Embed Size (px)
Citation preview

Free Radical Biology & Medicine, Vol. 8, pp. 145-152, 1990 0891-5849/90 $3.00 + .00 Printed in the USA. All rights reserved. © 1990 Pergamon Press plc
Original Contribution
MICROSOMAL LIPID PEROXIDATION: THE ROLE OF NADPH--CYTOCHROME P450 REDUCTASE AND CYTOCHROME P450
ALEX SEVANIAN, KERSTIN NORDENBRAND,* EUNJOO KIM, LARS ERNSTER,* and PAUL HOCHSTEIN
Institute for Toxicology, University of Southern California, Los Angeles, CA 90033, U.S.A.
(Received 13 April 1989; Revised 29 August 1989; Accepted 13 October 1989)
A b s t r a c t - - T h e role of NADPH - - cytochrome P450 reductase and cytochrome P450 in NADPH- and ADP - - Fe 3÷-dependent lipid peroxidation was investigated by using the purified enzymes and liposomes prepared from either total rat-liver phospholipids or a mixture of bovine phosphatidyl choline and phosphatidyl ethanolamine (PC/PE liposomes). The results suggest that NADPH- and A D P - Fe 3 ÷-dependent lipid peroxidation involves both N A D P H - cytochrome P450 reductase and cytochrome P450. Just as in the case of cytochrome P450-1inked monooxygenations, the role of these enzymes in lipid peroxidation may be to provide two electrons for 02 reduction. The first electron is used for reduction of ADP--Fe 3÷ and subsequent addition of O2 to the perferryl radical (ADP--Fe3+-O2-), which then extracts an H atom from a polyunsaturated lipid (LH) giving rise to a free radical (LH.) that reacts with 02 yielding a peroxide free radical (LOO.). The second electron is then used to reduce LOO. to the lipid hydroperoxide (LOOH). In the latter capacity, reduced cytochrome P450 can be replaced by EDTA--Fe 2+ or by the superoxide radical as generated through redox cycling of a quinone such as menadione.
Keywords--Lipid peroxidation, Cytochrome P450, Cytochrome P450 reductase, ADP-Iron, Superoxide, Liposomes, Free radicals
INTRODUCTION
Initiation of microsomal lipid peroxidation by NADPH and ADP--Fe 3+, first described 25 years ago, 1-3 is generally believed to involve NADPH--cytochrome P450 reductase. 4 Evidence supporting this concept is twofold: 1) Antibodies against the reductase inhibit lipid peroxidationS; 2) Purified P450 reductase can in- itiate NADPH- and ADP--Fe 3 ÷-dependent lipid per- oxidation in liposomes prepared from microsomal phospholipids. 6 However, there is growing evidence that other factors may be involved in initiation reac- tions, as suggested by the findings that 1) Purified P450 reductase is a poor catalyst of ADP--Fe 3+ reduction by NADPHT'S; 2) In the liposomal system E D T A - Fe 3 +, in addition to ADP--Fe 3 +, is required for the initiation of lipid peroxidation. Cytochrome P450 can partially replace EDTA--Fe 3+, even with reductase preparation that is not capable of reducing cytochrome P450. 9,j°
The purpose of the present study was to attempt to identify any additional component(s) involved in mi-
*Address correspondence to: Alex Sevanian, Department of Bio- chemistry, Arrhenius Laboratory, University of Stockholm, S-106 91 Stockholm, Sweden. (Permanent address)
crosomal lipid peroxidation, using purified, recon- stitutively active reductase and cytochrome P450 in combination with liposomes prepared from vari- ous phospholipids. Another objective was to explain the enhanced rate of NADPH oxidation found in microsomes undergoing active lipid peroxidation, 1-3 a finding that is not accounted for by current schemes for the mechanism of microsomal lipid perox- idation. 1,2,~°-~2 An abstract summarizing some parts of this work has been published. ~3
MATERIALS AND METHODS
Lipid peroxidation was measured by following NADPH oxidation spectrophotometrically at 340 nm, 02 consumption polaro-graphically, and by TBAR for- mation as described by Ernster and Nordenbrand. 14 Li- posomes were prepared either from a 4:1 (mol:mol) mixture of bovine phosphotidyl choline (PC) and phos- photidyl ethanolamine (PE) or from total rat-liver phos- pholipids as described by Sevanian et a1.15 Liposomes were mixed with N A D P H - cytochrome P450 reduc- tase and/or cytochrome P450 at room temperature prior to addition to the reaction medium. Purified rat-liver
145
146 A. SEVAN]AN et al.
E
O E t-. v "ID ¢1 N .D "ID x O I o.. < z
10-
5-
/ ADP-Fe3+(100:I)
EDT~-FeZ/ TA_Fe3+(1.1:1)
/ / ADd~P-Fe3+(100:1)
ADP-Fe3+(20:1) !
