Embed Size (px)
Citation preview

Biochem. J. (2009) 417, 791–801 (Printed in Great Britain) doi:10.1042/BJ20081149 791
Modulating serine palmitoyl transferase (SPT) expression and activityunveils a crucial role in lipid-induced insulin resistance in rat skeletalmuscle cellsMaria L. WATSON*, Matthew COGHLAN† and Harinder S. HUNDAL*1
*Division of Molecular Physiology, Sir James Black Centre, College of Life Sciences, University of Dundee, Dundee, DD1 5EH, Scotland, U.K., and †AstraZeneca, CVGI, Alderley Park,Macclesfield, Cheshire SK10 4TG, U.K.
Saturated fatty acids, such as palmitate, promote accumulationof ceramide, which impairs activation and signalling of PKB(protein kinase B; also known as Akt) to important end pointssuch as glucose transport. SPT (serine palmitoyl transferase) isa key enzyme regulating ceramide synthesis from palmitate andrepresents a potential molecular target in curbing lipid-inducedinsulin resistance. In the present study we explore the effects ofpalmitate upon insulin action in L6 muscle cells in which SPTexpression/activity has been decreased by shRNA (small-hairpinRNA) or sustained incubation with myriocin, an SPT inhibitor.Incubation of L6 myotubes with palmitate (for 16 h) increasesintramyocellular ceramide and reduces insulin-stimulated PKBactivation and glucose uptake. PKB inhibition was not associatedwith impaired IRS (insulin receptor substrate) signalling and wasameliorated by short-term treatment with myriocin. SilencingSPT expression (∼90%) by shRNA or chronic cell incubationwith myriocin (for 7 days) markedly suppressed SPT activityand palmitate-driven ceramide synthesis; however, challengingthese muscle cells with palmitate still inhibited the hormonal
activation of PKB. This inhibition was associated with reducedIRS1/p85-PI3K (phosphoinositide 3-kinase) coupling that arisesfrom diverting palmitate towards greater DAG (diacylglycerol)synthesis, which elevates IRS1 serine phosphorylation viaactivation of DAG-sensitive PKCs (protein kinase Cs). Treatmentof SPT-shRNA cells or those treated chronically with myriocinwith PKC inhibitors antagonized palmitate-induced loss ininsulin signalling. The findings of the present study indicatethat SPT plays a crucial role in desensitizing muscle cells toinsulin in response to incubation with palmitate. While short-term inhibition of SPT ameliorates palmitate/ceramide-inducedinsulin resistance, sustained loss/reduction in SPT expression/activity promotes greater partitioning of palmitate towards DAGsynthesis, which impacts negatively upon IRS1-directed insulinsignalling.
Key words: ceramide, diacylglycerol, insulin receptor substrate1 (IRS1), myriocin, palmitate, protein kinase B (PKB), proteinkinase C (PKC).
INTRODUCTION
Hyperlipidaemia is one of a cluster of abnormalities associatedwith the metabolic syndrome and one that is considered importantfor both development and progression of insulin resistance inskeletal muscle, a tissue that makes a major contribution to whole-body glucose homoeostasis. Sustained over supply of NEFAs(non-esterified fatty acids; ‘free’ fatty acids) to skeletal muscle hasbeen shown to result in the accumulation of IMTGs (intramusculartriacylglycerols) and that of fatty-acid-derived metabolites, suchas DAG (diacylglycerol) and ceramide. Although accumulationof IMTG has been positively correlated with a loss in skeletalmuscle insulin sensitivity, the finding that muscle of endurance-trained athletes exhibit improved insulin sensitivity in the faceof an increase in IMTG implies that increases in intramuscularlipid alone may not in itself be the primary driver of lipid-inducedinsulin resistance [1]. Recent work from our laboratory and workof others has shown that increased availability of palmitate, themost prevalent circulating saturated fatty acid, impairs the insulin-dependent regulation of glucose transport and glycogen synthesisin skeletal muscle cells [2,3]. This impairment is due, in large part,to an attendant accumulation in muscle cells of the sphingolipidceramide, for which the fatty acid is a key precursor. Increasesin intramyocellular ceramide have previously been linked to loss
of insulin sensitivity in cultured muscle cells [4,5] and in skeletalmuscle of both rodents [6,7] and humans [8]. However, it is onlyrecently that the mechanistic basis of ceramide-induced insulinresistance is beginning to be understood.
We [2,4,9], and others [5,10,11], have shown that increasesin intracellular ceramide promote a targeted loss in the insulin-dependent activation of the serine/threonine kinase PKB (proteinkinase B; also known as Akt). This kinase plays a pivotal rolein the hormonal activation of glucose transport and glycogensynthesis, and consequently its dysregulation is likely to impactsignificantly upon the homoeostatic control of blood glucose. Theloss of PKB activation by ceramide has been proposed to occurby one of two mechanisms involving either dephosphorylationof its two regulatory sites (Thr308 and Ser473) by a type 2A-likephosphatase activity [10] or alternatively, as we have shown,inhibition in the cell-surface recruitment and phosphorylationof PKB [4,12] by a process dependent upon activation ofatypical PKC (protein kinase C) isoforms (λ/ζ ) [9,11]. Increasingintracellular ceramide [by provision of short-chain (C2) ceramideor stimulating endogenous palmitic (C16)-ceramide synthesis fromfatty acids such as palmitate] results in potent activation ofPKCζ (for a review, see [13]) and, as such, this has importantimplications for PKB activation. In unstimulated cells, PKCζ hasbeen shown to directly interact with and repress PKB in numerous
Abbreviations used: DAG, diacylglycerol; DTT, dithiothreitol; FBS, foetal bovine serum; IMTG, intramuscular triacylglycerol; IRS, insulin receptor substrate;LCB, long-chain base; α-MEM, α-minimal essential medium; NEFA, non-esterified fatty acid; NF-κB, nuclear factor κB; PI3K, phosphoinositide 3-kinase;PKB, protein kinase B; PKC, protein kinase C; shRNA, small-hairpin RNA; SPT, serine palmitoyl transferase; TLR, Toll-like receptor.
1 To whom correspondence should be addressed (email [email protected]).
c© The Authors Journal compilation c© 2009 Biochemical Society

792 M. L. Watson, M. Coghlan and H. S. Hundal
cell types [9,11,14–17], but dissociates upon cell treatment withgrowth factors [14] and insulin [9], thus allowing PKB to becomeactivated. In contrast, ceramide stabilizes the interaction betweenthe two kinases and, moreover, ablates the ability of insulin to notonly dissociate the kinase complex, but to also activate PKB [9].It follows that limiting intramuscular ceramide production and/oraccumulation would confer an insulin-sensitizing effect and thatenzymes participating in ceramide synthesis may potentially serveas therapeutic targets for promoting insulin sensitivity of tissuessuch as skeletal muscle. Indeed, recent in vivo work suggeststhat attenuating endogenous ceramide synthesis ameliorates theloss in skeletal muscle insulin sensitivity induced in response tosaturated fat, glucocorticoids and obesity [18].
