Embed Size (px)
Citation preview

at SciVerse ScienceDirect
Crop Protection 35 (2012) 85e90
Contents lists available
Crop Protection
journal homepage: www.elsevier .com/locate/cropro
Melia azedarach controls Meloidogyne incognita and triggers plantdefense mechanisms on cucumber
I. Cavoski a, Z.Al. Chami a, F. Bouzebboudja a, N. Sasanelli b, V. Simeone a, D. Mondelli c,T. Miano c, G. Sarais d, N.G. Ntalli d, P. Caboni d,*a International Centre for Advanced Mediterranean Agronomic Studies, Valenzano (Ba), Italyb Institute for Plant Protection, C.N.R., Bari, ItalycDepartment of Environmental and Agro-Forestry Biology and Chemistry, Faculty of Agriculture, University of Bari, Bari, ItalydDepartment of Pharmaceutical Chemistry and Technology, University of Cagliari, Cagliari, Italy
a r t i c l e i n f o
Article history:Received 25 March 2011Received in revised form5 January 2012Accepted 17 January 2012
Keywords:Melia azedarach fruitsMeloidogyne incognitaNematicidal activityEnzymatic activity
* Corresponding author.E-mail address: [email protected] (P. Caboni).
0261-2194/$ e see front matter � 2012 Elsevier Ltd.doi:10.1016/j.cropro.2012.01.011
a b s t r a c t
Melia azedarach fruit extracts have recently raised a substantial interest for their use in crop protectionagainst phytoparasitic nematodes. The effect of M. azedarach on the root-knot nematode Meloidogyneincognita on cucumber, as well as the effect on the plant deference mechanism, is reported herein.Crushed fruits of M. azedarach, tested in the soil at the rates of 30 and 60 g kg�1, exhibited nematicidalactivity similar to the one of fenamiphos (0.02 g a.i. kg�1) in terms of nematode population in roots andsoil as well as reproduction rate. M. azedarach water extracts, rich in aldehydes, alcohols and carboxylicacids, showed nematicidal activity against M. incognita. Moreover, all M. azedarach treatments decreasedthe activities of catalase (CAT) and peroxidase (POX) involved in host H2O2 detoxification. Soil applicationof M. azedarach fruits could be favourably considered in the control of M. incognita on cucumber ina sustainable agriculture, since they act directly as nematicidals. Furthermore, M. azedarach elicits plantdefence and helps the host to fight the nematodes infestation in an indirect way.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Environmental and health concerns impose the reductionof conventional nematicides and the development of safer butcomparatively effective biorational nematode control measures(Noling and Becker, 1994). Plants may represent a source of natu-ral pesticides (Sasanelli and D’Addabbo, 1992, 1993; Isman,2000; Chitwood, 2002; Perez et al., 2003; Matthiessen andKirkegaard, 2006).
Melia azedarach L., commonly known as chinaberry, is a plantspecies of the Meliaceae family exhibiting a wide range of bio-logical activities of practical agricultural and pharmaceutical use.Extracts from various parts of M. azedarach are reported to exhibitinsecticidal (Bohnenstengela et al., 1999; Li, 1999; Céspedes et al.,2000; Carpinella et al., 2002; Abou-Fakhr Hammad et al., 2005;Charleston et al., 2005; Isman, 2006; Akhtar et al., 2008;Rachokarn et al., 2008), antifungal (Carpinella et al., 2003, 2005),nematicidal (Hasabo and Noweer, 2005; Ntalli et al., 2010a),antihelmintic (Maciel et al., 2006), cytotoxic and antiproliferative
All rights reserved.
activities (Ntalli et al., 2010b). Recently, carboxylic acids andaldehydes from M. azedarach fruits, and in particular furfural,5-hydroxymethylfurfural and furfurol have proven strong nema-ticides (Ntalli et al., 2010c). On the other hand, amending the soilwith chinaberry products has been proven to enhance the soilfertility. Toselli et al. (2010) concluded that M. azedarach leavesand fruits in combination with inorganic fertilizers and compostsstimulated mineral N release, root N uptake and peach (Prunuspersica L.) growth. Noble et al. (1996) reported that the leaf litter ofM. azedarach can significantly increase the soil ash alkalinity(an estimation of organic anion content) which results in anoverall pH increase. Furthermore, M. azedarach derivatives havebeen reported to stimulate soil microbial biomass (Marino et al.,2009; Spyrou et al., 2009).
Catalases (CAT) and peroxidases (APX) are H2O2 detoxificationenzymes in plants involved in the enzymatic scavenging of reactiveoxygen species (ROS) (Kim et al., 2005), occurring after plants’exposure to biotic or abiotic factors (pollutants, pathogen attacks,chilling and heating). Lowactivities of such enzymes inside the hostimply the enhanced defence system of the plant.
In the current study, we report on: a) the direct suppressiveeffect ofM. azedarach crushed fruits (CF) and their aqueous extracts(AEF) on the root-knot nematode Meloidogyne incognita (Kofoid

I. Cavoski et al. / Crop Protection 35 (2012) 85e9086
et White) Chitwood infecting cucumber, b) the GC/MS analysis ofthe chemical composition of AEF and c) the triggering of plantdefense system induced by the reduction of catalase (CAT) andascorbate peroxidases (APX) activities in host roots.
