Embed Size (px)
Citation preview

REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 32, No. 1 DE 2003
MODIFICACIÓN D E L E S Q U E L E T O P E P T I D I C O D E F R A G M E N T O S D E LA P R O T E Í N A M S P - 1 D E L
P l a s m o d i u m f a l c i p a r u m : SÍNTESIS EN FASE SÓLIDA D E P S E U D O P É P T I D O S AMIDA R E D U C I D A
B A C K B O N E M O D I F I C A T I O N O F P l a s m o d i u m f a l c i p a r u m ' s M S P - 1 P E P T I D E F R A G M E N T S : S O L I D
P H A S E SYNTHESIS O F R E D U C E D AMIDE P S E U D O P E P T I D E S
José Manuel Lozano'*, Luz Man,' Solazar*, Carlos Parra*. Nubia Barrera*, Fanny Guzmán*, José Libardo Torres-Castellanos** y Manuel Elkin Palarroyo*, ***
Recibido: 17/02/02 - Aceptado: 30/06/03
RESUMEN
Los estadios asexuales sanguíneos del Plasmodium fakiparum constituyen un blanco natural para el desarrollo de vacunas antimaláricas debido a la gran cantidad de antígenos expresados en su superficie que están implicados en el proceso de invasión del parásito a glóbulos rojos, desencadenando la enfermedad en el hospedero humano. Uno de estos antígenos es la proteína de superficie del merozoito MSP-I, una glicopro-teína de 195 kDa en su forma madura. Recientemente se identificaron los pép-tidos de esta proteína con alta capacidad de unión a glóbulos rojos (GR). Entre todos ellos, el péptido sintético codificado 1585, cuya estructura primaria
nece al amino terminal del fragmento de 42 kDa de la proteína MSP-I, mostró propiedades de unión específica a GR. Posteriormente, se identificó el motivo de unión cuya secuencia de aminoácidos es LKP-A. Con el objeto de evaluar el papel del enlace peptídico en la modulación estructural del esqueleto peptídico se diseñaron pseudopéptidos amida reducida, mediante la sustitución de enlaces peptí-dicos normales de la forma [CO-NH] del motivo de unión a GR por el enlace Í̂CHiNH]. De esta manera se obtuvieron
formas monoméricas y poliméricas pseu-dopeptídicas del 1585. En este estudio reportamos algunas de las propiedades físicas y químicas de los pseudopéptidos análogos del 1585.
-E VL YLKPLAG V Y R S L K K Q L E ' perte-
Autor corresponsaL Correo electrónico: [email protected] /[email protected] Fundación Instituto de Inmunología de Colombia (FIDIC). Carrera 50 No. 26-00. Bogotá. Colombia. Departamento de Química. Facultad de Ciencias. Universidad Nacional de Colombia. Departamento de Patología, Facultad de Medicina. Universidad Nacional de Colombia.

REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 32, No. 1 DE 2003
Palabras clave: enlace peptídico, esqueleto peptídico, pseudopéptido, estructura tridimensional
ABSTRACT
Due to the number of surface antigens expressed in Plasmodium fakiparum asexual stages which are associated to red blood cell (RBC) invasión and so causing human malaria infection, these erythrocyte parasite forms are thus, a natural target for malarial vaccine development. One of such antigens is the Merozoite Surface Protein-1 (MSP-I).
This is a 195 kDa glycoprotein in its matute expressed form. Recently, MSP-1 high binding capacity peptides to RBC were identified. Amongst all of them, the synthetic so called 1585 pep-tide, whose primary structure '-''-EVLYLKPLAGVYRSLKKQLE'^"' belongs to the MSPi's 42 kDa fragment amino termini, shown specific high binding ca
pacity to RBCS. Thereafter, the 1585 peptide LKP-A high binding motif was identified. To assess the role of such binding niotif peptide bonds respect to back-bone structure modulation, a set of reduced amide pseudopeptides were designed and synthesised. Thus, a single natural [CO-NH] peptide bond was replaced by a T[CH2-NH]. isoster bond. Then, 1585 pseudopeptide monomer as well as poly-meric forms were obtained. In this work we report some physichal and chemical properties of such 1585 surrogates.
Key words: Peptide bond, peptide back-bone, pseudopeptide, tridimensional structure.
