Pinatubo are effectively pioneer species, capable of occupying early second-growth habitat. We predict the future occurrence of other, less tolerant, native species (e.g. Crocidura grayi and Apomys microdon) with the development of suitable forested habitat on the mountain.
The 1991 eruption of Mt. Pinatubo was one of the most powerful in recent history; however, several more powerful eruptions occurred earlier in its existence (Newhall et al. 1996). Cataclysmic geological events that are seemingly rare from the perspective of human history are commonplace when viewed over geological time. As
Rickart et al.: Mammals of Mt. PinatuboPhilippine Journal of ScienceVol. 150 No. S1, Special Issue on Biodiversity
130

these events created the physical fabric of the Philippines, they also shaped the evolution of its remarkably rich biota. Throughout their existence, Philippine mammals have regularly experienced catastrophic disturbances, some more devastating and widespread than the recent eruption of Mt. Pinatubo, not only those associated with geological processes but also much more frequently by powerful typhoons (Heaney et al. 2016b).
Although many species of native Philippine mammals can tolerate substantial disturbance (Rickart et al. 2011, 2013; Heaney et al. 2016b), we nonetheless find it remarkable that so many native species were present in areas of early successional habitat on Mt. Pinatubo. Unfortunately, we have no data on the residual mammal fauna immediately following the eruption. However, a parallel event – the 1980 eruption of Mount St. Helens in SW Washington, USA – provides insight into the probable situation following the eruption of Mt. Pinatubo. In the months following the Mount St. Helens eruption, small mammal surveys conducted across a gradient of habitat damage revealed stark differences in species occurrence patterns and an inverse relationship between the severity of habitat damage and the number of residual species present (Andersen and MacMahon 1985a).
We postulate that mammals on Mt. Pinatubo exhibited a similar pattern of differential survival across a gradient of habitat damage, with the greatest numbers and diversity in areas where damage was least severe, and only the hardiest surviving in scattered sheltered refugia in areas of greater damage. Burrowing mammals such as Chrotomys may have had high survival rates even where habitat was severely damaged, as was the case with fossorial rodents (Thomomys) on Mount St. Helens that survived underground in areas with extreme eruption damage where they were instrumental in regenerating soil and promoting early revegetation (Andersen and MacMahon 1985b).
Bats and large mammals probably dispersed from surrounding intact areas shortly after the eruption of Mt. Pinatubo (del Moral and Grishin 1999; Crisafulli et al. 2015) and, as seed dispersers, undoubtedly played an important early role in plant succession (Marler and del Moral 2011). Fruit bats and frugivorous birds were probably responsible for the diversity and abundance of figs noted at our survey localities and will continue to influence future forest development on the mountain (Utzurrum 1995; Ingle 2003).
Perhaps the most remarkable aspect of our work on Mt. Pinatubo was the rediscovery of Apomys sacobianus. Prior to our surveys in central Luzon, we assumed that this species might be more broadly distributed within the Zambales Mountains. The fact that we did not find it either on Mt. Tapulao (Balete et al. 2009) or Mt. Natib (Rickart et
al. 2013) demonstrated that it had a restricted distribution and raised the specter that, if locally endemic to Mt. Pinatubo, it might have become extinct as a result of the eruption. Although A. sacobianus may indeed be restricted to Mt. Pinatubo and surrounding lowland areas, our results revealed it to be widespread and the most abundant species on the mountain. Among all of the species that we documented, it was the most successful in coping with extreme disturbance and thriving under the harsh conditions characteristic of early-successional habitats. Compared to species that are broadly distributed, range-restricted species such as A. sacobianus are widely considered to be less tolerant of habitat disturbance and competitively inferior, particularly those that are endemic to tropical island systems (Berglund et al. 2009; Walsh et al. 2012). This certainly is not the case for small mammal communities on Luzon that are dominated by endemic species, many of which are ecological specialists with restricted geographic distributions yet are tolerant of habitat disturbance and resist invasion by non-native species (Rickart et al. 2011; Heaney et al. 2016b). Among Philippine mammals, Apomys sacobianus is perhaps the most extreme example of a “disturbance specialist,” a range-restricted species that is highly adapted to life in a landscape that is periodically ravaged by catastrophic events.
As is the case with sites of other devastating volcanic eruptions (del Moral and Grishin 1999), Mt. Pinatubo provides an opportunity to study aspects of biotic recovery following extreme disturbance – including rates of habitat regeneration across different levels of local disturbance, changes in species diversity and community composition over time, and complexities of biotic interactions. Habitat recovery on Mt. Pinatubo will be a long and complex process, and coordination of research efforts – as envisioned under the rubric of long-term ecological research (Kim et al. 2018) – could greatly enhance efforts to document this process. Furthermore, understanding the process of ecological recovery from cataclysmic natural disturbances can augment reforestation and other efforts for ecological reclamation of human-disturbed landscapes. As such, lessons learned from the tragedy of Mt. Pinatubo could ultimately support efforts to protect and restore biological communities throughout the Philippines and beyond.