The synthesis of ceramide from saturated fatty acids, suchas palmitate, is crucially dependent upon the activity of SPT(serine palmitoyl transferase), which catalyses the first and rate-limiting step of sphingolipid biosynthesis involving the condens-ation of the amino acid L-serine with palmitoyl-CoA to form3-oxodihydrosphingosine [19]. Mammalian SPT belongs to thefamily of pyridoxal 5′-phosphate-dependent enzymes and is com-posed of two distinct protein subunits known as LCB1 (long-chainbase-1; 53 kDa) and LCB2 (long-chain base-2; 63 kDa), withstability of the latter being critically dependent upon expressionof the former. More recently, a third SPT subunit (LCB3) hadbeen identified, but given that its tissue expression is highlyvariable, being expressed predominantly in the placenta andcells of trophoblastic origin [20], it is likely to be dispensablewith regard to SPT function in other cell and tissue types. SPTactivity is susceptible to inhibition by serine analogues such ascycloserine and by molecules that structurally resemble transition-reactive intermediates such as the anti-fungal agent myriocin [19].In the present study we have modulated SPT expression andactivity in cultured rat skeletal muscle cells by either silencingthe expression of the LCB1 gene or subjecting muscle cellsto sustained incubation with myriocin in an attempt to assessthe impact of these interventions on palmitate-induced insulinresistance. We have shown previously that ceramide generatedde novo from palmitate is a major factor promoting insulinresistance in muscle cells and that short-term inhibition of SPThas an ameliorative effect on insulin signalling [2]. However,under circumstances when expression of SPT has been repressedor is chronically inhibited, we show that utilization of palmitate isdiverted towards greater synthesis of DAG, which subsequentlyactivates DAG-sensitive PKCs that in turn promote a reduction inIRS (insulin receptor substrate)-directed insulin signalling.
EXPERIMENTAL
Materials
α-MEM (α-minimal essential medium), FBS (foetal bovineserum) and antibiotic/antimycotic solution were from Life Tech-nologies. Most other reagent grade chemicals, myriocin,insulin, palmitate, palmitoyl-CoA, DAG, sn-1,2-DAG kinase andantibodies against actin were purchased from Sigma–Aldrich.Ro 31.8220 and GF109203X were from Calbiochem–Merck.C2-ceramide was from Tocris and C16-ceramide was purchasedfrom Avanti. Radioactive chemicals, 2-deoxy [3H]glucoseand [3H]serine were purchased from PerkinElmer. [γ -32P]ATP and protein-G–Sepharose beads were from AmershamBiosciences. Antibodies to PKB/Akt, phospho-PKB(Ser473),PKCθ and phospho-IRS1(Ser1101) were purchased from NewEngland Biolabs. Antibodies against the p85 subunit of PI3K(phosphoinositide 3-kinase) and anti-IRS1 were obtained fromUpstate–Millipore. Anti-LCB2 antibody was from Abcam and
antibodies against LCB1 were synthesized by AstraZeneca.Anti-α1 Na/K-ATPase (α6F) antibody was obtained from theUniversity of Iowa Hybridoma Bank and the anti-phosphoserineantibody was from Chemicon–Millipore. BSA-V (Fraction V,fatty-acid free) for conjugating with palmitate, and completeprotein phosphatase inhibitor tablets were purchased fromBoehringer–Roche Diagnostics.
Cell culture and treatments
L6 skeletal muscle cells were maintained in α-MEM anddifferentiated in culture to the stage of myotubes as previouslydescribed [21]. L6 myotubes were incubated for the time periodsand with the appropriate effectors as indicated in the Figurelegends. Plates were washed twice with ice-cold PBS and lysisbuffer was added [50 mM Tris/HCl (pH 7.4), 0.27 M sucrose,1 mM sodium orthovanodate, 1 mM EDTA, 1 mM EGTA, 10 mMsodium β-glycerophosphate, 50 mM NaF, 5 mM sodiumpyrophosphate, 1% (v/v) Triton X-100, 0.1% 2-mercaptoethanoland protease inhibitors]. Whole-cell lysates were centrifuged at4000 g at 4 ◦C for 10 min and were stored at −20 ◦C after beingsnap–frozen in liquid nitrogen. In some experiments, L6 myotubeswere sub-fractionated to isolate total membranes as previouslydescribed [21]. Briefly, cells were scraped off plates using a rubberpoliceman in PBS and centrifuged at 4000 g at 4 ◦C for 10 min.The resulting cell pellet was resuspended in buffer I [20 mMHepes (pH 7.4), 250 mM sucrose, 2 mM EGTA, 3 mM sodiumazide and protease inhibitors] and homogenized with a Dounceglass-to-glass homogenizer. The homogenates were centrifugedat 750 g for 5 min to remove the nuclear pellet. The supernatantwas centrifuged at 67000 rev./min (Beckman rotor TLA 100.3)for 60 min to pellet total cell membranes.
SPT silencing using shRNA (short-hairpin RNA)
A short RNA sequence (5′-GGAGCGTTCTGATCTTACA-3′)complimentary to the LCB1 subunit of SPT, was chosen fromsequences obtained from web tools on the Dharmacon (http://www.dharmacon.com), Promega (http://www.dharmacon.com)and Eurogentic (http://www.eurogentec.com) websites. Once asuitable sequence was identified, a BLAST search was performedto check and exclude those sequences that generated non-specific hits. The sequence was then cloned into the siSTRIKEvector (Promega) to create a hairpin loop sequence (5′-AAC-CGGAGCGTTCTGATCTTACATTCAAGAGATGTAAGATC-AGAACGCTCCTTTTT-3′). The vector was transfected (1 μg ofDNA) into L6 myoblasts using FuGENETM reagent (Boehringer–Roche). Myoblasts were washed free of transfection medium5 h later and were incubated for a further 2 days in α-MEMcontaining 2% (v/v) FBS and 1 % (v/v) antibiotic/antimycoticsolution. Stable transformants were subsequently selected byaddition of 800 μg/ml G418 (Geneticin) disulfate to the culturemedium. Transformed L6 myoblasts were selected using cloningdiscs (Sigma–Aldrich) and propagated to generate a workingstock cell line. Silencing of SPT was confirmed by usingantibodies against the LCB1 and LCB2 subunits and by analysisof cellular SPT activity.
SDS/PAGE and immunoblotting
Cell lysates and total membrane fractions (15–50 μg ofprotein) were reconstituted in Laemmli buffer and subjected toSDS/PAGE on 10% or 7.5% resolving gels and transferredon to Immobilon-P membranes as described previously [21].Membranes were probed with primary antibodies of interestand subsequently detected using either HRP (horseradish
c© The Authors Journal compilation c© 2009 Biochemical Society

SPT and insulin resistance 793
peroxidase)-conjugated anti-rabbit IgG or anti-mouse-IgG (NewEngland Biolabs). Membranes were visualized using ECL®
(enhanced chemiluminescence; Pierce) on Konica Minolta X-raymedical film.
Hexose uptake
2-Deoxyglucose uptake was assayed as previously described [21].Briefly, after relevant incubations, L6 myotubes were washedtwice with HBS [Hepes-buffered saline; 140 mM NaCl, 20 mMHepes, 5 mM KCl, 2.5 mM MgSO4 and 1 mM CaCl2 (pH 7.4)]and incubated for 10 min in HBS containing 10 μM [3H]2-deoxy-D-glucose (1 μCi/ml) at room temperature (20 ◦C). Non-specific binding was determined by quantifying cell-associatedradioactivity in the presence of 10 μM cytochalasin B. Mediumwas aspirated and adherent cells were washed twice with0.9% (w/v) ice-cold NaCl. Cells were subsequently lysed in50 mM NaOH and cell-associated radioactivity was quantifiedusing a Beckman LS 6000IC scintillation counter. The proteinconcentration in cell lysates was determined using Bradfordreagent [22].
