Embed Size (px)
Citation preview

Vol. 89, Part 3 [ 2 85 ] October 1987
Trans Br. mycol. Soc. 89 (3), 285-294 (1987) Printed in Great Britain
MATING BEHAVIOUR IN AN ISOLATE OF PHANEROCHAETECHR Y SOSPORIUM
By WENDY THOMPSON* AND PAUL BRODADept of Biochemistry and Applied Molecular Biology, UMIST, PO Box 88, Manchester M60 lQD
MATERIALS AND METHODS
The five heterokaryotic isolates of P. chrysosporiumused are listed in Table 1. These had beenmaintained on slopes of 2% (w/v) malt extract,1'5 % (wIv) agar, grown initially at 37°C and thenheld at 4°, and subcultured every three months.Conidia from slopes were transferred to 9 em diamPetri dishes containing IS ml Walseth cellulose(Gold & Cheng, 1979). The dishes were theninverted and incubated at 28° for between two andeight weeks.
cultures derived from single basidiospores mayproduce fruit bodies and basidiospores in culturehave been taken as indirect evidence of homothallism.
A method has been published which describesthe conditions necessary to induce fruit bodyformation by P. chrysosporium (Gold & Cheng,1979). Gold et al. (1980) stated that crosses madefrom colonies produced from individual basidiospores should determine whether the species isheterothallic or homothallic. Perhaps one reasonwhy this has not been done is that P. chrysosporiumdoes not produce clamps on its primary orsecondary mycelia. It is therefore not as easy toidentify secondary mycelia in this species as it is insome other basidiomycetes. In Schizophyllum commune Fr., for example, the primary mycelium doesnot form clamps but the secondary mycelium doesand is therefore easily distinguished. The primaryand secondary mycelia of P. chrysosporium aredifferent in gross morphology but it is very difficultto distinguish them reliably. This has necessitatedan analysis of the mating behaviour of P. chrysosporium similar to those employed by Coates et al.(1981) for Stereum hirsutum (Willd.: Fr.) S. F.Gray and Boddy & Rayner (1982) for S. gausapatum (Fr.) Fr. in both of which the primary andsecondary mycelia possess clamp connexions.
* To whom all correspondence should be sent.
~ttempts were made to produce basidiocarps of five isolates of Phanerochaete chrysosporiumIn pure culture. Only one isolate formed fruit bodies with basidiospores and the latter wereused to analyse th~i~ mating behaviour. A wide range of responses was observed in pairingsbetween monobasidiosporous cultures and the isolate was found to possess a unifactorialhomogenic incompatibility system.
Phanerochaete chrysosporium Burdsall is a thermotolerant, white rotting, saprobic basidiomycete. Itis an important experimental organism, used instudies of the biodegradation of lignin, e.g. Keyseret al. (1978), Fenn & Kirk (1979), Jeffries et al.(1981), Eriksson et al. (1983) and Tien & Kirk(1984) and the production of protein from lignocellulosic waste material, e.g, Ek & Eriksson(1980).
It differs from other Phanerochaete specieswhich have been studied in culture because it hasa Sporotrichum puluerulentum Novobranova anamorph (Burdsall, 1981; Stalpers, 1984). BecauseP. chrysosporium produces conidia prolifically onboth homokaryotic and heterokaryotic mycelia andcertain isolates produce basidospores in culture(Burdsall & Eslyn, 1974; Gold & Cheng, 1979) itcan be relatively easily manipulated in physiologicaland genetical studies. Despite these positive attributes, the mating system of this species has not yetbeen investigated thoroughly, even though anumber of studies have been concerned withproducing mutants and maintaining forced heterokaryons (Gold et al., 1982; Alic & Gold, 1985;Johnsrud & Eriksson, 1985). In order to interpretproperly the results and implications of geneticalexperiments it is essential that the basic life cycle,including the mating system, of the organism iselucidated.
There have been several reports that P. chrysosporium is primary homothallic (Eriksson et al.,.1978; Alic & Gold, 1985) and according to Boidin(1971) holocoenocytic (plurinucleate) fungi such asPhanerochaete spp., are invariably homothallic.However, such reports have not been based oncareful analyses of mating behaviour. Rather,observations such as the lack of clamp connexions,holocoenocytic nuclear behaviour and the fact that
Vol. 89, Part 2, was issued on 1 September, 198711 MYC89

286 Mating behaviour in Phanerochaete chrysosporium
Table 1. Isolates used in analysis of mating behaviour in Phanerochaete chrysosporium
P. chrysosporium(Lombard ME 446)
P. chrysosporium(Nicot Elphick)
P . chry sosporium(White)
P. chrysosporium(Burdsall)
P. chrysosporium(previously Sporotrichumpuluerulentum Novobranova(Beliakovaj).
OriginU.S.A.
France
Papua-New Guinea
U.S.A.
U .S.S.R.
Culture collectionreferenceATCC 34541
IMI74691
IMI Il0I20
IMI232175
ATCC24725
Only one of the P. chrysosporium isolates (ME446) formed basidocarps and basidiospores underthe conditions described above. The other fourisolates could not be induced to fruit even thoughseveral different media were used, e.g. tap wateragar, 0'1 % (w/ v) malt extract agar, media incorporating sawdust or wood shavings from beech(Fagus syluatica L.) and pine (Pinus sylvestris L.)and sterile beech and pine shavings. Neitherfluctuations in temperature nor different light/dark regimes induced fruiting in these four isolates.
Single basidiospores from P. chrysosporiumME 446 were isolated as described in Raeder &Broda (1986).
