Embed Size (px)
Citation preview

Archs oral Bid. Vol. 31, No. 6, pp. 351-356, 1986 Printed in Great Britain
OW3-9969/86 53.00 + 0.00 Pergamon Journals Ltd
MECHANICAL ANALYSIS OF THE MALFORMED, YET FUNCTIONAL, MANDIBULAR JOINTS OF A WILD TIMBER WOLF, CANS LUPUS
C. BARRETTE
Departement de Biologie, Universite Laval, Quebec, Canada GlK 7P4
Summary-In spite of grossly malformed jaw joints, presumably from birth, the animal survived in the wild for at least 3 years. Instead of a deep glenoid fossa folded over a cylindrical condyle to form a hinge, both joints had freely open articulating surfaces and a mandibular condyle without neck. The neckless condyle produced a shorter moment arm of resistance in all biting positions. The moment arm of the masseter, the jaw-adductor tending to disarticulate the jaw, was longer as a result of an elongated angular process. The combined effect at the carnassials was a 36 per cent improvement in the efficiency of the lever for which the joint is the fulcrum and thus an equivalent reduction in the disarticulating force. The joints were held together by an extra stout ligament about 5 mm in diameter and 8 mm long, linking the lateral pole of the condyle to the zygomaticarch.
INTRODUCTION
The mandibular joint (MJ) of mammals is without doubt one of the most interesting structures of verte- brate comparative anatomy (Crompton and Parker, 1978; Starck and Frick, 1984). The joint is also the fulcrum of the complicated lever that the mammalian jaw is considered to be (e.g. Davis, 1955; Maynard- Smith and Savage, 1959; Greaves, 1978; Hylander, 1975; Smith, 1978). The masticatory apparatus of mammals is a precision instrument, particularly in carnivores (e.g. Scapino, 1965, 1981; Emerson and Radinsky, 1980). This is best exemplified by the so-called Every effect which refers to the intricate and extraordinary accuracy needed in the adduction of the jaw to allow the proper working of the carnassial teeth (Mellet, 1981).
In view of that, it seems unlikely that a wild carnivore could function normally and survive with a diseased, injured or malformed MJ. My purpose is to describe an animal which did nevertheless survive.
