Embed Size (px)
Citation preview

This article was downloaded by: [Fondren Library, Rice University ]On: 13 November 2014, At: 08:44Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Bolletino di zoologiaPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tizo19
Microanatomy of the anterior median eyes and itspossible relation to polarized‐light reception inLycosa tarentula (Araneae, Lycosidae)Jacqueline Kovoor a , Arturo Muñoz Cuevas a & Joaquin Ortega Escobar ba Laboratoire de Zoologie‐Arthropodes , C.N.R.S.‐M.N.H.N.F , 61 rue de Buffon,Cedex05, Paris, F‐75231, Franceb Area de Psicobiologia , Universidad Autonoma de Madrid , Madrid, E‐28 049, SpainPublished online: 28 Jan 2009.
To cite this article: Jacqueline Kovoor , Arturo Muñoz Cuevas & Joaquin Ortega Escobar (1993) Microanatomy of theanterior median eyes and its possible relation to polarized‐light reception in Lycosa tarentula (Araneae, Lycosidae),Bolletino di zoologia, 60:4, 367-375, DOI: 10.1080/11250009309355841
To link to this article: http://dx.doi.org/10.1080/11250009309355841
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”)contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensorsmake no representations or warranties whatsoever as to the accuracy, completeness, or suitability for anypurpose of the Content. Any opinions and views expressed in this publication are the opinions and viewsof the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sources of information. Taylorand Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses,damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connectionwith, in relation to or arising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Boll. Zool. 60: 367-375 (1993)
Microanatomy of the anterior medianeyes and its possible relation topolarized-light reception inLycosa tarentula (Araneae, Lycosidae)
JACQUELINE KOVOORARTURO MUÑOZ CUEVASLaboratoire de Zoologie-Arthropodes, C.N.R.S.-M.N.H.N.,61 rue de Buffon, F-75231 Paris Cedex 05 (France)
JOAQUIN ORTEGA ESCOBARArea de Psicobiologia, Universidad Autonoma de Madrid,E-28 049 Madrid (Spain)
ABSTRACT
Microanatomy of the anterior median eyes (AME) of a lycosidspider (Lycosa tarentula) shows two classes of photoreceptors in theretina: dorsal and median visual cells (A) are polygonal in cross-section and bear rhabdomeres on all their faces; ventral visual cells(B), rectangular in section, bear rhabdomeres only on two oppositefaces; rhabdomes are aligned in parallel series and successive lines ofrhabdoms are oriented orthogonally to each other. Receptor A cellsare wider than Β cells, and about twice as numerous. In AME opticnerve, axons are also of two types, large and small, and the opticcentre shows thick and narrow fibres, from the second orderneurons, being separated by connective and glial cells. Anteriormedian eyes of second stage juveniles already present the samestructure as those of adults. A similar anatomical structure waspreviously described in the AME of an agelenid spider by Schröer(1974). Since the detection and analysis of polarized light by agelenidand lycosid AME have also been demonstrated, it is suggested thatthe analysers of the e-vector of sky-polarized light are in the receptorΒ cells and that the analysis is a successive process using a twisting ofthe retina due to the action of two muscles.
KEY WORDS: Spiders - Retina - Photoreceptor classes - Polarized-light reception - Histology.
ACKNOWLEDGEMENTS
We are very grateful to Prof. Michael F. Land who accepted thetask of improving the English. His fine comments and suggestions onthe manuscript are greatly appreciated. Thanks are also due toanonymous referees for their pertinent remarks.
(Received 16 February 1993 - Accepted 25 June 1993)
INTRODUCTION
The structure and functioning of the visual system arepoorly known in many families of spiders, mainly on thecommon assumption that most of these are found in darkenvironments and that chemo- and mechanoreceptionrather than vision would be the major senses involved inany guiding activity. Apart from the intensively studied,and thus best-known Salticidae, five other araneomorphfamilies - Lycosidae, Pisauridae, Thomisidae, Oxyopidaeand Deinopidae - show visually guided behaviour duringlocomotion, homing, prey capture and courtship(Bristowe & Locket, 1926; Homann, 1931; Kaston, 1936;Crane, 1949; Whitcomb & Eason, 1965; Robinson &Robinson, 1971; Blest & Land, 1977; Forster, 1982; Uetz& Stratton, 1982).
In lycosid spiders, a complex group of about 2500species very distinct in habitats and habits, fourcamera-like eyes are arranged in three rows; they are dif-ferently oriented, structured and sized, and it can beassumed that each pair of eyes is specialized for particularvisual tasks which are different from that of the otherpairs (Homann, 1931; Magni & Strata, 1965; Carricaburuet al., 1990), although the anterior lateral, posteriormedian and posterior lateral eyes do act as a unit in manycases.
Polarized light is known to be perceived through thevisual system of some spiders. Information relating to thedirection of the polarization plane (e-vector) of skylight isused at any time of the day when clear sky is visible forthe orientation of agelenids and lycosids to their retreator a particular part of their web (Papi, 1955a, b; Papi etal., 1957; Görner, 1958, 1962; Tretzel, 1961; Papi &Tongiorgi, 1963). Görner (1962) showed by experimentalblinding of the anterior median eyes that Agelenalabyrinthica could no longer orientate to polarized light.It was confirmed by electroretinography that the anteriormedian eyes (AME) of Arctosa variana C.L. Koch werethe main receptors for polarized light (Magni et al., 1964,1965; Magni, 1966) but Baccetti & Bedini (1964) failed todetect in the AME of the same species a specific structurewhich might be responsible for the detection of the pat-tern of polarization of the sky.
Two modes of arrangement of the rhabdoms havebeen found in the AME retina of Lycosa erythrognathaLucas ( = L. raptoria Walckenaer) by Melamed & TrujilloCenoz (1966). In the peripheral parts of the retina, therhabdoms are oriented either parallel or perpendicular tothe retinal radii; on the other hand, in the central region,rhabdoms are oriented in other directions. Theperipheral region might be suitable for the perception oflight polarization; but no physiological experiment hasbeen performed to confirm or invalidate this sup-position. It seemed unlikely to Waterman (1981, p. 423)for «adding all the radial or tangential inputs would can-cel out e-vector discrimination, at least optically, if thesystem is radially symmetrical».