2. Materials and methods
2.1. Chemicals
Ultrapure water was obtained from the Millipore (Billerica, MA)Milli-Q system. The neem extract derived from Azadirachta indicaA. Juss contained 11% azadirachtin A (w w�1). Fenamiphos CS 240was obtained from Makhteshim Agan Group.
2.2. Extracts preparation
Ripen fruits of M. azedarach were collected in Cagliari, Italy inJanuary 2010. A voucher specimen was deposited in the Depart-ment of Life and Environmental Sciences (Botany and BotanicalGarden Division, Herbarium CAG, Sardinian Section, University ofCagliari, Italy) for species identification. Mature fruits were initiallygrinded in a blender and were successively used to prepareaqueous extracts (AEF) at two different ratios, 1:5 and 1:10 (w v�1).Samples were soaked by sonication for 15 min at 40 �C and filteredthrough aWhatman No.1 filter paper (Whatman International Ltd.,Maidstone, England) before their use. In order to calculate theextraction yields, extracts were vacuum dried by rotary evaporator(Ika Werke, Germany) at 60 �C to constant weight and yields werecalculated as dry weights. Three replicates were carried out for eachextraction type.
2.3. GCeMS analysis
ATrace GC Ultra gas chromatograph (Thermo Finnigan), coupledwith a Trace DSQ mass spectrometry detector, a splitesplitlessinjector, and an Xcalibur MS platform, was used to analyse theM. azedarach aqueous extract (AEF). The column was a fused silicacapillary Varian CP-WAX 57CB (60 m � 0.25 mm; filmthickness ¼ 0.25 mm) (Varian Inc.). The injector and the transferline temperatures were set at 200 �C. The oven temperature wasprogrammed as follows: 50 �C (hold for 1 min), raised to 220 �C(3 �C min�1), and isothermally held for 30 min. Heliumwas used ascarrier gas at 1 mL min�1; 1 mL of AEF extract at a concentration of2500 mg mL�1 was injected in the splitless mode. MS conditionswere as follows: ionization mode EI positive from 40 to 300 amu.The components of AEF were identified by (a) comparison of theirrelative retention times and mass fragmentation with those ofauthentic standards and (b) computer matching against the NIST98library. Quantitative analysis of each component was carried outwith the external standard method.
2.4. Pot experiments
The Italian population of M. incognita host race 1 (Taylor andSasser, 1978), was reared on tomato plants (Solanum lycopersicumL.) cv. Rutgers in a glasshouse at 25 � 2 �C. Two months later thenumber of eggs and juveniles (J2) were quantified in roots (Husseyand Barker, 1973). The chopped infected roots were then thor-oughly mixed with 3 kg of steam sterilised sandy soil (pH 7.2;sand > 99%; silt < 1%; clay < 1% and organic matter ¼ 0.75%) andwere used as inoculum. Appropriate amounts of this inoculumwere added and mixed with steam sterilised soil to give an initialnematode population density of 5 eggs and juveniles mL�1 of soil(Pi). Seven treatments were established in the pot experiment:
a and b) crushed M. azedarach fruits incorporated into theM. incognita infested soil at rates of 30 and 60 g kg�1 soil (CF1 andCF2); c and d) aqueous extracts (1:5 and 1:10; w v�1) prepared fromthe same amount of fruits (AEF1 and AEF2); e) a commercialformulation of azadirachtin (AZA) derived from neem plants(A. indica A. Juss) applied at the dose of 0.03 g kg�1 soil (Ntalli et al.,2009); f) a fenamiphos treatment (FEN) at the rate of 0.02 g kg�1
soil (62.5 L of a commercial formulation at 24% a.i. ha�1) and g) aninfested untreated soil control (CON). All treatments were appliedat transplant. To evaluate the effect of the nematode suppression onroot enzymatic activity M. azedarach treatments were replicated ina nematode-infested and non-infested soil. Treatments of potswere performed at 26.8% of soil water holding capacity. Eachtreatment was replicated five times in a randomised block design at25 � 2 �C. In each pot a one-month-old cucumber (Cucumis sativusL.) seedling, hybrids Sakamari F1, was transplanted. During theexperiment plants were irrigated as needed but no fertilizers wereapplied.
Two months later, plants were uprooted and differentassessments were made. Root gall index (RGI) on each rootsystem was estimated according to a 0e5 scale, where 0 ¼ nogalls; 1 ¼ 1e2 galls; 2 ¼ 3e10 galls; 3 ¼ 11e30 galls; 4 ¼ 31e100galls and 5 > 100 galls with a root system completely deformedby the presence of numerous and large galls (Hussey and Barker,1973). Final nematode soil density in each pot was determined byprocessing 500 mL soil by the Coolen’s method (Coolen, 1979).M. incognita density in roots was assessed according to Husseyand Barker (1973). Final nematode population density (Pf) ineach pot was determined by summing nematodes recovered fromsoil and roots. The reproduction rate of the nematode (r) was alsocalculated as ratio between final and initial population density(Pf Pi�1).