ANIMALS STUDIED
The animal is an adult female timber wolf (Canis lupus) captured in the wild in Quebec (46”OO’ N, 75” 15’ W). When captured, the animal (no. 75-60) was 3 years old, as determined from cementum annuli count; it seemed healthy and quite normal. Only after the skull was cleaned and dried did the malformation become apparent. The skull will be compared with four skulls of the same size of wolves from the same region.
DESCRIPTION OF THE MALFORMATION
The malformation to the MJ was the same on both sides and the skull as a whole is as symmetrical as the other 4 wolf skulls. Except for the joints, the skull of 75-60 seemed quite normal; most dimensions, both of the cranium and mandible, were well within the range for normal wolves (Table 1). In addition, for the three measurements suggested by Emerson and Radinsky
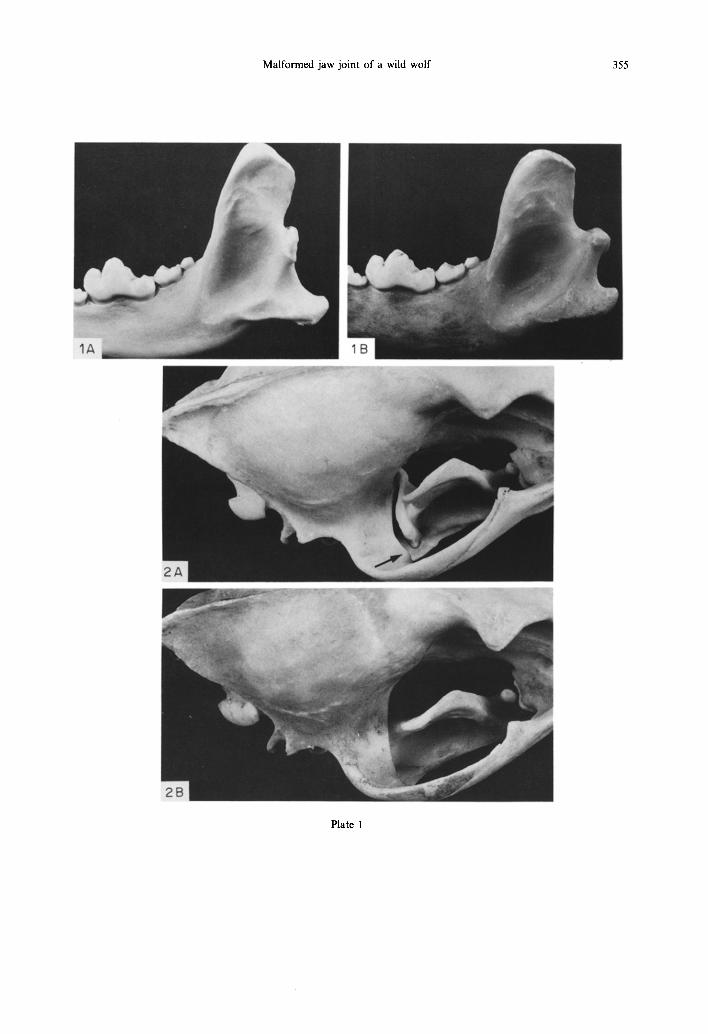
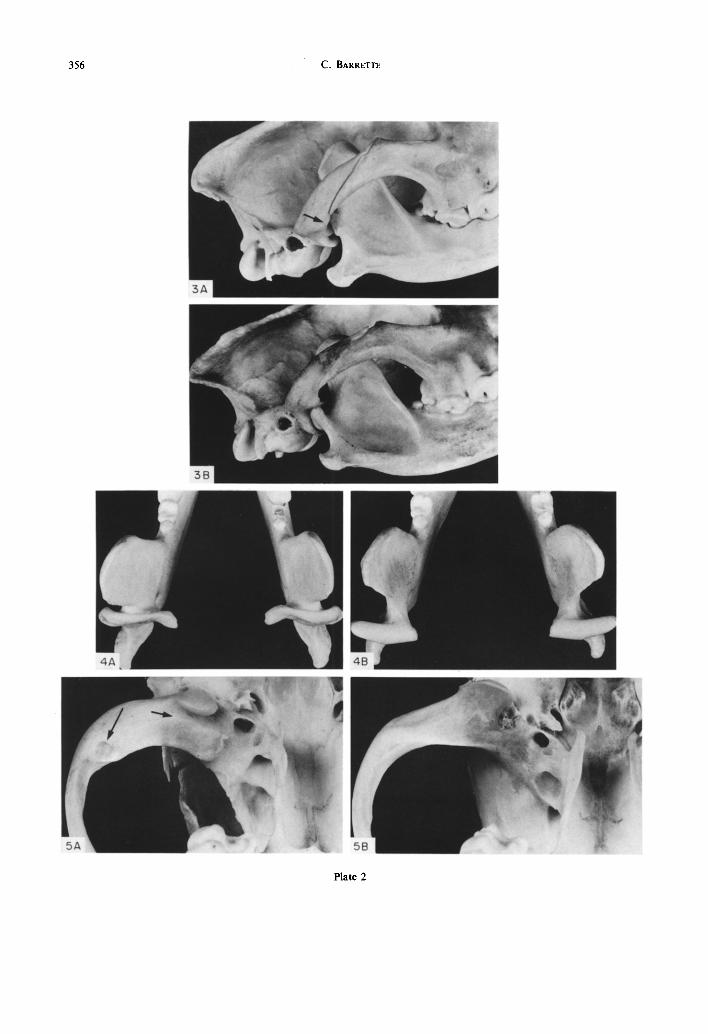
(1980) to evaluate the size of masticatory muscles from the morphology of their bony attachments, skull 75-60 was indistinguishable from the normal ones. The size of the carnassials (M, and Pm4) as well as that of Mi and the canines was normal. The main dimensions which in 75-60 differed markedly from the mean of the 4 normal wolves are those involving the MJ (Table 1). The mandibular condyle of 75-60 is without a neck (Plate Fig. lA), its articulating surface is thus close (about 5 mm) to the posterior edge of the coronoid process, compared with normal wolves in which it is about 15 mm behind the cor- onoid process. Seen from above, in 75-60 the condyle is in full view (Plate Fig. 2A) in front of the posterior end of the zygomatic arch and seen from the side its articulation was not at all the deep hinge of normal carnivores (Plate Fig. 3). Instead, the condyle has a complicated shape and its articulating surface is rather flat (Plate Fig. 4). The glenoid surface was different from the normal deep, semicircular fossa; it was shallow with the post-glenoid process separated from the articulating surface by a 2 mm wide and 3 mm deep gap (Plate Fig. 5A).