INTRODUCCIÓN
El Plasmodium falciparum, agente causal de la mayoría de los casos de malaria en el mundo, tiene un ciclo de vida complejo, el cual involucra dos hospederos:
CONSERVADO SEMICONSERV.\DO
Procesamiento Primario 83 kDa
Precursor MSPl
28-30kDa 38 kDa
Procesamiento secundario
42 kDa
y , ' 33kDa^v 19kDa
Péptido 1585 '=«=EVLVLKPLAGVYRSLKKQLE'"
Figura 1. Organización estructural de la proteína MSP-1 del Plasmodiumfalciparum y localización del péptido 1585 (MSP-1 1282-1301

REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 32, No. 1 DE 2003
la hembra del mosquito Anopheles y un ser humano susceptible de contraer la enfermedad. Los estadios asexuales sanguíneos del parásito constituyen un blanco natural para el desarrollo de vacunas antimaláricas debido a la gran cantidad de antígenos expresados en su superficie, y que a su vez estén implicados en el proceso de invasión del parásito a glóbulos rojos desencadenando la enfermedad en el hospedero humano.
Uno de estos antígenos es la proteína de superficie del merozoito-1 MSP-i, una glicoproteína de 195 kDa, cuya forma precursora sufre un procesamiento pro-teolítico primario hacia fragmentos de 83 kDa, 30 kDa, 38 kDa y 42 kDa, un segundo procesamiento proteolítico del fragmento de 42 kDa que libera un fragmento de 33 kDa, y un fragmento de 19 kDa que permanece unido a los merozoi-
tos (1) como se observa en la figura 1. Los fragmentos liberados en los dos procesos permanecen asociados de manera no covalente sobre la superficie de los merozoitos durante el proceso de invasión a sus células blanco. Recientemente se identificaron los péptidos de la MSP-I con alta capacidad de unión a glóbulos rojos (GR). Entre todos ellos, el péptido sintético codificado 1585, cuya estructura primaria EVLYLKPLAGVYRSLKKQLE pertenece al amino terminal del fragmento de 42 kDa, mostró propiedades de unión específica a GR y posteriormente se idenfiticó el motivo de unión del 1585 cuya secuencia de aminoácidos es LKP-A(2).
El objetivo en el diseño de pseudopéptidos es el enlace peptídico. Por inversión del enlace peptídico y manteniendo la quiralidad del carbono a se
Péptido natural L (enlace peptídico)
[CO-NH]
Pseudopéptidos Retro y Retroinverso
JL. I H
A_X„. 4'-[NH-C0]
Pseudopéptido amida reducida
0 R"
H
1 0
1 R H
H'-ÍCHjNH]
Figura 2. Algunas formas isostéricas del enlace peptídico [CO-NH].

REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 32, No 1 DE 2003
obtiene un pseudopéptido Retroinverso, si se invierte la quiralidad se obtendrá un pseudopépddo Retro. Al sustituir el átomo de oxígeno del carbono carbonílico del enlace peptídico por dos átomos de hidrógeno se obtiene un pseudopéptido amida reducida (parte inferior) (3) como se puede observar en la figura 2.
Con el objeto de evaluar el papel del enlace peptídico en la modulación estruc-uiral del esqueleto peptídico se diseñaron pseudopéptidos amida reducida, mediante la sustitución de enlaces peptídicos normales de la forma [CO-NH] del motivo de unión a GR por el enlace 4^[CH:-NH]. como se observa en la Figura 3. De esta manera se obtuvieron formas monoméricas y poliméricas pseudopeptídicas del 1585 (MSP-i'-**-'""), las primeras se emplearon para estudio de sus propiedades químicas y físicas, y las fonuas poliméricas para estudios biológicos y bioquímicos. En el presente estudio reportamos algunas de las propiedades químicas y físicas de los pseudopéptidos análogos del 1585.
En la figura 3 se observa la posición de los enlaces peptídicos normales del motivo de unión al glóbulo rojo (GR) del péptido 1585 de la proteína MSP-i del P. falciparum a ser sustituidos por enlaces isostéricos del tipo amida reducida a partir de su estrucaira primaria. Las flechas destacan desde el N hacia c terminal el enlace entre Ki:87-Pi288 para originar el
'-«E\ L\-L-K-P-L-A-GVVRSLKKQLE'""
Pse- 9475 ) 1'-|CH,NH|
Figura 3. Selección de enlaces peptídicos a modificar en el péptido 1585 (MSP-1 •-^- ' '^"") .
Pse-9473, y el enlace entre PI28«-LI:!(M para originar el Pse-9475. Ambos pseudopéptidos amida reducida se sintetizaron en forma de monómero y polímero.
