Embed Size (px)
Citation preview

AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 95:2740 (1994)
Maternal Mortality or Prolonged Growth: Age at Death and Pelvic Size in Three Prehistoric Amerindian Populations
ROBERT G. TAGUE Department of Geography and Anthropology, Louisiana State University, Baton Rouge, Louisiana 70803-4105
KEY WORDS Estrogen, Growth, Pubis, Sexual dimorphism, Survivorship
ABSTRACT This study evaluates the relationship between age at death and pelvic size among adults in three prehistoric Amerindian populations. The issue is to identify if the pelvis continues to grow and remodel in adult- hood, or if there is differential survivorship among adults, particularly among females, based on pelvic size. The samples used in this study are Indian Knoll, Pecos Pueblo, and Libben. Twenty-one measures of the pelvis are analyzed. A correlational analysis among individuals 18 years of age and older shows that the subpubic angle narrows with advancing age in both sexes. The suggested etiology is osteophytic growth on the ischiopubic ramus among older adults. A two-sample test comparing younger (ages 18-24) and older (25 years of age and older) adults shows that the linea terminalis (which represents the pubic and iliac components of the pelvic inlet) is significantly shorter in the younger age group, but this pattern is seen only in females. Two interpretations are consistent with this result. First, a female with a short linea terminalis (i.e., small pel.vic inlet) may have died at a young age due to childbirth-related complications. In prehistory, maternal mortality may have been a leading cause of death, with pelvic inlet capacity being a determinant of survivorship among parturients. Second, the linea terminalis may be unique by continuing to grow in early adulthood in females but not in males; the growth occurs at the medial border of the pubis. Longitudinal growth studies of modern hu- mans provide support for the second interpretation. The selective advantage of a later age at maturation of the pubis in females than males is that the period of growth is prolonged, thereby contributing to sexual dimorphism in pubic length and, correspondingly, linea terminalis length and pelvic inlet circumference. 0 1994 Wiley-Liss, Inc.
This study addresses the relationship be- tween age at death and pelvic size among adults in three samples of prehistoric Amer- indians. Many researchers have compared age with various measures of the cranial and infracranial skeleton among adults. These researchers have interpreted a direct relationship between age and a skeletal di- mension as evidence of continued periosteal apposition of bone throughout adulthood (for a review and references, see Lazenby, 1990). The relative magnitude of change in the skeletal dimensions ranges from approx- imately 1% to 12% (Lazenby, 1990, see his Table 3).
Studies on pelvic growth and remodelling in modern humans have been limited to in- dividuals 18 years of age and younger. The results of these studies suggest that the pel- vis completes growth before the third decade of life. For example, Greulich and Thoms (1944) stated that growth of the pelvic inlet during puberty is rapid and requires only about 18 months. Based on a sample of 23 individuals who had pelvic x-rays taken at both ages 18 and 20-22, Moerman (1981: 124) reported that “post-eighteen year old
Received May 20, 1993; accepted March 21,1994.
0 1994 WILEY-LISS, INC

28 R.G. TAGUE
growth in the pelvis averages less than two millimeters per measurement.” The present study considers whether or not the pelvis continues to grow and remodel after age 18.
However, there is an alternative interpre- tation to a statistically significant relation- ship between age at death and pelvic size. In prehistory, survivorship among females may have been related to pelvic capacity. A female with a small pelvis may have died at a young age due to childbirth-related com- plications. To interpret the results of this study as evidence of maternal mortality, the childbirth-related death would have to have been associated with the first, or perhaps second, birth event. If a woman successfully gave birth to several offspring, then mortal- ity in a subsequent delivery could not be as- cribed to pelvic size alone, unless either sig- nificant contraction in pelvic dimensions occurred with advancing age or fetal size in- creased significantly with advancing parity. Whereas non-first-born neonates weigh more than first-born neonates by an average of 5.7% (183 gm) (Meredith, 19501, differ- ences in cranial dimensions between first- born neonates and those of higher birth or- ders are negligible. Compared to first-born neonates, later born neonates are larger in occipitofrontal circumference (0.4%, 1.4 mm), occipitofrontal diameter (0.5%, 0.6 mm), and biparietal diameter (0.9%, 0.8 mm) (Meredith, 1950, see his Table 4). As age at first birth in prehistory was likely to have been late in the second decade of life or early in the third decade,’ this study com- pares pelvic size between younger adults
‘Among industrialized countries, the age a t menarche has de- clined by 2 to 3 years during the last century. In these countries, average age at menarche now ranges from 12 to 13.5 years of age (Eveleth and Tanner, 1990, see their Table 10 and Fig. 153). The secular trend is attributed to improved nutrition. Followingmen- arche there is a period of adolescent subfecundity, and reproduc- tive maturity is usually not attained until several years later (for discussion and references, see Ellison, 1990). The subfecundity associated with the first few years after menarche is illustrated in Weiss’ (1973, see his Fig. 3 ) model fertility schedule and in Woods (1989, see his Fig. 2.7) fecundability schedule. Both show that fecundability (and fertility) “is typically low in adolescence, but rises rapidly to a peak during the early- to mid-twenties” (Wood, 1989 76). If menarche occurred at mid-adolescence in prehistory, then reproductive maturity would not have been at- tained until late adolescence. Howell (1979) reported that the mean age at first birth among the Dobe !Kung (contemporary hunter-gatherers) is 18.8, and two thirds of the women had their first births between the ages of 17 and 20.
(18-24 years of age) and older adults (25 years of age and older) within each sex.
Maternal mortality may have been one of the leading causes of death among popula- tions that lacked modern obstetric care (but see Wells, 1975). In a sample of pre-Colum- bian Andean mummies, 14% of the females between the ages of 12 and 45 died as a result of childbirth-related complications (Arriaza et al., 1988). Most of the childbirth- related deaths were associated with the pu- erperium, which is the period after birth. Although puerperal infection can result from a difficult labor due to small pelvic size, Arriaza et al. (1988: 40) observed that “the pelves of [these females] . . . were normal in shape, and the metric measurements did not differ significantly from the established nor- mal patterns of gynecologic measurements.” Arriaza et al. (1988) concluded that mater- nal mortality was related to improper or un- sanitary methods of fetal delivery. High rates of maternal mortality also character- ized premodern Europe. Puerperal infection was the principal cause of death; obstructed labor due to small pelvic size was also a sig- nificant factor (Dobbie, 1982; Eccles, 1982; Shorter, 1982). In the United States in 1913, the second leading cause of death among women 1 5 4 4 years of age was mortality as- sociated with childbirth (Meigs, 1917). Among those females dying of childbirth-re- lated complications, the majority were asso- ciated with the puerperium; the second and third leading causes of death were eclamp- sia (or toxemia) during pregnancy and ob- struction to labor due to small pelvic size. In the late 1920s, the rate of maternal mortal- ity among Pueblo Indians was seven times greater than that for other women in the United States; again, mortality was related to the puerperium (Aberle, 1934). Finally, among the Ganda in the early 1950s, diffi- cult labor due to disproportion between the dimensions of the fetal cranium and mater- nal pelvic inlet was common, requiring sur- gical intervention in 12.2% of the parturi- ents (Allbrook and Sibthorpe, 1961).
With the exception of mummified remains (e.g., Arriaza et al., 1988), the most compel- ling evidence of maternal mortality in ar- chaeological material is a burial in which a fetal skeleton is found within an adult fe-

AGE AT DEATH AND PELVIC SIZE 29
male’s abdomino-pelvic cavity (e.g., Owsley and Bradtmiller, 1983). For the skeletal populations used in this study, there were two such burials in both Indian Knoll (Webb, 1974) and Libben (C.O. Lovejoy, pers. comm.). Burial records on Pecos Pueblo were not available.
MATERIALS AND METHODS Samples from three prehistoric Amerin-
dian skeletal populations were used: Indian Knoll, Pecos Pueblo, and Libben. Each popu- lation was temporally and geographically di- verse from the others (Hooton, 1930; Webb, 1974; Lovejoy et al., 1977).
To assign sex to the skeletons, I relied principally on Phenice’s technique (1969) and the shape of the greater sciatic notch. When the skull was present, sex was evalu- ated using standard cranial and mandibular traits (Krogman and Iacan, 1986, see their Table 6.3). From a sample of skeletons of known sex, Meindl et al. (1985a) reported a 4% error rate in the determination of sex when only the pelvis was used and a 3% error rate when both the pelvis and skull were evaluated.