The MJ of mammals is enclosed by a capsule strengthened by a medial and a lateral ligament (Scapino, 1965; Souteyrand-Boulanger, 1971). The skull of 75-60 must have had in addition a stout ligament inserted on the lateral pole of the condyle and originating on the ventro-anterior edge of the zygomatic arch. It could have been an independent ligament or a local modification of the normal lateral ligament. The marks left on the bones by that liga- ment were about 5mm in diameter (Fig. 5A), and were clearly visible on both condyles and as a slight flange (Fig. 2A) on both zygomatic arches. Such marks were nowhere to be seen on the hundreds of normal canid skulls I have examined (e.g. Fig. 5B), and I could find no description of a rope-like liga- ment of that nature, neither could I see anything like it in 2 wolf heads I dissected for that purpose. The lateral ligament of canids is a broad sheet-like struc- ture (Scapino, 1965) inserted on the lateral pole of the condyle and adjacent part of the condylar process

352 C. BARRETTE
Table 1. Measurements of normal wolf skulls and of the abnormal one Normal
x+SDcn=4) Abnormal A oercentaee*
1. 2. 3. 4. 5. 6. I. 8. 9.
10. 11. 12. 13.
14.
Condylobasal length Palatal length Post-palatal length Upper cheek-tooth row length Zygomatic width Post-orbital least width Occipital width Palatal width outside of M, Mandible length: from angular process to infradental Lower cheek-tooth row length Height of vertical ramus Height of mandible behind M, Total mandible length: from condyle processinfradentale Mandibular condyle length
211.2 +0.9 109.3 * 1.5 88.3 k 1.1 79.5 $- 1.1
120.1 & 3.8 36.3 + 0.8 14.3 f 2.7 72.9 k 1.5
164.3 rf: 1.9 94.6 + 0.6 62.9 + 2.9 26.6 + 1.8
163.7 f 1.9 25.5 + 0.9
211.0 0.0 110.1 +0.7 88.2 0.0 78.5 -1.3
118.5 -1.3 34.6 -4.7 75.1 +1.1 70.4 -3.4
164.2 0.0 94.2 0.0 70.3 +11.8 29.1 +9.4
156.5 -4.4 29.7 + 16.5
All measurements are in mm, taken with a steel vernier caliper. Measurements nos 1, 2, 3, 5 and 8 as in Jolicoeur (1959); nos 6 and 7 as in Emerson and
Radinsky (1980); nos. 4, 9, 10, 11, 12 and 13 as in von den Driesch (1976). z (abnormal - ji- normal)
x 100. R normal
(Scapino, 1965). By comparison, the insertion mark of the postulated additional ligament of 75-60 was limited to the pole of the condyle and was circular.
With the jaw closed, the lateral pole of the condyle was about 8 mm from the zygomatic flange (Fig. 2A); that must have been the length of the ligament at rest. That distance was shorter by virtue of the longer than normal condyle (see Table 1 and Fig. 4).
In addition to an extra ligament, the jaw of 75-60, regarded as a bent lever, was different from that of the normal wolves. The major difference derives from the absence in 75-60 of a neck on the condyle, i.e. a considerable shortening of the moment arm of re- sistance at the biting site (Table 2 and Text Fig. 6). The moment arm of resistance at the carnassials was about 8 mm shorter than the mean of 4 normal wolves which had the same condylobasal length as 75-60 (Table 2). The moment arm of the masseter, i.e. the muscle tending to disarticulate the jaw (Maynard- Smith and Savage, 1959) was longer in 75-60 (Table 2). This results mainly from the greater development of the angular process (Fig. 1).