10 20 30 iuM Fe 3+
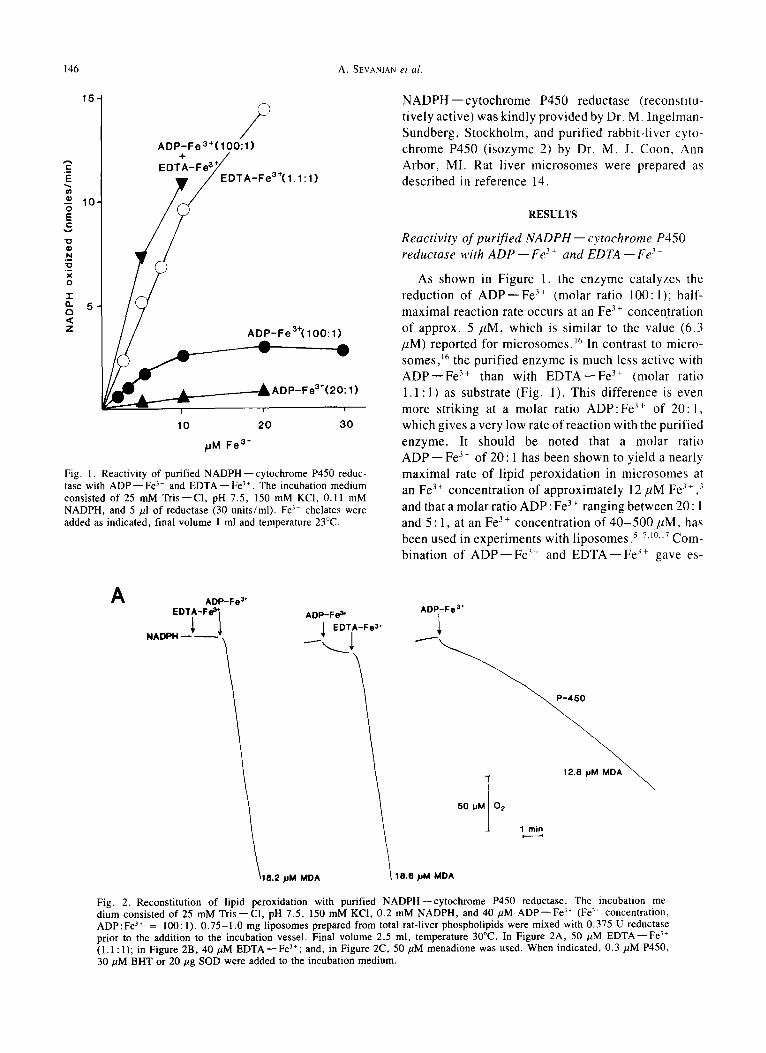
Fig. 1. Reactivity of purified NADPH--cytochrome P450 reduc- tase with A D P - - F e Z." and E D T A - - F e 3+. The incubation medium consisted of 25 mM Tris--C1, pH 7.5, 150 mM KCI, 0.11 mM NADPH, and 5/21 of reductase (30 units/ml). Fe 3÷ chelates were added as indicated, final volume 1 ml and temperature 23°C.
N A D P H - - c y t o c h r o m e P450 reductase (reconstitu- tively active) was kindly provided by Dr. M. Ingelman- Sundberg, Stockholm, and purified rabbit-liver cyto- chrome P450 (isozyme 2) by Dr. M. J. Coon, Ann Arbor, MI. Rat liver microsomes were prepared as described in reference 14.