Duplicate pairings were made between all combinations of monobasidiospore isolates derivedfrom isolate ME 446, the five isolates of P.chrysosportum in Table I, synthesized heterokaryons and also between selected monosporeisolates and secondary mycelia. These were doneusing 9 em diam Petri dishes containing 15 ml z %(wIv) malt extract, 1'5 % (wI v) agar. Discs of agarand mycelium were cut with a 7 mm cork borerfrom the margins of actively growing colonies andplaced 1'5-2 cm apart in pairings between monospore isolates and 2 ern apart in pairings betweensecondary mycelia, and secondary mycelia andmonospore isolates.
All pairings were incubated, lids uppermost, at30° in the dark and examined every two days for upto fourteen days, by which time all interactions haddeveloped fully and autolysis of cultures wasbeginning. Selected pairings were also incubated at37° as this is close to the optimum temperature(40°) for P. chrysosporium grown in culture (Burdsall & Eslyn, 1974). However, identical resultswere obtained for these pairings at 30 and 37° andtherefore to simplify the observation and recordingof results all further crosses were incubated at30°.
As reliable indicators of secondary mycelia werenot available it was necessary to perform a numberof further tests on the monospore pairings in orderto interpret the initial results. These involved subculturing from different regions of the interactionplates and examining the resulting colony morphologies, and also back pairing against the originalmonospore isolates.
RESUL TS AND DISCUSSION
Because only one of the five isolates tested (ME466) gave rise to any basidiospores in culture it wasonly possible to carry out sib-pairings between themonosporous progeny from P. chrysosporium ME466. The results of all pairings are summarized inTable 2 and examples of selected pairings areshown in Figs 1-5.
On the basis of their interactive behaviour thetwelve monospore isolates from P. chry sosporiumME 446 fell into two incompatibility groups(T able 3). Generally isolates within each groupwere incompatible with each other but compatiblewith members of the other group. There weresome exceptions to this, however, which cannot yetbe explained. For example, monospore 18 wascompatible with every other isolate with which itwas paired and cultures derived from five singleconidia of isolate 18 always gave the same result.Such exceptions were perhaps not surprising as thecriteria used for secondary mycelium identificationwere subjective.
Conidia from the five heterokaryotic isolateslisted in Table I, and from four selected monosporeisolates, were fixed in 5 % (vIv) glutaraldehyde(T aab, Reading, U.K.) in 0'1 M Tris/HCI buffer,pH7, for 30 min at room temperature. 100 pi ofthis solution was mixed, on a microscope slide,with an equal volume of 300 ng ml' DAPI (4,6Diamidino-z-phenylindole) (Sigma, U.K.) in 0'1M Tris/HCI buffer pH7, and kept in the dark for 5-

Wendy Thompson and P. Broda
10 min (Lin et al., 1977). The conidia were thenobserved with a Leitz Dialux 20EB fluorescencemicroscope.
DAPI-staining of conidia revealed that 90-94 %of conidia from the five natural isolates werebinucleate and 70-78 % of conidia from themonobasidiosporous cultures were binucleate.
In contrast to previous reports, which havesuggested that P. chrysosporium is probably homothallic (Eriksson et al., 1978; Alic & Gold, 1985),the present investigation shows that isolate ME446 has a homogenic incompatibility system underunifactorial control, i.e. it has bipolar heterothallism/unifactorial diaphoromixis.
The wide range of responses in pairings betweencombinations of compatible and incompatiblemonospore isolates (Figs 1, 2) can be interpretedin terms of the three fundamental functions whichRayner et al. (1984) have associated with secondarymycelium formation in basidiomycetes. These areaccess migration, acceptor migration and stabilization of the secondary mycelium. On this interpretation the formation of secondary myceliumresulted from access and/or acceptor migrationfollowed by stabilization. Deadlock and antagonistic interactions involving no overgrowth of oneisolate into another were incompatible with noaccess migration occurring.
The lytic interaction which developed aftersome unilateral encroachment of one isolate intoanother can be explained in terms of restrictedaccess function, and' blotchy' cultures may be themanifestations of unstabilized mycelia after accessand acceptor migration had occurred.
The box delimited by the dashed lines in Table3 includes paired isolates, most of which showbehaviour characteristic of restricted access orrestricted acceptor function. This observation maybe significant to any interpretation of the functionof the homogenic incompatibility system in P.chrysosporium. In Stereum hirsutum, for example,there is evidence of a multiallelic factor (the Bfactor) which is a homogenic incompatibility locusgoverning access migration and which may behomologous with the B-factor in tetrapolar species(Coates & Rayner, 1985a), e.g. Coprinus cinereusBeeli, in which the B factor controls fertility andnuclear migration (Kemp, 1980). It is thought thatthe compatibility locus (C-factor) in S. hirsutumalso has a component which allows access (Coates& Rayner, 1985a) and also that the access component of the C-factor is transposable and capableof being switched in a way that may be analagousto the cassette system in yeasts (Coates & Rayner,1985 b).
The compatibility of monospore isolate 18 withevery other monospore isolate with which it was
paired, could indicate that recombination hadoccurred within its mating type locus. Intrafactorrecombination has been demonstrated in Flammulina velutipes (Curtis: Fr.) Singer and Pleurotusostreatus (Jacquin) Kummer (Fincham et al.,1979), but not in a bipolar species. In thosematings involving isolate 18 in which unilateraldevelopment of secondary mycelium occurred,isolate 18 was always the mate which was migratedinto. The promiscuous isolate 18 must hold cluesto the control of homogenic incompatibility in thisspecies, but has not yet been further characterized.