MATERIALES Y MÉTODOS
Síntesis de pseudopéptidos amida reducida y determinación estructural
Derivados N-metoxi-N-metil-a- (t-Boc-amino)-carboxamidas y a-t-Boc-Aminoaldehídos
Para la obtención de cuatro pseudopéptidos amida reducida ^-[CHJ-NH], dos en forma monomérica y dos en forma poli-mérica, se prepararon los a-iV-í-Boc- ami-noaldehídos derivados de los aminoácidos comerciales protegidos a-.\'-r-Boc en dos etapas, empleando para ello estrategias en fase líquida tal como está descrito en la literatura (4). Inicialmente los a-A'-/-Boc-aminoácidos se transformaron a sus correspondientes A',(?-dime-tilcarboxamidas, los cuales se trataron posteriormente con LÍAIH4 para producir su correspondiente aldehido. Cada producto ínter- medio, así como cada derivado, se caracterizó por RMN-IH y "c . En ambos casos se muestra la caracterización de los dos derivados obtenidos para los residuos de Lisina (Lys) y Prolina (Pro), incluido el análisis por espectroscopia infrarroja (IR) para los productos finales.
Caracterización de los derivados N-metoxi-N-metil-a-(t-Boc-amino)-car-boxamidas
A^-^Boc-Lys-N-(OCH3)CH3: rendimiento: 98%; Rf. 0,65 (acetato de eti-

REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 32, No. 1 DE 2003
lo/hexano 2:1). RMN-'H, 8 (ppm) (CDCLJ/
TMSint): 1,45 (s,9 H, C(CH3)3), 1,5 (d, 2 H, Y-H) 1,6 (d, 2 H, 5-H), 1,8 (d, 2 H, P-H), 3.10 (d, 2 H, e-H), 3,25 (s, 3 H, N-CH3), 3,80 (s, 3 H, OCH3), 4.41 (dd, 1 H, aH), 8,01 (s, 1 H, NH), 7,14-7,30 (m, 5 H aromáticos del grupo protector cl-z). Aspecto oleoso, incoloro, inodoro.
Af-f-Boc-Pro-N-(OCH3) CH3: rendimiento: 97,5%; Rf. 0,55 (acetato de eti-lo/hexano 2:1). RMN-'H, 5 (ppm) (CDCLs/TMSint): 1,45 (S, 9 H, C(CH3)3), 2,0 (d, 2 H, y-H), 2,12 (s, 1 H, p- H), 3,25 (s, 3 H, N-CH3), 3,7 (d, 2 H, 5-H), 3,80 (s, 3 H, 0CH3), 4,41 (dd, 1 H, aH). Aspecto oleoso, tenuemente amarillo, inodoro.
Caracterización de los productos aldehido obtenidos
N-f-Boc-Lys-H: rendimiento 75,4%; Rf. 0,46 (acetato de etilo/hexano 2:1). RMN-'H, 5 (ppm) (CDCL3/ TMsint): 1,45 (s, 9 H, C(CH3)3), 1,5 (d, 2 H, Y-H) 1,6 (d, 2 H, 6-H), 1,8 (d, 2 H, p-H), 3.10 (d, 2 H, e-H), 4,41 (dd, 1 H, a H), 8,01 (s, 1 H. NH), 7,14-7,30 (m, 5 H, aromáticos del grupo protector CL-Z), 9,61 (s, 1 H CHO). RMN-"C (CDCI3/TMSint): 8= 200 ppm (c-1, CHO). Producto oleoso, de color levemente amarillo, de olor característico.
I.R. (película): v (cm') = 2934,16, 1694,16, 1522,52, 1366,32, 1248,68, 1164,79, 1041,37,753,07.
iV-f-Boc-Pro-H: rendimiento 78,6%; Rf. 0,5 (acetato de etilo/hexano 2:1). RMN-'H, 8 (ppm) (CDCI3/ TMSint): 1,45 (s, 9 H, C(CH3)3), 2,0 (d, 2 H, Y- H), 2,12 (s, 1 H, P-H), 3,7 (d, 2 H, 8-H), 4,41 (dd.
1 H, aH), 9,64 (S,l H CHO). RMN-'̂ c(CDCl3/TMSint): 8= 200 ppm (c-1, CHO). Producto oleoso, de color rojizo, de olor característico.
I.R. (película): v (cm-1) = 2973,70, 1694,16, 1391,39, 1365,35, 1161,90, 1120,44, 997,98, 918,91, 773,32.
Síntesis en fase sólida de los análogos amida reducida
Los péptidos se sintetizaron empleando las metodologías estándar para la síntesis de péptidos en fase sólida mediante la estrategia í-Boc (5, 6) y los pseudopéptidos del 1585 mediante modificaciones reportadas previamente (4). La pureza de cada molécula se analizó mediante cromatografía líquida de alta eficiencia HPLC-FR
provista de una columna de 5fim y 90Á de porosidad con un gradiente lineal de 0-70% AcN durante 30 min, con un flujo de elución de 1,0 mi por min. Las cromatografías preparativas se realizaron empleando una columna RPCI8 de S^m y 300A de porosidad con un gradiente lineal de 0-100% de AcN durante 45 min con un flujo de elución de 4,5 mi por min y detección a 210 nm.