ACKNOWLEDGMENTS We gratefully acknowledge our colleagues J. Sarmiento, R. Buenviaje, and R. Plutado, without whose help this project would not have been possible. We thank the mayors and staff of Mabalacat City and Municipality, and the barangay officials of San Vicente for their cooperation. We thank the Aeta of Sitio Burog for their
Rickart et al.: Mammals of Mt. PinatuboPhilippine Journal of ScienceVol. 150 No. S1, Special Issue on Biodiversity
131

assistance and for sharing their knowledge, especially O. Sanchez, C. Tolentino, and R. Tolentino. We appreciate the support of the DENR–Protected Areas and Wildlife Bureau (redesignated the Biodiversity Management Bureau), especially T.M. Lim, R.L. Calderon, J. de Leon, C. Custodio, M. Mendoza, A.M. Tagtag, and T. Tenazas. DENR Wildlife Gratuitous Permit No. 262 was issued to Dr. Lawrence Heaney of the Field Museum. The figures were prepared by A. Niedzielski and L. Nassef. We thank A. Goldman and A. Niedzielski for preparing specimens for study, and J. Phelps and the late W. Stanley for technical support. This project was supported by the Negaunee Foundation, the Grainger Foundation, the Brown Fund for Mammal Research, and the Ellen Thorne Smith Fund of the Field Museum.
REFERENCESALVIOLA PA, DUYA MRM, DUYA MV, HEANEY
LR, RICKART EA. 2011. Mammalian diversity pat-terns on Mount Palali, Caraballo Mountains, Luzon. Fieldiana: Life and Earth Sciences 2: 61–74.
ANDERSEN DC, MACMAHON JA. 1985a. The effects of catastrophic ecosystem disturbance: the residual mammals of Mount St. Helens. Journal of Mammal-ogy 66: 581–589.
ANDERSEN DC, MACMAHON JA. 1985b. Plant suc-cession following the Mount St. Helens volcanic erup-tion: Facilitation by a burrowing rodent, Thomomys talpoides. American Midland Naturalist 114: 62–69.
BALETE DS, HEANEY LR, RICKART EA. 2013. The mammals of Mt. Irid, southern Sierra Madre, Luzon Island. National Museum of the Philippines: Journal of Natural History 1: 15–29.
BALETE DS, HEANEY LR, VELUZ MJ, RICKART EA. 2009. Diversity patterns of small mammals in the Zambales Mts., Luzon, Philippines. Mammalian Biology 74: 456–466.
BARBEHENN K, SUMANGIL JP, LIBAY JL. 1973. Rodents of the Philippine croplands. Philippine Agri-culturalist 56: 217–242.
BERGLUND H, JÄREMO J, BENGTSSON G. 2009. En-demism predicts intrinsic vulnerability to nonindigenous species on islands. The American Naturalist 174: 94–101.
CRISAFULLI CM, SWANSON FJ, HALVORSON JJ, CLARKSON B. 2015. Volcano ecology: Disturbance characteristics and assembly of biological communi-ties. In: Sigurdsson H, Houghton B, McNutt S. eds. Encyclopedia of Volcanoes, 2nd ed. London: Elsevier Publishers. p. 1265–1284.
DEL MORAL R, GRISHIN SY. 1999. Volcanic dis-turbances and ecosystem recovery. In: Ecosystems of Disturbed Ground. Ecosystems of the World, Vol. 16. Walker LR ed. Amsterdam: Elsevier. p. 137–160.
GENOWAYS HH, MCLAREN SB, TIMM RM. 2020. Innovations that changed mammalogy: The Japanese mist net. Journal of Mammalogy 101: 627–629.
HEANEY LR, BALETE DS, ALVIOLA PA, DUYA MRN, RICKART EA. 2013. The mammals of Mt. Anacuao, NE Luzon Island, Philippines: a test of predictions of Luzon small mammal biodiversity pat-terns. National Museum of the Philippines: Journal of Natural History 1: 1–13.
HEANEY LR, BALETE DS, DUYA MRM, DUYA MV, JANSA SA, STEPPAN SJ, RICKART EA. 2016a. Doubling diversity: a cautionary tale of previously unsuspected mammalian diversity on a tropical oceanic island. Frontiers of Biogeography. p. 1–19.
HEANEY LR, BALETE DS, RICKART EA. 2016b. The Mammals of Luzon: Biogeography and Natural History of a Philippine Fauna. Baltimore: The Johns Hopkins University Press. 287p.
HEANEY LR, BALETE DS, VELUZ MJ, STEPPAN SJ, ESSELSTYN JA, PFEIFFER AW, RICKART EA. 2014. Two new species of Philippine forest mice (Apomys, Muridae, Rodentia) from Lubang and Luzon islands, with a redescription of Apomys sacobianus Johnson, 1962. Proceedings of the Biological Society of Washington 126: 395–413.