It was unfortunate that four of the five isolates ofP. chrysosporium would not fruit in culture. It isimportant that attempts to obtain fruiting in culturecontinue or that collections be made of monosporeisolates from naturally occurring sporophores.Data from analyses of the homogenic incompatibility systems of these additional isolates of P.chrysosporium must be obtained in order to determine whether the types of interactive behaviourobserved for isolate ME 446 are typical of thespecies. Other isolates may differ in their matingbehaviour, especially if they originate from widelyseparated locations. It is known for example thatthe British population of S. hirsutum has a unifactorial homogenic incompatibility system. Incontrast the Finnish population appears to be nonoutcrossing (Rayner et al., 1984), these populationsbeing considered as members of the same specieson grounds of morphology.
Johnsrud & Eriksson (1985) observed that bothhomokaryotic and heterokaryotic basidiosporeswere produced in culture by P. chrysosporium K-3(derived from a culture of S. puluerulentum P-127I, Novobranova, ATCC 32629). Although all theirbasidiospores were derived from 4-spored basidia,Johnsrud & Eriksson's observation contrasts withour own data which suggest that all basidiosporesproduced by P. chrysosporium ME 446 were homokaryotic. Our data are further supported by thedemonstration by Raeder & Broda (1986), whoused restriction fragment length polymorphism asa criterion, that cultures derived from twelve singleconidia of secondary mycelium of ME 446 wereheterokaryotic, whilst cultures derived from 53single basidiospores (including the twelve used inthis study) from ME 446 were homokaryotic.Although we could not be certain of the origin ofour basidiospore-derived cultures (whether theywere from basidia bearing any number of sterigmata and basidiospores between 2 and 8), the dataof Raeder & Broda (1986) demonstrate conclusivelythat our isolates are homokaryotic.
The only other Phanerochaete species whosemating behaviour has been studied, are P. velutina(DC.) Parmasto and P. laevis (Fr.) Erikss. & Ryv.,
11-2

Table 2. Summary of pairings in culture between mono-basidiosporous and secondary mycelia
N0000
Paired isolates
A.Mono-basidiosporous
isolates (78 diff. pairings)
B.Synthesized secondarymycelia (15 diff.pairings)
Resultant morphology
(a ) Uniform intermingling of isolates
(b) (i) Encroachment of one isolate intothe other
(ii) Encroachment of one isolateinto the other followed byantagonism (pigment production)along the advancing edge ofencroachment region
(c) Formation of a band of varying width(2-10 mm), marked by heavyconidiation, between the paired isolatesNarrow (up to I mm), yellowpigmented line extending centrallyalong band (when plates viewedfrom below)
(d) (i) Deadlock ; paired isolates simplyceased growing on contact with oneanother(ii) Deadlock followed by development of lytic zone on one side of theinteraction zone
(e) 'Blotchy' cultures ; patches of twomorphologically distinct types ofmycelia throughout plates
(a ) Intermingling
(b) Narrow lytic zones (weak lytic zones(weak antagonism) between pairedisolates)
Further tests to help characterize interaction
On subculturing morphology is identical with that oforiginally paired isolates
On subculturing morphology consistently different fromthat of originally paired isolates
On subculturing from the encroachment region resultantmorphology is persistent and consistently different fromthat of originally paired isolates
On subculturing, colonies resemble one or other of theoriginally paired isolates
On back-pairing with original mates get interminglingwith one or other of these
On subculturing from either side of the interaction zone(band) and re-pairing these get same type of interaction