2.5. Enzymes extraction and assays
Fresh cucumber root samples (5 g) were homogenized in anOsterizer Sunbeam, Designer homogenizer in 50 mL 100 mMpotassium phosphate buffer (pH 7.6) containing 1 mM EDTA-Na2and 0.5 mM ascorbate. The homogenized samples were centrifugedat 10,000 rpm for 5 min. The supernatant was used as a crudeenzyme extract in catalase (CAT) and ascorbate peroxidase (APX)enzyme analyses. All measurements were made at 20 �C in a UNI-CAM BS DISC PD 2000-1 spectrophotometer. APX activity (EC1.11.1.11) was determined by following the decrease of ascorbateand measuring the change in absorbance at 290 nm for 1 min in2 mL of a reaction mixture containing 50 mM potassium phosphatebuffer (pH 7.0),1mMEDTA-Na2, 0.5 mMascorbic acid, 0.1mMH2O2and 50 mL of crude enzyme extract at 25 �C (Nakano and Asada,1981). The activity was calculated from the extinction coefficient(2.8 mM�1 cm�1) for the ascorbate. CAT activity (EC 1.11.1.6) wasdetermined as a decrease in absorbance at 240 nm for 1 minfollowing the decomposition of H2O2 (Cakmak et al., 1993).The reaction mixture (3 mL) contained 50 mM phosphate buffer(pH 7.0), 15 mM H2O2 and 50 mL of crude enzyme extract at 25 �C.The activity was calculated from the extinction coefficient(40 mM�1 cm�1) for H2O2.
2.6. Statistical analysis
Enzymatic and nematological data were subjected to statisticalanalysis of variance (ANOVA) and means were compared byStudent’s t test and least significant difference (LSD) test at 0.05 and0.01 levels of significance. Statistical analysis was performed usingthe PlotIT program.
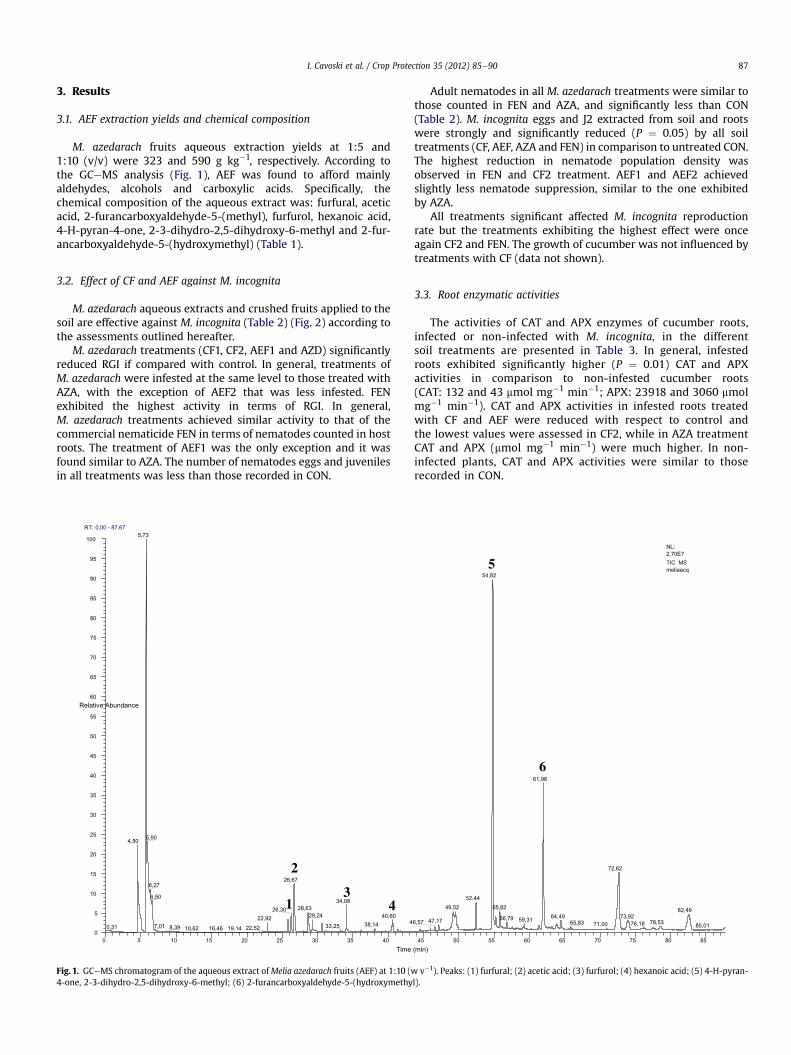
I. Cavoski et al. / Crop Protection 35 (2012) 85e90 87
3. Results
3.1. AEF extraction yields and chemical composition
M. azedarach fruits aqueous extraction yields at 1:5 and1:10 (v/v) were 323 and 590 g kg�1, respectively. According tothe GCeMS analysis (Fig. 1), AEF was found to afford mainlyaldehydes, alcohols and carboxylic acids. Specifically, thechemical composition of the aqueous extract was: furfural, aceticacid, 2-furancarboxyaldehyde-5-(methyl), furfurol, hexanoic acid,4-H-pyran-4-one, 2-3-dihydro-2,5-dihydroxy-6-methyl and 2-fur-ancarboxyaldehyde-5-(hydroxymethyl) (Table 1).
3.2. Effect of CF and AEF against M. incognita
M. azedarach aqueous extracts and crushed fruits applied to thesoil are effective against M. incognita (Table 2) (Fig. 2) according tothe assessments outlined hereafter.
M. azedarach treatments (CF1, CF2, AEF1 and AZD) significantlyreduced RGI if compared with control. In general, treatments ofM. azedarach were infested at the same level to those treated withAZA, with the exception of AEF2 that was less infested. FENexhibited the highest activity in terms of RGI. In general,M. azedarach treatments achieved similar activity to that of thecommercial nematicide FEN in terms of nematodes counted in hostroots. The treatment of AEF1 was the only exception and it wasfound similar to AZA. The number of nematodes eggs and juvenilesin all treatments was less than those recorded in CON.