HEANEY LR, DOLAR ML, BALETE DS, ESSEL-STYN JA, RICKART EA, SEDLOCK JL. 2010. Synopsis of Philippine Mammals. Retrieved on 19 Sep 2020 from http://www.fieldmuseum.org/philip-pine_mammals/
INGLE NR. 2003. Seed dispersal by wind, birds, and bats between Philippine montane rainforest and succes-sional vegetation. Oecologia 134: 251–261.
JOHNSON DH. 1962. Two new murine rodents. Pro-ceedings of the Biological Society of Washington 75: 317–319.
JUSTINIANO R, SCHENK JJ, BALETE DS, RICKART EA, ESSELSTYN JA, HEANEY LR, STEPPAN SJ. 2015. Testing diversification models of endemic Philippine forest mice (Apomys) with nuclear phy-logenies across elevational gradients reveal repeated colonization of isolated mountain ranges. Journal of Biogeography 42: 51–64.
KIM ES, TRISURAT Y, MURAOKA H, SHIBATA H, AMOROSO V, BOLDGIV B, HOSHIZAKI K, KASSIM AR, KIM YS, NGUYEN HQ, OHTE N,
Rickart et al.: Mammals of Mt. PinatuboPhilippine Journal of ScienceVol. 150 No. S1, Special Issue on Biodiversity
132

ONG PS, WANG CP. 2018. The International Long-Term Ecological Research–East Asia–Pacific Regional Network (ILTER-EAP): history, development, and perspectives. Ecological Research 33: 19–34.
MARLER TE, DEL MORAL R. 2011. Primary suc-cession along an elevation gradient 15 years after the eruption of Mount Pinatubo, Philippines. Pacific Sci-ence 65: 157–173.
[MGB] Mines and Geosciences Bureau. 2010. Geology of the Philippines, 2nd ed. Quezon City, Philippines.
NEWHALL CG, DAAG AS, DELFIN FG, HOBLITT RP, MCGEEHIN J, PALLISTER JS, REGALADO MT, RUBIN M, TUBIANOSA BS, TAMAYO RA, UMBAL JV. 1996. Eruptive history of Mount Pina-tubo. In: Fire and Mud: Eruptions and Lahars of Mount Pinatubo, Philippines. Newhall CG, Punongbayan RS eds. Quezon City: Philippine Institute of Volcanology and Seismology; Seattle: University of Washington Press. p. 165–195.
PUNONGBAYAN RS, NEWHALL CG, HOBLITT RP. 1996. Photographic record of rapid geomorphic change at Mount Pinatubo, 1991–94. In: Fire and Mud: Eruptions and Lahars of Mount Pinatubo, Philippines. Newhall CG, Punongbayan RS eds. Quezon City: Philippine Institute of Volcanology and Seismology; Seattle: University of Washington Press. p. 21–66.
REGINALDO AA, ONG PS. 2020. Structure of small non-flying mammal communities in disturbed habitats in the Central Cordillera, Luzon Island, Philippines. Philippine Science Letters 13: 81–94.
RICKART EA, BALETE DS, ROWE RJ, HEANEY LR. 2011. Mammals of the northern Philippines: toler-ance for habitat disturbance and resistance to invasive species in an endemic fauna. Diversity and Distribu-tions 17: 530–541.
RICKART EA, HEANEY LR, BALETE DS, AL-VIOLA PA, DUYA MRM, DUYA MV, ROSELL-AMBAL G, SEDLOCK JL. 2013. The mammals of Mt. Natib, Bataan Province, Luzon, Philippines. National Museum of the Philippines: Journal of Natural History 1: 31–44.
RICKART EA, HEANEY LR, UTZURRUM RCB. 1991. Distribution and ecology of small mammals along an elevational transect in southeastern Luzon, Philippines. Journal of Mammalogy 72: 458–469.
UTZURRUM RCB. 1995. Feeding ecology of Philippine fruit bats: Patterns of resource use and seed dispersal. Symposia of the Zoological Society of London 67: 63–77.
WALSH JC, VENTER O, WATSON JEM, FULLER RA, BLACKBURN TM, POSSINGHAM HP. 2012. Exotic species richness and native species endemism increase the impact of exotic species on islands. Global Ecology and Biogeography 21: 841–850.
WOLFE EW, SELF S. 1983. Structural lineaments and Neogene volcanism in southwestern Luzon. In: The Tectonic and Geological Evolution of Southeast Asian Seas and Islands, Part 2. Hayes DE ed. American Geophysical Union, Geophysical Monograph 27. p. 157–172.
Rickart et al.: Mammals of Mt. PinatuboPhilippine Journal of ScienceVol. 150 No. S1, Special Issue on Biodiversity
133