Analysis of cellular ceramide and DAG content
After appropriate cell treatments, myotubes were extracted fromplates using a rubber policeman in ice-cold PBS for analysisof ceramide and DAG using the DAG kinase method, whichrelies on the kinase catalysing the formation of ceramide 1-phosphate and phosphatidic acid respectively. Briefly, proteincontent in the harvested cells was assayed using the Bradfordmethod [22] and lipids were extracted from a cell aliquotcontaining 400 μg of protein by the addition of chloroform/methanol [1:2 (v/v)] with the phases being broken by theaddition of chloroform and 1 M NaCl. The organic phase wasseparated, dried and used for ceramide and DAG measurementsas previously described [2]. The lipids were incubated at roomtemperature for 30 min in a reagent mix containing sn-1,2-DAGkinase in diluted enzyme buffer [10 mM imidazole/HCl and1 mM diethylenetriaminepenta-acetic acid (pH 6.6)], assay buffer[0.1 M imidazole/HCl, 0.1 M NaCl, 25 mM MgCl2, 2 mM EGTA(pH 6.6)] and 20 mM DTT (dithiothreitol), and the reaction wasinitiated by the addition of 5 mM ATP (mixed with [γ -32P]ATP;1 μCi/reaction) and 1 μl of DAG kinase, which equates to anactivity of at least 1.3 nmol/min. The reaction was terminated bythe addition of 20 μl of 1% (v/v) perchloric acid and 450 μl ofchloroform/methanol [1:2 (v/v)]. The organic phase was isolatedand washed twice with 1 % (v/v) perchloric acid. The sampleswere dried and reconstituted in chloroform/methanol [95:5 (v/v)]and spotted on to TLC plates. Reaction products were separatedby placing the plate into a chamber equilibrated with a solution ofcholoroform/methanol/acetic acid (65:15:5, by vol.). Radioactiveproducts were visualized on Konica Minolta X-ray medical filmusing an X-ograph imaging system. Changes in ceramide andDAG content were expressed as a fold change relative to thatmeasured in untreated cells.
Analysis of SPT activity
SPT activity was analysed using the Merrill method as describedby Holleran et al. [23]. Briefly, 80–100 μg of total membraneprotein from L6 myotubes was incubated at 37 ◦C for 10 min ina reaction buffer (100 mM Hepes, 2.5 mM EDTA, 5 mM DTT,50 μM pyridoxal phosphate and 1 mM serine mixed with 1μCi/100 μl [3H]serine). The reaction was initiated by the additionof 20 μl of 1 mM palmitoyl-CoA and terminated by the addi-tion of 200 μl 0.5 M NH4OH and cooled on ice. The lipid phases
were broken by the addition of 1.5 ml of chloroform/methanol[1:2 (v/v)], 25 μl of sphingonine (1 mg/ml, as a carrier) and 2 mlof NH4OH. Samples were vortexed and centrifuged (3000 g at4 ◦C for 5 min) to separate the phases. The aqueous phase wasremoved and the organic phase was washed twice with deionizedwater. The organic phase (300 μl) was then dried under vacuumand reconstituted in chloroform/methanol/acetic acid (6.5:1:1, byvol.) and the radioactivity was quantified using a Beckman LS6000IC scintillation counter. Non-specific activity was assessedby assaying activity in wild-type cells in which the SPT substrate,palmitoyl-CoA, was excluded from the assay.
IRS1 immunoprecipitation
Following appropriate treatments, L6 myotubes were lysed inlysis buffer and IRS1 was immunoprecipitated using an antibodyagainst the C-terminal domain of IRS1. Immunocomplexes wereincubated with protein-G–Sepharose beads overnight at 4 ◦C andsolubilized in Laemmli sample buffer prior to immunoblotting asdescribed above.
Statistical analyses
Data analysis was performed using GraphPad Prism software andresults were considered statistically significant at P values lessthan 0.05.
RESULTS
Cellular silencing of SPT using shRNA
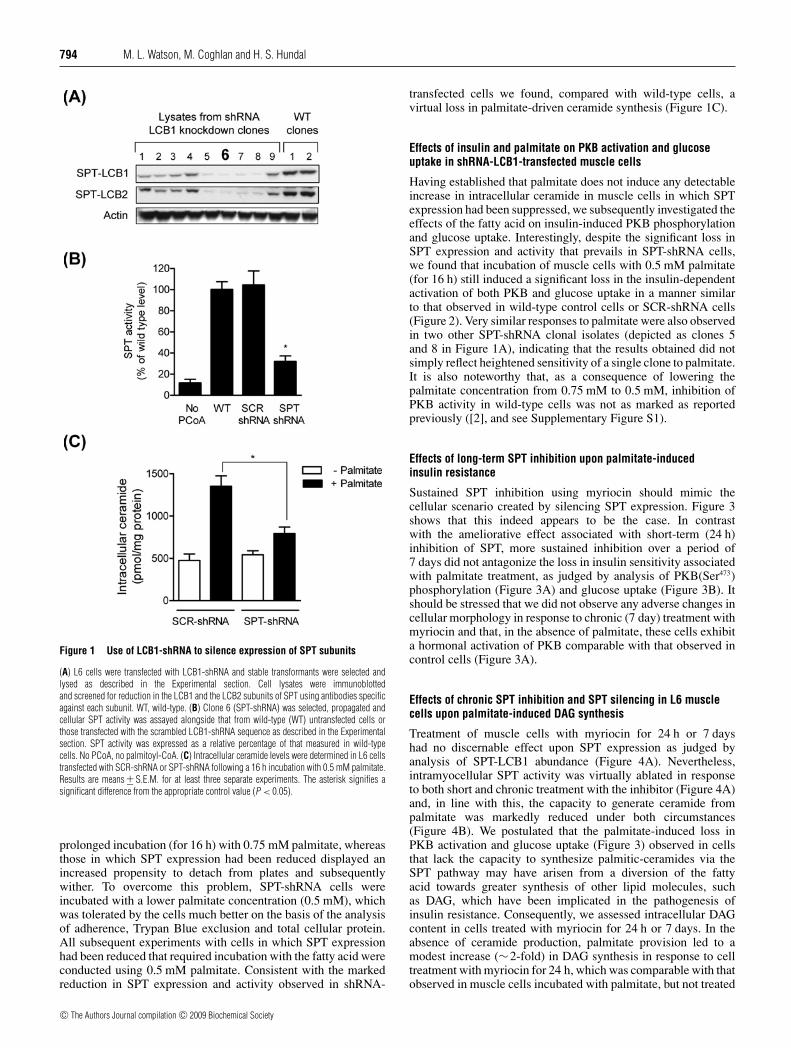
Previous work from our laboratory has shown that incu-bation of L6 myotubes for 16 h with 0.75 mM palmitateinduces a greater than 2-fold increase in intracellular ceramide,which promotes a significant decrease in the hormonal(insulin) activation of PKB and glucose uptake [2]. Attenuatingceramide synthesis from palmitate by co-incubation of cellswith myriocin (an SPT inhibitor) significantly curtails theceramide synthesis from palmitate and the associated decreasein PKB activation ([2] and see Supplementary Figure S1at http://www.BiochemJ.org/bj/417/bj4170791add.htm). On thebasis of the above observations we hypothesized that a sustainedsuppression in SPT expression and/or activity may protect musclecells against the insulin-desensitizing effects of palmitate. Totest this hypothesis, we generated a stable L6 muscle cell linein which cellular SPT expression was decreased substantiallythrough silencing the expression of the LCB1-SPT subunit usingshRNA. Nine positive L6 transformants were identified in whichexpression of LCB1 had been reduced by between 60 and 90%compared with wild-type clones. Of these, clone 6 (Figure 1A)exhibited the greatest reduction in LCB1 expression and wassubsequently used for further experimental analysis. Given thatthe stability of the LCB2-SPT subunit is inherently dependenton the expression of LCB1 and both subunits associate witha 1:1 molar stoichiometry [19], silencing of LCB1 also led toan attendant reduction in the expression of LCB2 (Figure 1A).Consistent with the loss in cellular SPT expression observedin shRNA-treated cells, analysis of SPT activity revealed thatthis was suppressed significantly (by ∼70%) in cells transfectedwith the shRNA-LCB1 sequence (SPT-shRNA) compared withwild-type L6 cells or those that had been transfected with thescrambled (SCR-shRNA) shRNA-LCB1 sequence (Figure 1B).An apparent consequence of silencing SPT expression in L6muscle cells was increased cellular sensitivity towards palmitate,which manifests itself as an increase in palmitate-induced celldeath. Wild-type L6 myotubes or SCR-shRNA cells tolerate
c© The Authors Journal compilation c© 2009 Biochemical Society
794 M. L. Watson, M. Coghlan and H. S. Hundal
Figure 1 Use of LCB1-shRNA to silence expression of SPT subunits
(A) L6 cells were transfected with LCB1-shRNA and stable transformants were selected andlysed as described in the Experimental section. Cell lysates were immunoblottedand screened for reduction in the LCB1 and the LCB2 subunits of SPT using antibodies specificagainst each subunit. WT, wild-type. (B) Clone 6 (SPT-shRNA) was selected, propagated andcellular SPT activity was assayed alongside that from wild-type (WT) untransfected cells orthose transfected with the scrambled LCB1-shRNA sequence as described in the Experimentalsection. SPT activity was expressed as a relative percentage of that measured in wild-typecells. No PCoA, no palmitoyl-CoA. (C) Intracellular ceramide levels were determined in L6 cellstransfected with SCR-shRNA or SPT-shRNA following a 16 h incubation with 0.5 mM palmitate.Results are means +− S.E.M. for at least three separate experiments. The asterisk signifies asignificant difference from the appropriate control value (P < 0.05).