The opportunity of associating anatomical,
Dow
nloa
ded
by [
Fond
ren
Lib
rary
, Ric
e U
nive
rsity
] a
t 08:
44 1
3 N
ovem
ber
2014

368 J. KOVOOR, A. MUÑOZ CUEVAS, J. ORTEGA ESCOBAR
physiological and behavioural studies on the visualsystem of Lycosa tarentula leads us to examine moreprecisely the structure of AME retina, optic nerve and op-tic centres, with the aim of finding the structures respon-sible for the detection and analysis of natural polarizedlight.
MATERIAL AND METHODS
Specimens of Lycosa tarentula (L.), a burrowing lycosid spider,were collected in the vicinity of Canto Blanco (Madrid, Spain). Adultmales and females were kept in terrariums prior to use. Somejuveniles (first and second instars) hatched from egg cases collectedwith the females in the field were also used for the histophysiologicalanalysis of the visual system.
Prosomas of specimens fixed in Bouin's fluid for 16 h (juveniles)or 24 h (adults) were dehydrated in ethanol and preserved for threeweeks (juveniles) or four months (adults) in n-butanol before embed-ding in paraffin; serial sections (5-6 μιτι thick) were made in the threeorthogonal planes. As in a previous paper (Kovoor et al., 1992), sec-tions were stained using Heidenhain's axan and Gabe's one-steptrichrome alone or in combination with aldehyde fuchsin.Histochemical reactions and other staining methods were used tovisualize a) anionic groups and polysaccharidic substances: P.A.S.reaction, alcian blue at pH 2.5 and toluidine blue at pH 4.2; b)proteins: Danielli's coupled tetrazonium reaction, aldehyd fuchsinstaining of sections previously oxydized by permanganate, leadhematoxylin and phosphotungstic hematoxylin stainings whichwere particularly useful to demonstrate, respectively, nerve fibresand glial cells, and Adams's ferric ferricyanide reaction for pigmentcells and reducing metabolic products. Technical procedures aredetailed in the handbooks of Gabe (1968) and Lillie & Fullmer(1976).
Semithin sections of juvenile AME retinal cells were obtained fromspecimens fixed for transmission electron microscopy. Juveniles,just after leaving the egg case (second instar) were fixed in 3%glutaraldehyde in 0.1 M cacodylate buffer at pH 7.2-7.4, at roomtemperature for two hours, and after washing in buffer, postfixed in2% osmium tetroxide in the same buffer, for one hour. Specimenswere then dehydrated in an alcohol series and propylene oxide andembedded in Epon. Transverse and sagittal sections, 0.5 to 0.8 μπιthick, were stained by 1 % toluidine blue-l % azur II in 1 % borax,mounted in a synthetic resin, observed and photographed through aLeitz Ortholux microscope.
Numbers of AME photoreceptor cells of one male and five adultfemales were determined by counting the axons in transverse sec-tions of the optic nerve. Rhabdoms of the ventral part of adult AMEretinae were also counted on complete series of transverse sections.
RESULTS
General aspect of the anterior median eyes (AME)
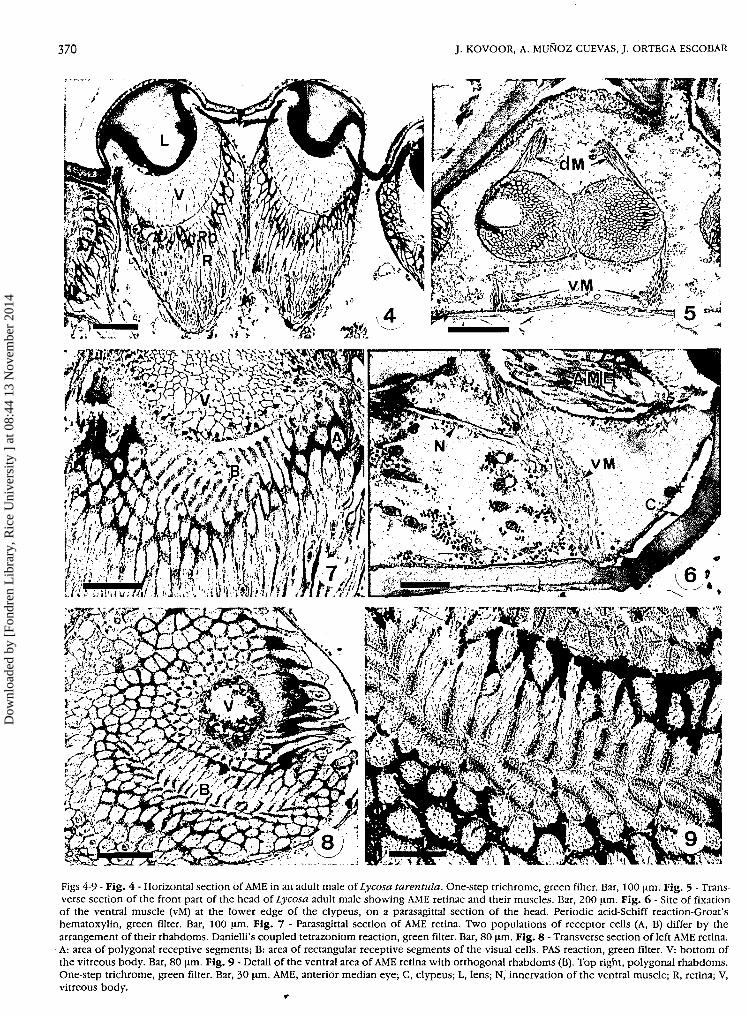
A general side view of the prosoma of Lycosa taren-tula adult female showed two small anterior eyes of thefront row: one median (AME) and one lateral (ALE); overthem, a very large eye of the second row (posteriormedian eye, PME) and one of the third row (posteriorlateral, PLE) on the rear part of the head (Fig. 1). Opticaxes of AME were oriented 20° upwards and 15° lateralto the sagittal plane of the prosoma, as shown on Figures2 and 3· In transverse sections, AME were almost circular:both retinae presented a contact zone in the sagittal plane
of the prosoma (Figs 4, 5). Histological studies showedthat the AME of L. tarentula were not radially sym-metrical.