RESULTS
R e a c t i v i t y o f p u r i f i e d N A D P H - - c y t o c h r o m e P450 r e d u c t a s e w i t h A D P - - F e 3 + a n d E D T A - - F e 3 +
As shown in Figure 1, the enzyme catalyzes the reduction of A D P - - F e 3+ (molar ratio 100: l); half- maximal reaction rate occurs at an Fe 3 + concentration of approx. 5 /tM, which is similar to the value (6.3 /~M) reported for microsomes. J6 In contrast to micro- somes, j6 the purified enzyme is much less active with A D P - - F e 3+ than with E D T A - - F e 3+ (molar ratio 1.1: 1) as substrate (Fig. 1). This difference is even more striking at a molar ratio A D P : F e 3+ of 20 :1 , which gives a very low rate of reaction with the purified enzyme. It should be noted that a molar ratio A D P - - Fe 3 + of 20: 1 has been shown to yield a nearly maximal rate of lipid peroxidation in microsomes at an Fe 3+ concentration of approximately 12/LM Fe 3÷ ,~ and that a molar ratio ADP: Fe 3 + ranging between 20 : 1 and 5 :1 , at an Fe 3+ concentration of 40 -500 /~M, has been used in experiments with l iposomes? 7,10.17 Com- bination of A D P - - Fe 3 + and E D T A - - Fe 3 + gave es-
A ADP-Fe 3" EDTff-F~ i ADP(FEe~DTA_Fe3"
NADPH . . . . . 1 " " ~ ' ~ ' ~
18.2 ~uM MDA / 18.6
ADP-Fe 3"
50 pM] 02 1 rain
pM MDA
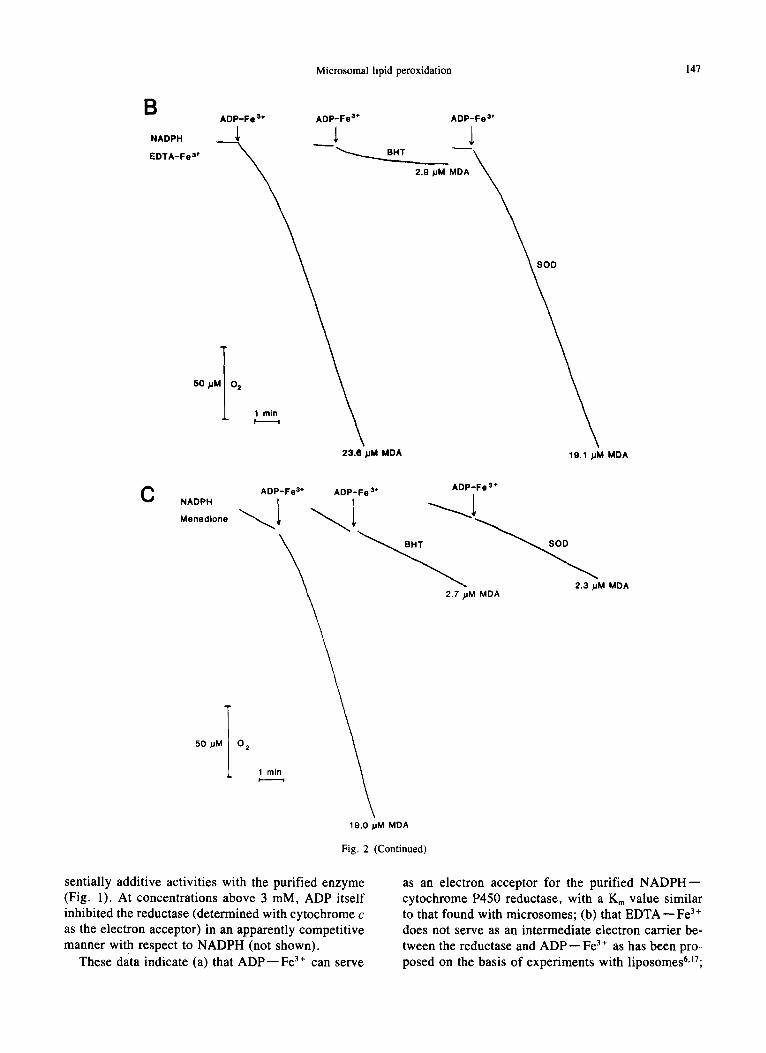
Fig. 2. Reconstitution of lipid peroxidation with purified NADPH--cytochrome P450 reductase. The incubation me- dium consisted of 25 mM Tris--C1, pH 7.5, 150 mM KCI, 0.2 mM NADPH, and 40/2M A D P - - F e 3+ (Fe Z+ concentration, ADP:Fe 3+ = 100: 1). 0 .75-1.0 mg liposomes prepared from total rat-liver phospholipids were mixed with 0.375 U reductase prior to the addition to the incubation vessel. Final volume 2.5 ml, temperature 30°C. In Figure 2A, 50/2M E D T A - - F e ~* (1.1 : 1); in Figure 2B, 40/2M EDTA--Fe3+; and, in Figure 2C, 50/2M menadione was used. When indicated, 0.3/2M P450, 30/2M BHT or 20/2g SOD were added to the incubation medium.
Microsomal lipid peroxidation 147
B ADP-Fe 3~" ADP-Fe a" ADP-Fe"*
EDTA_Fe3~ ~ ~ BHT - - ~ 2.9 jJM MDA
50 p
± ~ m,°, \
23.6 ~M MDA 19.1 pM MDA
C ADP_Fe3÷ ADP_Fe 3. ADP-Fe 34-
Menedlone M ~ M MDA
50 IJM I 0 2 ~ mi
19.0 uM MDA
Fig. 2 (Continued)
sentially additive activities with the purified enzyme (Fig. 1). At concentrations above 3 mM, ADP itself inhibited the reductase (determined with cytochrome c as the electron acceptor) in an apparently competitive manner with respect to NADPH (not shown).