prolonged incubation (for 16 h) with 0.75 mM palmitate, whereasthose in which SPT expression had been reduced displayed anincreased propensity to detach from plates and subsequentlywither. To overcome this problem, SPT-shRNA cells wereincubated with a lower palmitate concentration (0.5 mM), whichwas tolerated by the cells much better on the basis of the analysisof adherence, Trypan Blue exclusion and total cellular protein.All subsequent experiments with cells in which SPT expressionhad been reduced that required incubation with the fatty acid wereconducted using 0.5 mM palmitate. Consistent with the markedreduction in SPT expression and activity observed in shRNA-
transfected cells we found, compared with wild-type cells, avirtual loss in palmitate-driven ceramide synthesis (Figure 1C).
Effects of insulin and palmitate on PKB activation and glucoseuptake in shRNA-LCB1-transfected muscle cells
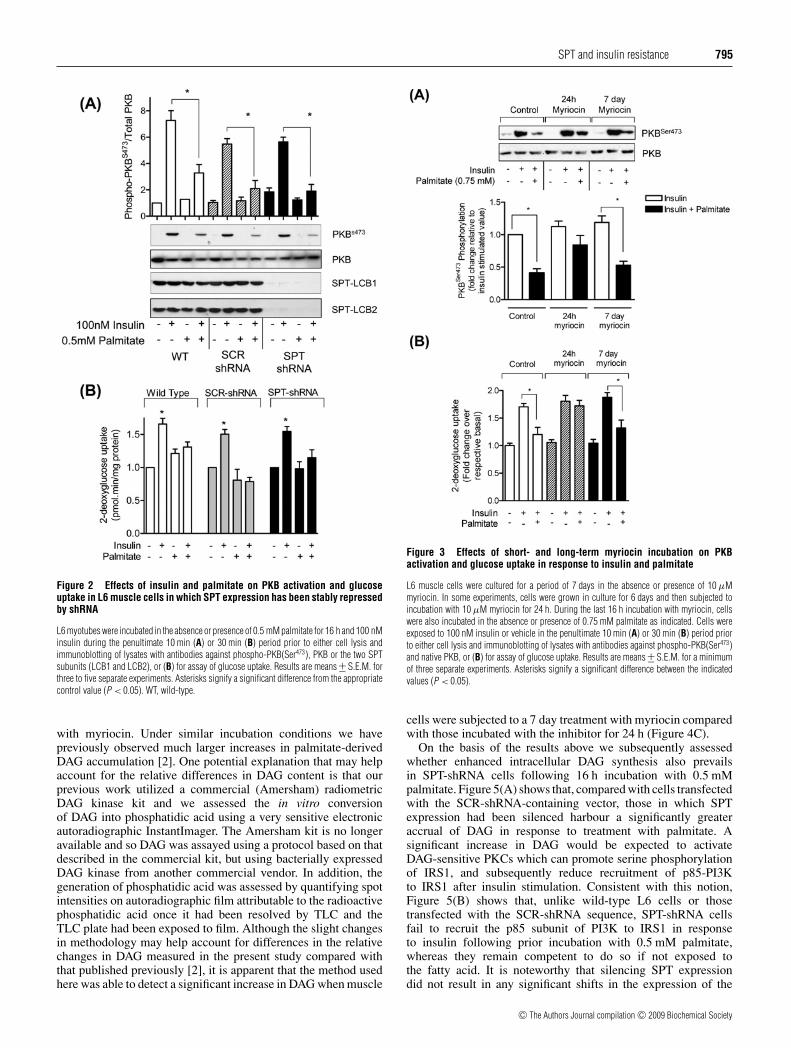
Having established that palmitate does not induce any detectableincrease in intracellular ceramide in muscle cells in which SPTexpression had been suppressed, we subsequently investigated theeffects of the fatty acid on insulin-induced PKB phosphorylationand glucose uptake. Interestingly, despite the significant loss inSPT expression and activity that prevails in SPT-shRNA cells,we found that incubation of muscle cells with 0.5 mM palmitate(for 16 h) still induced a significant loss in the insulin-dependentactivation of both PKB and glucose uptake in a manner similarto that observed in wild-type control cells or SCR-shRNA cells(Figure 2). Very similar responses to palmitate were also observedin two other SPT-shRNA clonal isolates (depicted as clones 5and 8 in Figure 1A), indicating that the results obtained did notsimply reflect heightened sensitivity of a single clone to palmitate.It is also noteworthy that, as a consequence of lowering thepalmitate concentration from 0.75 mM to 0.5 mM, inhibition ofPKB activity in wild-type cells was not as marked as reportedpreviously ([2], and see Supplementary Figure S1).
Effects of long-term SPT inhibition upon palmitate-inducedinsulin resistance
Sustained SPT inhibition using myriocin should mimic thecellular scenario created by silencing SPT expression. Figure 3shows that this indeed appears to be the case. In contrastwith the ameliorative effect associated with short-term (24 h)inhibition of SPT, more sustained inhibition over a period of7 days did not antagonize the loss in insulin sensitivity associatedwith palmitate treatment, as judged by analysis of PKB(Ser473)phosphorylation (Figure 3A) and glucose uptake (Figure 3B). Itshould be stressed that we did not observe any adverse changes incellular morphology in response to chronic (7 day) treatment withmyriocin and that, in the absence of palmitate, these cells exhibita hormonal activation of PKB comparable with that observed incontrol cells (Figure 3A).