When paired with the originally paired mates the subcultured isolates intermingled with one or other of theformer
On subculturing isolates intermingle with one or other oforiginally paired mates
On subculturing from lytic zone cultures havemorphologies resembling one or other of originallypaired mates
On subculturing from either type of mycelium culturesresemble one or the other of originally paired matesOn pairing with the latter the subcultured coloniesintermingle with one or other of them
On subculturing morphology is identical with that oforiginally paired isolates
On subculturing from lytic zone morphologies resembleone or the other of originally paired isolates
Interaction type
Self-Self
Compatible, i.e. secondarymycelium development
Compatible
Incompatible
Incompatible
Incompatible
Incompatible
Incompatible
Self-self
Antagonism
~....~'
0n:.::to~ec'l::'-t
~'
d2~(ll"'1o
fr(ll...(ll
g.~00o00
"C:Io"'1
a'S
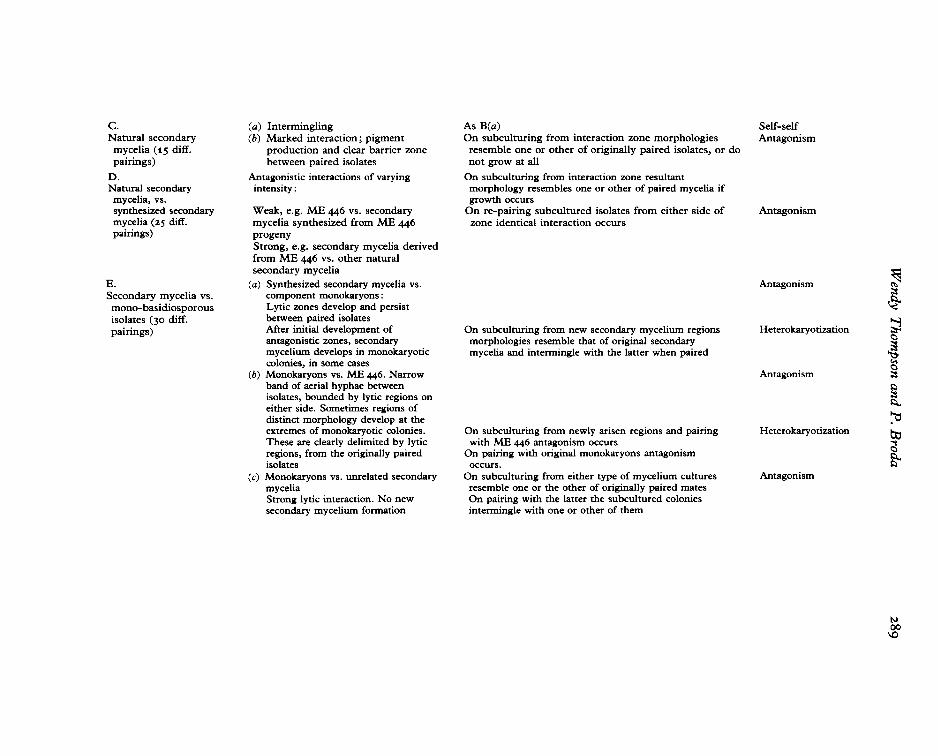
c.Natural secondary
mycelia (15 diff.pairings)
D.Natural secondarymycelia, vs.synthesized secondarymycelia (25 diff.pairings)
E.Secondary mycelia vs.mono-basidiosporousisolates (30 diff.pairings)
(a) Intermingling(b) Marked interaction; pigment
production and clear barrier zonebetween paired isolates
Antagonistic interactions of varyingintensity :
Weak, e.g. ME 446 vs. secondarymycelia synthesized from ME 446progenyStrong, e.g. secondary mycelia derivedfrom ME 446 vs. other naturalsecondary mycelia
(a) Synthesized secondary mycelia vs,component monokaryons:Lytic zones develop and persistbetween paired isolatesAfter initial development ofantagonistic zones, secondarymycelium develops in monokaryoticcolonies, in some cases
(b) Monokaryons vs. ME 446. Narrowband of aerial hyphae betweenisolates, bounded by lytic regions oneither side. Sometimes regions ofdistinct morphology develop at theextremes of monokaryotic colonies.These are clearly delimited by lyticregions, from the originally pairedisolates
(c) Monokaryons vs, unrelated secondarymyceliaStrong lytic interaction. No newsecondary mycelium formation
As B(a)On subculturing from interaction zone morphologiesresemble one or other of originally paired isolates, or donot grow at all
On subculturing from interaction zone resultantmorphology resembles one or other of paired mycelia ifgrowth occurs
On re-pairing subcultured isolates from either side ofzone identical interaction occurs
On subculturing from new secondary mycelium regionsmorphologies resemble that of original secondarymycelia and intermingle with the latter when paired
On subculturing from newly arisen regions and pairingwith ME 446 antagonism occurs
On pairing with original monokaryons antagonismoccurs.
On subculturing from either type of mycelium culturesresemble one or the other of originally paired matesOn pairing with the latter the subcultured coloniesintermingle with one or other of them
Self-selfAntagonism
Antagonism
Antagonism
Heterokaryotization
Antagonism
Heterokaryotization
Antagonism
~;:l
%~c~~§~
~b::I""tC
~
N00\0
Mating behaviour in Phanerochaete chrysosporium
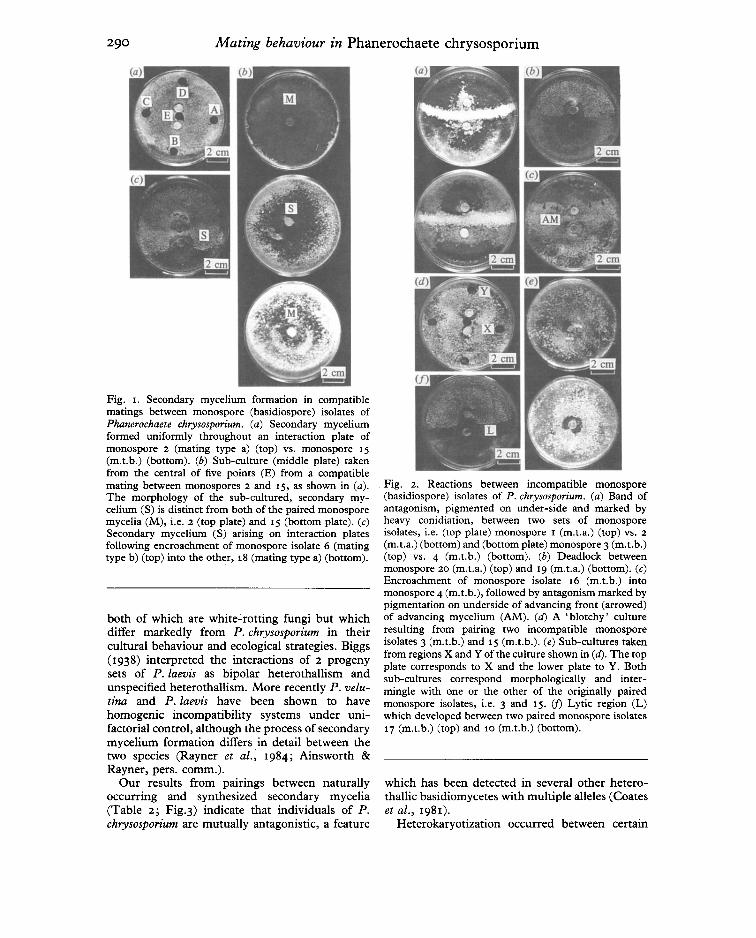
Fig. I. Secondary mycelium formation in compatiblematings between monospore (basidiospore) isolates ofPhaneroehaete ehrysosporium. (a) Secondary myceliumformed uniformly throughout an interaction plate ofmonospore 2 (mating type a) (top) vs. monospore 15(m.t.b.) (bottom). (b) Sub-culture (middle plate) takenfrom the central of five points (E) from a compatiblemating between monospores 2 and 15, as shown in (a).The morphology of the sub-cultured, secondary mycelium (S) is distinct from both of the paired monosporemycelia (M), i.e, 2 (top plate) and 15 (bottom plate). (e)Secondary mycelium (S) arising on interaction platesfollowing encroachment of monospore isolate 6 (matingtype b) (top) into the other, 18 (mating type a) (bottom).