Ocular muscles
AME retina is movable by the action of two antagonistmuscles attached obliquely to its lateral external surface(Scheuring, 1914). The dorsal muscle is fastened to thefront upper epidermis of the clypeus at 450 μπι, from theoptic axis of AME, and 80-90 μπι lateral to the generalsagittal plane (All measurements were made on adultfemales). The ventral muscle runs to the ventral edge ofthe clypeus at 500 μπι from the optic axis where it joinsthe epidermis at ca 400 μπι from the sagittal plane (Figs 5,6). The alternating contraction of these two muscles cangenerate rotative and, to some extent, up and downmovements of the retinal cup.
Microanatomy of AME retina and optic nerve in adultmales and females
The lens is biconvex, with a flattened corneal surface.Beneath this, the high cells of the vitreous body form acrescent, the basal part of which rests on the preretinalmembrane. The retina underlies this connective mem-brane, forming a thick cup (250 μπι deep at the most)made up of two types of cells: photoreceptor andpigment or glial cells, enveloped by the postretinal mem-brane. The axonal extensions of the photoreceptor cellsjoin together in a bunch at the bottom of the retinal cup,forming the optic nerve which ends in the optic lobes ofthe supraoesophageal ganglion (brain), where synapticconnections take place with ganglionic cells (Fig. 3)·
Photoreceptor cells are in close contact with eachother in their apical part where rhabdomeres are borne.The medial segments of the photoreceptors, which con-tain the nuclei, are clearly separated from each other bythe ramified extensions of the pigment cells. In the apicalsegment, ramifications of pigment cells were detectedonly as more or less thick intensely stained lines betweenthe rhabdomeres of neighbouring cells (Figs 4, 7-9).
Two distinct modes of arrangement of the photorecep-tor cells were found in AME retina. In the median anddorsal parts, polygonal visual cells (A) surrounded bypigment cells were juxtaposed on three to five faces, allof which bore rhabdomeres. Transverse sections of theseregions showed a mosaic aspect (Fig. 8A). In the ventralpart of the retina, visual cells had a rectangular transversesection and were differently structured and arranged. Anumber of them were aligned in two sets of rows, oneextending on each side of the sagittal plane of the eye;cells were joined to one another by their two oppositelateral faces bearing rhabdomeres and forming series ofparallel rhabdoms (B). The apical end of these cells facedthe light. Two successive rows of these cells seemed con-tinuous ventrally, where still another arrangement of thephotoreceptor cells occurred (Fig. 7). Here other rows ofcells were found whose parallel rhabdoms were oriented
Dow
nloa
ded
by [
Fond
ren
Lib
rary
, Ric
e U
nive
rsity
] a
t 08:
44 1
3 N
ovem
ber
2014

MEDIAN EYES AND POLARIZED LIGHT IN A LYCOSID SPIDER 369
Γ P M E ALEPLE>~
Figs 1 -2 - Side and front views of the head of a Lycosa tarentula adult female. Fig. 1 - The optic axis of the anterior median eye (AME) is orien-ted at 20° upward. Fig. 2 - Each anterior median eye is oriented at 15° lateral to sagittal plane of the prosoma. ALE, anterior lateral eye; PLE,posterior lateral eye; PME, posterior median eye. Fig. 1: Bar, 470 μηΐ; Fig. 2: Bar, 230 μπι.
ALE
14,15
Fig. 3 - Diagrammatic horizontal scheme from the AME to the opticlobes of the CNS. Levels of sections illustrated in Figures 7, 8, 10-15are indicated with arrows and corresponding numbers. ALE, anteriorlateral eye; AME, anterior median eye; L, lens; OL, optic lobe of thebrain.
at right angles to those of the preceding rows, and theyseparated these rows from the mosaic type of the centralregion (Figs 7-9). Light entered these intermediary cellsfrom the side.
For all receptor cells, the rhabdomere height, from 20to 30 μπι, and the length, from 1.2 to 2.1 μπι, were foundto depend on the general size of the individuals and thephase of the circadian rhythm of illumination duringwhich they were fixed (Fig. 9)· The dorsal and medianphotoreceptor cells appeared generally wider (A cells, 13to 15 μπι in adult females) than the ventral ones (B cells,8.5 to 9.5 μηι) (Figs 7, 8, 10).