These data indicate (a) that A D P - - F e 3÷ can serve
as an electron acceptor for the purified N A D P H - - cytochrome P450 reductase, with a K,~ value similar to that found with microsomes; (b) that E D T A - - F e 3+ does not serve as an intermediate electron carrier be- tween the reductase and A D P - - F e 3 ÷ as has been pro- posed on the basis of experiments with liposomes6,17;
148 A. SEVANIAN et al.
and (c) that the ADP:Fe 3+ ratio, as well as the net concentration of ADP, (the latter probably relative to the NADPH concentration) are critical parameters in determining the ability of A D P - - F e 3 + to serve as an electron acceptor for the reductase.
Reconstitution of lipid peroxidation with purified NADPH--cytochrome P450 reductase
02 consumption and TBAR formation (indicated as MDA) are observed with liposomes prepared from total liver phospholipids in the presence of P450 reductase, NADPH, A D P - - F e 3+ and E D T A - - F e 3+ (Fig. 2A). ADP - - Fe 3 + in the absence of EDTA - - Fe 3 + gave mea- surable 02 consumption and MDA formation, whereas E D T A - - F e 3+ in the absence of A D P - - F e 3+ gave none. In the ADP--Fe3+-supplemented system, E D T A - - F e 3+ could be partially replaced by cyto- chrome P450. These results are in essential agreement with those by Aust et al. 5'6'1°'17
The A D P - - Fe 3 ÷ and E D T A - - Fe 3 ÷-dependent 02 consumption and MDA formation were inhibited by the antioxidant butylated hydroxytoluene (BHT) but not by superoxide dismutase (SOD) (Fig. 2B). Men- adione (2-methyl-l ,4-naphthoquinone) is reduced by NADPH in microsomes, giving rise to an 02 con- sumption through "redox cycl ing" via N A D P H - - cytochrome P450 reductase with the formation of the autoxidable semiquinone ~8 but no TBAR formation. J2
In the case of liposomes, menadione gave an effect similar to that of E D T A - - F e 3+, promoting, in com- bination with A D P - - Fe 3 ~, O2 consumption and TBAR formation (Fig. 2C). This effect was abolished by both BHT and SOD.
These results show that both E D T A - - F e -~ and menadione are efficient in supporting lipid peroxida- tion in liposomes in the presence of NADPH and NADPH--cy toch rome P450 reductase; however, in the former case, the effect probably is due to EDTA- Fe 2 + (rather than to O j formed through autooxidation of the latter), whereas in the latter case, it is due to 02" itself, possibly by interaction with an endogenous iron component of the microsomes.
Effects o fADP Fe 3+ ED T A- -Fe 3+ cvtochrome P450 and liposomes on the rate of oxidation of NADPH by purified NADPH -- cytochrome P450 reductase
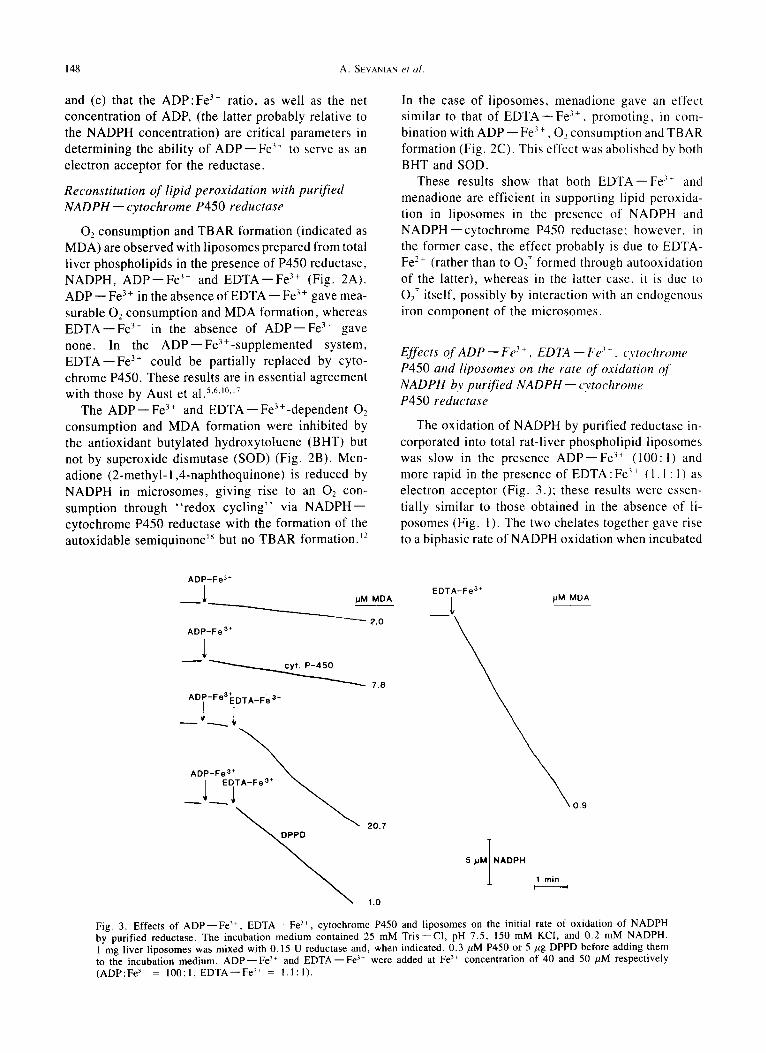