Fig. 6. Diagram of the lateral side of the left mandible of a wolf. 1, Moment arm of the masseter; 2, moment arm of the temporal; 3, resistance arm at the lower carnassial (M,).
Modified from Emerson and Radinsky (1980).
DISCUSSION
The skull of 75-60, being normal in all respects except the MJ, provides support for the idea of Moss (1968) that the mammalian jaw is made of many units, having the capacity to develop independently from each other. As the deformity is symmetrical it
Table 2. The moment arms of masticatory muscles and of the resistance at the carnassials (M, and Pm4) of wolves Normal jtkSD Abnormal A-N in mm A percentage*
1. Moment arm of masseter 29.6 f 2.1 36.4 +6.8 +23.0 2. Moment arm of temporal 36.6 f 1.2 34.1 -2.5 -6.8 3. Moment arm of resistance at M, 78.6 + 1.1 70.1 -8.5 - 10.8 4. Moment arm of resistance at Pm4 88.5 + 0.8 80.6 -7.9 -8.9 5. Efficiency of the masseter of M, 0.38 0.52 +0.14 +31 per cent 6. Efficiency of the masseter at Pm4 0.33 0.45 +0.12 +36 per cent 7. Efficiency of the temporal at M, 0.47 0.49 +0.02 f4 per cent 8. Efficiency of the temporal at Pm4 0.41 0.42 +0.01 f2 per cent
Measurements nos 1, 2 and 3 as in Fig. 6; no. 4 was taken from the vertical wall of the glenoid fossa to the apex of Pm4. Computation no. 5 = no. l/no. 3, no. 6 = no. l/no. 4, no. 7 = no. 2/no. 3, no. 8 = no. 2/no. 4.
*As in Table 1.

Malformed jaw joint of a wild wolf 353
seems likely that the abnormality is not the result of an injury but that it arose as an error of development. It is unknown whether the deformity was initially limited to the condyle or the glenoid fossa or both. In either case, there would have been “. . . secondary, compensatory and mechanically obligatory changes in the related skeletal unit” Moss (1968) to produce a functional joint.
The mammalian MJ is a stress-bearing joint during mastication (Brehman et al., 1981; Hylander, 1975, 1979; Smith, 1978; Kiltie, 1981; Weijs, 1980). The combined forces of the powerful jaw-adductor muscles and the resistance of the food at the site of biting, produce a large vertical component at the joint (on the balancing side). The vertical force is usually reslsted by the stout and wide glenoid fossa that acts as a buttress. On a normal wolf skull, it is obvious that opening the jaw, even widely, leaves the condyle stably tucked into the glenoid fossa. To remove the condyle from the fossa at a gape of about 20”, it is necessary first to slide it about 2-3 mm downward, and about 10 mm forward, a trajectory impossible in a living wolf as the large temporal muscle (about 60 per cent of the jaw adductors weight in carnivores; Davis, 1964) has a large backward-directed com- ponent. However, in skull 75-60, as soon as the jaw starts to open, the condyle slips upward, until the angular process of the mandible comes to rest against the zygomatic arch. Even without any muscular action, the mere rotation of the jaw causes the condyle to travel upward and the jaw to disarticulate. In 75-60, it is not only the action of the masseter that would have tended to pull the condyle out of the glenoid fossa, but the action of the temporal as well, because of the lack of the bony buttress against which the temporal muscle presses the condyle in the normal wolf.
Two adaptations which distinguish 75-60 con- tributed to its survival in spite of its abnormal MJ: a strong extra ligament and an efficient mechanical arrangement. The extra ligament must have been essential to limit disarticulation during biting. But its position is not the best that could presumably have been achieved. The lateral pole of the mandibular condyle is higher than normal (12 versus 7 mm) and the attachment of the ligament is at the lowest part of the pole of the condyle, which is the opposite of what would be predicted. Indeed, seen from the side, with the jaw closed, the condylar end of the ligament is .ibout 34mm below the zygomatic flange (Fig. 3A), not the best position to resist the upward excursion of the condyle during jaw opening and biting. Thus at every bite, the jaw must have been slightly disarticulated, i.e. up to the point where the insertion of the ligament moved 5-6 mm upward above its zygomatic origin and began to resist the vertical component of jaw muscles action at the joint.