Only adults were analyzed in the present study. A young specimen was classified as an adult if the epiphyses of the distal femur and proximal tibia had fused (for females) or had begun to fuse (for males), and if the epi- physes of the iliac crest and ischial ramus had begun to fuse (for both sexes). Based on Johnston’s (1961) study of the sequence of epiphyseal fusion in the Indian Knoll skele- tal population, the stage of epiphyseal fusion described above characterized indi- viduals who were approximately 18 years of age.
To assign an age at death for each speci- men, the symphyseal surface of the pubis and the auricular surface of the ilium were evaluated, in addition to observations on ep- iphyseal fusion. The methods of Meindl et al. (1985b) for the pubis and Lovejoy et al. (1985b) for the auricular surface were used. Lovejoy et al. (1985a, see their Table 5) re- ported that the correlations in a modern ca- daver sample between actual age at death and the assigned age based only on the pubis or auricular surface were 0.78 and 0.71, re- spectively. In the present study, the pubes
and auricular surfaces were analyzed inde- pendently of each other, with the male and female samples being evaluated separately. For example, before the pubes were ana- lyzed, the auricular surfaces were covered with tape. The pubes were then seriated from youngest to oldest. Following seriation, an age was assigned to each specimen based on pubic symphyseal morphology. The au- ricular surfaces were then seriated and aged, with the pubes covered with tape.
After both the pubes and auricular sur- faces were analyzed, the two age estimates were averaged to provide a final age at death for each specimen. In the minority of cases when only one joint surface was pre- served, that single age indicator was used to assign an age at death. This reliance on ei- ther joint surface for assigning an age at death is appropriate because Lovejoy et al. (1985a, see their Table 4) reported that among the five skeletal and dental traits used in their study to estimate age at death, the highest correlation was between the pu- bis and auricular surface (r = 0.75). In the present study, the mean Pearson’s product- moment correlation coefficient between the assigned ages for the pubis and auricular surface is 0.73, which compares favorably to that reported by Lovejoy et al. (1985a). The correlations for each sample are 0.83 for males and 0.73 for females in Indian Knoll, 0.68 for both sexes in Pecos Pueblo, and 0.72 for both sexes in Libben.
Some of the measurements in this study were taken with reference points on the symphyseal surface of the pubis and auricu- lar surface of the ilium (Fig. 1). As these two joint surfaces may be involved in the hy- pothesized growth and remodelling of the pelvis in adulthood, their accuracy in assess- ing age at death may be compromised. To determine whether or not the assigned ages based on the pubis and auricular surface were biased, a product-moment correlation coefficient was calculated using the Libben sample between the ages in this study and those assigned by Lovejoy (1985) based on dental wear. Dental wear is regarded as an accurate indicator of age at death (Lovejoy, 1985; Lovejoy et al., 1985a). The correlation is 0.78 (N = 101, P < 0.001). The implication is that the pubis and auricular surface pro-
rn P P
e
a C
f
b
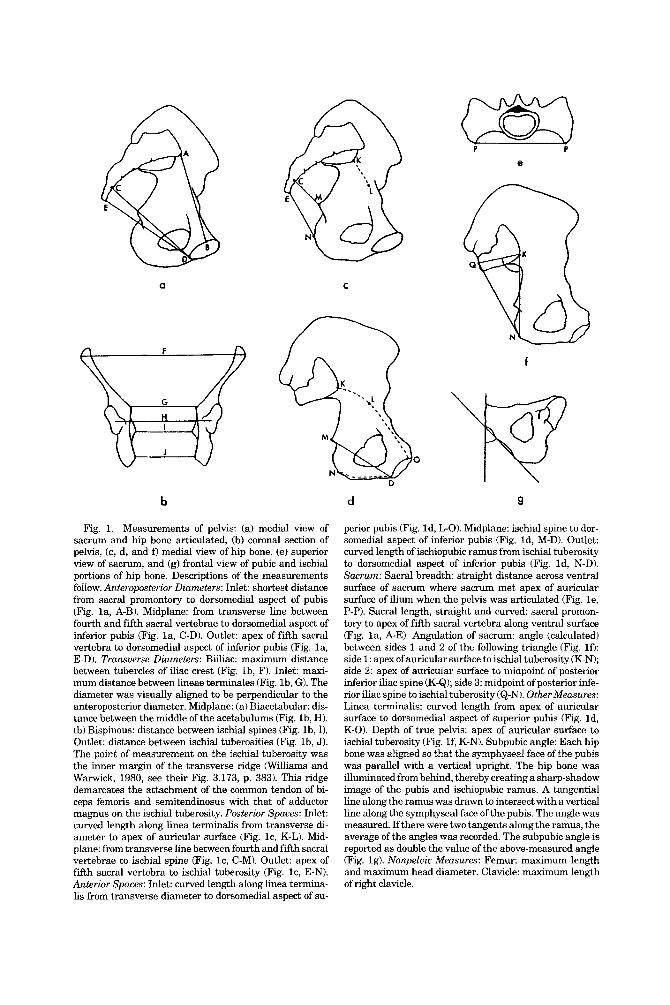
Fig. 1. Measurements of pelvis: (a) medml view of sacrum and hip bone articulated, (b) coronal section of pelvis, (c, d, and f) medial view of hip bone, (e) superior view of sacrum, and (g) frontal view of pubic and ischial portions of hip bone. Descriptions of the measurements follow. Anteroposterwr Diameters: Inlet: shortest distance from sacral promontory to dorsomedial aspect of pubis (Fig. l a , A-B). Midplane: from transverse line between fourth and fifth sacral vertebrae to dorsomedial aspect of inferior pubis (fig. l a , C-D). Outlet: apex of fifth sacral vertebra to dorsomedial aspect of inferior pubis (Fig. l a , E-D). Transverse Diameters: Biiliac: maximum distance between tubercles of iliac crest (Fig. lb , F). Inlet: maxi- mum distance between lineae terminales (Fig. lb, G). The diameter was visually aligned to be perpendicular to the anteroposterior diameter. Midplane: (a) Biacetabular: dis- tance between the middle of the acetabulums (Fig. lb , H). (b) Bispinous: distance between ischial spines (Fig. lb, I). Outlet: distance between ischial tuberosities (Fig. lb , J). The point of measurement on the ischial tuberosity was the inner margin of the transverse ridge (Williams and Warwick, 1980, see their Fig. 3.173, p. 383). This ridge demarcates the attachment of the common tendon of bi- ceps femoris and semitendinosus with that of adductor magnus on the ischial tuberosity. Posterior Spaces: Inlet: curved length along linea terminalis from transverse di- ameter to apex of auricular surface (Fig. lc, K-L). Mid- plane: from transverse line between fourth and fifth sacral vertebrae to ischial spine (Fig. lc, C-M). Outlet: apex of fifth sacral vertebra to ischial tuberosity (Fig. lc, E-N). Anterior Spaces: Inlet: curved length along linea termina- lis from transverse diameter to dorsomedial aspect of su-
d 9
perior pubis (Fig. Id, L-0). Midplane: ischial spine to dor- somedial aspect of inferior pubis (Fig. Id, M-D). Outlet: curved length of ischiopubic ramus from ischial tuberosity to dorsomedial aspect of inferior pubis (Fig. Id, N-D). Sacrum: Sacral breadth straight distance across ventral surface of sacrum where sacrum met apex of auricular surface of ilium when the pelvis was articulated (Fig. le, P-P). Sacral length, straight and curved: sacral promon- tory to apex of fifth sacral vertebra along ventral surface (Fig. l a , A-E). Angulation of sacrum: angle (calculated) between sides 1 and 2 of the following triangle (Fig. If): side 1 : apex of auricular surface to ischial tuberosity (K-N); side 2: apex of auricular surface to midpoint of posterior inferior iliac spine (K-Q); side 3: midpoint of posterior infe- rior iliac spine to ischial tuberosity (Q-N). OtherMeasures: Linea terminalis: curved length from apex of auricular surface to dorsomedial aspect of superior pubis (Fig. Id, K-0). Depth of true pelvis: apex of auricular surface to ischial tuberosity (Fig. lf , K-N). Subpubic angle: Each hip bone was aligned so that the symphyseal face of the pubis was parallel with a vertical upright. The hip bone was illuminated from behind, thereby creating a sharp-shadow image of the pubis and ischiopubic ramus. A tangential line along the ramus was drawn to intersect with a vertical line along the symphyseal face of the pubis. The angle was measured. Ifthere were two tangents along the ramus, the average of the angles was recorded. The subpubic angle is reported as double the value of the above-measured angle (Fig. lg). Nonpeluic Measures: Femur: maximum length and maximum head diameter. Clavicle: maximum length of right clavicle.