The oxidation of NADPH by purified reductase in- corporated into total rat-liver phospholipid liposomes was slow in the presence A D P - - F e 3+ (100:1) and more rapid in the presence of EDTA:Fe 3+ (1.1: 1) as electron acceptor (Fig. 3.); these results were essen- tially similar to those obtained in the absence of li- posomes (Fig. 1). The two chelates together gave rise to a biphasic rate of NADPH oxidation when incubated
ADP-Fe 3+
ADP-Fe 3+
Cyt. P-450
ADP-Fe3~DTA_Fe 3÷
_<_l\
pM MDA
2.0
7.8
20.7
1.0
EDTA-Fe 3÷ ~M MDA
0.9
/JMINADPHI 1 rain 5
I q
Fig. 3. Effects of A D P - - F e 3+, EDTA--Fe 3~, cytochrome P450 and liposomes on the initial rate of oxidation of NADPH by purified reductase. The incubation medium contained 25 mM Tr is - -Cl , pH 7.5, 150 mM KCI, and 0.2 mM NADPH. 1 mg liver liposomes was mixed with 0.15 U reductase and, when indicated, 0.3 ,uM P450 or 5 pg DPPD before adding them to the incubation medium. A D P - - F e 3+ and EDTA--Fe 3+ were added at Fe 3+ concentration of 40 and 50 pM respectively (ADP:Fe 3+ = 100:1, EDTA--Fe 3+ = 1.1:1).

Microsomal lipid peroxidation 149
in the presence of total liver phospholipid liposomes (Fig. 3) with a concomitant TBAR formation (indicated as MDA). These findings are consistent with the ob- servation (Figs. 2A and 2B) that lipid peroxidation is initiated by the two chelates together and ceases when 02 is exhausted, after which NADPH oxidation pro- ceeds at.a diminished rate through reduction of the Fe 3+ chelates. As expected, the antioxidant diphenyl- p-phenylenediamine (DPPD) abolished the initial rapid phase of NADPH oxidation. Just as in the experiment shown in Figure 2A, cytochrome P450 could partially replace EDTA--Fe 3+ in initiating lipid peroxidation in the reconstituted liposomal system.
The effects described above were not observed with bovine PC/PE liposomes. However, these liposomes did exhibit a high rate of lipid peroxidation in the pres- ence of ADP--Fe 3+ and ascorbate, provided that a critical concentration of deoxycholate (DOC) was added (Fig. 4). EDTA--Fe 3+ could not replace
8SC
Liposomes ] A D P - F e 3+ " ~ - ~ . . . . . no D O C
50 luM 02
1 min k I
lO . l , uM MDA
Fig. 4. Ascorbate- and ADP--Fe3+-induced lipid peroxidation in liposomes containing phosphatidyl choline and bovine phosphatidyl ethanolamine (molar ratio 4: 1). The incubation medium contained 25 mM Tris-- CI, pH 7.5, 50 mM KCI, 0.2 mM NADPH, and 2.5 mg liposomes in a final volume of 2.5 ml. When indicated, 1 mM ascorbate (asc.) and deoxycholate (DOC) in the final concentration indicated were added.
ADP--Fe 3+. In the total-liver phospholipid lipo- somes, DOC was not required for initiation of lipid peroxidation (not shown).