RT: 0,00 - 87,67
0 5 10 15 20 25 30 35 40Time
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100
Relative Abundance
5,73
5,904,50
26,676,27
6,5034,08
28,6326,3029,24 40,6022,92
438,147,01 33,250,31 8,39 22,5210,62 19,1416,46
1
2
43
Fig. 1. GCeMS chromatogram of the aqueous extract ofMelia azedarach fruits (AEF) at 1:10 (w4-one, 2-3-dihydro-2,5-dihydroxy-6-methyl; (6) 2-furancarboxyaldehyde-5-(hydroxymethy
Adult nematodes in all M. azedarach treatments were similar tothose counted in FEN and AZA, and significantly less than CON(Table 2). M. incognita eggs and J2 extracted from soil and rootswere strongly and significantly reduced (P ¼ 0.05) by all soiltreatments (CF, AEF, AZA and FEN) in comparison to untreated CON.The highest reduction in nematode population density wasobserved in FEN and CF2 treatment. AEF1 and AEF2 achievedslightly less nematode suppression, similar to the one exhibitedby AZA.
All treatments significant affected M. incognita reproductionrate but the treatments exhibiting the highest effect were onceagain CF2 and FEN. The growth of cucumber was not influenced bytreatments with CF (data not shown).
3.3. Root enzymatic activities
The activities of CAT and APX enzymes of cucumber roots,infected or non-infected with M. incognita, in the differentsoil treatments are presented in Table 3. In general, infestedroots exhibited significantly higher (P ¼ 0.01) CAT and APXactivities in comparison to non-infested cucumber roots(CAT: 132 and 43 mmol mg�1 min�1; APX: 23918 and 3060 mmolmg�1 min�1). CAT and APX activities in infested roots treatedwith CF and AEF were reduced with respect to control andthe lowest values were assessed in CF2, while in AZA treatmentCAT and APX (mmol mg�1 min�1) were much higher. In non-infected plants, CAT and APX activities were similar to thoserecorded in CON.
45 50 55 60 65 70 75 80 85 (min)
54,82
61,98
72,62
52,4449,52 55,82 82,49
73,9264,4956,79 59,3147,176,57 78,5365,83 76,1871,00 85,01
NL:2,70E7TIC MS meliaacq5
6
v�1). Peaks: (1) furfural; (2) acetic acid; (3) furfurol; (4) hexanoic acid; (5) 4-H-pyran-l).
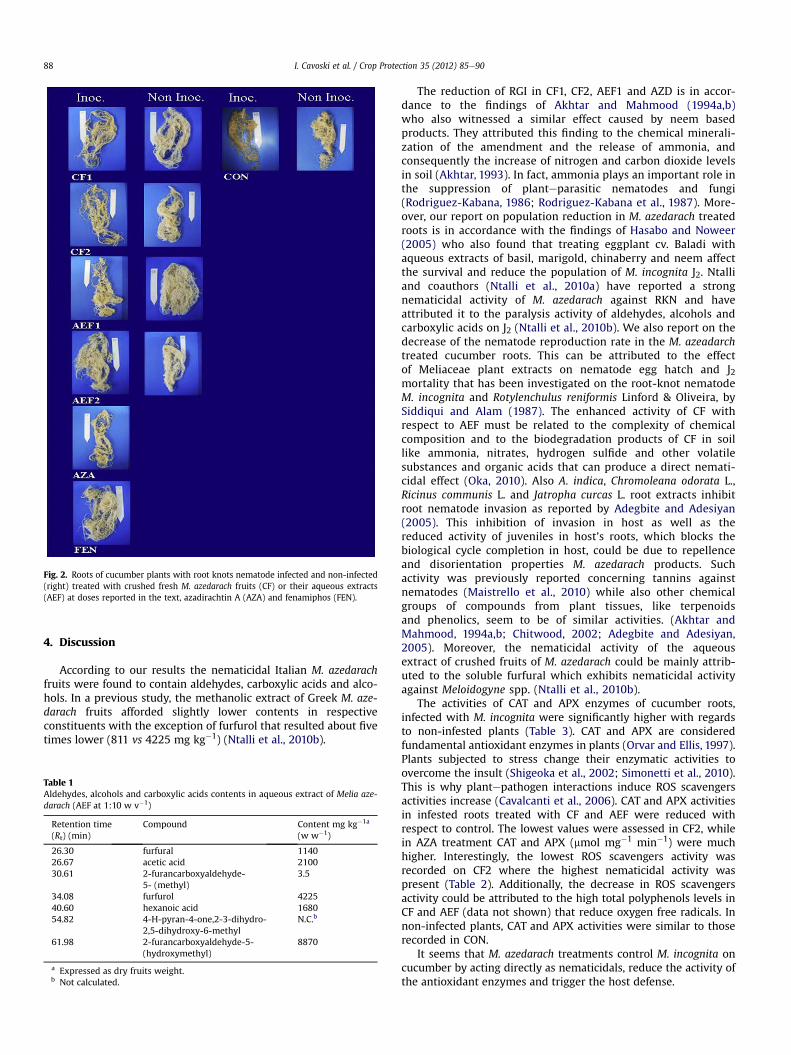
Fig. 2. Roots of cucumber plants with root knots nematode infected and non-infected(right) treated with crushed fresh M. azedarach fruits (CF) or their aqueous extracts(AEF) at doses reported in the text, azadirachtin A (AZA) and fenamiphos (FEN).