31 AGE AT DEATH AND PELVIC SIZE
vide an estimate of age at death comparable to that of dental wear.
For a few of the specimens in this study, both the pubis and the auricular surface were damaged, thereby precluding assign- ment of a specific age at death. However, if one or both of the joint surfaces were suffi- ciently preserved to permit assignment of an age at death of at least 25 years, then the specimen was included in one of the statistical analyses (Student’s t-test, see below). Therefore, sample sizes differed slightly between the two statistical anal- yses.
This study tested for a statistical associa- tion between age at death and pelvic size in three independent samples. The research strategy was to identify a pattern to the re- sults among the samples. The reasonable in- ference is that a uniform pattern of results reflects a common biological process. Prod- uct-moment correlation coefficients were calculated between age at death and a suite of pelvic (and nonpelvic) measurements, and a two-tailed test of significance (P s 0.05) was applied. A two-tailed test of significance was used because the dimensions of the pel- vis may either increase or decrease if remod- elling occurs throughout adulthood. In cal- culating the correlation coefficient, the specimens used were those for which a spe- cific age at death was assigned. In the Stu- dent’s t-test, pelvic (and nonpelvic) dimen- sions were compared between individuals 18-24 years of age and those 25 years of age and older. A one-tailed test of significance (P 0.05) was applied as both hypotheses (continued pelvic growth in early adulthood and differential survivorship based on pelvic size) specify that younger adults will have smaller dimensions. Included in this analy- sis were both those specimens for which a specific age at death was assigned and those who were assigned a minimum age at death of at least 25 years. However, there are two fundamental assumptions in interpreting the results of the statistical analyses. As the skeletons within each burial population were accumulated over a period of several hundred years (Hooton, 1930; Webb, 1974; Lovejoy et al., 19771, one must assume within each population (1) genetic related- ness among the individuals and (2) absence of a significant secular trend in pelvic size.
The hip bones and sacra were frequently damaged in the material used in this study. Consequently, sample sizes for the pelvic measurements varied. To measure each pel- vis, the hip bones and sacrum were articu- lated, and the pelvis was encircled with sev- eral heavy rubber bands. The pubes touched in the midline; no compensation was made for a symphyseal disk. Measurements were taken with sliding and spreading calipers, a curvometer, and an osteometric board. For Indian Knoll, Charles Snow’s measure- ments for maximum diameter of the femoral head were used. A description of the mea- surements accompanies Figure l .
RESULTS The first issue is whether or not there is a
relationship between age at death and pel- vic size among adults. Table 1 presents the product-moment correlation coefficients. The results show that neither sex demon- strates an unequivocal (i.e., consistent) as- sociation between age at death and a pelvic (or nonpelvic) measure. There is, however, a significant negative association between age at death and subpubic angle in four of the six samples (both sexes in Pecos Pueblo and Libben), and the correlation coefficient approaches significance (P = 0.073) in a fifth sample (Indian Knoll females).
The second issue is whether or not there is a difference in pelvic size between younger (18-24 years of age) and older (25 years of age and older) adults. Table 2 presev-ts the results of Student’s t-tests. The results show that only the linea terminalis is signifi- cantly shorter in the younger age group in all three samples, and this pattern is seen only in females. Among males, only one sam- ple (Libben) shows a significant difference in linea terminalis length between younger and older adults. The results are equivocal for other pelvic measures. Finally, the clavi- cle is significantly shorter among younger adults in four samples (males in Indian Knoll, females in Pecos Pueblo, and both sexes in Libben) and nonsignificantly shorter in a fifth sample (females in Indian Knoll).
DISCUSSION Although the results of the correlational
analysis are not uniform, the subpubic angle
32 R.G. TAGUE
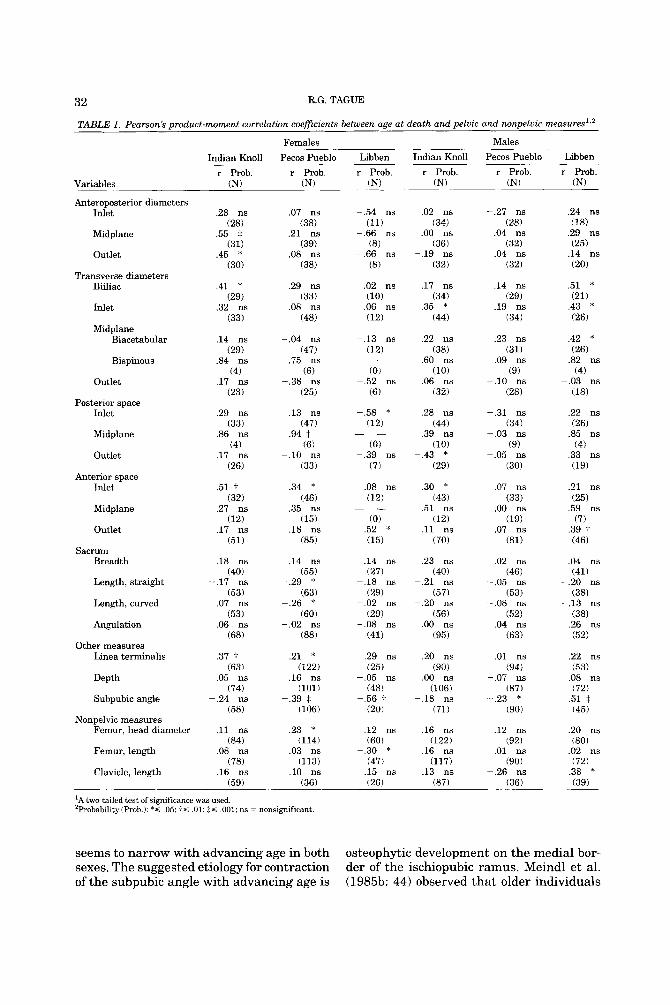
TABLE 1. Pearson's product-moment correlation coefficients between age at death and pelvic and nonpelvie measures'.'
Females Males
Indian Knoll Pecos Pueblo Libben Indian Knoll Pecos Pueblo Libben r Prob. r Prob. r Prob. r Prob. r Prob. r Prob.
Variables (N) (N) (N) (N) (N) (N)
Anteroposterior diameters Inlet
Midplane
Outlet
Transverse diameters Biiliac
Inlet
Midplane Biacetabular
Bispinous
Outlet
Posterior space Inlet
Midplane
Outlet
Anterior space Inlet
Midplane
Outlet
Sacrum Breadth
Length, straight
Length, curved
Angulation
Other measures Linea terminalis
Depth
Subpubic angle
Nonpelvic measures Femur, head diameter
Femur, length
Clavicle, length
.28 ns (281
.55 $ (31)
.45 * (30)
.41 * (29)
.32 ns (33)
.14 ns (29)
.84 ns (4)
.17 ns (23)
.29 ns (331
.86 ns (4)
.17 ns (26)
.51 t (32)
.27 ns (12)
.17 ns (51)
.18 ns (401
-.17 ns (53)
.07 ns (53)
.06 ns (68)
.37 t (63)
.05 ns (74)
-.24 ns (58)
.ll ns (84)
.08 ns (78)
.16 ns (59)
.07 ns (38)
21 ns (39)
.08 ns (38)
.29 ns (33)
.08 ns (48)
-.04 ns (47)
.75 ns (6)
-.38 ns (25)
-.13 ns (47)
.94 t (61
- . lo ns (33)
.34 * (46)
.35 ns (15)
.18 ns (85)
.14 ns (55)
-.29 * (63)
-.26 * (60)
-.02 ns (88)
.21 * (122)
.16 ns (101)
(106) -.39 p
.23 * (114)
.03 ns (113)
.10 ns (36)
-.54 ns (11)
-.66 ns (8)
-.66 ns (8)
.02 ns (10)
.06 ns (12)
-.13 ns (12)
(0) -.52 ns
(6)
_ _
-.58 * (12)
(0) -.39 ns
(7)
_ _
.08 ns (12)
(0) .52 * (15)
.14 ns (27)
-.18 ns (29)
-.02 ns (29)
-.08 ns (41)
.29 ns (25)
p.05 ns (48)
p.56 t (20)
.12 ns (60)
-.30 * (47)
.15 ns (26)
- -
.02 ns (341
.OO ns (36)
-.19 ns (32)
.17 ns (34)
.35 * (44)
.22 ns (38)
.60 ns (10)
.06 ns (32)
.28 ns (44)
.39 ns (10)
-.43 *.