Effects of chronic SPT inhibition and SPT silencing in L6 musclecells upon palmitate-induced DAG synthesis
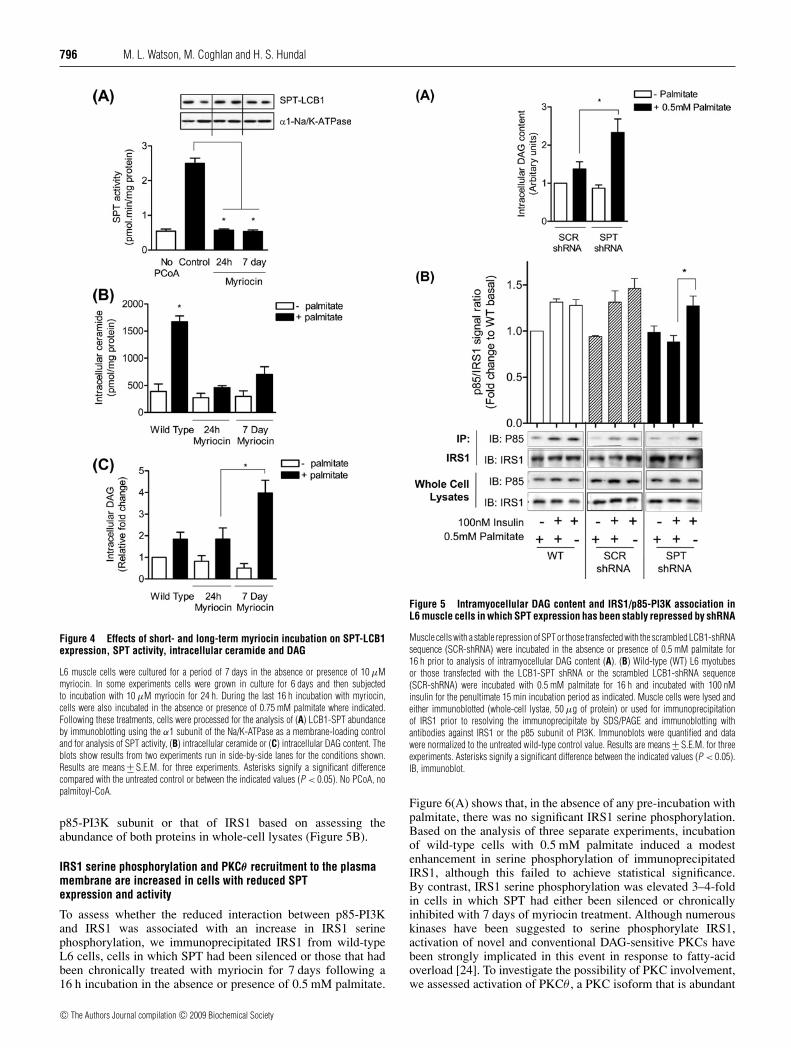
Treatment of muscle cells with myriocin for 24 h or 7 dayshad no discernable effect upon SPT expression as judged byanalysis of SPT-LCB1 abundance (Figure 4A). Nevertheless,intramyocellular SPT activity was virtually ablated in responseto both short and chronic treatment with the inhibitor (Figure 4A)and, in line with this, the capacity to generate ceramide frompalmitate was markedly reduced under both circumstances(Figure 4B). We postulated that the palmitate-induced loss inPKB activation and glucose uptake (Figure 3) observed in cellsthat lack the capacity to synthesize palmitic-ceramides via theSPT pathway may have arisen from a diversion of the fattyacid towards greater synthesis of other lipid molecules, suchas DAG, which have been implicated in the pathogenesis ofinsulin resistance. Consequently, we assessed intracellular DAGcontent in cells treated with myriocin for 24 h or 7 days. In theabsence of ceramide production, palmitate provision led to amodest increase (∼2-fold) in DAG synthesis in response to celltreatment with myriocin for 24 h, which was comparable with thatobserved in muscle cells incubated with palmitate, but not treated
c© The Authors Journal compilation c© 2009 Biochemical Society
SPT and insulin resistance 795
Figure 2 Effects of insulin and palmitate on PKB activation and glucoseuptake in L6 muscle cells in which SPT expression has been stably repressedby shRNA
L6 myotubes were incubated in the absence or presence of 0.5 mM palmitate for 16 h and 100 nMinsulin during the penultimate 10 min (A) or 30 min (B) period prior to either cell lysis andimmunoblotting of lysates with antibodies against phospho-PKB(Ser473), PKB or the two SPTsubunits (LCB1 and LCB2), or (B) for assay of glucose uptake. Results are means +− S.E.M. forthree to five separate experiments. Asterisks signify a significant difference from the appropriatecontrol value (P < 0.05). WT, wild-type.
with myriocin. Under similar incubation conditions we havepreviously observed much larger increases in palmitate-derivedDAG accumulation [2]. One potential explanation that may helpaccount for the relative differences in DAG content is that ourprevious work utilized a commercial (Amersham) radiometricDAG kinase kit and we assessed the in vitro conversionof DAG into phosphatidic acid using a very sensitive electronicautoradiographic InstantImager. The Amersham kit is no longeravailable and so DAG was assayed using a protocol based on thatdescribed in the commercial kit, but using bacterially expressedDAG kinase from another commercial vendor. In addition, thegeneration of phosphatidic acid was assessed by quantifying spotintensities on autoradiographic film attributable to the radioactivephosphatidic acid once it had been resolved by TLC and theTLC plate had been exposed to film. Although the slight changesin methodology may help account for differences in the relativechanges in DAG measured in the present study compared withthat published previously [2], it is apparent that the method usedhere was able to detect a significant increase in DAG when muscle
Figure 3 Effects of short- and long-term myriocin incubation on PKBactivation and glucose uptake in response to insulin and palmitate
L6 muscle cells were cultured for a period of 7 days in the absence or presence of 10 μMmyriocin. In some experiments, cells were grown in culture for 6 days and then subjected toincubation with 10 μM myriocin for 24 h. During the last 16 h incubation with myriocin, cellswere also incubated in the absence or presence of 0.75 mM palmitate as indicated. Cells wereexposed to 100 nM insulin or vehicle in the penultimate 10 min (A) or 30 min (B) period priorto either cell lysis and immunoblotting of lysates with antibodies against phospho-PKB(Ser473)and native PKB, or (B) for assay of glucose uptake. Results are means +− S.E.M. for a minimumof three separate experiments. Asterisks signify a significant difference between the indicatedvalues (P < 0.05).
cells were subjected to a 7 day treatment with myriocin comparedwith those incubated with the inhibitor for 24 h (Figure 4C).
On the basis of the results above we subsequently assessedwhether enhanced intracellular DAG synthesis also prevailsin SPT-shRNA cells following 16 h incubation with 0.5 mMpalmitate. Figure 5(A) shows that, compared with cells transfectedwith the SCR-shRNA-containing vector, those in which SPTexpression had been silenced harbour a significantly greateraccrual of DAG in response to treatment with palmitate. Asignificant increase in DAG would be expected to activateDAG-sensitive PKCs which can promote serine phosphorylationof IRS1, and subsequently reduce recruitment of p85-PI3Kto IRS1 after insulin stimulation. Consistent with this notion,Figure 5(B) shows that, unlike wild-type L6 cells or thosetransfected with the SCR-shRNA sequence, SPT-shRNA cellsfail to recruit the p85 subunit of PI3K to IRS1 in responseto insulin following prior incubation with 0.5 mM palmitate,whereas they remain competent to do so if not exposed tothe fatty acid. It is noteworthy that silencing SPT expressiondid not result in any significant shifts in the expression of the
c© The Authors Journal compilation c© 2009 Biochemical Society
796 M. L. Watson, M. Coghlan and H. S. Hundal
Figure 4 Effects of short- and long-term myriocin incubation on SPT-LCB1expression, SPT activity, intracellular ceramide and DAG
L6 muscle cells were cultured for a period of 7 days in the absence or presence of 10 μMmyriocin. In some experiments cells were grown in culture for 6 days and then subjectedto incubation with 10 μM myriocin for 24 h. During the last 16 h incubation with myriocin,cells were also incubated in the absence or presence of 0.75 mM palmitate where indicated.Following these treatments, cells were processed for the analysis of (A) LCB1-SPT abundanceby immunoblotting using the α1 subunit of the Na/K-ATPase as a membrane-loading controland for analysis of SPT activity, (B) intracellular ceramide or (C) intracellular DAG content. Theblots show results from two experiments run in side-by-side lanes for the conditions shown.Results are means +− S.E.M. for three experiments. Asterisks signify a significant differencecompared with the untreated control or between the indicated values (P < 0.05). No PCoA, nopalmitoyl-CoA.
p85-PI3K subunit or that of IRS1 based on assessing theabundance of both proteins in whole-cell lysates (Figure 5B).
IRS1 serine phosphorylation and PKCθ recruitment to the plasmamembrane are increased in cells with reduced SPTexpression and activity
To assess whether the reduced interaction between p85-PI3Kand IRS1 was associated with an increase in IRS1 serinephosphorylation, we immunoprecipitated IRS1 from wild-typeL6 cells, cells in which SPT had been silenced or those that hadbeen chronically treated with myriocin for 7 days following a16 h incubation in the absence or presence of 0.5 mM palmitate.