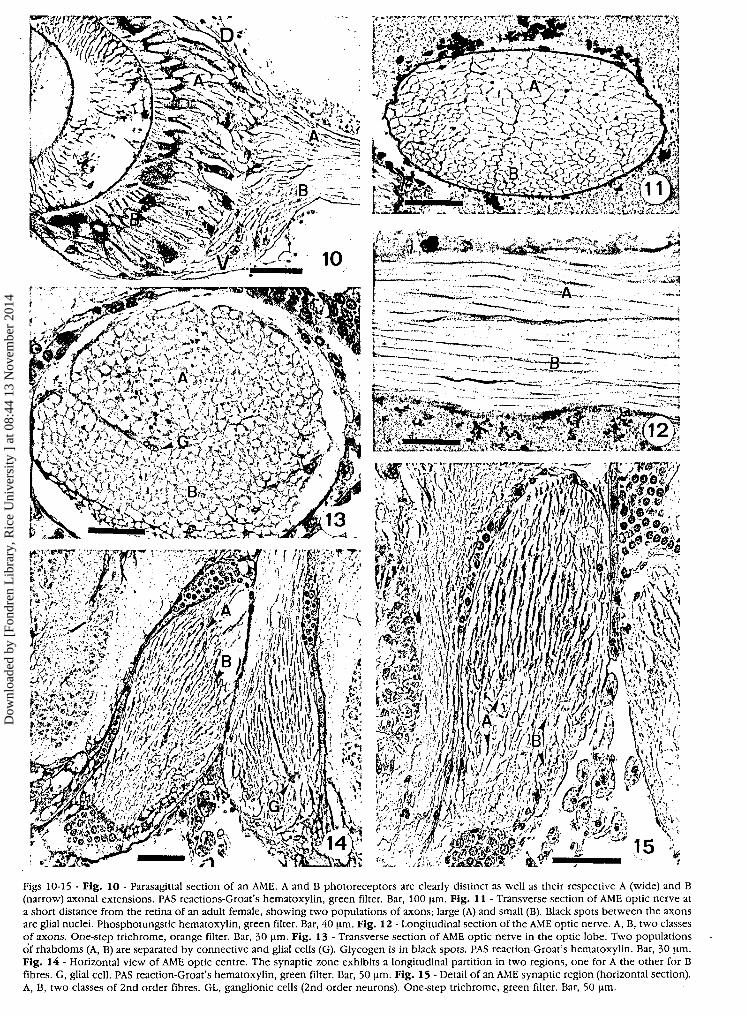
The retinal cup was limited externally by the connec-tive postretinal membrane which continued as the en-velope of the optic nerve from the bottom of the AMEretina. Transverse sections of the optic nerve, close tothe retina, showed more or less flattened oval profiles.Sections of axons could be assigned to two distinctclasses (A and B) according to their diameter (Fig. 11).
Total counts of axons in AME optic nerve of one maleand several females varied little between individuals(Table I). Counts of Β rhabdoms on serials sections ofAME retina were identical for a couple of specimens. Infemale 21, direct counts of Β rhabdoms were nearly thesame as the number of B axons in the optic nerve(Table I).
In longitudinal sections of the nerve, two types offibres could be recognized (Fig. 12). Inside the opticlobes, the optic nerve was partitioned in two regions,one for A fibres, the other for Β fibres, by a strand of con-nective tissue and glial cells (Fig. 13)· This partition wasstill visible in the synaptic zone, where thick and narrowfibres corresponded, respectively, to A and Β populationsof the photoreceptor cells (Figs 14, 15).
Dow
nloa
ded
by [
Fond
ren
Lib
rary
, Ric
e U
nive
rsity
] a
t 08:
44 1
3 N
ovem
ber
2014
370 J. KOVOOR, A. MUÑOZ CUEVAS, J. ORTEGA ESCOBAR
m$&&
Figs 4-9 - Fig. 4 - Horizontal section of AME in an adult male of Lycosa tarentula. One-step trichrome, green filter. Bar, 100 μπι. Fig. 5 - Trans-verse section of the front part of the head of Lycosa adult male showing AME retinae and their muscles. Bar, 200 μηπ. Fig. 6 - Site of fixationof the ventral muscle (vM) at the lower edge of the clypeus, on a parasagittal section of the head. Periodic acid-Schiff reaction-Groat'shematoxylin, green filter. Bar, 100 μΐη. Fig. 7 - Parasagittal section of AME retina. Two populations of receptor cells (A, B) differ by thearrangement of their rhabdoms. Danielli's coupled tetrazonium reaction, green filter. Bar, 80 μΐη. Fig. 8 - Transverse section of left AME retina.A: area of polygonal receptive segments; B: area of rectangular receptive segments of the visual cells. PAS reaction, green filter. V: bottom ofthe vitreous body. Bar, 80 μηι. Fig. 9 - Detail of the ventral area of AME retina with orthogonal rhabdoms (B). Top right, polygonal rhabdoms.One-step trichrome, green filter. Bar, 30 μπι. ΑΜΕ, anterior median eye; C, clypeus; L, lens; N, innervation of the ventral muscle; R, retina; V,vitreous body.
Dow
nloa
ded
by [
Fond
ren
Lib
rary
, Ric
e U
nive
rsity
] a
t 08:
44 1
3 N
ovem
ber
2014
^„.„^
lifiI ¿ f t * ' Λ · ' ' · «ν » ' · , • >•
Figs 10-15 - Flg. 10 - Parasagittal section of an AME. A and Β photoreceptors are clearly distinct as well as their respective A (wide) and Β(narrow) axonal extensions. PAS reactions-Groat's hematoxylin, green filter. Bar, 100 μΐη. Fig. 11 - Transverse section of AME optic nerve ata short distance from the retina of an adult female, showing two populations of axons; large (A) and small (B). Black spots between the axonsare glial nuclei. Phosphotungstic hematoxylin, green filter. Bar, 40 μΐη. Fig. 12 - Longitudinal section of the AME optic nerve. A, B, two classesof axons. One-step trichrome, orange filter. Bar, 30 μΐη. Fig. 13 - Transverse section of AME optic nerve in the optic lobe. Two populationsof rhabdoms (A, B) are separated by connective and glial cells (G). Glycogen is in black spots. PAS reaction-Groat's hematoxylin. Bar, 30 μΐη.Fig. 14 - Horizontal view of AME optic centre. The synaptic zone exhibits a longitudinal partition in two regions, one for A the other for Βfibres. G, glial cell. PAS reaction-Groat's hematoxylin, green filter. Bar, 50 μΐη. Fig. 15 - Detail of an AME synaptic region (horizontal section).A, B, two classes of 2nd order fibres. GL, ganglionic cells (2nd order neurons). One-step trichrome, green filter. Bar, 50 μηι.