both of which are white-rotting fungi but whichdiffer markedly from P. chrysosporium in theircultural behaviour and ecological strategies. Biggs(1938) interpreted the interactions of 2 progenysets of P. laevis as bipolar heterothallism andunspecified heterothallism. More recently P. velutina and P. laevis have been shown to havehomogenic incompatibility systems under unifactorial control, although the process of secondarymycelium formation differs in detail between thetwo species (Rayner et ai., 1984; Ainsworth &Rayner, pers. comm.).
Our results from pairings between naturallyoccurring and synthesized secondary mycelia(Table 2; Fig.g) indicate that individuals of P.chrysosporium are mutually antagonistic, a feature
Fig. 2. Reactions between incompatible monospore(basidiospore) isolates of P. ehrysosporium. (a) Band ofantagonism, pigmented on under-side and marked byheavy conidiation, between two sets of monosporeisolates, i.e. (top plate) rnonospore I (m.t.a.) (top) vs. 2
(m.t.a.) (bottom) and (bottom plate) monospore 3 (m.t.b.)(top) vs. 4 (m.t.b.) (bottom). (b) Deadlock betweenmonospore 20 (rn.t.a.) (top) and 19 (m.t.a.) (bottom). (e)Encroachment of monospore isolate 16 (m.t.b.) intomonospore 4 (m.t.b.), followed by antagonism marked bypigmentation on underside of advancing front (arrowed)of advancing mycelium (AM). (d) A 'blotchy' cultureresulting from pairing two incompatible monosporeisolates 3 (m.t.b.) and 15 (m.t.b.), (e) Sub-cultures takenfrom regions X and Y of the culture shown in (d). The topplate corresponds to X and the lower plate to Y. Bothsub-cultures correspond morphologically and intermingle with one or the other of the originally pairedmonospore isolates, i.e. 3 and 15. if) Lytic region (L)which developed between two paired monospore isolates17 (m.t.b.) (top) and 10 (m.t.b.) (bottom).
which has been detected in several other heterothallic basidiomycetes with multiple alleles (Coateset al., 1981).
Heterokaryotization occurred between certain
Wendy Thompson and P. Broda
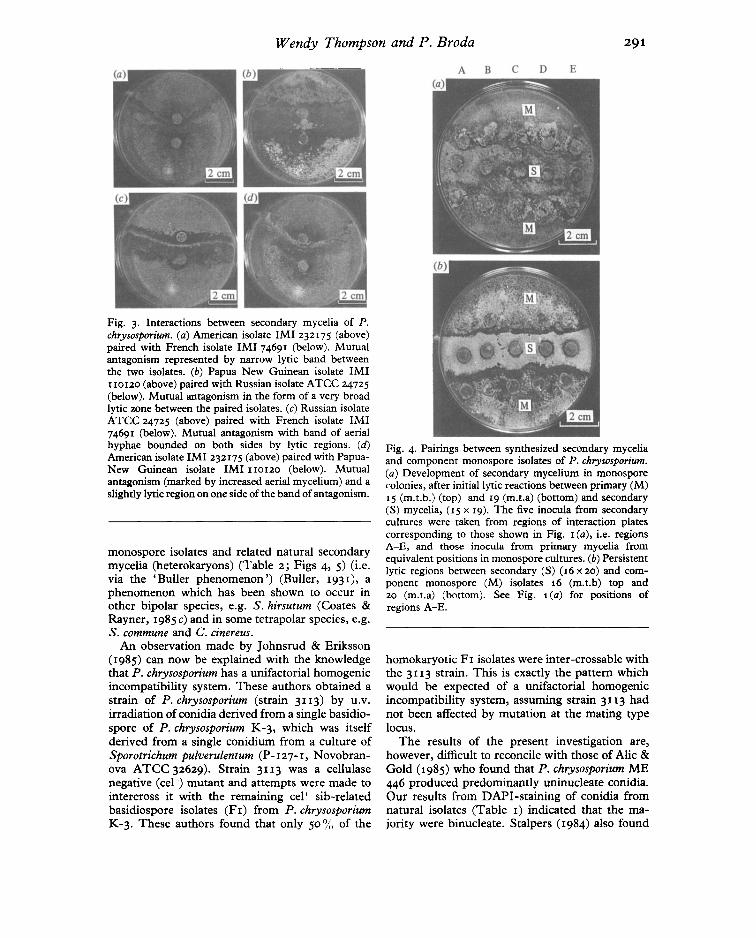
Fig. 4. Pairings between synthesized secondary myceliaand component monospore isolates of P. chrysosporium,(a) Development of secondary mycelium in monosporecolonies, after initial lytic reactions between primary (M)15 (m.t.b.) (top) and 19 (rn.t.a) (bottom) and secondary(S) mycelia, (15 x 19). The five inocula from secondarycultures were taken from regions of interaction platescorresponding to those shown in Fig. I (a), i.e. regionsA-E, and those inocula from primary mycelia fromequivalent positions in monospore cultures. (b) Persistentlytic regions between secondary (S) (16 x 20) and component monospore (M) isolates 16 (m.t.b) top and20 (m.t.a) (bottom). See Fig. 1(a) for positions ofregions A-E.