Análisis de espectrometría de masas MALDI-TOF
Para identificar cada pseudopéptido se registraron espectros de masas empleando un equipo Bruker Protein TOF espectrofotómetro (Billerica, MA. USA). LOS experimentos MALDi ("Matrix Assisted Láser Desorptionlonization") se realizaron usando la técnica TOF ("Time of Flight"), empleando las matrices y los procedimientos estándar.

REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 32, No. 1 DE 2003
Espectroscopia RMN bidimensional
Se emplearon 10 mg del liofilizado de cada pseudopéptido purificado, disueltos en 500ml de una solución acuosa que contenía TFED3 al 30% tal como está descrito (7).
Los experimentos de RMN se realizaron en un espectrómetro Bruker DRX-600 MHZ, a 295 K, y la señal de agua se suprimió usando la estrategia NoesyPrlD. Los datos se procesaron y analizaron empleando el programa xwinnmr 1.3 en estaciones de trabajo Silicon Graphics. Los sistemas de "spin" se identificaron mediante experimentos bidimensionales TOCSY (8) y eos Y (9). Los experimentos NOESY (10) permitieron la asignación de señales de aminoácidos vecinos en el espacio. El tiempo de mezclado escogido fue 400 ms, y el ancho espectral 12 ppm en ambas dimensiones.
Cálculo de estructura molecular
Las distancias interprotónicas se calcularon a partir de los experimentos NOESY. Los voliímenes de los "cross-peaks" se ajustaron de acuerdo con la relación: dij/drcf = (Vij/Vrcf)"*' (11). Las distancias interprotónicas se calcularon empleando la distancia protónica 8-8 en la Pro7 como dref para el 1585 y sus análogos Pse-9473 y Pse-9475. Posteriormente, se generaron familias de estructuras mediante el procesamiento de los datos en estaciones de trabajo índigo 2 para mo-delización molecular.
Los espectros ID y 2D se procesaron usando el programa FÉLIX 97.0, y las familias estructurales se generaron mediante el procesamiento de datos con NMRchitect, por combinación de los pro
gramas "distance geometry" (DGII) y anillamiento simulado. Las intensidades de los "cross-peaks" NOE se clasificaron en rangos, así: fuertes 1,8 A-2,8 A, me-dianos"2,8 Á-3,5 A y débiles 3,5 Á-5,0 Á. Los ángulos dihedros y las asignaciones estereoespecíficas se ajustaron como está descrito (12). Los enlaces peptídicos se forzaron a configuración trans y los carbonos a a quiralidad L. Las estructuras obtenidas se refinaron y se seleccionaron aquellas con geometrías razonables y bajas violaciones a las restricciones de conformación.
RESULTADOS Y DISCUSIÓN
Síntesis e identificación de pseudopéptidos del 1585
Los pseudopéptidos M^[CH:-NH] amida reducida derivados del péptido 1585 se diseñaron tomando como base la secuencia de aminoácidos de la proteína MSP-i esquematizada en la Figura 1, en la cual se ubica la estructura primaria del fragmento correspondiente al péptido blanco de este estudio (MSP-I '-*-"" ') ; éstos se codificaron como Pse-9473 (modificado entre los residuos KnsT-Piiss) y Pse-9475 (modificado entre los residuos Pl288-L|2S9)-
En la Figura 2 se observa la comparación directa entre la conformación de un péptido en su forma natural constituido por residuos de configuración L (en la parte superior de la misma) respecto a análogos pseudopeptídicos de los tipos Retro y Retroinverso, y en la parte inferior los análogos objeto de este estudio del tipo amida reducida *P-[CH:-NH], resaltados por un marco sólido. Los enla-
10

REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 32, No. 1 DE 2003
a)
b)
c)
>r"Y
BocNH CH
(3) W (50-70%)
Figura 4. Síntesis de iV-í-Boc-aminoaldehídos a partir de Aí-í-Boc-aminoácidos.