(29)
.30 * (43)
.51 ns (121
.ll ns (70)
.23 ns (40)
-.21 ns (57)
-.20 ns (56)
.OO ns (95)
.20 ns (901
.OO ns (106)
-.18 ns (71)
.16 ns (122)
.16 ns (117)
.13 ns (87)
-.27 ns 128)
.04 ns (32)
.04 ns (32)
.14 ns (29)
.19 ns (34)
.23 ns (31)
-.09 ns (9)
- . lo ns (28)
-.31 ns (34)
-.03 ns (9)
-.05 ns (30)
.07 ns (33)
.OO ns (19)
.07 ns (81)
.02 ns (46)
-.05 ns (531
-.08 ns (52)
.04 ns (63)
.01 ns (941
-.07 ns (87)
-.23 * (90)
.12 ns (92)
.01 ns (90)
-.26 ns (36)
.24 ns (181
.29 ns (25)
.14 ns (20)
.51 * (21)
.43 * (26)
.42 * (26)
.82 ns (4)
-.03 ns (18)
.22 ns (26)
.85 ns (4)
.33 ns (19)
.21 ns (25)
.59 ns (7)
.39 t (46)
.04 ns (411
-.20 ns (38)
-.13 ns (38)
.26 ns (52)
.22 ns
.08 ns (53)
(72) -51 $
(45)
.20 ns (80)
.02 ns (72)
.38 * (39)
'A two-tailed test of significance was used. 'Probability(Pr0b.): * S .05: t s .01; IS ,001; ns = nonsignificant.
seems to narrow with advancing age in both sexes. The suggested etiology for contraction of the subpubic angle with advancing age is
osteophytic development on the medial bor- der of the ischiopubic ramus. Meindl et al. (198513: 44) observed that older individuals

AGE AT DEATH AND PELVIC SIZE 33
develop “bony outgrowths of the pubis and especially.. . of the ischium. . . . These in- clude the tendonous attachments of the me- dial adductor group (especially the gracilis) and . . . the ischial attachment of the sacro- tuberous ligament.” Collectively, however, the results of the correlational analysis do not support a generalization that through- out adulthood there is either differential survivorship based on pelvic size or contin- ual growth and remodelling of the pelvis. While continual periosteal apposition may occur during adulthood, the magnitude of apposition is not sufficient to be detected in the correlational analysis.
The results of the t-tests show that the clavicle is significantly shorter among younger adults relative to older adults in four of six samples, with both sexes evidenc- ing this age group difference. Although the results are not uniform, the implication is that the clavicle continues to lengthen dur- ing early adulthood in males and females. This explanation is supported by studies on samples of individuals of known age, sex, and race that show that the majority of indi- viduals in both sexes do not evidence com- plete fusion of the medial epiphysis of the clavicle until age 25 (Todd and D’Errico, 1928; McKern and Stewart, 1957; Webb and Suchey, 1985).
The present study shows that only the linea terminalis is significantly shorter among younger adults relative to older adults in all three samples, and this pattern is seen only in females. As the results for the linea terminalis are statistically significant only in the two-sample test and not in the correlational analysis, the implication is that the biological significance of differen- tial linea terminalis length is manifested in early adulthood. Two interpretations of these results-maternal mortality and con- tinued growth of the linea terminalis in early adulthood in females but not in males-will be considered.
The linea terminalis is composed of pubic length and lower iliac height (i.e., the dis- tance between the apex of the auricular sur- face and the iliopubic junction). Both compo- nents of the linea terminalis are longer in females than males (Straus, 1927; Wash- burn, 1948, 1949). Moreover, the linea ter-
minalis is the sole determinant of sexual di- morphism in pelvic inlet circumference (Tague, 1992) because the sexes do not differ in the dimensions of the other components of inlet circumference-sacral breadth (Flander, 1978; Segebarth-Orban, 1980; Moerman, 1981; Hager, 1989; Tague, 1992) and width of the pubic symphyseal cartilage (Heyman and Lundqvist, 1932; Abramson et al., 1934). The present study shows that the linea terminalis is 6-7 mm shorter in younger females relative to older females (Table 2). For the three samples, the mean percentage difference in linea termi- nalis length between younger and older adults is 4.6% for females and 2.2% for males. Moreover, whereas older females have a linea terminalis that is an average of 6.4% longer than that of older males, the linea terminalis in younger females is only an average of 1.6% longer than that of older males. Collectively, these results show that younger females had a linea terminalis sig- nificantly shorter than that of older females and, on average, only slightly longer than that of older males. As humans are charac- terized by neonatal cephalo: :female pelvic constriction (i.e., the dimensions of the new- born’s cranium closely approximate those of the adult female’s pelvic inlet [Schultz, 1949]), a small inlet circumference in younger females may have promoted obstet- ric difficulty.
A spacious circumference to the pelvic in- let is requisite for obstetric success because pregnancy-related changes in inlet circum- ference are minimal. During pregnancy, the hormone relaxin is secreted. Relaxin alters the mechanical properties of the pelvic liga- ments so that these ligaments become more pliable (Shenvood, 1988). The attendant in- crease in pelvic joint mobility enhances ob- stetric success because the dimensions of the bony birth canal can temporarily en- large to accommodate the fetus. The dimen- sions of the lower pelvic planes can enlarge significantly as a result of increased mobil- ity of the sacroiliac joints (Bore11 and Fern- strom, 1957; Ohlsen, 1973); Russell (1969: 820) estimated that the “area of the outlet increases . . . by approximately 20 to 30 per cent.” Conversely, change in the circumfer- ence of the pelvic inlet is minimal. The sym-
Var
iabl
es
Ant
erop
oste
rior
dia
met
ers
Inle
t (m
m)
Mid
plan
e (m
m)
Out
let (
mm
)
Tra
nsve
rse
diam
eter
s B
iilia
c (m
m)
Inle
t (m
m)
Mid
plan
e B
iace
tabu
lar
(mm
)
Bis
pino
us (m
m)
Out
let (
mm
)
Post
erio
r sp
ace
Inle
t (m
m)
Mid
plan
e (m
m)
Out
let
(mm
)
Ant
erio
r sp
ace
Inle
t (m
m)
Mid
plan
e (m
m)
Out
let l
mm
)
Indi
an K
noll
Fem
ales
Peco
s Pu
eblo
L
ibbe
n
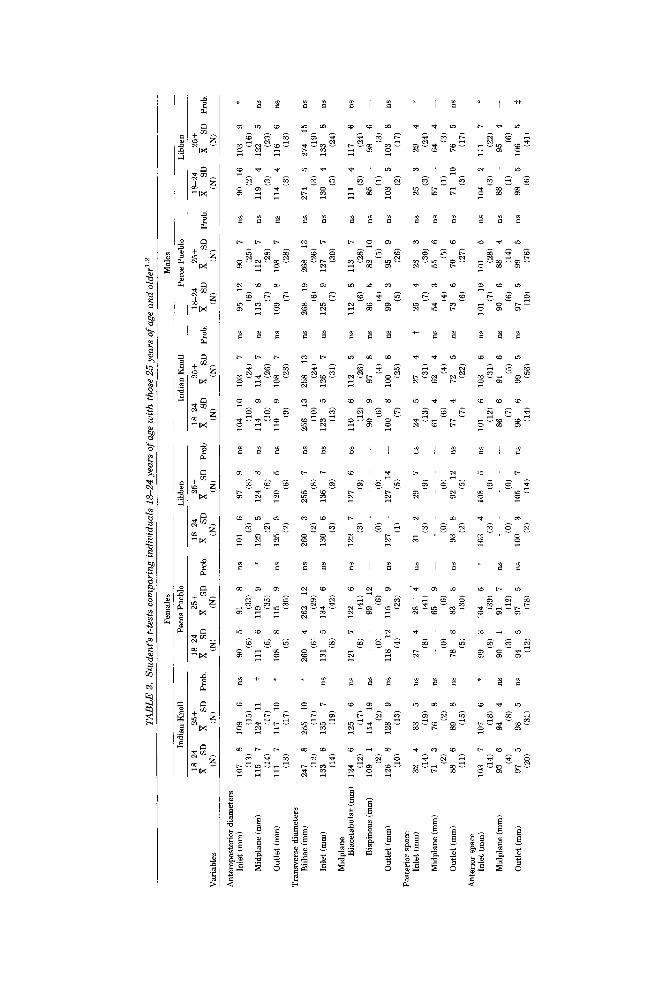
TAB
LE 2
. St
uden
t's t-
test
s com
pari
ng in
divi
dual
s 18
-24
year
s of
age
wit
h th
ose
25 y
ears
of
age
and
olde
r's2
~ ~
la2
4
25+
X SD
x
SD
(N)
(N)
107
8 10
9 6
(13)
(1
5)
115
7 12
4 11
(1
4)
(17)
11
1 7
117
10
(13)
(1
7)
247
8 25
5 10
(1
2)
(17)
13
3 6
135
7 (1
4)
119)
124
6 12
5 6
(12)
(1
7)
109
1
114
19
(2)
(2)
126
8
128
9 (1
0)
(13)
32
4 33
5
(14)
(1
9)
71
3 76
8
12)
(2)
88
6 89
8
(11)
(1
5)
103
7 10
7 6
(14)
(1
8)
93
6 94
4
(4)
(8)
97
5 98
5
(20
) (3
1)
~
1S
24
25
+ Pr
ob.