Dow
nloa
ded
by [
Fond
ren
Lib
rary
, Ric
e U
nive
rsity
] a
t 08:
44 1
3 N
ovem
ber
2014
372 J. KOVOOR, A. MUÑOZ CUEVAS, J. ORTEGA ESCOBAR
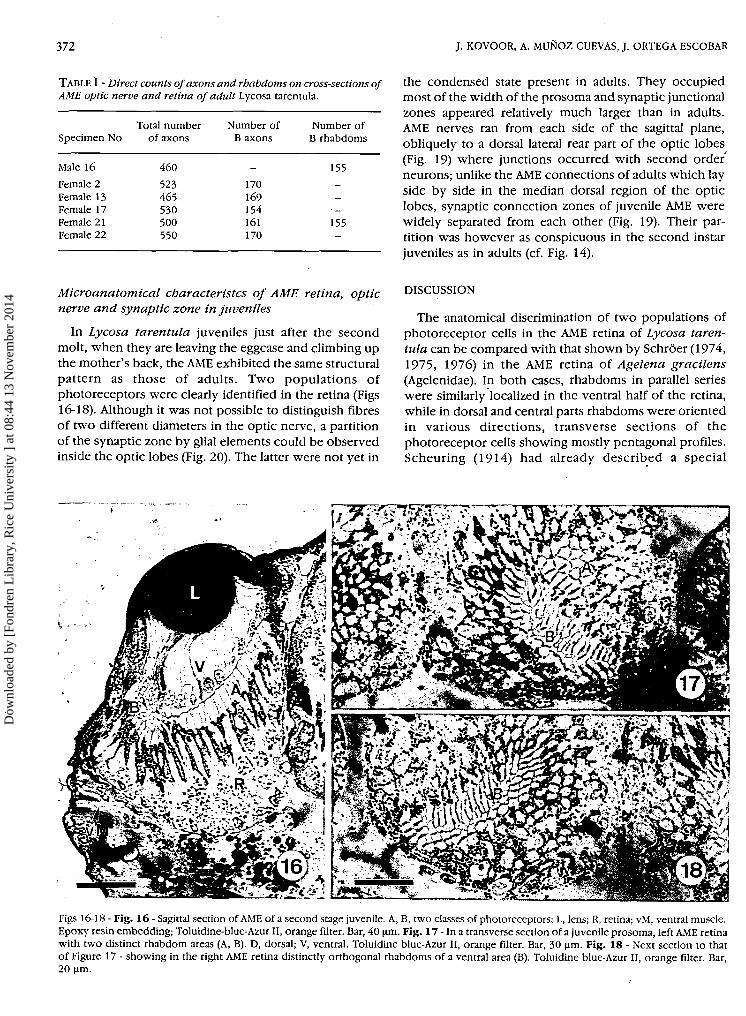
TABLE I - Direct counts of axons and rhabdoms on cross-sections ofAME optic nerve and retina of adult Lycosa tarentula.
Specimen No
Male 16
Female 2Female 13Female 17Female 21Female 22
Total numberof axons
460
523465530500550
Number ofΒ axons
_
170
169154161170
Number ofΒ rhabdoms
155__
· -
155-
the condensed state present in adults. They occupiedmost of the width of the prosoma and synaptic junctionalzones appeared relatively much larger than in adults.AME nerves ran from each side of the sagittal plane,obliquely to a dorsal lateral rear part of the optic lobes(Fig. 19) where junctions occurred with second order'neurons; unlike the AME connections of adults which layside by side in the median dorsal region of the opticlobes, synaptic connection zones of juvenile AME werewidely separated from each other (Fig. 19). Their par-tition was however as conspicuous in the second instarjuveniles as in adults (cf. Fig. 14).
Microanatomical characteristcs of AME retina, opticnerve and synaptic zone in juveniles
In Lycosa tarentula juveniles just after the secondmolt, when they are leaving the eggcase and climbing upthe mother's back, the AME exhibited the same structuralpattern as those of adults. Two populations ofphotoreceptors were clearly identified in the retina (FigsI6-I8). Although it was not possible to distinguish fibresof two different diameters in the optic nerve, a partitionof the synaptic zone by glial elements could be observedinside the optic lobes (Fig. 20). The latter were not yet in
DISCUSSION
The anatomical discrimination of two populations ofphotoreceptor cells in the AME retina of Lycosa taren-tula can be compared with that shown by Schröer (1974,1975, 1976) in the AME retina of Agelena gracilens(Agelenidae). In both cases, rhabdoms in parallel serieswere similarly localized in the ventral half of the retina,while in dorsal and central parts rhabdoms were orientedin various directions, transverse sections of thephotoreceptor cells showing mostly pentagonal profiles.Scheuring (1914) had already described a special
ΜΜΜΜ,WÉËMË
^• ÎÂ^ Ï 'V ·^ - -^ ' - i- > ¿ ' - * . ' -
Figs 16-18-Fig. 16-Sagittal section of AME of a second stage juvenile. A, B, two classes of photoreceptors; L, lens; R, retina; vM, ventral muscle.Epoxy resin embedding; Toluidine-blue-Azur II, orange filter. Bar, 40 μπι. Fig. 17 - In a transverse section of a juvenile prosoma, left AME retinawith two distinct rhabdom areas (A, B). D, dorsal; V, ventral. Toluidine blue-Azur II, orange fllter. Bar, 30 μπι. Fig. 18 - Next section to thatof Figure 17 - showing in the right AME retina distinctly orthogonal rhabdoms of a ventral area (B). Toluidine blue-Azur II, orange fllter. Bar,20 μΐη.
Dow
nloa
ded
by [
Fond
ren
Lib
rary
, Ric
e U
nive
rsity
] a
t 08:
44 1
3 N
ovem
ber
2014

MEDIAN EYES AND POLARIZED LIGHT IN A LYCOSID SPIDER 373
Figs 19-20 - Transverse section of the prosoma of a second instar juvenile. Fig. 19 - A synaptic zone (arrows) of AME on each side, surroundedby glial cells, in the rear part of the optic lobes of the brain. Left side, connection with the neuropile is visible; right side, the synaptic zoneis clearly bipartite. NS, large neurosecretory cell just next to the synaptic area. Toluidine blue-Azur II, orange filter. Bar, 80 μηι. Flg. 20 - Detailof Figure 19. The AME synaptic region in the optic lobe shows two distinct parts. NS, neurosecretory cell. Bar, 20 μηι.
arrangement of the photoreceptor cells in the AME of cer-tain Lycosidae {Pardosa and Trochosa genera). Thispeculiarity was illustrated by a camera lucid drawing of asagittal section of Trochosa ruricola AME but was not takeninto account in more recent publications on the subject.