Figure 5 Intramyocellular DAG content and IRS1/p85-PI3K association inL6 muscle cells in which SPT expression has been stably repressed by shRNA
Muscle cells with a stable repression of SPT or those transfected with the scrambled LCB1-shRNAsequence (SCR-shRNA) were incubated in the absence or presence of 0.5 mM palmitate for16 h prior to analysis of intramyocellular DAG content (A). (B) Wild-type (WT) L6 myotubesor those transfected with the LCB1-SPT shRNA or the scrambled LCB1-shRNA sequence(SCR-shRNA) were incubated with 0.5 mM palmitate for 16 h and incubated with 100 nMinsulin for the penultimate 15 min incubation period as indicated. Muscle cells were lysed andeither immunoblotted (whole-cell lystae, 50 μg of protein) or used for immunoprecipitationof IRS1 prior to resolving the immunoprecipitate by SDS/PAGE and immunoblotting withantibodies against IRS1 or the p85 subunit of PI3K. Immunoblots were quantified and datawere normalized to the untreated wild-type control value. Results are means +− S.E.M. for threeexperiments. Asterisks signify a significant difference between the indicated values (P < 0.05).IB, immunoblot.
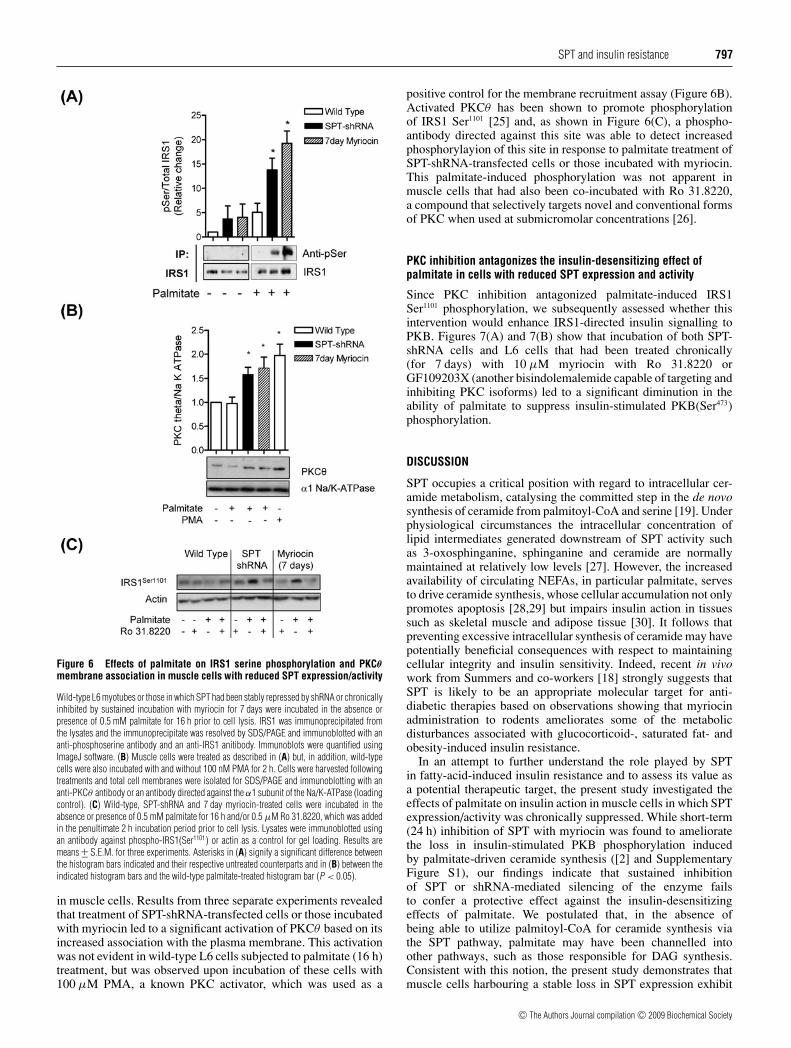
Figure 6(A) shows that, in the absence of any pre-incubation withpalmitate, there was no significant IRS1 serine phosphorylation.Based on the analysis of three separate experiments, incubationof wild-type cells with 0.5 mM palmitate induced a modestenhancement in serine phosphorylation of immunoprecipitatedIRS1, although this failed to achieve statistical significance.By contrast, IRS1 serine phosphorylation was elevated 3–4-foldin cells in which SPT had either been silenced or chronicallyinhibited with 7 days of myriocin treatment. Although numerouskinases have been suggested to serine phosphorylate IRS1,activation of novel and conventional DAG-sensitive PKCs havebeen strongly implicated in this event in response to fatty-acidoverload [24]. To investigate the possibility of PKC involvement,we assessed activation of PKCθ , a PKC isoform that is abundant
c© The Authors Journal compilation c© 2009 Biochemical Society
SPT and insulin resistance 797
Figure 6 Effects of palmitate on IRS1 serine phosphorylation and PKCθmembrane association in muscle cells with reduced SPT expression/activity
Wild-type L6 myotubes or those in which SPT had been stably repressed by shRNA or chronicallyinhibited by sustained incubation with myriocin for 7 days were incubated in the absence orpresence of 0.5 mM palmitate for 16 h prior to cell lysis. IRS1 was immunoprecipitated fromthe lysates and the immunoprecipitate was resolved by SDS/PAGE and immunoblotted with ananti-phosphoserine antibody and an anti-IRS1 anitibody. Immunoblots were quantified usingImageJ software. (B) Muscle cells were treated as described in (A) but, in addition, wild-typecells were also incubated with and without 100 nM PMA for 2 h. Cells were harvested followingtreatments and total cell membranes were isolated for SDS/PAGE and immunoblotting with ananti-PKCθ antibody or an antibody directed against the α1 subunit of the Na/K-ATPase (loadingcontrol). (C) Wild-type, SPT-shRNA and 7 day myriocin-treated cells were incubated in theabsence or presence of 0.5 mM palmitate for 16 h and/or 0.5 μM Ro 31.8220, which was addedin the penultimate 2 h incubation period prior to cell lysis. Lysates were immunoblotted usingan antibody against phospho-IRS1(Ser1101) or actin as a control for gel loading. Results aremeans +− S.E.M. for three experiments. Asterisks in (A) signify a significant difference betweenthe histogram bars indicated and their respective untreated counterparts and in (B) between theindicated histogram bars and the wild-type palmitate-treated histogram bar (P < 0.05).
in muscle cells. Results from three separate experiments revealedthat treatment of SPT-shRNA-transfected cells or those incubatedwith myriocin led to a significant activation of PKCθ based on itsincreased association with the plasma membrane. This activationwas not evident in wild-type L6 cells subjected to palmitate (16 h)treatment, but was observed upon incubation of these cells with100 μM PMA, a known PKC activator, which was used as a
positive control for the membrane recruitment assay (Figure 6B).Activated PKCθ has been shown to promote phosphorylationof IRS1 Ser1101 [25] and, as shown in Figure 6(C), a phospho-antibody directed against this site was able to detect increasedphosphorylayion of this site in response to palmitate treatment ofSPT-shRNA-transfected cells or those incubated with myriocin.This palmitate-induced phosphorylation was not apparent inmuscle cells that had also been co-incubated with Ro 31.8220,a compound that selectively targets novel and conventional formsof PKC when used at submicromolar concentrations [26].