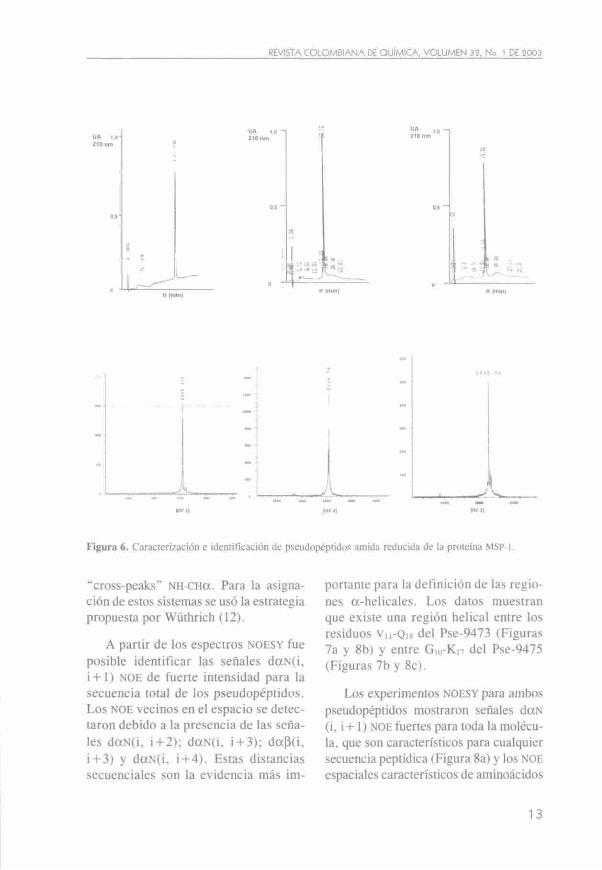
ees peptídicos seleccionados para su modificación se observan en la Figura 3. La letra griega Pse (^) representa a la expresión Pseudo. Tales análogos se obtuvieron en cantidades normales con rendimientos mayores al 70%, de acuerdo con los procedimientos descritos en las Figuras 4 y 5. Cuando fue necesario, se purificaron por HPLC-FR preparativa hasta homogeneidad > 98 %. Todas las moléculas fueron solubles en agua. Las masas moleculares experimentales para cada molécula estuvieron acordes con los valores esperados; para el péptido no modificado 1585 se encontró experimentalmente una masa molecular relativa 2340,21 m/z (valor esperado: 2348,04 Da). Para el Pse-9473 se obtuvo un valor de 2334,79 m/z (valor esperado: 2334,0 Da) y para el Pse-9475 se obtuvo un valor de 2335,52 m/z (valor esperado: 2334,0 Da), como puede observarse en la Figura 6.
En la figura 4 se observa (a) el carbono carbonílico de un N-Boc-a-aminoaci-
do comercial 1 se protege con O.A'-di-metilhidroxilamina formando así el derivado íV.o-dimetilcarboxamida 2. Posteriormente, la Míí-dimetilcarboxamida 2 se trata con LÍAIH4 a -10"C, para producir la sal de Litio 3, como puede observarse en (b). La sal de Litio 3 se hidroliza en medio ácido a temperatura ambiente (TA) para dar lugar a la formación del .v-r-Boc-a-aminoaldehido 4 como se observa en (c).
La síntesis de los enlaces isostéricos T[CH2-NH] se lleva a cabo mediante ami-nación reductiva in situ, luego de la condensación de los grupos CHO de 4 y el grupo amino libre de 5 (figura 5). Al realizar la desprotección y neutralización de la función amino del úUimo residuo de la cadena peptídica, anclada previamente a la resina o soporte sólido, representado en la gráfica como una esfera gris 5, se hace reaccionar con el A'-í-Boc-a-ami-noaldehido 4 obtenido en la etapa anterior (Figura 4) para formar el aducto 6. Posteriormente, este intermediato se tra-
11

REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 32, No 1 DE 2003
1. A c O H 1 % / D M F
2. NaCNBHs
3. Síntesis en
1. Acido tr i f luoroacético
2. HF (bajo/alto)
V 3. AcOH 5% H2O
NH2 (60-70%)
Figura 5. Síntesis en fase sólida de isósteros H'lCH:- NH] amida reducida.
ta con NaCNBHí para reducir el ion iminio y formar el grupo metileno. Finalmente, el pseudopéptido 7 se obtiene mediante las estrategias normales de síntesis de péptidos en fase sólida como está descrito en la literatura.
En la figura 6 se observa: a) En la parte superior, el perfil cromatográfico (HPLC-FR) obtenido para el péptido no modificado 1585 (MSP-1'-«"»') luego de la elución de una muestra de material liofilizado y reconstituido en solvente A en las condiciones descritas en Materiales y Métodos; en la parte inferior aparece el espectro de masas (MALDI) correspondiente, b) El Pse-9473 (Ki:87- Piiss). c) El Pse-9475 (P1288- L^ss). Los valores expe
rimentales de la masa molecular en cada caso correspondieron con el valor esperado.