homokaryotic FI isolates were inter-crossable withthe 3 II 3 strain. This is exactly the pattern whichwould be expected of a unifactorial homogenicincompatibility system, assuming strain 31I3 hadnot been affected by mutation at the mating typelocus.
The results of the present investigation are,however, difficult to reconcile with those of Alic &Gold (1985) who found that P. chrysosporium ME446 produced predominantly uninucleate conidia.Our results from DAPI-staining of conidia fromnatural isolates (Table I) indicated that the majority were binucleate. Stalpers (1984) also found
EDcBA
monospore isolates and related natural secondarymycelia (heterokaryons) (Table 2; Figs 4, 5) (i.e.via the 'Buller phenomenon ') (Buller, 1931), aphenomenon which has been shown to occur inother bipolar species, e.g. S. hirsutum (Coates &Rayner, 1985 c) and in some tetrapolar species, e.g.S. commune and C. cinereus.
An observation made by Johnsrud & Eriksson(1985) can now be explained with the knowledgethat P. chrysosporium has a unifactorial homogenicincompatibility system. These authors obtained astrain of P. chrysosporium (strain 3II3) by u.v.irradiation of conidia derived from a single basidiospore of P. chrysosporium K-3, which was itselfderived from a single conidium from a culture ofSporotrichum pulverulentum (P-127-1, Novobranova ATCC 32629). Strain 3II3 was a cellulasenegative (cel) mutant and attempts were made tointercross it with the remaining eel" sib-relatedbasidiospore isolates (FI) from P. chrysosporiumK-3. These authors found that only 50 % of the
Fig. 3. Interactions between secondary mycelia of P.chrysosporium, (a) American isolate IMI 232175 (above)paired with French isolate IMI74691 (below). Mutualantagonism represented by narrow lytic band betweenthe two isolates. (b) Papua New Guinean isolate 1M1110120(above) paired with Russian isolate ATCC 24725(below). Mutual antagonism in the form of a very broadlytic zone between the paired isolates. (c) Russian isolateATCC 24725 (above) paired with French isolate 1M174691 (below). Mutual antagonism with band of aerialhyphae bounded on both sides by lytic regions. (d)American isolate 1M1 232175(above) paired with PapuaNew Guinean isolate 1M1 110120 (below). Mutualantagonism (marked by increased aerial mycelium) and aslightly lytic region on one side of the band of antagonism.
Mating behaviour in Phanerochaete chrysosporium
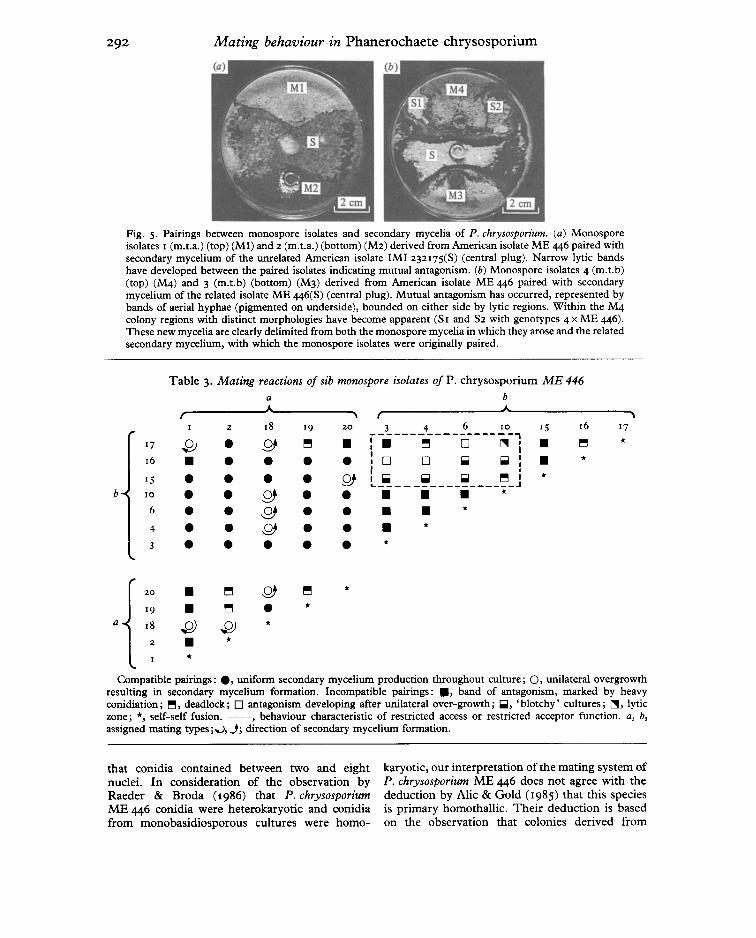
Fig. 5. Pairings between monospore isolates and secondary mycelia of P. chrysosporium . (a) Monosporeisolates 1 (m.t.a.) (top) (M I) and 2 (m.t.a.) (bottom) (M2) derived from American isolate M E 446 paired withsecondary mycelium of the unrelated American isolate IM I 232175(S) (central plug). Narrow lytic band shave developed between the paired isolates indicating mutual antagonism. (b) Monospore isolates 4 (rn.t .b)(top) (M4) and 3 (rn.t.b) (bottom) (M3) derived from Amer ican isolate ME 446 paired with secondarymycelium of the related isolate ME 446(S) (cent ral plug). Mutual antagonism has occurred, represented bybands of aerial hyphae (pigmented on underside), bounded on either side by lytic regions. Within the M4colony regions with distinct morphologies have become apparent (S I and S2 with genotypes 4 x ME 446).These new mycelia are clearly delimited from both the monospore mycelia in which they arose and the relatedsecondary mycelium, with which the monospore isolates were originally paired.