Also, the condyle may have slipped a few milli- metres downward under the action of the digastric muscle in opening the jaw. In the normal wolf, the post-glenoid process hinders this downard movement (Ewer, 1973) whereas in 75-60 the post-glenoid pro- cess is too short to do so (Fig. 3). The concavity on the posterior surface of the condyle (Fig. 4) has exactly the same size and curvature as the post- glenoid process which suggests that the condyle did
slip downward and, possibly rotated, against the post-glenoid process whenever the jaw opened.
In terms of lever efficiency, the shorter moment arm of resistance and the longer moment arm of the masseter combined to yield a 36 per cent im- provement in the jaw lever of 75-60 at the camassials (Table 2; the same analysis applies to the canines). As the masseter has a component likely to dis- articulate the jaw (Maynard-Smith and Savage, 1959), 36 per cent improvement in its mechanical efficiency means that, to achieve a given bite force between the carnassials, 36 per cent less power was needed than in a normal wolf. That means a 36 per cent reduction in the upward reactive force at the condyle and thus a 36 per cent reduction in the force of disarticulation. No such improvement applies to the temporal muscle (Table 2), which may seem surprising as the temporal is usually regarded as the muscle which resists the disarticulation effect of the masseter (Maynard-Smith and Savage, 1959). How- ever, that can be only true where the temporal has a bony structure against which to press the condyle. In 75-60, a stronger or more efficient temporal would only tend to disarticulate the jaw further because no such bony structure is present.
The moment arm of the resistance at the car- nassials of 75-60 is shorter not only because the fulcrum of the lever is further forward, but also because the carnassials are further back. Whether measured from the anterior edge of the canine or the posterior edge of the coronoid process above the condyle (two landmarks presumably unaffected by the abnormal condylar process), the lower carnassial of 75-60 is about 2-3 mm more posterior than that of the 4 normal wolves (the same is true for the upper carnassial). The difference is small, but it would contribute to the reduction of the tendency to dis- articulate the jaw.
To work efficiently, the carnassials must be prop- erly aligned and strongly pressed against one another like the blades of a pair of scissors (Maynard-Smith and Savage, 1959). The deep and tight hinge-like joint of many carnivores is probably the main factor responsible for keeping the blades together. As wolf 75-60 did not have such a joint, muscular action (pterygoid muscles), interlocking canines (see Kiltie, 1981) and the Every effect (Mellet, 1981), provided resistance to lateral displacements of its jaw during biting. On the other hand, there was little beside the backward component of the adductor muscles, and the joint ligaments, to resist the longitudinal forward displacement and horizontal rotation of the man- dible.
Expressed simply, the MJ of 75-60 violates at least two mechanical properties considered to be essential to the proper working of a carnivore’s masticatory apparatus: it lacks the bony buttress thought to be essential to resist disarticulation, and its jaw joint is so open that it must have allowed horizontal (mostly longitudinal) displacements of the mandible of an amplitude sufficient to hinder the working of the carnassials. Nevertheless, the animal’s masticatory apparatus must have functioned with sufficient efficiency to allow its survival to 3 years of age.
This abnormal specimen may inspire a partial re-examination of some assumptions at the basis of

354 C. BARREN
the mechanical analysis of carnivores masticatory apparatus.
Acknowledgements-I am very grateful to D. Banville for providing the malformed skull, and to H. Jolicoeur and C. Pichette (all three from the Ministere du Loisir de la Chasse et de la P&he, of Quebec) for the normal skulls and the two wolf heads dissected. M. Bourassa photographed the specimens, and C. Dallaire drew the mandible.