I. Cavoski et al. / Crop Protection 35 (2012) 85e9088
4. Discussion
According to our results the nematicidal Italian M. azedarachfruits were found to contain aldehydes, carboxylic acids and alco-hols. In a previous study, the methanolic extract of Greek M. aze-darach fruits afforded slightly lower contents in respectiveconstituents with the exception of furfurol that resulted about fivetimes lower (811 vs 4225 mg kg�1) (Ntalli et al., 2010b).
Table 1Aldehydes, alcohols and carboxylic acids contents in aqueous extract of Melia aze-darach (AEF at 1:10 w v�1)
Retention time(Rt) (min)
Compound Content mg kg�1a
(w w�1)
26.30 furfural 114026.67 acetic acid 210030.61 2-furancarboxyaldehyde-
5- (methyl)3.5
34.08 furfurol 422540.60 hexanoic acid 168054.82 4-H-pyran-4-one,2-3-dihydro-
2,5-dihydroxy-6-methylN.C.b
61.98 2-furancarboxyaldehyde-5-(hydroxymethyl)
8870
a Expressed as dry fruits weight.b Not calculated.
The reduction of RGI in CF1, CF2, AEF1 and AZD is in accor-dance to the findings of Akhtar and Mahmood (1994a,b)who also witnessed a similar effect caused by neem basedproducts. They attributed this finding to the chemical minerali-zation of the amendment and the release of ammonia, andconsequently the increase of nitrogen and carbon dioxide levelsin soil (Akhtar, 1993). In fact, ammonia plays an important role inthe suppression of planteparasitic nematodes and fungi(Rodriguez-Kabana, 1986; Rodriguez-Kabana et al., 1987). More-over, our report on population reduction in M. azedarach treatedroots is in accordance with the findings of Hasabo and Noweer(2005) who also found that treating eggplant cv. Baladi withaqueous extracts of basil, marigold, chinaberry and neem affectthe survival and reduce the population of M. incognita J2. Ntalliand coauthors (Ntalli et al., 2010a) have reported a strongnematicidal activity of M. azedarach against RKN and haveattributed it to the paralysis activity of aldehydes, alcohols andcarboxylic acids on J2 (Ntalli et al., 2010b). We also report on thedecrease of the nematode reproduction rate in the M. azeadarchtreated cucumber roots. This can be attributed to the effectof Meliaceae plant extracts on nematode egg hatch and J2mortality that has been investigated on the root-knot nematodeM. incognita and Rotylenchulus reniformis Linford & Oliveira, bySiddiqui and Alam (1987). The enhanced activity of CF withrespect to AEF must be related to the complexity of chemicalcomposition and to the biodegradation products of CF in soillike ammonia, nitrates, hydrogen sulfide and other volatilesubstances and organic acids that can produce a direct nemati-cidal effect (Oka, 2010). Also A. indica, Chromoleana odorata L.,Ricinus communis L. and Jatropha curcas L. root extracts inhibitroot nematode invasion as reported by Adegbite and Adesiyan(2005). This inhibition of invasion in host as well as thereduced activity of juveniles in host’s roots, which blocks thebiological cycle completion in host, could be due to repellenceand disorientation properties M. azedarach products. Suchactivity was previously reported concerning tannins againstnematodes (Maistrello et al., 2010) while also other chemicalgroups of compounds from plant tissues, like terpenoidsand phenolics, seem to be of similar activities. (Akhtar andMahmood, 1994a,b; Chitwood, 2002; Adegbite and Adesiyan,2005). Moreover, the nematicidal activity of the aqueousextract of crushed fruits of M. azedarach could be mainly attrib-uted to the soluble furfural which exhibits nematicidal activityagainst Meloidogyne spp. (Ntalli et al., 2010b).
The activities of CAT and APX enzymes of cucumber roots,infected with M. incognita were significantly higher with regardsto non-infested plants (Table 3). CAT and APX are consideredfundamental antioxidant enzymes in plants (Orvar and Ellis, 1997).Plants subjected to stress change their enzymatic activities toovercome the insult (Shigeoka et al., 2002; Simonetti et al., 2010).This is why plantepathogen interactions induce ROS scavengersactivities increase (Cavalcanti et al., 2006). CAT and APX activitiesin infested roots treated with CF and AEF were reduced withrespect to control. The lowest values were assessed in CF2, whilein AZA treatment CAT and APX (mmol mg�1 min�1) were muchhigher. Interestingly, the lowest ROS scavengers activity wasrecorded on CF2 where the highest nematicidal activity waspresent (Table 2). Additionally, the decrease in ROS scavengersactivity could be attributed to the high total polyphenols levels inCF and AEF (data not shown) that reduce oxygen free radicals. Innon-infected plants, CAT and APX activities were similar to thoserecorded in CON.
It seems that M. azedarach treatments control M. incognita oncucumber by acting directly as nematicidals, reduce the activity ofthe antioxidant enzymes and trigger the host defense.