x SD
fi
SD
Prob
. %
SD
fi
SD
Prob
. (N
) (N
J (N
J (N
)
la2
4
25+
ns
90
5 91
8
ns
101
6 97
9
ns
(6)
(33)
13
) (8
) t
111
6 11
9 9
* 12
9 5
124
8 ns
(6
) (3.5)
(2)
(6)
* 10
8 8
115
9 ns
12
5 8
120
5 ns
(5
) (3
5)
(2)
(6)
* 26
0 4
262
12
ns
260
3 25
5 7
ns
(6)
(29)
(2
) (8)
ns
131
5 13
4 6
ns
130
6 13
6 7
ns
(8)
142)
(3
) (9
)
ns
121
7 12
2 6
ns
122
7 12
7 6
ns
(8)
(41)
(3
) (9
) ns
-
- 99
12
-
..
(0
) (6
) (0
) (0)
ns
118
12
115
9 ns
12
7 -
127
14 -
(4)
(23)
(1
) (5
)
ns
27
4 28
4
ns
31
2 29
7
ns
(8)
(41)
(3
) (9
) ns
--
65
9-
-
.
(0)
(6)
(0)
(0)
ns
78
8 83
8
ns
93
8 92
12
ns
(5
) (3
0)
(2)
(5)
* 99
3
104
5 *
103
4 10
8 5
ns
(8)
139)
(3
) (9
) n
s9
01
91
7n
s
--
(3
) (1
2)
(0)
(0)
ns
94
5 97
5
ns
100
3 10
5 7
ns
(12)
(7
8)
(2)
(14)
Mal
es
Indi
an K
noll
Peco
s Pu
eblo
fi SD
T
SD
Prob
. SD
fi
SD
Prob
. 18
-24
25+
Is2
4
25+
(N)
(N)
(N)
(N)
104
10
103
7 ns
95
12
90
7
ns
114
9 11
4 7
ns
113
8 11
2 7
ns
110
9 10
8 7
ns
109
8 10
8 7
ns
(10)
(2
4)
(6)
(25)
(10)
(26)
(7
) (2
8)
(9)
(23)
(7
) (2
8)
256
13
258
13
ns
268
19
268
13
ns
123
5 12
6 7
ns
125
9 12
7 7
ns
(10)
(2
4)
(6)
(26)
(13)
(3
1)
(7)
(30)
110
6 11
2 5
ns
112
8 11
3 7
ns
90
9 97
8
ns
86
5
82
10
ns
100
8 10
0 8
ns
99
3 95
9
ns
(12)
(2
6)
(6)
(28)
(6)
(4)
(4)
(5)
(7)
(25)
(5
) (2
6)
24
5 27
4
t 25
4
23
3 ns
61
4 62
4
nf
54
3 55
6
ns
77
4 72
5
ns
73
6 70
6
ns
(13)
(3
1)
(7)
(30)
(6)
(4)
(4)
(5)
(6)
(27)
(7
) (2
2)
101
6 10
3 6
ns
101
10
101
5 ns
86
6 91
6
ns
90
6 88
4
ns
96
6 99
5
ns
97
5 99
5
ns
(12)
(3
1)
(7)
(28)
(7)
(5)
(6)
114)
110)
(7
6)
(14)
(5
6)
Lib
ben
la2
4
25+
X SD
SD
Pr
ob.
(NJ
(NJ
90
16
103
9 *
119
4 12
2 5
ns
114
4 11
6 6
ns
(2)
(16)
(3)
(23)
(3)
(18)
271
5 27
4 15
ns
130
4 13
3 8
ns
(3)
(19)
(3)
(24)
111
4 11
7 6
ns
85
- 98
6
-
103
5 10
3 8
ns
(3)
(24)
(1)
(3)
(2)
(17)
25
3 29
4
*
57
- 64
4
-
71
10
76
5 ns
(3)
(24)
(1)
(3)
(3)
(17)
104
2 11
1 7
* 88
-
95
4 -
98
5 10
6 5
f
(3)
(22)
(1)
(6)
(6)
(41)

Sacr
um
Bre
adth
(m
m)
Len
gth,
str
aigh
t (m
m)
Len
gth,
cur
ved
(mm
)
Ang
nlat
ion
(deg
rees
)
Oth
er m
easu
res
Lin
ea te
rmin
alis
(mm
)
Dep
th (m
mJ
Subp
ubic
ang
le
(deg
rees
)
Non
pelv
ic m
easu
res
Fem
ur, h
ead
Fem
ur, l
engt
h (m
m)
Cla
vicl
e, le
ngth
(mm
)
diam
eter
(m
m)
105
6 (1
1)
102
10
(19)
10
9 10
(1
9)
68
5 (2
2)
135
8 (2
3)
100
4 (2
5)
101
7 (1
9)
39
2 (2
7)
408
19
(26)
12
9 5
(16)
106
6 (2
9)
97
10
(34)
10
9 7
(34)
68
6
(46)
141
9 (4
0)
103
6 (4
9)
96
8 (3
9)
39
2 (5
8)
411
18
(53)
13
1 6
(44)
ns
109
4 10
9 5
ns
100
7 96
10
ns
109
7 10
7 7
ns
67
5 70
6
(7)
(51)
(11)
(56)
(11)
(53)
(11)
(8
2)
t 12
6 9
132
7
* 97
5
99
5
ns
95
9 85
9
(19)
(1
08)
(12)
(9
4)
(16)
(9
5)
ns
38
1
39
2
ns
391
19
393
18
ns
128
4 13
3 7
(15)
(1
05)
(22)
(9
7)
ns
109
9 11
2 5
us
(5)
(24)
ns
97
5
101
8 ns
(5
) (2
6)
ns
106
7 11
1 7
ns
(5)
(26)
ns
72
3
73
5 ns
(6
) (3
6)
t 13
6 5
143
9 *
(6)
(23)
ns
10
4 3
103
5 ns
(7
) (4
3)
ns
98
5 95
11
ns
(3
) (1
8)
* 42
3
42
2 ns
(9
) (5
4)
ns
436
25
427
19
ns
(9)
(40)
*
137
2 14
3 8
t
107
4 10
6 6
(10)
(3
0)
108
9 10
3 7
(15)
(4
2)
117
8 11
3 6
(15)
(4
1)
61
5 62
4
(18)
(7
9)
12fi
9 12
9 6
(20)
(7
1)
103
7 10
4 6
(20)
(8
9)
78
12
73
7 (1
5)
(56)
43
2 43
2
(22)
(1
03)
433
24
437
20
(27)
(9
3)
138
8 14
4 8
ns
106
6 10
6 6
ns
102
9 10
1 8
(6)
(42)
(8)
(51)
(8)
(50)
ns
11
0 10
10
9 6
ns
61
5 62
5
(10)
(58)
ns
125
11
126
7
ns
103
7 10
2 5
ns
68
10
61
8
(14)
(8
5)
(10)
(8
3)
(12)
(8
6)
ns
44
2 44
2
ns
424
16
430
18
t 15
7 8
150
7
(12)
(8
7)
(12)
(8
4)
(4)
(36)
ns
110
5 11
0 6
ns
105
13
105
8
ns
114
13
114
8
ns
59
5 61
4
(5)
(38)
(5)
(34)
(5)
(34)
(4)
(50)
ns
131
7 13
7 8
ns
101
4 10
6 5
ns
78
11
68
7
(9)
(45)
(6)
(69)
(4)
(42)
ns
45
2 46
2
ns
453
22
460
20
(11)
(7
4)
(9)
(68)
(4)
(37)
ns
148
6 15
8 8
ns
ns
ns
ns * ns * ns
(7)
(33)
(4
) (2
2)
(16)
(7
3)
'A o
ne-t
aile
d te
st o
f sig
nifi
canc
e was
use
d.