REFERENCES
Brehman K., Boyd R. L., Laskin J., Gibbs C. H. and Mahan P. (1981) Direct measurement of loads at the temporomandibular joint in Macaca arctoides. J. dent. Res. 60, 182&1824.
Crompton A. W. and Parker P. (1978) Evolution of the mammalian masticatory apparatus. Am. Sci. 66, 192-201.
Davis D. D. (1955) Masticatory apparatus in the spectacled bear Tremarctos ornatus. Fieldiana Zool. 37, 25-46.
Davis D. D. (1964) The giant panda-A morphological study of evolutionary mechanisms. Fieldiana Zool. Memoirs 3, l-339.
Driesch A. von den (1976) A guide to the measurement of animal bones from archaelogical sites. Peabody Museum Bull. 1, l-137.
Emerson S. B. and Radinsky L. (1980) Functional analysis of sabertooth cranial morphology. Paleobiology 6, 295312.
Ewer R. F. (1973) The Curnoivores. Wiedenfeld & Nicolson, London.
Greaves W. S. (1978) The jaw lever system in ungulates: a new model. J. Zool., Lond. 184, 271-285.
Hylander W. L. (1975) The human mandible: lever or link? Am. J. phys. Anthrop. 43, 227-242.
Hylander W. L. (1979) An experimental analysis of tempo- romandibular joint reaction force in macaques. Am. J. phys. Anthrop. 51, 433456.
Jolicoeur P. (1959) Multivariate geographical variation in the wolf, Cunis lupus L. Evolution 13, 283-299.
Kiltie R. A. (1981) The function of interlocking canines in rain forest peccaries (Tayassuidae). J. Mammal. 62, 459469.
Maynard-Smith J. and Savage R. J. G. (1959) The mechan- ics of mammalian jaws. Sch. Sci. Rev. 141, 289301.
Mellet J. S. (1981) Mammalian camassial function and the “Every effect”. J. Mammal. 62, 164166.
Moss M. L. (1968) Functional cranial analysis of mam- malian mandibular ramal morphology. Acta anat. 71, 4234t7.
Scapino R. P. (1965) The third joint of the canine jaw. J. Morph. 116, 23-50.
Scapino R. P. (1981) Morphological investigation into functions of the jaw symphysis in camivorans. J. Morph. 167, 339-375.
Starck D. and Frick H. (1984) Vergleichende Anatomie heute-Aufgaben, Ergebnisse, Ziele. Z. Siiugetierk. 49, 257-268.
Smith R. J. (1978) Mandibular biomechanics and tempo- romandibular joint function in primates. Am. J. phys. Anthrop. 49, 341-350.
Souteyrand-Boulenger J. D. (1971) Arthrologie. In: Trait6 de Zoologie (Edited by Grass& P. P.) Vol. 16(3), pp. 837-l 157. Masson, Paris.
Weijs W. A. (1980) Biomechanical models and the analysis of form: a study of the mammalian masticatory appara- tus. Am. Zool. 20, 707-719.
Plate 1 Fig. 1. Lateral view of left mandible. A: abnormal; B: normal (as in all the photographs). Note the neckless
condyle, the longer angular process, and the longer coronoid process in A. Fig. 2. Dorsal view of the skulls with the mandibles in place and closed, with the teeth in occlusion. Note the fully-visible condyle in A. The gap between the lateral pole of the condyle and the slight flange (arrow)
on the zygomatic arch is where the extra ligament was located.
Plate 2
Fig. 3. Lateral view of the right side of the skulls with mandibles in place. Note the open articulation and long angular process in A, as well as the gap where the extra ligament was (arrow).
Fig. 4. Dorsal view of the condyles. The posterior convex curvature in A is the main articular surface (see Fig. 2); the concave one fits against the post-glenoid process.
Fig. 5. Ventral view of the left glenoid fossa. Note the gap between the articular surface and the post-glenoid process (short arrow), as well as the circular scar (5 mm in diameter) on the zygoma, in A
(long arrow).
Malformed jaw joint of a wild wolf 355
Plate 1
356 C. BARRETTE
Plate 2





