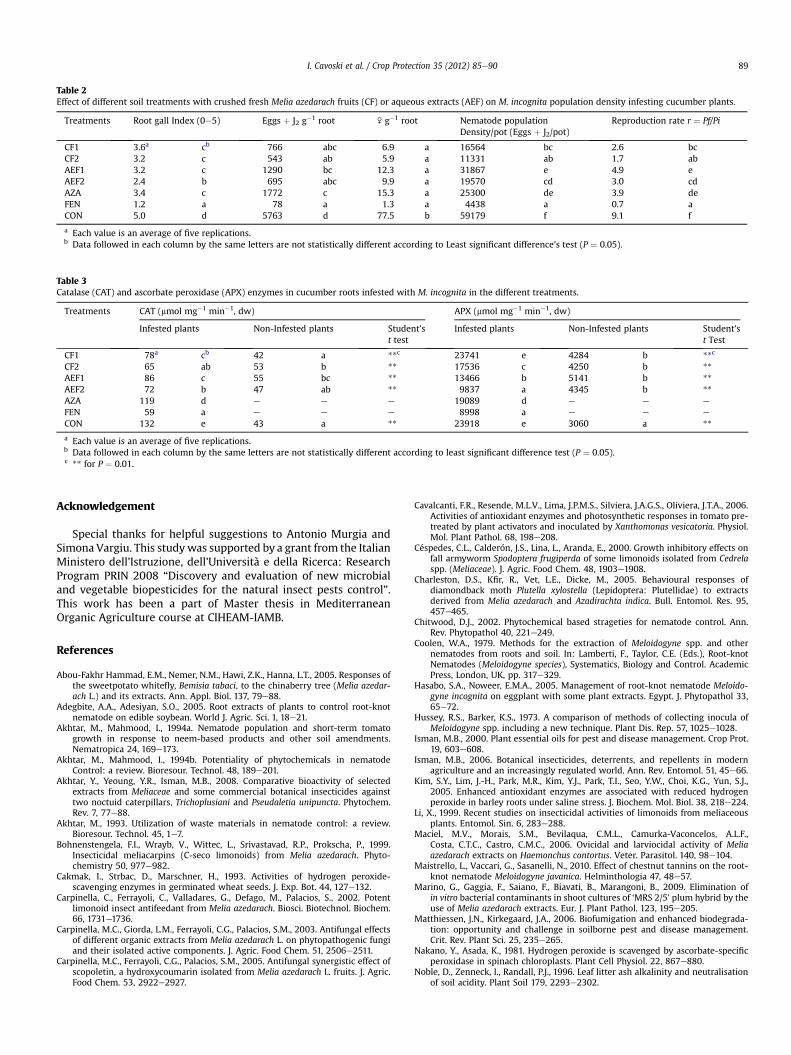
Table 3Catalase (CAT) and ascorbate peroxidase (APX) enzymes in cucumber roots infested with M. incognita in the different treatments.
Treatments CAT (mmol mg�1 min�1, dw) APX (mmol mg�1 min�1, dw)
Infested plants Non-Infested plants Student’st test
Infested plants Non-Infested plants Student’st Test
CF1 78a cb 42 a **c 23741 e 4284 b **c
CF2 65 ab 53 b ** 17536 c 4250 b **
AEF1 86 c 55 bc ** 13466 b 5141 b **
AEF2 72 b 47 ab ** 9837 a 4345 b **
AZA 119 d e e e 19089 d e e e
FEN 59 a e e e 8998 a e e e
CON 132 e 43 a ** 23918 e 3060 a **
a Each value is an average of five replications.b Data followed in each column by the same letters are not statistically different according to least significant difference test (P ¼ 0.05).c ** for P ¼ 0.01.
Table 2Effect of different soil treatments with crushed fresh Melia azedarach fruits (CF) or aqueous extracts (AEF) on M. incognita population density infesting cucumber plants.
Treatments Root gall Index (0e5) Eggs þ J2 g�1 root \ g�1 root Nematode populationDensity/pot (Eggs þ J2/pot)
Reproduction rate r ¼ Pf/Pi
CF1 3.6a cb 766 abc 6.9 a 16564 bc 2.6 bcCF2 3.2 c 543 ab 5.9 a 11331 ab 1.7 abAEF1 3.2 c 1290 bc 12.3 a 31867 e 4.9 eAEF2 2.4 b 695 abc 9.9 a 19570 cd 3.0 cdAZA 3.4 c 1772 c 15.3 a 25300 de 3.9 deFEN 1.2 a 78 a 1.3 a 4438 a 0.7 aCON 5.0 d 5763 d 77.5 b 59179 f 9.1 f
a Each value is an average of five replications.b Data followed in each column by the same letters are not statistically different according to Least significant difference’s test (P ¼ 0.05).
I. Cavoski et al. / Crop Protection 35 (2012) 85e90 89
Acknowledgement
Special thanks for helpful suggestions to Antonio Murgia andSimona Vargiu. This studywas supported by a grant from the ItalianMinistero dell’Istruzione, dell’Università e della Ricerca: ResearchProgram PRIN 2008 “Discovery and evaluation of new microbialand vegetable biopesticides for the natural insect pests control”.This work has been a part of Master thesis in MediterraneanOrganic Agriculture course at CIHEAM-IAMB.
References
Abou-Fakhr Hammad, E.M., Nemer, N.M., Hawi, Z.K., Hanna, L.T., 2005. Responses ofthe sweetpotato whitefly, Bemisia tabaci, to the chinaberry tree (Melia azedar-ach L.) and its extracts. Ann. Appl. Biol. 137, 79e88.
Adegbite, A.A., Adesiyan, S.O., 2005. Root extracts of plants to control root-knotnematode on edible soybean. World J. Agric. Sci. 1, 18e21.