PKC inhibition antagonizes the insulin-desensitizing effect ofpalmitate in cells with reduced SPT expression and activity
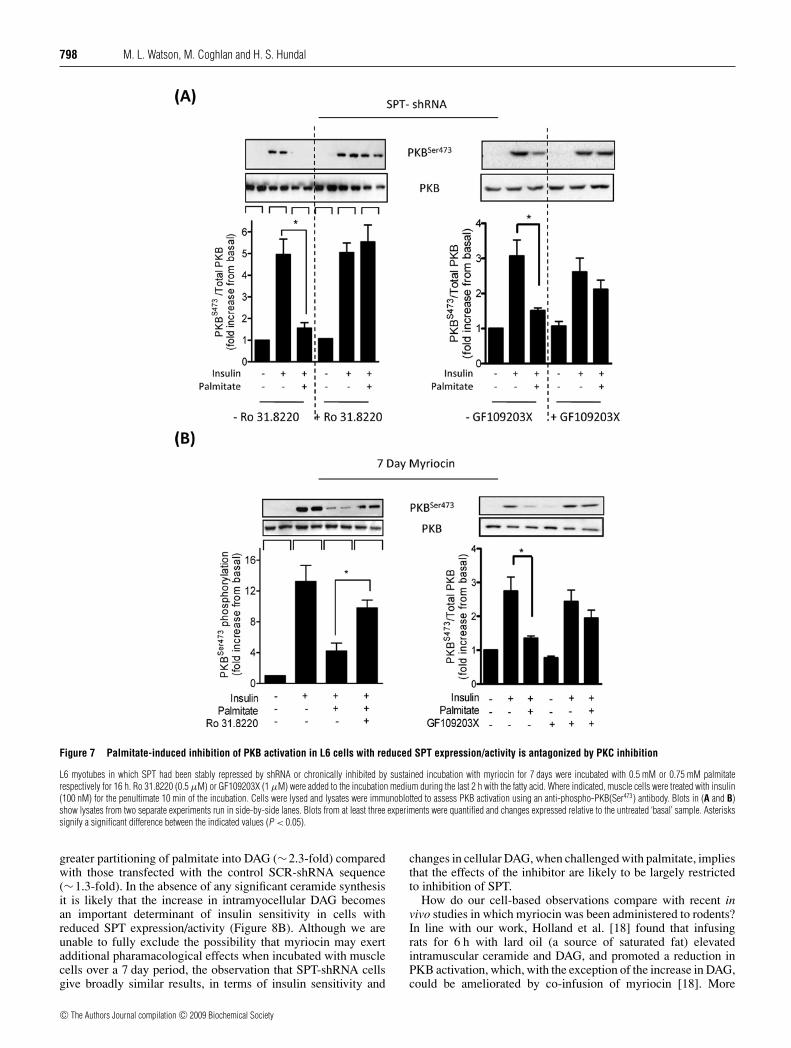
Since PKC inhibition antagonized palmitate-induced IRS1Ser1101 phosphorylation, we subsequently assessed whether thisintervention would enhance IRS1-directed insulin signalling toPKB. Figures 7(A) and 7(B) show that incubation of both SPT-shRNA cells and L6 cells that had been treated chronically(for 7 days) with 10 μM myriocin with Ro 31.8220 orGF109203X (another bisindolemalemide capable of targeting andinhibiting PKC isoforms) led to a significant diminution in theability of palmitate to suppress insulin-stimulated PKB(Ser473)phosphorylation.
DISCUSSION
SPT occupies a critical position with regard to intracellular cer-amide metabolism, catalysing the committed step in the de novosynthesis of ceramide from palmitoyl-CoA and serine [19]. Underphysiological circumstances the intracellular concentration oflipid intermediates generated downstream of SPT activity suchas 3-oxosphinganine, sphinganine and ceramide are normallymaintained at relatively low levels [27]. However, the increasedavailability of circulating NEFAs, in particular palmitate, servesto drive ceramide synthesis, whose cellular accumulation not onlypromotes apoptosis [28,29] but impairs insulin action in tissuessuch as skeletal muscle and adipose tissue [30]. It follows thatpreventing excessive intracellular synthesis of ceramide may havepotentially beneficial consequences with respect to maintainingcellular integrity and insulin sensitivity. Indeed, recent in vivowork from Summers and co-workers [18] strongly suggests thatSPT is likely to be an appropriate molecular target for anti-diabetic therapies based on observations showing that myriocinadministration to rodents ameliorates some of the metabolicdisturbances associated with glucocorticoid-, saturated fat- andobesity-induced insulin resistance.
In an attempt to further understand the role played by SPTin fatty-acid-induced insulin resistance and to assess its value asa potential therapeutic target, the present study investigated theeffects of palmitate on insulin action in muscle cells in which SPTexpression/activity was chronically suppressed. While short-term(24 h) inhibition of SPT with myriocin was found to amelioratethe loss in insulin-stimulated PKB phosphorylation inducedby palmitate-driven ceramide synthesis ([2] and SupplementaryFigure S1), our findings indicate that sustained inhibitionof SPT or shRNA-mediated silencing of the enzyme failsto confer a protective effect against the insulin-desensitizingeffects of palmitate. We postulated that, in the absence ofbeing able to utilize palmitoyl-CoA for ceramide synthesis viathe SPT pathway, palmitate may have been channelled intoother pathways, such as those responsible for DAG synthesis.Consistent with this notion, the present study demonstrates thatmuscle cells harbouring a stable loss in SPT expression exhibit
c© The Authors Journal compilation c© 2009 Biochemical Society
798 M. L. Watson, M. Coghlan and H. S. Hundal
Figure 7 Palmitate-induced inhibition of PKB activation in L6 cells with reduced SPT expression/activity is antagonized by PKC inhibition
L6 myotubes in which SPT had been stably repressed by shRNA or chronically inhibited by sustained incubation with myriocin for 7 days were incubated with 0.5 mM or 0.75 mM palmitaterespectively for 16 h. Ro 31.8220 (0.5 μM) or GF109203X (1 μM) were added to the incubation medium during the last 2 h with the fatty acid. Where indicated, muscle cells were treated with insulin(100 nM) for the penultimate 10 min of the incubation. Cells were lysed and lysates were immunoblotted to assess PKB activation using an anti-phospho-PKB(Ser473) antibody. Blots in (A and B)show lysates from two separate experiments run in side-by-side lanes. Blots from at least three experiments were quantified and changes expressed relative to the untreated ‘basal’ sample. Asteriskssignify a significant difference between the indicated values (P < 0.05).
greater partitioning of palmitate into DAG (∼2.3-fold) comparedwith those transfected with the control SCR-shRNA sequence(∼1.3-fold). In the absence of any significant ceramide synthesisit is likely that the increase in intramyocellular DAG becomesan important determinant of insulin sensitivity in cells withreduced SPT expression/activity (Figure 8B). Although we areunable to fully exclude the possibility that myriocin may exertadditional pharamacological effects when incubated with musclecells over a 7 day period, the observation that SPT-shRNA cellsgive broadly similar results, in terms of insulin sensitivity and
changes in cellular DAG, when challenged with palmitate, impliesthat the effects of the inhibitor are likely to be largely restrictedto inhibition of SPT.