Observations by Blest & O'Carroll (1989) of two tiersof receptive segments in the principal retina of Geolycosagodeffroyi ( = Lycosa godeffroyi L. Koch) referred to asingle small unspecified portion of the AME retina andthus could neither be compared nor combined with ourown data. However, as the structure described in L.tarentula seems to be present in the AME of at least threelycosid vagrant or burrowing species belonging to threedistinct genera, it can be inferred that such a structure isa general feature for the family Lycosidae. Preliminarystudies on Peucetia (Oxyopidae) juveniles and adults alsoindicated a double structure of AME retinae (Kovoor etal., unpubl.).
Both classes of photoreceptor cells were detectable inadults as well as in juveniles just after leaving the eggsac(2nd instar). The principal eyes of L. tarentula developduring the first instar that lasts about two weeks. It seemsthat they are able to function only after the second moltwhen both the eyes and the optic centres are fullyorganized. Observations on the predatory behaviour of L.tarentula juveniles actually showed a «maturation» of thebehavioural pattern during the second stage of develop-ment (Ortega Escobar & Fernandez Montraveta, 1985).
The anatomical partitioning of the synaptic zone, at allstages of the development from the second instar, mightindicate that information transferred from each class ofAME receptor cells in the optic lobes is integrated in adistinct area of the brain. Thus, the efferent responsesmight also be of two different types.
The anatomical features of AME retina and optic centredescribed above in L. tarentula can be related to theresults of behavioural and physiological studies on thevisual system of other lycosid spiders. Papi (1955a, b,1959) and Papi et al. (1957) demonstrated the astro-nomical orientation of three Arctosa species to the riverbanks where they lived. From the studies by Magni et al.(1964, 1965) it was known that AME, and also to someextent the PME, were concerned with the detection ofpolarized light. These authors postulated that the abilityto analyze the polarization plane of the light was presentin retinal elements of AME and PME that were «spatiallyarranged in such a way that the majority of them wouldbe activated for a given direction of the polarizationplane» (Magni et al, 1965, p. 155). Without any otherstructural data than those of Baccetti & Bedini (1964) onthe photoreceptor cells of Arctosa which were shown incross section to have a pentagonal profile, with each facebearing a rhabdomere, Görner (cited by von Frisch,1965) concluded that the irregular polygonal sections ofAME photoreceptors might be suited to the reception ofpolarized light. Although AME structure has never beenreinvestigated in Arctosa variana, it seems very unlikelythat it differs markedly from that of other lycosid speciesalready studied.
The large ventral area of AME retinae of L. tarentula,provided with rhabdoms aligned in orthogonal rowsseems to be much better suited to perceive the pattern ofsky polarized light than the polygonal receptor cells inthe central and dorsal regions. Many examples have beendescribed in other arthropods, and particularly in insects,which utilize the polarized light pattern of the sky fornavigation, as demonstrated by von Frisch (1949, 1951)for the bee Apis mellifera (review by Wehner, 1982). In
Dow
nloa
ded
by [
Fond
ren
Lib
rary
, Ric
e U
nive
rsity
] a
t 08:
44 1
3 N
ovem
ber
2014

374 J. KOVOOR, A. MUÑOZ CUEVAS, J. ORTEGA ESCOBAR
bees and ants (Schinz, 1975; Aepli et al, 1985; Wehner,1989) as well as in Orthoptera (Burghause, 1979; Egelhaaf& Dambach, 1983), Díptera (Wunderer & Smola, 1982;Hardie, 1984; Strausfeld & Wunderer, 1985; Fortini &Rubin, 1991) and Lepidoptera (Kolb, 1986, Hämmerle &Kolb, 1987) compound eyes comprise a special area atthe dorsal rim of the retina which contains ommatidiawith rhabdoms arranged in two orthogonal directions.The photoreceptors of this special area which has beencalled «POL area» by Wehner & Strasser (1985) wouldconstitute a two-channel system analyzing e-vectors ofpolarized light - via the oriented molecules of rhodopsinpacked inside the microvillar membrane (Wehner &Rössel, 1985).
As Schröer (1974-1976) pointed out for Agelenagracilens, the movements of the AME retina cup elicitedby two muscles make it possible that the analysis ofe-vector of polarized light is a successive process at leastin agelenid and lycosid spiders. In Lycosa tarentula, thelocation of particular rhabdoms in the ventral part of theretina, and the orientation of AME optic axis at 20° up-ward are suitable for the spider to scan the sky fromeither inside or outside its burrow.
Although the organization of the retinae in mostspiders would permit the analysis of polarized light(Land, 1985), this ability was demonstrated only in twocases other than those of agelenids and lycosids:Aphonopelma californica, a mygalomorph spider, couldorientate to polarized light (Henton & Crawford, 1966);the process of analysis was thought to be intra-ocular butthe structure and physiology of the retinae have not yetbeen studied in Mygalomorphae. From structural obser-vations, Land (1969) suggested that Layer IV of AMEretinae of salticids (Phidippus sp.) might be concernedwith the detection of the polarization plane of light(Eakin & Brandenburger, 1971); but an experimentalstudy of the orientation of Phidippus pulcherrimus toprey position during indirect pursuit (Hill, 1979) did notprovide evidence to indicate the utilization of thepolarized light pattern. On the other hand, experimentson single receptor cells of the posterior lateral eyes ofPlexippus sp. actually showed a weak reaction to therotation of the polarization plane (Hardie & Duelli, 1978).
It will be of interest to elucidate precisely the im-plications of the structure described in the anteriormedian eyes of Lycosa tarentula, and in particular thedevelopmental aspects, on the visually guided behaviourof these lycosid spiders in both phases of the light cycle.