Akhtar, M., Mahmood, I., 1994a. Nematode population and short-term tomatogrowth in response to neem-based products and other soil amendments.Nematropica 24, 169e173.
Akhtar, M., Mahmood, I., 1994b. Potentiality of phytochemicals in nematodeControl: a review. Bioresour. Technol. 48, 189e201.
Akhtar, Y., Yeoung, Y.R., Isman, M.B., 2008. Comparative bioactivity of selectedextracts from Meliaceae and some commercial botanical insecticides againsttwo noctuid caterpillars, Trichoplusiani and Pseudaletia unipuncta. Phytochem.Rev. 7, 77e88.
Akhtar, M., 1993. Utilization of waste materials in nematode control: a review.Bioresour. Technol. 45, 1e7.
Bohnenstengela, F.I., Wrayb, V., Wittec, L., Srivastavad, R.P., Prokscha, P., 1999.Insecticidal meliacarpins (C-seco limonoids) from Melia azedarach. Phyto-chemistry 50, 977e982.
Cakmak, I., Strbac, D., Marschner, H., 1993. Activities of hydrogen peroxide-scavenging enzymes in germinated wheat seeds. J. Exp. Bot. 44, 127e132.
Carpinella, C., Ferrayoli, C., Valladares, G., Defago, M., Palacios, S., 2002. Potentlimonoid insect antifeedant from Melia azedarach. Biosci. Biotechnol. Biochem.66, 1731e1736.
Carpinella, M.C., Giorda, L.M., Ferrayoli, C.G., Palacios, S.M., 2003. Antifungal effectsof different organic extracts from Melia azedarach L. on phytopathogenic fungiand their isolated active components. J. Agric. Food Chem. 51, 2506e2511.
Carpinella, M.C., Ferrayoli, C.G., Palacios, S.M., 2005. Antifungal synergistic effect ofscopoletin, a hydroxycoumarin isolated from Melia azedarach L. fruits. J. Agric.Food Chem. 53, 2922e2927.
Cavalcanti, F.R., Resende, M.L.V., Lima, J.P.M.S., Silviera, J.A.G.S., Oliviera, J.T.A., 2006.Activities of antioxidant enzymes and photosynthetic responses in tomato pre-treated by plant activators and inoculated by Xanthomonas vesicatoria. Physiol.Mol. Plant Pathol. 68, 198e208.
Céspedes, C.L., Calderón, J.S., Lina, L., Aranda, E., 2000. Growth inhibitory effects onfall armyworm Spodoptera frugiperda of some limonoids isolated from Cedrelaspp. (Meliaceae). J. Agric. Food Chem. 48, 1903e1908.
Charleston, D.S., Kfir, R., Vet, L.E., Dicke, M., 2005. Behavioural responses ofdiamondback moth Plutella xylostella (Lepidoptera: Plutellidae) to extractsderived from Melia azedarach and Azadirachta indica. Bull. Entomol. Res. 95,457e465.
Chitwood, D.J., 2002. Phytochemical based strageties for nematode control. Ann.Rev. Phytopathol 40, 221e249.
Coolen, W.A., 1979. Methods for the extraction of Meloidogyne spp. and othernematodes from roots and soil. In: Lamberti, F., Taylor, C.E. (Eds.), Root-knotNematodes (Meloidogyne species), Systematics, Biology and Control. AcademicPress, London, UK, pp. 317e329.
Hasabo, S.A., Noweer, E.M.A., 2005. Management of root-knot nematode Meloido-gyne incognita on eggplant with some plant extracts. Egypt. J. Phytopathol 33,65e72.
Hussey, R.S., Barker, K.S., 1973. A comparison of methods of collecting inocula ofMeloidogyne spp. including a new technique. Plant Dis. Rep. 57, 1025e1028.
Isman, M.B., 2000. Plant essential oils for pest and disease management. Crop Prot.19, 603e608.
Isman, M.B., 2006. Botanical insecticides, deterrents, and repellents in modernagriculture and an increasingly regulated world. Ann. Rev. Entomol. 51, 45e66.
Kim, S.Y., Lim, J.-H., Park, M.R., Kim, Y.J., Park, T.I., Seo, Y.W., Choi, K.G., Yun, S.J.,2005. Enhanced antioxidant enzymes are associated with reduced hydrogenperoxide in barley roots under saline stress. J. Biochem. Mol. Biol. 38, 218e224.
Li, X., 1999. Recent studies on insecticidal activities of limonoids from meliaceousplants. Entomol. Sin. 6, 283e288.
Maciel, M.V., Morais, S.M., Bevilaqua, C.M.L., Camurka-Vaconcelos, A.L.F.,Costa, C.T.C., Castro, C.M.C., 2006. Ovicidal and larviocidal activity of Meliaazedarach extracts on Haemonchus contortus. Veter. Parasitol. 140, 98e104.
Maistrello, L., Vaccari, G., Sasanelli, N., 2010. Effect of chestnut tannins on the root-knot nematode Meloidogyne javanica. Helminthologia 47, 48e57.
Marino, G., Gaggia, F., Saiano, F., Biavati, B., Marangoni, B., 2009. Elimination ofin vitro bacterial contaminants in shoot cultures of ‘MRS 2/5’ plum hybrid by theuse of Melia azedarach extracts. Eur. J. Plant Pathol. 123, 195e205.
