Embed Size (px)
Citation preview

J . Zool. Syst. Evol. Research 33, 97-100 (1995) 0 1995 Blackwell Wissenschafts-Verlag, Berlin ISSN 0044-3808
Received on 6 March 1995
Mitochondrial DNA restriction-fragment-length monomorphism in the Italian wolf (Canis lupus) population
E. RANDI, F. FRANCISCI and V. LUCCHINI
Abstract Mitochondrial-DNA (mtDNA) restriction patterns were studied in 22 wolves (Canis lupus) sampled in central-northern Italy. A total of 60 restriction sites were detected, encompassing about 2 % of the mitochondria1 genome of canids. All wolves showed the same restriction pattern. Therefore, a single mtDNA haplotype was detected in the Italian wolf population. Historical information on peninsular isolation and demographic decline suggest that low genetically effective population size and random drift may have strongly reduced the mtDNA variability of wolves in Italy over the last 100-200 years. A different mtDNA restriction pattern in feral dogs sampled from a wolf range in central Italy was detected. These findings suggest that the hybridization and introgression of female dog genomes into the Italian wolf population may be rare or absent.
Key words: Mitochondria1 DNA - RFLP - Canis lupus - Dog - Population genetics - Hybridization - Conservation genetics
Introduction Wolves were largely exterminated in western Europe over the last 2 centuries but isolated remnants have survived in Iberia and Italy (Delibes 1990; Promberger and Schroder 1993). According to Cagnolaro et al. (1974), wolves in Italy have been confined south of the Po River for the last 100 years, following their eradication in the Alps and adjacent regions throughout the 1800s. Occasionally, some have dispersed into the southern and northern Alps, and some kills were recorded up to 1987 (Cagnolaro et al. 1974; Delibes 1990), but this has had little effect on the declining population trend. Therefore, gene flow across the Alps could have been increasingly restricted, and finally completely interrupted, over the last 2 centuries.
The recent dynamics of the Italian wolf population have been only partially reconstructed because the trends cannot easily be estimated from the incomplete and inaccurate his- torical records. Nationwide surveys of wolves in Italy in 1970- 1973 provided different interpretations of distribution, ranging patterns, and population size (Cagnolaro et al. 1974; Zimen and Boitani 1975). Subsequent updates of living wolf popu- lations were based on broad extrapolations of small-scale, infrequent winter-tracks or howling surveys (Boitani 1992), and did not correlate with available kill figures and specimens (Francisci and Guberti 1993). Qualitative information indi- cates that the decline has continued since the end of the 1960s, and has been followed by a recent recovery by wolves in peninsular Italy. Wolf distribution throughout the Apennines and branching ranges shrank and re-expanded in 1950-1980: areas in northern Italy, seemingly deserted in 195G1970, have since been recolonized, probably following the increase in available populations of prey species; evidence for wolves increasing where their populations had previously been per- sistent, i.e. in central and southern latitudes, is still scanty. The minimum wolf population size should have not been lower than 100 (Zimen and Boitani 1975), and has recently been estimated to have increased to 300-400 in Italy (Boitani et al. 1989). Analyses of kill figures refemng to 1960-1992 (Cagnolaro et al. 1974; Francisci and Guberti 1993), however, suggest that a population size of 300 wolves in Italy may well
Fig. 1. Approximate distribution range of wolves in Italy
have been in existence for several decades. The current Italian wolf population is actively dispersing northward along the Apennine divide, and a few individuals have already reached the southern borders of the French Alps (Fig. 1). Expansion of the Italian wolf population could have been helped by the full legal protection accorded to the species since 1975, and by increasing numbers of ungulate prey species, particularly wild boar (Sus scrofa), in the Apennines (Francisci and Gub- erti 1993). Large numbers of free-ranging dogs are thought to compete with wolves (Boitani 1992), or to have indirectly favoured their demographic recovery by crossbreeding (Boit- ani 1984).