REFERENCES
Aepli F., Labhart T., Meyer Ε. P., 1985 - Structural specializations ofthe cornea and retina at the dorsal rim of the compound eyes inhymenopteran insects. Cell Tissue Res., 239: 19-24.
Baccetti B., Bedini C., 1964 - Research on the structure andphysiology of the eyes of a lycosid spider. I. Microscopic andultramicroscopic structure. Arch. ital. Biol., 102: 97-112.
Blest A. D., Land M. E., 1977 - The physiological optics of Dinopissubrufus L. Koch: a fish-lens in a spider. Proc. Roy. Soc. London,Ser. B, 196: 198-222.
Blest D. Α., O'Carroll D., 1989 - The evolution of the tiered principalretinae of jumping spiders (Araneae: Salticidae). In: R. N. Singh &N. J. Strausfeld (eds), Neurobiology of sensory systems. PlenumPress, New York, pp. 155-170.
Bristowe W. S., Locket G. H., 1926 - The courtship of British lycosidspiders, and its probable significance. Proc. zool. Soc., Lond., Pt.2, N° 22: 317-347.
Burghause F. M. H. R., 1979 - Die strukturelle Spezialisierung des dor-salen Augenteils der Grillen (Orthoptera, Grylloidea). Zool.Jahrb., Physiol., 83: 502-525.
Carricaburu P., Muñoz Cuevas Α., Ortega Escobar J., 1990 - Elec-troretinography and circadian rhythm in Lycosa tarentula(Araneae, Lycosidae). Acta zool. fenn., 190: 63-67.
Crane J., 1949 - Comparative biology of salticid spiders at RanchoGrande, Venezuela. Part IV. An analysis of display. Zoologica, 34:159-214.
Eakin R. M., Brandenburger J. L., 1971 - Fine structure of the eyes ofjumping spiders. J. Ultrastr. Res., 37: 618-663.
Egelhaaf Α., Dambach M., 1983 - Giant rhabdoms in a specializedregion of the compound eye of a cricket: Cycloptiloidescanariensis (Insecta, Gryllidae). Zoomorphol., 102: 65-77.
Forster L., 1982 - Visual communication in jumping spiders(Salticidae). In: P. N. Witt & J. S. Rovner (eds), Spider com-munication. Mechanisms and ecological significance. PrincetonUniversity Press, Princeton, pp. 161-212.
Fortini Μ. Ε., Rubin G. M., 1991 - The optic lobe projection patternof polarization-sensitive photoreceptor cells in Drosophilamelanogaster. Cell Tis. Res., 265: 185-191.
Frisch Κ. von, 1949 - Die Polarisation des Himmelslichtes als orien-tierender Faktor bei den Tanzen der Bienen. Experientia, 5:142-148.
Frisch K. von, 1951 - Orientierungsvermögen und Sprache derBienen. Naturwiss., 38: 105-112.
Frisch K. von, 1965 - Tanzsprache und Orientierung der Bienen.Springer-Verlag, Berlin, 578 pp.
Gabe M., 1968 - Techniques histologiques. Masson, Paris, 1113 pp.Görner P., 1958 - Die optische und kinästhetische Orientierung der
Trichterspinne Agelena labyrinthica (Cl.). Z. vergl. Physiol., 41:111-153.
Görner P., 1962 - Die Orientierung der Trichterspinne nachpolariziertem Licht. Z. vergl. Physiol., 45: 307-314.
Hämmerle B., Kolb G., 1987 - Structure of the dorsal eye region ofthe moth, Adoxophyes reticulana Hb. (Lepidoptera, Tortricidae).Int. J. Ins. Morphol. Embryol., 16: 121-129.
Hardie R. C., 1984 - Properties of photoreceptors R7 and R8 in dorsalmarginal ommatidia in the compound eyes of Musca andCalliphora. J. comp. Physiol., 154: 157-165.
Hardie R. C., Duelli P., 1978 : Properties of single cells of posteriorlateral eyes of jumping spiders. Z. Naturforsch., 33c: 156-158.
Henton W. W., Crawford F. T., 1966: The discrimination of polarizedlight by the Tarantula. J. comp. Physiol., 52: 26-32.
Hill D. E., 1979 : Orientation by jumping spiders of the genusPhidippus (Salticidae) during the pursuit of prey. Behav. Ecol.Sociobiol., 5: 301-322.
Homann H., 1931: Beiträge zur Physiologie der Spinnen Augen. III.Das Sehvermögen der Lycosiden. Z. vergl. Physiol., 11: 40-67.
Kaston B. J., 1936 - The senses involved in the courtship of somevagabond spiders. Entomol. Am., 16: 97-166.
Kolb G., 1986 - Retinal ultrastructure in the dorsal rim and large dor-sal area of the eye of Aglais urticae (Lepidoptera). Zoomorphol.,106: 244-246.
Kovoor J., Muñoz Cuevas Α., Ortega Escobar J., 1992 - Le systèmevisuel de Lycosa tarentula fasciiventris (Araneae, Lycosidae). I.Organisation des nerfs et des premiers ganglions optiques. Ann.Sci. nat., Zool., Paris, 13e Ser., 13: 25-36.
Land M. F., 1969 - Structure of the retinae of the principal eyes ofjumping spiders (Salticidae: Dendryphantinae) in relation tovisual optics. J. exp. Biol., 51: 443-470.
Dow
nloa
ded
by [
Fond
ren
Lib
rary
, Ric
e U
nive
rsity
] a
t 08:
44 1
3 N
ovem
ber
2014

MEDIAN EYES AND POLARIZED LIGHT IN A LYCOSID SPIDER 375
Land M. F., 1985 - The morphology and optics of spider eyes. In: G.H. Barth (ed.). Neurobiology of arachnids. Springer-Verlag,Heidelberg, New-York, Tokyo, pp. 58-78.