Table 3. Mating reactions of sib monospore isolates of P. chrysosporium ME 446
a b, A\ , A ,
1 2 18 19 20 3 4 6 10 15 16 17------------------,17 ~ • y. ~ • :. ~ 0 ["IIIJ I • ~ *16 • • • • • : 0 0 Iiiiil Iiiiil: • *15 • • • • Q) ! Iiiiil Iiiiil Iiiiil ~ I *~ _________________ J
b 10 • • g) • • • • • *6 • • y • • • • *4 • • g • • • *3 • • • • • *
20 • ~ ,Q) ~ *19 • ~ • *
a 18 .[2) ~ *2 • *
*Compatible pairings : . , uniform secondary mycelium production throughout culture ; 0 , unilateral overgrowth
resulting in secondary mycelium formation. Incompatible pairings: . , band of antagonism, marked by heavyconidiat ion; ~, deadlock ; 0 antagonism developing after unilateral over-growth; Iiiiil, ' blotchy' cultures; ["IIIJ, lyticzone ; *, self-self fusion. --- - , behaviour characteristic of restricted access or restricted accepto r function. a, b,assigned mating types;~, j ; direct ion of secondary mycelium form ation .
that conidia contained between two and eightnuclei. In consideration of the observation byRaeder & Broda (1986) that P. chrysosporiumME 446 conidia were heterokaryotic and conidiafrom monobasidiosporous cultures were homo-
karyotic, our interpretation of the mating system ofP . chrysosporium ME 446 does not agree with thededu ction by Alic & Gold (1985) that thi s speciesis primary homothallic. Their deduction is basedon the observation that colonies derived from

Wendy Thompson and P. Broda 293uninucleate conidia could fruit in culture, givingrise to large numbers of basidiospores. However,these features are not conclusive evidence ofhomothallism and homokaryotic fruiting is knownto occur in many basidiomycetes (Elliott , 1985).Homokaryotic fruiting with the production ofviable basidiospores has been observed in P. laevis(Ainsworth, pers. comm.).
Although the present investigation has concentrated on the mating behaviour of one isolate ofP. chrysosporium our results demonstrate that isolate ME 446 is not homothallic but that it has aunifactorial homogenic incompatibility system.This knowledge should facilitate the interpretationof previous genetical studies on this species andwill provide a better basis for the design of futureexperiments which may be concerned with obtaining and manipulating mutants with alteredligninolytic and cellulolytic activities.
This work was part of a programme supportedjointly by British Petroleum's Venture ResearchUnit and the Agricultural and Food ResearchCouncil. We thank Allan Brown and Dr UteRaeder for cultures derived from single basidiospores of P. chrysosporium ME 446, and DrsMartyn Ainsworth and Alan Rayner for valuablediscussion.
REFERENCES
ALIC, M. & GOLD, M. H. (1985). Genetic recombinationin the lignin-degrading basidiomycete Phanerochaetechrysosporium. Applied and Environmental Microbiology50,27-30.
BIGGS, R. (1938). Cultural studies in the Thelephoraceaeand related fungi. Mycologia 30, 64-78.
BODDY, L. & RAYNER, A. D . M. (1982). Populationstructures, intermycelial interactions and infectionbiology of Stereum gausapatum. Transactions of theBritish Mycological Society 78, 337-35 I.
BOlDIN, J. (1971). Nuclear behaviour in the myceliumand the evolution of the Basidiomycetes. In Evolutionin the Higher Ba sidiomycetes (ed. R. H. Peterson),p. 134. Knoxville : University of Tennessee Press.
BULLER, A. H. R. (1931). Researches on Fungi 4 p. 329.London: Longmans, Green and Co.
BURDSALL, H. H., JR (1981). The taxonomy of Sporotrichum pruinosum and Sporotrichum puloerulentum]Phanerochaete chrysosporium. Mycologia 73, 675-680.
BURDSALL, H . H ., JR & ESLYN, W. E. (1974). A newPhanerochaete with a Chrysosporium imperfect state .Mycotaxon 1, 123-133.
COATES, D. & RAYNER, A. D . M. (1985a). Genetic controland variation in expression of the 'bow-tie reactionbetween homokaryons of Stereum hirsutum. Transactions of the British Mycological Society 84, 191-205.
COATES, D. & RAYNER, A. D. M. (1985 b). Evidence for acytoplasmically transmissible factor affecting recognition and sornato-sexual differentiation in the basidiomycete Stereum hirsutum. Journal of General Microbiology 131,207-219.