Estudios de identificación y caracterización de los pseudopéptidos del fragmento MSP-l'̂ ^^-'̂ "' de malaria
Asignación de los espectros 2D-RMN protónicos
Los "cross-peaks" entre HN y el protón CHa se identificaron por análisis de los espectros COSY obtenidos en TFED3 al 30% en H:0. Los espectros TOCSY se emplearon para correlacionar los sistemas de "spin" de las cadenas laterales con los
12
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 32, No. 1 DE 2003
UA 1.0 210 nm
UA 1.0 210 n m
V : . ; ift;^^
;,L }
Figura 6. Caracterización e identificación de pseudopéptidos amida reducida de la proteína MSP-1.
"cross-peaks" NH-CHa. Para la asignación de estos sistemas se usó la estrategia propuesta por Wüthrich (12).
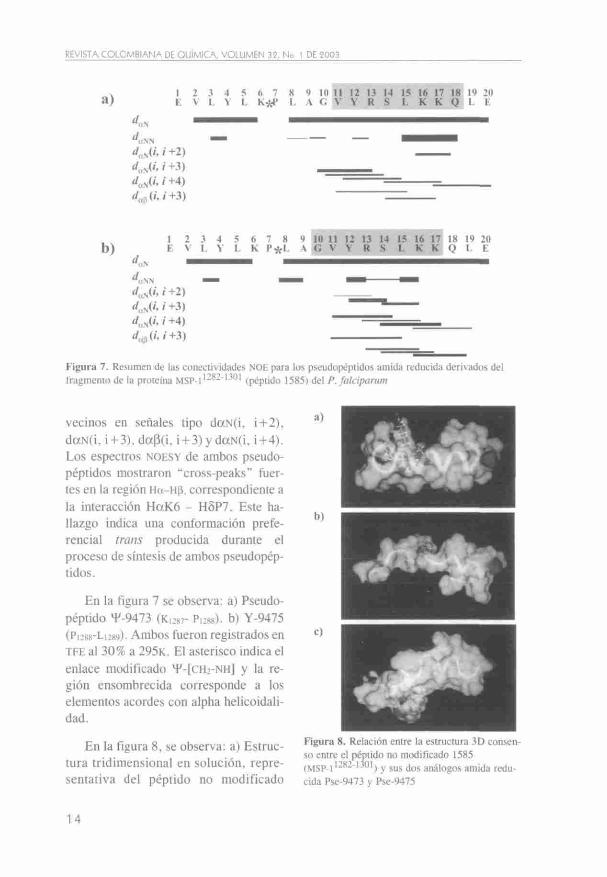
A partir de los espectros NOESY fue posible identificar las señales daN(i, i-1-1) NOE de fuerte intensidad para la secuencia total de los pseudopéptidos. Los NOE vecinos en el espacio se detectaron debido a la presencia de las señales daN(i, i-f2); daN(i, i-f3); dap(i, i-l-3) y daN(i, i-l-4). Estas distancias secuenciales son la evidencia más im
portante para la definición de las regiones a-helicales. Los datos muestran que existe una región helical entre los residuos Vn-Qig del Pse-9473 (Figuras 7a y 8b) y entre G10-K17 del Pse-9475 (Figuras 7b y 8c).
Los experimentos NOESY para ambos pseudopéptidos mostraron señales daN (i, i -h 1) NOE fuertes para toda la molécula, que son característicos para cualquier secuencia peptídica (Figura 8a) y los NOE espaciales característicos de aminoácidos
13
REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 32, No 1 DE 2003
a) 1 2 3 4 5 6 7 8 9 10 II 12 U 14 15 16 17 18 I"» 20 E V L Y L K*P L A G V Y R S L K K Q I E
"aNN *'aN('.'+2) < N ( ' . « - ^ 3 )
< N ( ' ' . ' + 4 ) í/„p(/,/+3)
b)
"oNN ' ' a^( ' • . ' •+2) do,-,(i,i+3) dai'i+4) í/„p(i, í-(-3)
I 2 3 4 5 6 7 8 9 W H P W I P W Í P P 18 19 20 E V L Y L K P ^ L A G V Y R S L K K Q L E
Figura 7. Resumen de las conectividades NOE para los pseudopéptidos amida reducida derivados del fragmento de la proteína MSP-I'-^^''-^'" (péptido 1585) del P. falciparum
vecinos en señales tipo daN(i, \+2), daN(i, i-l-3), da(i(i, i-l-3)ydaN(i, i-l-4). Los espectros NOESY de ambos pseudopéptidos mostraron "cross-peaks" fuertes en la región Ha-Hp, correspondiente a la interacción HaK6 - H5P7. Este hallazgo indica una conformación preferencial trans producida durante el proceso de síntesis de ambos pseudopéptidos.