Lillie R. D., Fullmer H. M., 1976 - Histopathologic technic and prac-tical histochemistry (4th ed.). Mc Graw Hill Book Company, NewYork, 942 pp.
Magni F., 1966 - Analysis of polarized light in wolf spiders. In: C. G.Bernhard (ed.), The functional organization of the compoundeye. Pergamon Press, Oxford, London, pp. 171-186.
Magni F., Strata P., 1965 - Electroretinographic responses from theeyes of the wolf spider Lycosa tarentula (Rossi). Arch. ital. Biol.,103: 694-704.
Magni F., Papi F., Savely H. E., Tongiorgi P., 1964 - Research on thestructure and physiology of the eyes of a lycosid spider. II. Therole of different pairs of eyes in astronomical orientation. Arch.ital. Biol., 102: 123-136.
Magni F., Papi F., Savely H. E., Tongiorgi P., 1965 - Research on thestructure and physiology of the eyes of a lycosid spider. III. Elec-troretinographic responses to polarized light. Arch. ital. Biol.,103: 136-158.
Melamed J., Trujillo-Cenoz O., 1966 - The fine structure of the visualsystem of Lycosa (Araneae: Lycosidae). Part I. Retina and opticnerve. Z. Zellforsch., 74: 12-31.
Ortega Escobar J., Fernandez Montraveta C., 1985 - Study aboutdevelopment of predatory behaviour in a wolf spider (Lycosatarentula fasciiventris). Proc. 19th Int. ethol. Conf., Toulouse,338.
Papi F., 1955 a - Astronomische Orientierung bei der Wolfspinne, Ar-ctosa perita (Latr.). Z. vergl. Physiol., 3: 230-233.
Papi F., 1955b - Ricerche sull'orientamento astronomico di Arctosaperita (Latr.) (Araneae-Lycosidae). Pubbl. Stn. Zool. Napoli, 27:76-103.
Papi F., 1959 - Sull'orientamento astronomico in specie del gen. Ar-ctosa (Araneae, Lycosidae). Z. vergl. Physiol., 41: 481-489.
Papi F., Tongiorgi P., 1963 - Innate and learned components in theastronomical orientation of wolf spiders. In: H. Autrum (ed.),Orientierung der Tiere. Symp. Garmisch-Partenkirchen, Ergebn.Biol., 26: 259-280.
Papi F., Serretti L., Parrini S., 1957 - Nuove ricerche sull'orientamen-to e il senso del tempo di Arctosa perita (Latr.) (Araneae-Lycosidae). Ζ. vergl. Physiol., 39: 531-561.
Robinson M. H., Robinson B., 1971 - The predatory behaviour of theogre-faced spider Dinopis longipes F. Cabridge. Am. Midi. Nat,85: 85-96.
Scheuring L., 1914 - Die Augen der Arachnoiden. II. Zool. Jahrb.,Anat., 37: 369-464.
Schinz R. H., 1975 - Structural specialization in the dorsal retina ofthe bee, Apis mellifera. Cell Tissue. Res., 162: 23-34.
Schröer W. D., 1974 - Zum Mechanismus der Analyse polariziertenLichtes bei Agelena gracilens C. L. Koch (Araneae, Agelenidae).Z. Morphol. Tiere, 79: 215-231.
Schröer W. D., 1975 - Polarized light detection in an agelenid spider,Agelena gracilens (Araneae, Agelenidae). Proc. 6th Int.Arachnological Congress, Amsterdam, pp. 191-194.
Schröer W. D., 1976 - Polarisations Empfindlichkeit rhabdomeralerSystem in der Hauptaugen der Trichterspinne Agelena gracilens(Arachnida: Araneae: Agelenidae). Entomol. gen., 3: 88-92.
Strausfeld N. J., Wunderer H., 1985 - Optic lobe projections ofmarginal ommatidia in Calliphora erythrocephala specialized fordetecting polarized light. Cell Tissue Res., 242: 163-178.
Tretzel Ε., 1961 - Biologie, Ökologie und Brutpflege von Coelotesterrestris (Wider) (Araneae, Agelenoidae). I. Biologie undÖkologie. Z. Morphol. Ökol. Tiere, 49: 658-745.
Uetz G. W., Stratton G. E., 1982 - Acoustic communication andreproductive isolation in spiders. In: P. N. Witt & J. S. Rovner(eds), Spider communication. Mechanisms and ecologicalsignificance. Princeton University Press, Princeton, U.S.A., pp.123-469.
Waterman T. H., 1981 - Polarization sensitivity. In: H. Autrum (ed.),Comparative physiology and evolution of vision in invertebrates.I., Handb. sens. Physiol. VII/6B, Springer-Verlag, Berlin, pp.283-469.
Wehner R., 1982 - Himmelsnavigation bei Insekten. Neurophysiologieund Verhalten. Neujahrblatt naturf. Ges., Zürich, 184: 1-132.
Wehner R., 1989 - The hymenopteran skylight compass: matchedfiltering and parallel coding. J. exp. Biol., 146: 63-85.
Wehner R., Rössel S., 1985 - The bee's celestial compass. A casestudy in bee neurobiology. Fortschr. Zool., 31: 11-53.
Wehner R., Strasser S., 1985 - The POL area of the honey bee's eye:behavioral evidence. Physiol. Entomol., 10: 337-349.
Whitcomb W. H., Eason R., 1965 - The mating behaviour of Peucetiaviridans (Araneida, Oxyopidae). Florida Entomol., 48: 163-167.
Wunderer H., Smola U., 1982 - Fine structure of ommatidia at thedorsal eye margin of Calliphora erythrocephala Meigen (Diptera:Calliphoridae): an eye region specialized for the detection ofpolarized light. Int. J. Ins. Morphol. Embryol., 11: 25-38.
Dow
nloa
ded
by [
Fond
ren
Lib
rary
, Ric
e U
nive
rsity
] a
t 08:
44 1
3 N
ovem
ber
2014





