COATES, D. & RAYNER, A. D . M. (1985c). Heterokaryon- homokaryon interactions in Stereum hirsutum. Transactions of the British Mycological Society 84, 637645·
COATES, D ., RAYNER, A. D. M. & TODD, N . K. (1981).Mating behaviour, mycelial antagonism and the establishment of individuals in Stereum hirsutum, Transactions of the British Mycological Society 76, 41-51.
EK, M. & ERIKSSON, K. E. (1980). Utilization of thewhite-rot fungus Sporotrichum puluerulentum for waterpurification and protein production on mixed lignocellulosic wastewaters. Biotechnology and Bioengineering 22, 2273-2284.
ELLIOTT, T . J. (1985). Developmental genetics - fromspore to sporophore. In Developmental Biology ofHigher Fungi ( roth Symposium of the British Mycological Society, April, 1984) (ed. D. Moore, L. A.Casselton, D . A. Wood & J. C. Frankland), pp . 451465. Cambridge, U.K . : Cambridge University Press .
ERIKSSON, J., HJORTSTAM, K . & RYVARDEN, L. (1978).The Corti ciaceae of North Europe 5, p. 989.
ERIKSSON, K. E., JOHNSRUD, S. C. & VALLANDER, L.(1983). Degradation of lignin and lignin model compounds by various mutants of the white-rot fungusSporotrichum puluerulentum. Archives of Microbiology135, 161-168 .
FENN, P. & KIRK, T. K. (1979). Ligninolytic system ofPhanerochaete chrysosporium : inhibition by a-phthalate. Archives of Microbiology 123, 307-309.
FINCHAM, J. R. S., DAY, P. R. & RADFORD, A. (1979).Fungal Genetics. Botanical Monographs 4 (ed. J. H.Burnett, H . G. Baker, H . Beevers & F. R. Whatley),p. 185. Oxford, U.K. : Blackwell Publications.
GOLD, M. H . & CHENG, T . M. (1979). Conditions forfruit body formation in the white rot basidiomycetePhanerochaete chrysosporium, Archives of Microbiology121,37-41.
GOLD, M. H ., CHENG, T. M ., KRISNANGKURA, K., MAYFIELD, M. B. & SMITH, L. M . (1980). Genetic andbiochemical studies on Phanerochaete chrysosporiumand their relation to lignin degradation. In LigninBiodegradation : Microbiology, Chemistry and PotentialApplications 2 (ed. T. K. Kirk, T. Higuchi & H . M.Chang), pp . 65-71. Boca Raton, Florida: CRC Press,Inc .
GoLD, M. H., CHENG, T. M. & MAYFIELD, M . B. (1962).Isolation and complementation studies of auxotrophicmutants of the lignin-degrading basidiomycetePhanerochaete chrysosporium. Applied and Environmental Microbiology 44, 996- 1000.
JEFFRIES, T . M ., CHOI, S. & KIRK, T . K. (1981).Nutritional regulation of lignin degradation byPhanerochaete chrysosporium, Applied and Environmental Microbiology 42, 290-296.
JOHNSRUD, S. C. & ERIKSSON, K. E. (1985). Crossbreeding of selected and mutated homokaryotic strainsof Phanerochaete chrysosporium K-3; new cellulosedeficient strains with increased ability to degrade lignin.Applied Microbiology and Biotechnology 21, 320-327.
KEMP, R. F . O. (1980). Bifactorial incompatibility without clamp connexions in the Coprinus patouillardiigroup. Transactions of the British Mycological Society74, 357-362.

294 Mating behaviour in Phanerochaete chrysosporiumKEYSER, P., KIRK, T. K. & ZEIKUS, J. G. (1978). Lig- D. H. Jennings & A. D. M. Rayner), pp. 383-417.
ninolytic enzyme system of Phanerochaete chryso- Cambridge, U.K.: Cambridge University Press.sporium:synthesisedintheabsenceoflignininresponse RAYNER, AD. M., COATES, D., AINSWORTH, A. M.,to nitrogen starvation. Journal of Bacteriology 135, ADAMS, T. J. H., WILLIAMS, E. N. D. & TODD, N. K.790-797. (1984). The biological consequences of the indivi-
LIN, M. S., COMINGS, D. E. & ALFI,0. S. (1977). Optical dualistic mycelium. In The Ecology and Physiology ofstudies of the interaction of 4'-6-Diamidino-2-phenyl- the Fungal Mycelium (8th Symposium of the Britishindole with DNA and metaphase chromosomes. Chro- Mycological Society 1983) (ed. D. H. Jennings & A. D.mosoma (Berlin) 60, 15-25. M. Rayner), pp. 509-540. Cambridge, U.K.: Cam-
RAEDER, U. & BRODA, P. (1986). Meiotic segregation bridge University Press.analysis of restriction site polymorphisms allows rapid STALPERS, J. A. (1984). A revision of the Genus Sporo-genetic mapping. European Molecular Biology Organis- trichum. Studies in Mycology (CBS) 24, 1-248.ation Journal, (In the Press). TIEN, M. & KIRK, T. K. (1984). Lignin degrading
RAYNER, A. D. M. & WEBBER, J. F. (1984). Interspecific enzyme from Phanerochaete chrysosporium. Purifica-mycelial interactions - an overview. In The Ecology tion, characterization, and catalytic properties of aand Physiology of the Fungal Mycelium (8th Symposium unique H 202-requiring oxygenase. Proceedings of theof the British Mycological Society, April 1983) (ed. National Academy of Science, USA 81, 2280-2284.
(Received for publication 30 October 1986)





