En la figura 7 se observa: a) Pseudopéptido H'-9473 (K,287- Pi:88). b) Y-9475 (Pi288-Li289). Ambos fueron registrados en TFE al 30% a 295K. El asterisco indica el enlace modificado T-[CH2-NH] y la región ensombrecida corresponde a los elementos acordes con alpha helicoidali-dad.
En la figura 8, se observa: a) Estructura tridimensional en solución, representativa del péptido no modificado
a)
b)
c)
Figura 8. Relación entre la estructura 3D consenso entre el péptido no modificado 1585 (MSP-l'-^'"'-'^') y sus dos análogos amida reducida Pse-9473 y Pse-9475
14

REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 32, No. 1 DE 2003
1585. b) Pseudopéptido y[CH2-NH] amida reducida Pse-9473. c) Pseudopéptido Pse-9475. El esqueleto peptídico de cada molécula se muestra en cinta color claro que indica la estructura molecular más representativa. La superficie accesible al agua de cada molécula se observa en transparencia.
Determinación estructural de pseudopéptidos
Luego de generar familias estructurales para cada una de las moléculas, se obtuvieron las estrucmras consenso representativas para la estructura tridimensional de cada pseudopéptido, como se observa en la Figura 8. En ella se muestran las estrucmras tridimensionales en solución, representativas del péptido no modificado 1585 en (a), del pseudopéptido T[CH2- NH] amida reducida Pse-9473 en (b), y del pseudopéptido Pse-9475 en (c). Cada estructura se caracteriza por tener el mínimo valor de energía posible y por cumplir con las restricciones de conformación permitidas. La introducción de un solo enlace isóstero del tipo 4̂ [CH2-NH] amida reducida, no sólo indujo cambios estructurales fuertes en los análogos del péptido 1585 (MSP-i'-*- '̂ '") sino que además, los dos nuevos grados de libertad debidos al grupo metileno al gobernar las propiedades conformacionales de este fragmento indujeron de manera sitio-dirigida nuevas propiedades inmuno-lógicas a cada análogo (13).
CONCLUSIONES
En fase sólida se sintetizaron diferentes pseudopéptidos amida reducida derivados del fragmento de 42 kDa de la proteí
na MSP-1 del Plasmodium falciparum. Tanto los análogos como el péptido nativo se caracterizaron por sus propiedades físicas y químicas mediante técnicas cromatográficas y espectrofotométricas de HPLC-FR, espectrometría de masas y RMN. Se identificaron plenamente los elementos de estructura secundaria para cada molécula mediante los análisis y el procesamiento computacional de los datos espectrales obtenidos y dinámica molecular restringida. La sustitución de un sólo enlace peptídico por un enlace ^[CHi-NH] amida reducida produjo fuertes cambios estrucmrales en la conformación global de la molécula respecto a la no modificada y, de esta manera, se afectó el esqueleto peptídico de las mismas. Los datos aquí presentados, junto con los estudios sobre las propiedades biológicas de estas moléculas, permiten afirmar que la síntesis en fases líquida y sólida de este tipo de compuestos es factible arrojando rendimientos deseables en condiciones normales de laboratorio. Por sus propiedades químicas y físicas es posible pensar en la obtención de cantidades mucho mayores en escalas piloto e industrial hacia la obtención de nuevas moléculas candidatas a vacunas contra la malaria y otras enfermedades transmisibles.
AGRADECIMIENTOS
Este trabajo fue financiado por la Presidencia de la República de Colombia, a través del Ministerio de Protección Social. Agradecemos a la doctora Fabiola Espejo por permitirnos emplear para este trabajo la estructura tridimensional consenso del péptido no modificado 1585.
15

REVISTA COLOMBIANA DE QUÍMICA, VOLUMEN 32, No 1 DE 2003
BIBLIOGRAFÍA
1. Holder A.; LockyerM. J.; OdinkK. G. (1985). Primary stmcture of the precursor to the three major antigens of Plasmodium falciparum merozoi-tes. Nature 311. 270-273.
2. U rquiza M.; Rodríguez L.; Suárez J. et al. (1996). Identification Plasmodiumfalciparum MSP-1 peptides able to bind to human red blood cells. Parasite Immunology 18, 505-526.
3. Guichard G.; Calbo S.; Muller S.; Kourilsky P.; Abastado J. P. (1995). Efficient binding of reduced peptide bond pseudopeptides to major histo-compatibility complex class I molecule. J. Biol. Chem. 270, 26057- 26064.