'Pro
babi
lity(
Prob
.):
*C
.05;
7s .
01; t
s ,0
01; n
s =
non
sign
ific
ant.

R.G. TAGUE 36
physis pubis widens by an average of 2-3 mm during pregnancy, and there is no fur- ther increase during labor (Heyman and Lundqvist, 1932; Abramson et al., 1934). This increase in symphyseal width repre- sents less than a 1% change in inlet circum- ference.' Based on an analysis of radio- graphs, Thorp and Fray (1938) were unable to detect any change in the area of the pelvic inlet in women during labor, despite pelvic joint relaxation. Therefore, whereas in- creased pelvic joint mobility can convert a confined pelvic outlet in a nonpregnant woman into one that is obstetrically suffi- cient in a parturient, obstetric adequacy of the inlet is determined by skeletal anatomy. These arguments support an interpretation that linea terminalis length was associated with differential survivorship among adult females.
An alternative interpretation to the sig- nificant difference in linea terminalis length between younger and older adult females is that the pubis continues to grow in females, but not in males, in early adulthood. The linea terminalis comprises the anterior and posterior spaces of the inlet (Fig. Id). Whereas the anterior space of the inlet is significantly shorter in younger females in Indian Knoll and Pecos Pueblo and nonsig- nificantly shorter in Libben, there is no sig- nificant difference between younger and older adult females in the posterior space of the inlet (Table 2). A reasonable inference is that the differences between younger and older adult females in linea terminalis length and anterior space of the inlet are due to the same process-continued growth of the pubis. Studies by Coleman (1969) and Moerman (1981) support this interpreta- tion. Both studies were longitudinal in de- sign, with pelvic growth being evaluated for samples of both sexes between the ages of 8 (Moerman) or 9 (Coleman) and 18. Coleman and Moerman examined pelvic radiographs, which were part of the longitudinal growth study at the Fels Research Institute.
2Mean pelvic inlet circumferences for each sample of adult females (18 years of age and older) are 383 mm (N = 33) in Indian Knoll, 373 mm (N = 59) in Pecos Pueblo, and 391 mm (N = 16) in Libben (Tape, 1992, see his Table I).
Moerman (1981) compared growth veloci- ties for 20 pelvic measurements between males and females for a 7 year period: the year of peak growth velocity (PGV) in stat- ure and 3 years preceding and succeeding the year of PGV. In males, the majority of pelvic measurements have growth velocity curves similar to that of stature. In females, however, many pelvic dimensions show sig- nificant growth in late adolescence extend- ing beyond that of stature (Moerman, 1981 [see her Table 6.111, 1982). These dimen- sions include: pubic length, ischium height, biiliac diameter, and the transverse diame- ters of the inlet and midplane (including bi- acetabular and bispinous diameters). Even for those pelvic dimensions in which both sexes share a pattern of late adolescent growth extending beyond that of stature (in- cluding breadth of sacrum, length of ischium [equivalent to anterior space of outlet in the present study], and transverse diameter of outlet), females show a greater amount of growth than do males. Of the pelvic mea- sures listed above, the present study demon- strates that only linea terminalis length (i.e., pubic length) differs significantly be- tween 18-24 year olds and those 25 years of age and older, and only females consistently evidence this difference. The implication is that, with the exception of the linea termi- nalis (and, correspondingly, the anterior space of the inlet), growth of the pelvis is essentially complete by age 18 in both sexes.
Moerman (1981, see her Fig. 6.9) showed that the age at PGV for the pubis coincides with that of stature in both sexes. PGV for both stature and the pubis occurs about 2 years earlier in females than males. During the year of PGV, the increase in pubic length is greater in females than males. Further- more, as stated above, the pubis continues to grow by a greater amount in females than males in the years following PGV.
While Moerman (1981) analyzed change in total length of the pubis, she did not dis- tinguish between growth at the acetabular and symphyseal borders. Coleman (1969), however, analyzed growth velocities for dif- ferent points on the pelvis. Coleman showed that, between the ages of 9 and 18, growth at all points on the pubis is greater in females than males, though the only statistically sig-

AGE AT DEATH AND PELVIC SIZE 37
nificant difference is at the superomedial point. Importantly, Coleman (1969, see his Fig. 15) showed that PGV at the superome- dial point of the pubis occurs absolutely later in females than males, the difference being about 4 years. Therefore, the implication is that PGV of the pubis occurs earlier in fe- males than males at the acetabular border3 but later in females than males a t the sym- physeal border. The asynchrony in growth between the acetabular and symphyseal borders of the pubis during adolescence im- plies that these growth centers respond dif- ferentially to e~t rogen .~ This differential re- sponse suggests that the osteoblasts at the symphyseal aspect of the pubis are rela- tively refractory to the growth-stimulating effects of estrogen. The later age of onset of PGV at the medial border of the pubis im- plies that the osteoblasts require either pro- longed exposure to estrogen or higher levels of estrogen to stimulate growth.
Not only is there asynchrony in PGV be- tween the acetabular and symphyseal bor- ders of the pubis, but there is also asyn- chrony in growth between the superior and inferior borders of the symphyseal aspect of the pubis. In contrast with the linea termi- nalis (which includes the superior border of the pubis), the anterior space of the outlet (which includes the inferior border of the pubis) does not show a consistent, statisti- cally significant difference in length be- tween younger and older adults in either sex (Table 2). The implication is that growth at the inferior border of the pubis is generally completed by age 18 in both sexes, whereas growth at the superior border continues into the third decade of life in females. This in-
3As PGV at the medial border of the pubis occurs later in females than males (Coleman, 19691, the sexual difference re- ported by Moerman (1981) in PGV of the pubis as a whole must have occurred principally at the acetabular border.
4Crelin and colleagues (Crelin, 1960,1969; Crelin and Blood, 1961; Bernstein and Crelin, 1967) demonstrated in rats, mice, and rabbits that sexual dimorphism of the pelvis is due to the effects of testosterone, not estrogen (but see Vesugi et al., 1992). The results of studies on humans (Greulich and Thoms, 1939; Morton and Gordan, 1952; Morris and Mahesh, 1963) are equiv- ocal as to the relative influence of testosterone and estrogen on pelvic dimorphism. However, because PGV of the pubis is syn- chronized with that of stature in both sexes (Moerman, 1981 ), the implication is that the pubis does respond to both steroids with accelerated growth.
terpretation is supported by Coleman’s (1969: 141) observation that for both sexes “at the inferior border of the pubic symphy- sis . . . there is a larger amount of total growth [between the ages of 9 and 181 than . . . [at] the superior border of the pubic symphysis.”
The argument that the pubis continues to grow in early adulthood in females, but not males, can be used to explain the develop- mental basis for sexual dimorphism in the ventral arc of the pubis. Budinoff and Tague (1990) argued that the periosteal attach- ment for the gracilis and adductor brevis muscles is replaced by an osseous attach- ment in late adolescence in both sexes. In females, continued growth of the pubis at its symphyseal border “laterally displaces” the site of attachment for these muscles, thereby creating the ventral arc. Todd (1920) suggested that growth of the pubis in males ceases at approximately age 18, and “the prominence of the ventral rampart [see discussion below] in the female suggests more vigorous growth or a longer period of growth at the symphysis” (Todd, 1921a: 38).