The recent demographic history of the Italian wolves may have produced, predictably, genetic consequences at the population level. Population genetic structure may have been shaped by isolation, demographic fluctuations, and by cross- breeding with feral dogs. In this paper, mtDNA sequence variability in the Italian wolf population is analysed so as to
98 RANDI, FRANCISCI and LUCCHINI
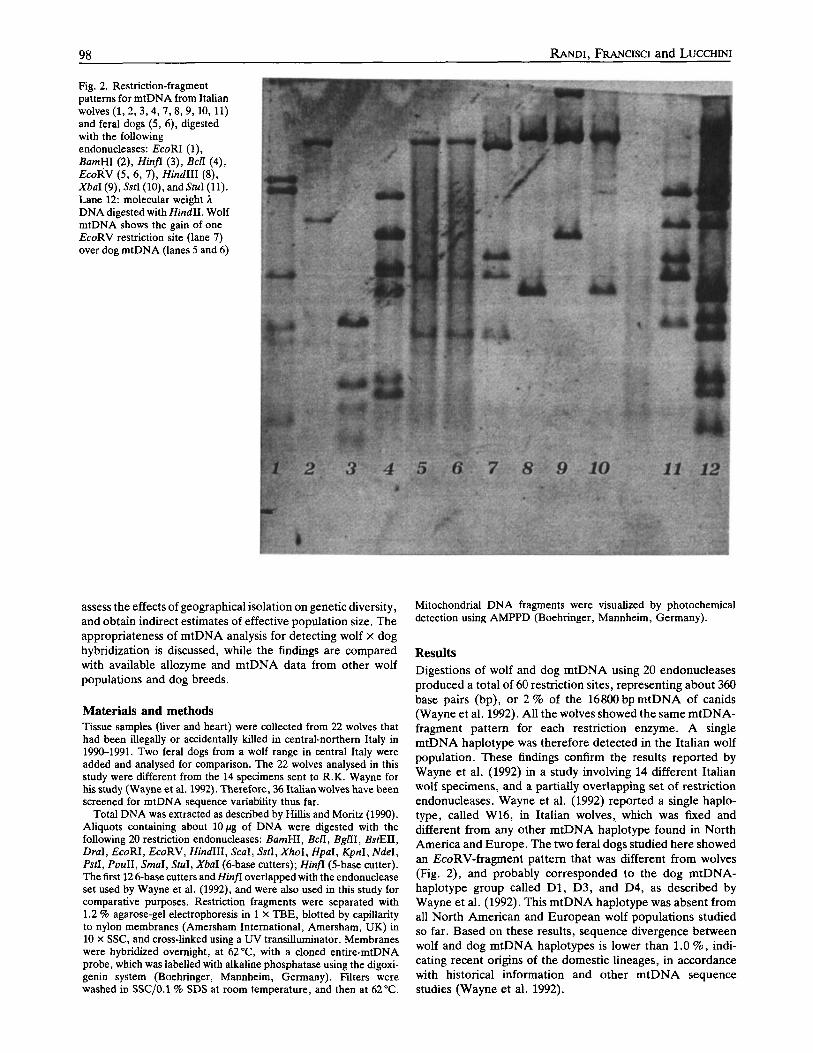
Fig. 2. Restriction-fragment patterns for mtDNA from Italian wolves (1, ? ,3 ,4 ,7 ,8 ,9 ,10, 11) and feral dogs (5, 6), digested with the following endonucleases: EcoRI (l) , BamHI ( 2 ) , Hinfr (3), BclI (4), EcoRV (5, 6, 7), HindIII (8), XbuI(9),SstI(lO),andSruI(ll). Lane 12: molecular weight A DNA digested with HindII. Wolf mtDNA shows the gain of one EcoRV restriction site (lane 7) over dog mtDNA (lanes 5 and 6)
assess the effects of geographical isolation on genetic diversity, and obtain indirect estimates of effective population size. The appropriateness of mtDNA analysis for detecting wolf x dog hybridization is discussed, while the findings are compared with available allozyme and mtDNA data from other wolf populations and dog breeds.
Materials and methods Tissue samples (liver and heart) were collected from 22 wolves that had been illegally or accidentally killed in central-northern Italy in 1990-1991. Two feral dogs from a wolf range in central Italy were added and analysed for comparison. The 22 wolves analysed in this study were different from the 14 specimens sent to R.K. Wayne for his study (Wayne et al. 1992). Therefore, 36 Italian wolves have been screened for mtDNA sequence variability thus far.
Total DNA was extracted as described by Hillis and Moritz (1990). Aliquots containing about 10% of DNA were digested with the following 20 restriction endonucleases: BamHI, Bcn, BglII, BstEII, DraI, EcoRI, EcoRV, HindIII, ScaI, SsrI, XhoI, HpaI, KpnI, NdeI, PstI, PuuII, SmaI, SruI, XbaI (6-base cutters); Hinfr (5-base cutter). The first 12 6-base cutters andHinfr overlapped with the endonuclease set used by Wayne et al. (1992), and were also used in this study for comparative purposes. Restriction fragments were separated with 1.2 % agarose-gel electrophoresis in 1 X TBE, blotted by capillarity to nylon membranes (Amersham International, Amersham, UK) in 10 X SSC, and cross-linked using a UV transilluminator. Membranes were hybridized overnight, at 6 2 T , with a cloned entire-mtDNA probe, which was labelled with alkaline phosphatase using the digoxi- genin system (Boehringer, Mannheim, Germany). Filters were washed in SSC/O.l % SDS at room temperature, and then at 62 "C.
Mitochondrial DNA fragments were visualized by photochemical detection using AMPPD (Boehringer, Mannheim, Germany).
Results Digestions of wolf and dog mtDNA using 20 endonucleases produced a total of 60 restriction sites, representing about 360 base pairs (bp), or 2 % of the 16800 bp mtDNA of canids (Wayne et al. 1992). All the wolves showed the same mtDNA- fragment pattern for each restriction enzyme. A single mtDNA haplotype was therefore detected in the Italian wolf population. These findings confirm the results reported by Wayne et al. (1992) in a study involving 14 different Italian wolf specimens, and a partially overlapping set of restriction endonucleases. Wayne et al. (1992) reported a single haplo- type, called W16, in Italian wolves, which was fixed and different from any other mtDNA haplotype found in North America and Europe. The two feral dogs studied here showed an EcoRV-fragment pattern that was different from wolves (Fig. 2), and probably corresponded to the dog mtDNA- haplotype group called D1, D3, and D4, as described by Wayne et al. (1992). This mtDNA haplotype was absent from all North American and European wolf populations studied so far. Based on these results, sequence divergence between wolf and dog mtDNA haplotypes is lower than 1.0 %, indi- cating recent origins of the domestic lineages, in accordance with historical information and other mtDNA sequence studies (Wayne et al. 1992).

![Mitochondrial genomes of African pangolins and …...Spotted Hyena Crocuta crocuta JF894378.1 [78] Grey Wolf Canis lupus KU696410.1 [79] Mitochondrial genomes of African pangolins](https://img.pdfslide.net/doc/110x75/5e70e1c5aed16c1ea26f8fce/mitochondrial-genomes-of-african-pangolins-and-spotted-hyena-crocuta-crocuta.jpg)













![Lobo [canis lupus]](https://img.pdfslide.net/doc/110x75/588270221a28ab470c8b6969/lobo-canis-lupus.jpg)