4. Lozano J. M.; Espejo F.; Díaz D.; Salazar L. M.; Rodríguez J.; Pinzón C ; Calvo J. C ; Guzmán F.; Patarro-yo M. E. (1998). Reduced amide pseudopeptide analogues of a malaria peptide possess secondary structural elements responsible for induction of functional antibodies which react with native proteins expressed in Plasmodium falciparum erythrocyte stages. 772̂ Journal of Peptide Research 52, (6):457-469.
5. Merrifield R.B. (1963). Solid phase peptide synthesis. 1. The synthesis of a tetrapeptide J. Am. Chem. Soc. 85, 2149.
6. Houghten R.A. (1985). General method for the rapid solid-phase peptide synthesis of the large number of peptides: specificity of antigen-antibody interactions of the level of individual
amino acids. Proc. Nati, Acad. Sci. USAS2,5l3l.
1. Walgers R.; Lee T.; Cam-mers-Goodwin A. (1998). An indi-rect chaotropic mechanism for the stabilisation of helix conformation of peptides in aqueous trifluoroethanol and hexafluoro-2-propanol. J. Am. Chem. Soc. 12, 5073-5079.
8. Piotto M.; Saudek K. V.; Sklenar V. (1992). Gradient tailored excitation for single quantum NMR spectroscopy of aqueous solution. J. Biolmol. NMR. 2, 661-666.
9. Ranee M.; Sorenssen O.; Bodenhau-sen G.; Wagner G.; Ernst R.; Wüthrich, K. (1983). Improved spectral resolution in COSY'H-NMR spectra of proteins via Double Quantum Filte-ring. Biochem. Biophys. Res. Commun. 117, 479-485.
lO.Jeener J.; Meier B.; Backman P.; Ernst R. (1979). Investigation of en-hance processes by two dimensional NMR spectroscopy. J.Chem.Phys. 71, 4546-4553.
ll.Plugariu C ; Stockner T.; Sterk H.; HaegenauerG. (1998). Conformatio-nal behaviour of the TraMheadpiece. J.Peptide Res. 51, 244-250.
12. Wüthrich, K. (1986). NMR of Proteins and Nucleic Acids, 2"'̂ edition. New York: Willey and Sons, p. 207-215.
13. Lozano J. M.; Alba M. P., Vanegas M.; Silva Y.; Torres-Castellanos J. L; Patarroyo M. E. (2003). MSP-i Malaria Pseudopeptide Analogs: Biological and Immunological Significance and Three-Dimensional Strucmre. Biol, Chem. 384, 71-82.
16



![fJZíg u}uEvþt a± ¶YísÝ[Y · fJZíg pÉp Iu)fIo uEv Iuýu1wuî Esí`Ñ uñu}uEi!\ fJua\ fJZíg pÉp Iu)fIt¡sîb \ tñ ao ` \ fJZíg hA~a]ua t oybÑuapÉp Io Z±s½jsîbs¡si](https://img.pdfslide.net/doc/110x75/5ec429ebc2f29309057eb286/fjzg-uuevt-a-ysy-fjzg-pp-iufio-uev-iuu1wu-es-uuuei.jpg)





![fJZíg u}uEvþt a± ¶YísÝ[Ykoreascience.or.kr/article/JAKO201307954512177.pdfuñu}uEi!\ fJZíg pÉp IZíf \ fJZíg hA~ a]ua t oybÑs½`í IuEi!\ fJZíg u}uEvþt a± YísÝ[Y A Study](https://img.pdfslide.net/doc/110x75/6091f24926db85780c6713fe/fjzg-uuevt-a-ys-uuuei-fjzg-pp-izf-fjzg-ha-aua-t-oybs.jpg)
![%põkm %gip ¶km %fIYíw` ZÞkm %uEjâ %gitñ I u IsÝ ýva]ua Z Vccl.snu.ac.kr/papers/journal_dome/journaldome2012_01.pdf · ¢pÍ pÉp Is½o uño b` pÍ %` `Ñge pÍ %t-uE Yño](https://img.pdfslide.net/doc/110x75/5ebb9bc1e6603a63c61ee417/pkm-gip-km-fiyw-zkm-uej-git-i-u-is-vaua-z-vcclsnuackrpapersjournaldomejournaldome201201pdf.jpg)


![Morfología e inmunofenotipo de las células plasmáticas en el ......teína M, lo que da lugar a citopenias y resor-ción ósea [1-3]. Esta alteración hematológica fue clasificada,](https://img.pdfslide.net/doc/110x75/60c8e3365cf9ea336052dbb8/morfologa-e-inmunofenotipo-de-las-clulas-plasmticas-en-el-tena-m.jpg)