Two explanations are offered as to the se- lective advantage of a later age at matura- tion of the pubis in females than males. First, in females the asynchrony in PGV be- tween the acetabular and symphyseal bor- ders of the pubis extends the period of growth. This prolonged growth of the pubis, coupled with more vigorous growth in fe- males than males, results in sexual dimor- phism in pubic length and, correspondingly, linea terminalis length and inlet circumfer- ence. Consequently, sexual dimorphism in pubic length results from differences be- tween the sexes in both the duration and magnitude of growth. Second, late matura- tion of the symphyseal border of the pubis may be a general phenomenon in primates. The interpubic joint synostoses in some pri- mate species, but not in humans (Todd, 1921b; Washburn, 1942; Schultz, 1944; Tague, 1990, 1993); the sacroiliac joint rarely fuses in nonhuman primates (pers. obs.). In nonhuman primates, synostosis of the interpubic joint results from fusion of the two medial epiphyses of the pubes to the pubic corpora and to each other (Todd, 1921b; Tague, 1993). Pubic symphyseal

38 R.G. TAGUE
synostosis not only obviates pregnancy-re- lated mobility a t the interpubic joint, but also restricts mobility at the sacroiliac joints. In mice, inhibition of pelvic joint mo- bility results in an increase in maternal and fetal mortality rates (Gardner and Van Heu- verswyn, 1940). A selective advantage to late maturation of the pubis is that synosto- sis of the interpubic joint is delayed. Among some mammals, elevated levels of estrogen (such as during pregnancy) induce resorp- tion of the pubis at its symphyseal border (for a review and references, see Tague, 1988). Therefore, if age at first pregnancy precedes maturation of the pubis, then the osteoclastic effects of estrogen may disrupt fusion of the medial epiphyses and, thereby, prevent synostosis of the interpubic joint. Although humans do not have a separate epiphysis at the symphyseal border of the pubis, the ventral rampart of the pubis is considered to be a delayed and “retrogres- sive” epiphysis (Todd, 1921c; Meindl et al., 1985b). The rampart represents an accre- tion of bone to the ventral demiface of the pubis. Rampart formation commences in the third decade of life and is typically com- pleted in the fourth decade (Todd, 1920, 1921a; Meindl et al., 1985b; Brooks and Suchey, 1990). In humans, therefore, forma- tion of the medial epiphysis of the pubis (i.e., the ventral rampart) typically occurs subse- quent to a female’s first pregnancy among natural fertility populations.
The discussion above illustrates that the symphyseal aspect of the pubis has an inter- esting biology. For the acetabular border of the pubis and for other bones, females ma- ture earlier than males (Krogman and Igcan, 1986, see their Tables 3.10 and 3.11). However, maturation of the symphyseal as- pect of the pubis occurs later in females than males. As stated above, the implication is that the osteoblasts at the symphyseal bor- der of the pubis are relatively refractory to low levels of estrogen. Conversely, high lev- els of estrogen induce resorption of the pubis at its symphyseal border; other bones in- crease in density upon exposure to estrogen (Gardner and Pfeiffer, 1938; Silberberg and Silberberg, 1971). Consequently, not only must different bones be evaluated as indi- vidual tissues subject to natural selection, but even loci within the same bone (e.g., the
pubis) may become differentiated as a result of selection.
CONCLUSIONS This study shows that the linea terminalis
is significantly shorter in younger females (18-24 years of age) relative to older females (25 years of age and older) in all three sam- ples of prehistoric Amerindians. Two expla- nations are offered-maternal mortality associated with a confined pelvic inlet cir- cumference and later age at maturation of the pubis in females than males. This study cannot provide a definitive answer as to which explanation is correct. This author, however, regards the studies by Coleman (1969) and Moerman (1981) as providing strong support for the latter explanation, that is, growth of the pubis is prolonged in females relative to males. Nevertheless, ma- ternal mortality was likely to have been a significant factor in prehistoric demogra- phy. High rates of maternal mortality do characterize some populations that lacked modern obstetric care. Although continued growth of the pubis in females but not males better explains the results of this study, the results also show that pelvic inlet circumfer- ence was relatively confined among young adult females. The implication is that birth among these females was probably difficult.
ACKNOWLEDGMENTS I thank the following individuals and in-
stitutions for allowing me to study skeletal material in their care: Lathe1 Duffield, Uni- versity of Kentucky; C. Owen Lovejoy, Kent State University; David Pilbeam, Harvard University. C. Owen Lovejoy generously provided his age estimates based on dental wear for the Libben sample. I thank Linda Budinoff for the artwork and Judy Suchey for other assistance in the preparation of the manuscript. I am grateful to Karen Rosen- berg and Marquisa LaVelle for providing helpful suggestions on a previous draft of the manuscript. Sigma Xi provided partial funding for the research.
LITERATURE CITED Aberle SBD (1934) Maternal mortality among the Pueb-
los. Am. J. Phys. Anthropol. 18(old series):431435. Abramson D, Roberts SM, and Wilson PD (1934) Relax-
ation of the pelvic joints in pregnancy. Surg. Gynecol. Obstet. 58t595-613.

39 AGE AT DEATH AND PELVIC SIZE
Allbrook D, and Sibthorpe EM (1961) A study of pelvic dimensions related to infant size in the Ganda of East Africa. S. Afr. J . Med. Sci. 26:73-83.
Arriaza B, Allison M, and Gerszten E (1988) Maternal mortality in pre-Columbian Indians of Arica, Chile. Am. J . Phys. Anthropol. 77~35-41.
Bernstein P, and Crelin ES (1967) Bony pelvic sexual dimorphism in the rat. Anat. Rec. 157517626.
Bore11 U, and Fernstrom I(1957) The movements a t the sacro-iliac joints and their importance to changes in the pelvic dimensions during parturition. Acta Obstet. Gynecol. Scand. 36:42-57.
Brooks S, and Suchey JM (1990) Skeletal age determi- nation based on the 0s pubis: A comparison of the Acsadi-Nemeskeri and Suchey-Brooks methods. Hum. Evol. 5:227-238.
Budinoff LC, and Tague RG (1990) Anatomical and de- velopmental bases for the ventral arc of the human pubis. Am. J. Phys. Anthropol. 82:73-79.
Coleman WH (1969) Sex differences in the growth of the human bony pelvis. Am. J. Phys. Anthropol. 31t125- 152.
Crelin ES (1960) The development of bony pelvic sexual dimorphism in mice. Ann. N.Y. Acad. Sci. 84:479-512.
Crelin ES (1969) The development of the bony pelvis and its changes during pregnancy and parturition. Trans. N.Y. Acad. Sci. 31:1049-1058.
Crelin ES, and Blood DK (1961) The influence of the testes on the shaping of the bony pelvis in mice, Anat. Rec. 140:375-379.
Dobbie BMW (1982) An attempt to estimate the true rate of maternal mortality, sixteenth to eighteenth centuries. Med. Hist. 26:79-90.
Eccles A (1982) Obstetrics and Gynaecology in Tudor and Stuart England. Kent, OH: Kent State University Press.
Ellison PT (1990) Human ovarian function and repro- ductive ecology: New hypotheses. Am. Anth. 92:933- 952.
Eveleth PB, and Tanner JM (1990) Worldwide Variation in Human Growth. New York: Cambridge University Press.
Flander LB (1978) Univariate and multivariate meth- ods for sexing the sacrum. Am. J. Phys. Anthropol. 49:103-110.
Gardner WLT, and Pfeiffer CA (1938) Skeletal changes in mice receiving estrogens. Proc. SOC. Exp. Biol. Med. 37:678-679.
Gardner WU, and Van Heuverswyn J (1940) Inhibition of pelvic changes occurring during pregnancy in mice by testosterone propionate. Endocrinology 26t833- 836.
Greulich WW, and Thoms H (1939) An x-ray study of male pelves. Anat. Rec. 75:289-305.
Greulich WW, and Thoms H (1944) The growth and development of the pelvis of individual girls be- fore, during and after puberty. Yale J . Biol. Med. 17:
Hager LD (1989) The Evolution of Sex Differences in the Hominid Bony Pelvis. Ph.D. dissertation, University of California, Berkeley.