Stimulation of NADPH oxidation in liver microsomes by ADP- -Fe 3 +
In agreement with earlier findings, 1 ADP--Fe 3+ stimulated NADPH oxidation which was depressed by inhibiting lipid peroxidation with DPPD (Fig. 5), al- though the latter effect was less pronounced than earlier reported. Note that these previous studies were per- formed with an A D P " Fe 3 + chelate of a molar ratio of 20: 1. In accordance with unpublished observations by F. Ursini and M. Maiorino (personal communi- cation), the stimulation of NADPH oxidation by ADP--Fe 3+ varied with the ADP:Fe 3+ ratio as well as the amount of microsomes used. Obviously, these observations will require further study. In any case, the present data show that ADP--Fe 3+ can serve as an active electron acceptor for NADPH in microsomes.
DISCUSSION
The results confirm earlier conclusions (cf. Intro- duction) that microsomal lipid peroxidation involves a component (or components) in addition to P450 reductase, which is replaceable by E D T A - Fe 3 ÷ and,
ADP-Fe a, /
0.5 mg micros 1 0.18 mM NADPH
I ' 10 pM NADPH
1 rain J i ,uM MDA 8.9
\ DPPD \
\ \
\ \
\
Fig. 5. Stimulation of NADPH oxidation in liver microsomes by ADP--Fe 3+. The incubation medium contained 25 mM Tris--C1, pH 7.5, 150 mM KC1, 0.2 mM NADPH, and 0.5 mg microsomal protein. ADP--Fe 3+ was added at a concentration of 20/~M Fe 3~ (ADP:Fe 3+ = 100: 1). When indicated, the reaction medium con- tained 5 mg DPPD. Final volume, 1 ml. Temperature, 30°C.
150 A. SEVANIAN et al.
1,/2 NADP++I/2 H +
1/2 N A D P H AD P - F e 2 ~ 2 i2
2+ ADP-Fe s+ ADP-Fe -0 2
2 ~ ADP '/'F-eS+ O~ -
L LH
O 2 "X'x~~5 ~"~P R O PAGATI O N ~ _
LOO.
TERMINATION
LOOH - - -
1,/2 NADPH
>MDA
1/2 NADP++I,/2 H +
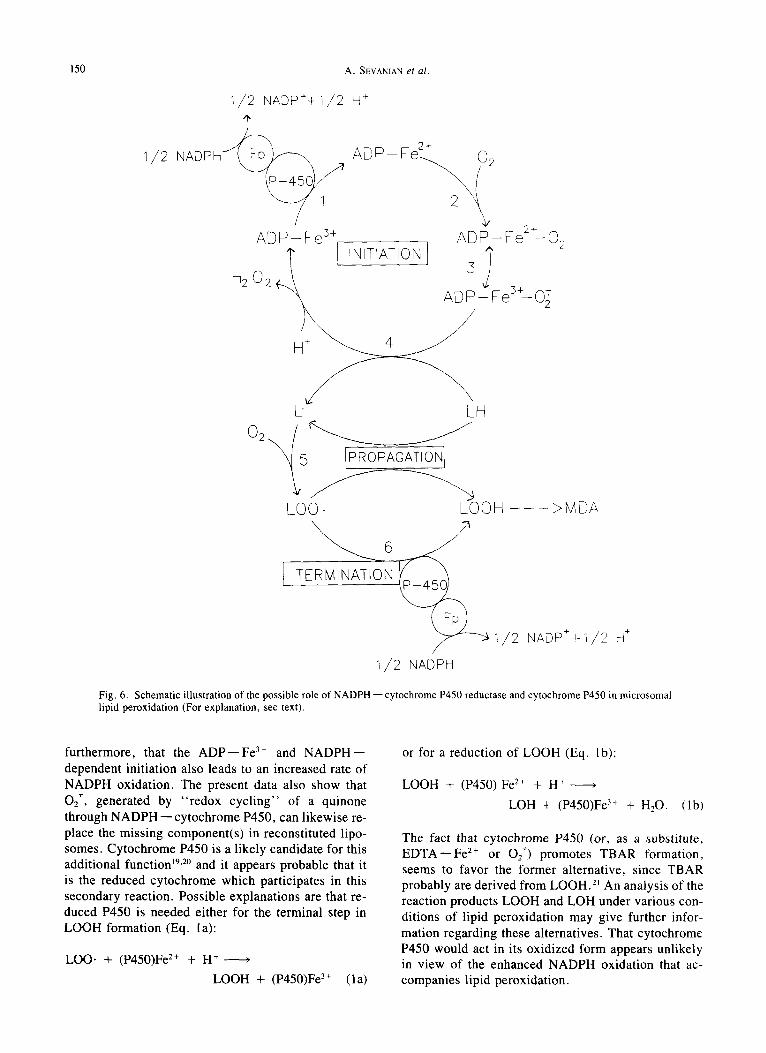
Fig. 6. Schematic illustration of the possible role of NADPH - - cytochrome P450 reductase and cytochrome P450 in microsomal lipid peroxidation (For explanation, see text).