Matthiessen, J.N., Kirkegaard, J.A., 2006. Biofumigation and enhanced biodegrada-tion: opportunity and challenge in soilborne pest and disease management.Crit. Rev. Plant Sci. 25, 235e265.
Nakano, Y., Asada, K., 1981. Hydrogen peroxide is scavenged by ascorbate-specificperoxidase in spinach chloroplasts. Plant Cell Physiol. 22, 867e880.
Noble, D., Zenneck, I., Randall, P.J., 1996. Leaf litter ash alkalinity and neutralisationof soil acidity. Plant Soil 179, 2293e2302.

I. Cavoski et al. / Crop Protection 35 (2012) 85e9090
Noling, J.W., Becker, J.O., 1994. The challenge of research and extension to defineand implement alternatives to methyl bromide. J. Nematol. 26, 573e586.
Ntalli, N.G., Menkissoglu-Spiroudi, U., Giannakou, I.O., Prophetou-Athanasiadou, D.A.,2009. Efficacyevaluationof a neem (Azadirachta indicaA. Juss) formulationagainstroot-knot nematodes Meloidogyne incognita. Crop Prot. 28, 489e494.
Ntalli, N.G., Menkisoglu-Spiroudi, U., Giannakou, I., 2010a. Nematicidal activity ofpowder and extracts of Melia azedarach fruits against Meloidogyne incognita.Ann. Appl. Biol. 156, 309e317.
Ntalli, N.G., Cottiglia, F., Bueno, C.A., Alché, L.E., Leonti, M., Vargiu, S., Bifulco, E.,Menkissoglu-Spiroudi, U., Caboni, P., 2010b. Cytotoxic tirucallane triterpenoidsfrom Melia azedarach fruits. Molecules 15, 5866e5877.
Ntalli, N.G., Vargiu, S., Menkisoglu-Spiroudi, U., Caboni, P., 2010c. NematicidalCarboxylic acids and aldehydes fromMelia azedarach fruits. J. Agric. Food Chem.58, 11390e11394.
Oka, Y., 2010. Mechanisms of nematode suppression by organic soil amendments.Appl. Soil Ecol. 44, 101e115.
Orvar, B.L., Ellis, B.E., 1997. Transgenic tobacco plants expressing antisense RNA forcytosolic ascorbate peroxidase show increased susceptibility to ozone injury.Plant J. 11, 1297e1305.
Perez, M.P., Navas-Cortes, J.A., Pascual-Villalobos, M.J., Castillo, P., 2003. Nematicidalactivity of essential oils and organic amendments from Asteraceae against root-knot nematodes. Phytopathology 52, 395e401.
Rachokarn, S., Piyasaengthong, N., Bullangpoti, V., 2008. Impact of botanicalextracts derived from leaf extracts Melia azedarach L. (Meliaceae) and Amar-anthus viridis L. (Amaranthaceae) on populations of Spodoptera exigua (Hübner)(Lepidoptera: Noctuidae) and detoxification enzyme activities. Commun. Agric.Appl. Biol. Sci. 73, 451e457.
Rodriguez-Kabana, R., Morgan-Jones, G., Chet, I., 1987. Biological control of nema-todes: soil amendments and microbial antagonists. Plant Soil 100, 237e247.
Rodriguez-Kabana, R., 1986. Organic and inorganic nitrogen amendments to soil asnematode suppressants. J. Nematol. 18, 129e135.
Sasanelli, N., D’Addabbo, T., 1992. The effect of Cineraria maritima, Ruta graveolensand Tagetes erecta extracts on the hatching of Heterodera schachtii. Nematol.Medit. 20, 49e51.
Sasanelli, N., D’Addabbo, T., 1993. Effect of Cineraria maritima, Ruta graveolens andTagetes erecta leaf and root extracts on Italian populations of Meloidogynespecies. Nematol. Medit. 21, 21e25.
Shigeoka, S., Ishikawa, T., Tamoi,M.,Miyagawa, Y., Takeda, T., Yabuta, Y., Yoshimura, K.,2002. Regulation and function of ascorbate peroxidase isoenzymes. J. Exp. Bot.372, 1305e1319.
Siddiqui, M.A., Alam, M.M., 1987. Efficacy of seed dressing with extracts of neem andPersia lilak against Meloidogyne incognita and Rotylenchulus reniformis. NematolMedit. 15, 399e403.
Simonetti, E., Alba, E., Montes, M.J., Delibes, A., Lopez-Branca, I., 2010. Analysis of ascor-bate peroxidase genes expressed in resistant and susceptible wheat lines infectedby the cereal cyst nematode, Heterodera avenae. Plant Cell Rep. 29, 1169e1178.
Spyrou, I.M., Karpouzas, D.G.,Menkissoglu-Spiroudi, U., 2009. Do botanical pesticidesalter the structure of the soil microbial community? Microb. Ecol. 58, 715e727.
Taylor, A.L., Sasser, J.N., 1978. Biology, Identification and Control of Root-knotNematodes (Meloidogyne spp.). North Carolina State University Graphic,Raleigh, N.C. (U.S.A.), 111 pp.
Toselli, M., Baldi, E., Sorrenti, G., Quartieri, M., Marangon, B., 2010. Evaluation of theeffectiveness of soil-applied plant derivatives of Meliaceae species on nitrogenavailability to peach trees. Sci. Hortic.-Amsterdam 124, 183e188.





