Heyman J , and Lundqvist A (1932) The symphysis pubis in pregnancy and parturition. Acta Obstet. Gynecol. Scand. 12:191-226.
91-97.
Hooton EA (1930) The Indians of Pecos Pueblo: A Study of Their Skeletal Remains. New Haven: Yale Univer- sity Press.
Howell N (1979) Demography of the Dobe !Kung. New York: Academic Press.
Johnston FE (1961) Sequence of epiphyseal union in a prehistoric Kentucky population from Indian Knoll. Hum. Biol. 33~66-81.
Krogman WM, and Igcan MY (1986) The Human Skele- ton in Forensic Medicine. Springfield, IL: Charles C. Thomas.
Lazenby RA (1990) Continuing periosteal apposition I: Documentation, hypotheses, and interpretation. Am. J. Phys. Anthropol. 82:451-472.
Lovejoy CO (1985) Dental wear in the Libben popula- tion: Its functional pattern and role in the determina- tion of adult skeletal age at death. Am. J. Phys. An- thropol. 82:47-56.
Lovejoy CO, Meindl RS, Pryzbeck TR, Barton TS, Heiple KG, and Kotting D (1977) Paleodemography of the Libben site, Ottawa County, Ohio. Science 198:291- 293.
Lovejoy CO, Meindl RS, Mensforth RP, and Barton T J (1985a) Multifactorial determination of skeletal age at death: A method and blind tests of its accuracy. Am. J. Phys. Anthropol. 68:l-14.
Lovejoy CO, Meindl RS, Pryzbeck TR, and Mensforth RP (1985b) Chronological metamorphosis of the auricular surface of the ilium: A new method for the determina- tion of adult skeletal age at death. Am. J. Phys. An- thropol. 68:15-28.
McKern TW, and Stewart TD (1957) Skeletal Age Changes in Young American Males. Analysed from the Standpoint of Age Identification. Technical Re- port EP-45. Natick, MA: Quartermaster Research and Development Center, U.S. Army, Environmental Pro- tection Research Division.
Meigs GL (1917) Maternal mortality from all conditions connected with childbirth in the United States and certain other countries. In R Bremmer (ed.) (1974): Children’s Bureau Studies. New York: Arno Press.
Meindl RS, Lovejoy CO, Mensforth RP, and Don Carlos L (1985a) Accuracy and direction of error in the sexing of the skeleton: Implications for paleodemography. Am. J. Phys. Anthropol. 68:79-85.
Meindl RS, Lovejoy CO, Mensforth RP, and Walker RA (198513) A revised method of age determination using the 0s pubis, with a review and tests of accuracy of other current methods of pubic symphyseal aging. Am. J . Phys. Anthropol. 68:29-45.
Meredith HV (1950) Birth order and body size. Am. J. Phys. Anthropol. 8:195-224.
Moerman ML (1981) A Longitudinal Study of Growth in Relation to Body Size and Sexual Dimorphism in the Human Pelvis. Ph.D. dissertation, University of Mich- igan.
Moerman ML (1982) Growth of the birth canal in adoles- cent girls. Am. J. Obstet. Gynecol. 143528-532.
Morris JL, and Mahesh VB (1963) Further observations on the syndrome, “testicular feminization.” Am. J . Ob- stet. Gynecol. 87:731-745.
Morton DG, and Gordan G (1952) Observations upon the role of sex hormones in the development ofbony pelvic conformation. Am. J. Obstet. Gynecol. 64:292-300.

40 R.G. TAGUE
Ohlsen H (1973) Moulding of the pelvis during labour. Acta Radiol. Diagn. 14t417434.
Owsley DW, and Bradtmiller B (1983) Mortality of preg- nant females in Arikara villages: Osteological evi- dence. Am. J. Phys. Anthropol. 61:331-336.
Phenice TW (1969) A newly developed visual method of sexing the 0s pubis. Am. J . Phys. Anthropol. 30:297- 302.
Russell JGB (1969) Moulding of the pelvic outlet. J . Ob- stet. Gynaecol. Br. Cmwlth. 76:817-820.
Schultz AH (1944) Age changes and variability in gib- bons. Am. J. Phys. Anthropol. 2(new series):l-l29.
Schultz AH (1949) Sex differences in the pelves of pri- mates. Am. J . Phys. Anthropol. 7:401423.
Segebarth-Orban R (1980) An evaluation of the sexual dimorphism of the human innominate bone. J. Hum. Evol. 9:601-607.
Sherwood OD (1988) Relaxin. In E Knobil and J D Neil1 (eds.): The Physiology of Reproduction, Vol. 1. New York: Raven Press, pp. 5 8 H 7 3 .
Shorter E (1982) A History of Women’s Bodies. New York Basic Books.
Silberberg M, and SilberbergR (1971) Steroid hormones and bone. In GH Bourne (ed.): The Biochemistry and Physiology of Bone, Vol. 111. New York: Academic Press, pp. 401484.
Straus WL, J r . (1927) The human ilium: Sex and stock. Am. J . Phys. Anthropol. 11:l-28.
Tague RG (1988) Bone resorption of the pubis and preauricular area in humans and nonhuman mam- mals. Am. J. Phys. Anthropol. 76.251-267.
Tague RG (1990) Morphology of the pubis and preauric- ular area in relation to parity and age a t death in Macaca mulatta. Am. J. Phys. Anthropol. 82.517-525.
Tague RG (1992) Sexual dimorphism in the human bony pelvis, with a consideration of the Neandertal pelvis from Kebara cave, Israel. Am. J . Phys. Anthropol. 88: 1-21.
Tague RG (1993) Pubic symphyseal synostosis and sex- ual dimorphism of the pelvis in Presbytis cristata and Presbytis rubicunda. Int. J . Primatol. 14:637454.
Thorp DJ, and Fray WE (1938) The pelvic joints during pregnancy and labor. JAMA 111:1162-1166.
Todd TW (1920) Age changes in the pubic bone: I. The male White pubis. Am. J . Phys. Anthropol. 3t285-334.
Todd TW (1921a) Age changes in the pubic bone: 11. The pubis of the male Negro-White hybrid. 111. The pubis of the White female. IV. The pubis of the female Ne- gro-White hybrid. Am. J . Phys. Anthropol. 4:l-70.
Todd TW (1921b) Age changes in the pubic bone. V. Mammalian pubic metamorphosis. Am. J. Phys. An- thropol. 4:333-406.
Todd TW (1921~) Age changes in the pubic bone: VI. The interpretation of variations in the symphyseal area. Am. J. Phys. Anthropol. 4:407-424.
Todd TW, and DErrico J, Jr. (1928) The clavicular epi- physes. Am. J . Anat. 41:2560.
Vesugi Y, Taguchi 0, Noumura T, and Iguchi T (1992) Effects of sex steroids on the development of sexual dimorphism in mouse innominate bone. Anat. Rec. 234541648,
Washburn SL (1942) Skeletal proportions of adult lan- gurs and macaques. Hum. Biol. 143444472,
Washburn SL (1948) Sex differences in the pubic bone. Am. J. Phys. Anthropol. 6t199-207.
Washburn SL (1949) Sex differences in the pubic bone of Bantu and Bushman. Am. J. Phys. Anthropol. 7:425- 432.
Webb WS (1974) Indian Knoll. Knoxville: University of Tennessee Press.
Webb PAO, and Suchey JM (1985) Epiphyseal union of the anterior iliac crest and medial clavicle in a modern multiracial sample of American males and females. Am. J . Phys. Anthropol. 68:457-466.
Weiss KM (1973) Demographic models for anthropology. Memoirs of the Society for American Archaeology, No. 27, Am. Antiquity Vol. 38, No. 2.
Wells C (1975) Ancient obstetric hazards and female mortality. Bull. N.Y. Acad. Med. 51:1235-1249.
Williams PL, and Warwick R (1980) Gray’s Anatomy, 36th British ed. Philadelphia: W.B. Saunders.
Wood JW (1989) Fecundity and natural fertility in hu- mans. In SR Milligan (ed.): Oxford Reviews of Repro- ductive Biology, Vol. 11. New York: Oxford University Press, pp. 61-109.















![Major Amerindian Cultures [Inca]](https://img.pdfslide.net/doc/110x75/58ef640f1a28ab6f048b45bf/major-amerindian-cultures-inca.jpg)