furthermore, that the A D P - - F e 3+ and N A D P H - - dependent initiation also leads to an increased rate of NADPH oxidation. The present data also show that 027, generated by "redox cycl ing" of a quinone through N A D P H - - cytochrome P450, can likewise re- place the missing component(s) in reconstituted lipo- somes. Cytochrome P450 is a likely candidate for this additional function ~ga° and it appears probable that it is the reduced cytochrome which participates in this secondary reaction. Possible explanations are that re- duced P450 is needed either for the terminal step in LOOH formation (Eq. la):
LOO. + (P450)Fe 2+ + H +
LOOH + (P450)Fe 3 + (la)
or for a reduction of LOOH (Eq. lb):
LOOH + (P450)Fe 2+ + H +
LOH + (P450)Fe 3+ + H 2 0 . (lb)
The fact that cytocbrome P450 (or, as a substitute, E D T A - - F e 2+ or O j ) promotes TBAR formation, seems to favor the former alternative, since TBAR probably are derived from LOOH. 21 An analysis of the reaction products LOOH and LOH under various con- ditions of lipid peroxidation may give further infor- mation regarding these alternatives. That cytochrome P450 would act in its oxidized form appears unlikely in view of the enhanced NADPH oxidation that ac- companies lipid peroxidation.

Microsomal lipid peroxidation 151
Figure 6 summarizes in a schematic form our current and tentative view on the possible role of cytochrome P450 in microsomal lipid peroxidation. Just as in the case of mixed-function oxidation, cytochrome P450, in conjunction with NADPH--cytochrome P450 re- ductase, may catalyze the reduction of a molecule of 02 by two electrons.22 In the case of lipid peroxidation, one of the electrons may be required to reduce ADP--Fe 3+ (reaction 1) which, in turn, gives rise to a perferryl radical (reactions 2 and 3) which is widely postulated to be capable of converting LH into L. (re- action 4)2'3A0'12; the latter reacts with 02 to form LOO. (reaction 5).
The second electron can be utilized in either one of two possible ways: (1) For the reduction of LOO. to LOOH (reaction 6 and Eq. la) and in this capacity, cytochrome P450 may be replaced by EDTA--Fe 2+ or by O2-; (2) For reductive decomposition of LOOH (Eq. 2a) which derives from propagation reactions in- volving LOO. and LH, producing new radical chain reactions (Eq. 2b). The latter possibility follows from the well known radical chain reactions described for lipid peroxides in the presence of transition metals (Eqs. 3a and 3b).
LOOH + P450--Fe 2+ )
LO" + P450---Fe 3+ (2a)
LOO. + LH ~ LOOH + L. (2a)
L- + 02 ~ LOO. (3a)
LO. + LH ~ LOH + L. (3b)
P450---Fe 3÷ + 1/2 NADPH ~-
P450---Fe 2÷ + 1/2 NADP ÷ + 1/2 H ÷ (4)
In this manner, cytochrome P450, in conjunction with NADPH -- cytochrome P450 reductase, catalyzes the decomposition of LOOH (reaction 4 and Eq. 2a) producing new lipid radicals capable of sustaining lipid peroxidation and promoting the accumulation of TBAR from LOOH decomposition. The involvement of the second electron, thereby, promotes further utilization of NADPH with concomitant O2 consumption and in- creased lipid peroxidation.
The data with PC/PE liposomes (Fig. 5) show that, in addition to the catalytic components involved in lipid peroxidation, the process is also dependent on the phospholipid composition and/or the physical state of the membrane including the net charge of the phos-
pholipids. These relationships require further inves- tigation.
Acknowledgements--This work has been supported by grants from the National Institutes of Health, USA (ES03816), the Swedish Nat- ural-Science Research Council, and Pharmacia AB, Uppsala. We acknowledge the valuable participation of Mr. R. Kammula in the initial phase of this investigation.
REFERENCES
1. Hochstein, P.; Ernster, L. ADP-activated lipid peroxidation cou- pled to the TPNH oxidase system of microsomes. Biochim. Biophys. Res. Commun. 12:388-394; 1963.
2. Hochstein, E: Ernster, L. Microsomal peroxidation of lipids and its possible role in cellular injury. Ciba Foundation Sym- posium on Cell Injury, Churchill, London; 1964:123-134.
3. Hochstein, E; Nordenbrand, K.; Ernster, L. Evidence for the involvement of iron in the ADP-activated peroxidation of lipids in microsomes and mitochondria. Biochem. Biophys. Res. Com- mun. 14:323-328, 1964.
4. Aust, S. D.; Morehouse, L. A.; Thomas, C. E. Role of metals in oxygen radical reactions. J. Free Rad. Med. 1:3-25; 1985.
5. Pedersen, T. C.; Buege, J. A.; Aust, S. D. Microsomal electron transport. The role of reduced nicotinamide adenine dinucleotide phosphate-cytochrome c reductase in liver microsomal lipid per- oxidation. J. Biol. Chem. 248:7134-7141; 1973.
6. Pedersen, T. C.; Aust, S. D. The mechanism of liver microsomal lipid peroxidation. Biochim. Biophys. Acta 385:232-241; 1975.
7. Buchler, J. R.; Tien, M.; Aust, S. D. The requirement for ferric iron in the initiation of lipid peroxidation by chelated ferrous iron. Biochem. Biophys. Res. Commun. 111:777-784; 1983.
8. Vegh, M.; Morton, A.; Horvath, I. Reduction of Fe(III) ADP complex by liver microsomes. Biophys. Acta 964:146-150.
9. Svingen, B. A.; Buege, J. A.; O'Neal, E O.; Aust, S. D. The mechanism of NADPH-dependent lipid peroxidation. J. Biol. Chem. 254:5892-5899; 1979.
10. Tien, M.; Aust, S. D. Comparative aspects of several model lipid peroxidation systems. In: Yagi, K., ed. Lipid peroxides in biology and medicine. New York: Academic Press; 1982:23- 39.
l l . Ernster, L.; Nordenbrand, K.; Orrenius, S. Microsomal lipid peroxidation: Mechanism and some biomedical implications. In: Yagi, K., ed. Lipid peroxides in biology" and medicine. New York: Academic Press; 1982:55-79.
12. Ernster, L.; Lind, C.; Nordenbrand, K.; Thor, H.; Orrenius, S. NADPH-cytochrome P450 reductase as an oxygen-radical gen- erator. In: Nozaki, M.; Yamamoto, S.; Ishimura, Y.; Coon, M. J.; Ernster, L.; Estabrook, R. W.; eds. Oxygenases and oxygen metabolism. New York: Academic Press; 1982:357-370.
13. Kammula, R.; Nordenbrand, K.; Sevanian, A.; Hochstein, P.; Ernster, L. Microsomal lipid peroxidation: the role of NADPH - - cytochrome P-450 reductase and cytochrome P-450. J. Cell Biol. 107:406a; 1988.
14. Ernster, L.; Nordenbrand, K. Microsomal lipid peroxidation. Meth. Enzymol. 10:574-580; 1967.
15. Sevanian, A.; Wratten, M. L.; McLeod, L, L.; Kim, E. Lipid peroxidation and phospholipase A 2 activity in liposomes com- posed of unsaturated phospholipids: a structural basis for en- zyme activation. Biochim. Biophys. Acta. 916:316-327.
16. Ursini, F.; Maiorino, M.; Hochstein, P.; Ernster, L. Lipid per- oxidation: Mechanisms of initiation. The role of iron and iron chelators. Free Rad. Biol. Med. 6:31-36; 1989.
17. Morehouse, L. A.; Aust, S. D. Reconstituted microsomal lipid peroxidation. A D P - Fe 3+-dependent peroxidation of phospho- lipid vesicles containing NADPH - - cytochrome P450 reductase and cytochrome P450. Free Radical Biol. Med. 4:269-277; 1988.
18. Lind, C.; Hochstein, E; Ernster, L. DT-diaphorase as a quinone reductase: a cellular control device against semiquinone and

152 A. SEVANIAN et al.
superoxide radical formation. Arch. Biochem. Biophys. 216:178-185; 1982.
19. Ekstrom, G.; Ingelman-Sundberg, M. Cytochrome P-450 de- pendent lipid peroxidation in reconstituted membrane vesicles. Biochem. Pharmacol. 33:2521-2523; 1984.
20. Coon, M. J.; Vaz, A. D. N. Mechanism of microsomal electron transfer reactions: Role of cytochrome P-450. Chemica Scripta 27A: 17-19, 1987.
21. Terao, J. and Matsushita, S. Thiobarbituric acid reaction of methyl archadonate monohydroperoxide isomers. Lipids 16:98- 101; 1981.
22. White, R. E.; Coon, M. J. Oxygen activation by cytochrome P-450. Annu. Rev. Biochem. 49:315-356: 1980.
ABBREVIATIONS
BHT--butylated hydroxytoluene
DOC--deoxycholate
DPPD--diphenyl-p-phenylene diamine
MDA--malondialdehyde
PC--phosphatidyl choline
PE--phosphatidyl ethanolamine
SOD--superoxide dismutase
TBAR--thiobarbituric acid reacting product~





