How do our cell-based observations compare with recent invivo studies in which myriocin was been administered to rodents?In line with our work, Holland et al. [18] found that infusingrats for 6 h with lard oil (a source of saturated fat) elevatedintramuscular ceramide and DAG, and promoted a reduction inPKB activation, which, with the exception of the increase in DAG,could be ameliorated by co-infusion of myriocin [18]. More
c© The Authors Journal compilation c© 2009 Biochemical Society

SPT and insulin resistance 799
Figure 8 Schemes outlining the effects of manipulating SPTexpression/activity upon palmitate-induced changes in insulin signalling
(A) Increased provision of palmitate results in enhanced synthesis of ceramide via the SPTpathway, which suppresses insulin-dependent activation of PKB and glucose uptake via amechanism that depends upon activation of atypical PKCs [2,9,11]. Short-term inhibition of SPTameliorates the loss in insulin sensitivity by antagonizing the synthesis and accumulation ofceramide. DAG synthesis is also elevated in response to an increase in palmitate provision, butthis does not disrupt insulin signalling upstream of PKB [2]. (B) Repressing SPT expressionby gene silencing or subjecting the enzyme to sustained inhibition with myriocin alters themetabolic fate of palmitate. The fatty acid is shunted towards greater synthesis of DAG to athreshold that promotes activation of DAG-sensitive PKCs that negatively regulate insulin actionvia increased serine phosphorylation of IRS1.
long-term administration of myriocin was found to improvewhole-body insulin sensitivity in rodent models of glucocorticoid-and obesity-induced insulin resistance. Although the authorsnoted that myriocin treatment markedly reduced tissue ceramide,they did not directly assess the chronic effects of myriocinadministration on intramuscular DAG or insulin action in skeletalmuscle. Therefore it is not implausible that the improved whole-body glucose tolerance observed by these authors in responseto long-term myriocin treatment may partly reflect a greaterefficacy of the drug in tissues other than skeletal muscle.Indeed, the observation that pre-treatment of rats with myriocincompletely prevents glucocorticoid-induced increases in hepaticglucose output, while only being partially effective in blockingdexamethasone-induced insulin resistance in skeletal muscle,is fully consistent with this proposition [18]. Several kinases,including members of the PKC family, have been implicatedin serine phosphorylation of IRS1 on multiple sites including,for example, Ser307, Ser312, Ser612, Ser636/639 and Ser1101, whichare widely accepted to negatively regulate IRS1-directed insulinsignalling [31]. Since DAG accrual has been linked to activation
of novel and conventional PKCs, this family of proteins is likelyto be an important effector of lipid-induced insulin resistance[31]. Indeed, the expression of PKCθ , a novel PKC isoform thatis very abundant in skeletal muscle, has been shown to not onlybe elevated in response to an increase in circulating NEFAs, butactivated by increases in fatty acyl-CoA and DAG [24,32–34].The full array of serine sites that may be targeted by PKCs on IRSproteins still remain poorly defined, but PKCθ has been suggestedto directly promote phosphorylation of Ser1101 [25] and indirectlythat of Ser307 via activation of JNK (c-Jun N-terminal kinase)and IKKβ {IκB [inhibitor of NF-κB (nuclear factor κB)] kinaseβ} [35,36]. The notion that a PKC-mediated increase in IRS1serine phosphorylation may underpin palmitate-induced insulinresistance in cells in which SPT expression/activity has beenreduced is supported by the following observations. First, ananti-phosphoserine antibody detects increased serine phos-phorylation within IRS1 immunoprecipitates prepared from palm-itate-treated SPT-shRNA-transfected L6 myotubes and fromthose chronically treated with myriocin. Secondly, palmitatetreatment of these cells led to activation of PKCθ and enhancedphosphorylation of IRS1 Ser1101; a reported PKCθ target site[25]. It is noteworthy that neither PKCθ activation nor increasedIRS1 Ser1101 were evident in wild-type cells following palmitatetreatment, consistent with the idea that palmitate does notdisrupt IRS1/PI3K association in such cells (Figure 5). Thirdly,palmitate-induced phosphorylation of Ser1101 was sensitive to Ro31.8220, a bisindolylmaleimide that selectively targets novel andconventional PKCs at submicromolar concentrations [37] and, assuch, this inhibitor had a restorative effect on insulin-stimulatedPKB phosphorylation. It is important to stress, however, that sincemuscle cells also express other DAG-sensitive PKCs, such asPKCα and PKCδ [38,39], these are likely to act in concert withPKCθ to promote a net increase in IRS1 serine phosphorylation[40,41]. Since bisindolylmaleimides such as Ro 31.8220 andGF109203X do not discriminate between novel and conventionalPKC isoforms, the ability of these compounds to antagonizepalmitate-induced inhibition of PKB in SPT-shRNA-transfectedcells and those treated with myriocin should, therefore, be viewedin the context of an overall reduction in the activation of DAG-sensitive PKCs.
In addition to ceramide and DAG there is growing evidencelinking components of the innate immune system to lipid-induced insulin resistance. Saturated fatty acids can serve asligands for TLR (Toll-like receptor)-2 and TLR-4, which,when activated, stimulate the pro-inflammatory NF-κB pathway[42,43]. Activation of NF-κB promotes increased expression ofpro-inflammatory cytokines, such as IL-6 (interleukin-6) andTNF-α (tumour necrosis factor-α), which negatively regulateskeletal muscle insulin sensitivity, as seen in rodents placed ona high-fat diet for periods of up to 16 weeks [43,44]. In separatestudies we have observed that palmitate can also induce NF-κBactivation in L6 myotubes, but our findings suggest that insulinresistance induced by the fatty acid is not, at least over the 16 hpalmitate incubation used in the present studies, dependent onNF-κB, given that inhibition of PKCs ameliorate the loss in PKBactivation without any significant reduction in NF-κB activation(C. Lipina and H. Hundal, unpublished work). Nevertheless, giventhat suppression of TLR-4 and NF-κB signalling [44–46] hasbeen suggested to antagonize diet-induced insulin resistance, itis likely that the loss in insulin sensitivity associated with fatty-acid overload involves contribution from diverse pathways, whichexert both spatial and temporal effects upon cellular responsesinfluencing the fidelity of insulin action within tissues such asskeletal muscle [47]. For example, the finding that the insulin-dependent activation of PKB can be acutely down-regulated
c© The Authors Journal compilation c© 2009 Biochemical Society

800 M. L. Watson, M. Coghlan and H. S. Hundal
within 10 min of raising intracellular ceramide, using a cell-permeant analogue of this sphingolipid [4], supports the idea thatceramide synthesis from palmitate may represent a key early eventinitiating fatty-acid-induced insulin resistance. However, insulinsensitivity will also be strongly influenced by responses that maytemporally cause a lag in the effects associated with an increasein cell ceramide and DAG, such as changes in gene expressioninduced in response to pro-inflammatory (NF-κB and cytokine)signalling that will affect important cellular functions such asinsulin signalling, nutrient uptake and mitochondrial function[47].
In summary, the findings of the present study demonstratethat ceramide synthesis from palmitate is critically dependentupon SPT and that increased flux through this enzyme makesan important contribution to the insulin-desensitizing effects ofpalmitate. Ceramide generated via this pathway induces a targetedinhibition of PKB by a mechanism that has been shown previouslyto involve activation of atypical PKCs [2,9,48]. The importanceof ceramide generated via this pathway is underscored by thefinding that short-term inhibition of SPT helps alleviate theinsulin-desensitizing effects of palmitate. However, a sustainedloss or reduction in SPT expression/activity, although reducing thecapacity for generating ceramide from palmitate, has the effect ofdiverting use of the fatty acid towards greater synthesis of DAG,which then becomes a more important determinant of insulinsensitivity through its ability to activate DAG-sensitive PKCs thatsuppress IRS-directed insulin signalling. Consequently, althoughtargeting SPT with inhibitors improves insulin sensitivity bothin vitro (the present study and [2,49]) and in vivo [18] inthe short-term, the perceived long-term benefits may need tobe carefully assessed based on the impact that SPT inhibitionhas upon accumulation of other fatty-acid-derived metaboliteswith the potential to promote insulin resistance within skeletalmuscle.
ACKNOWLEDGEMENTS
We thank Dr Nikolaos Dimopoulos, Emma Cwiklinski and Charlotte Green for assistancewith some of the experimental work reported and Dr Chris Lipina for providing helpfulcomments on the manuscript prior to submission.
FUNDING
This work was supported by the Biotechnology and Biological Sciences ResearchCouncil/Industrial (AstraZeneca) CASE Quota Award [grant number BBS/2005/12337];the European Commission [contract LSHM-CT-20004-005272]; Diabetes Research &Wellness Foundation [open funding programme]; and Diabetes UK [grant numberBDA:RD07/0003405].
REFERENCES
1 Bruce, C. R., Kriketos, A. D., Cooney, G. J. and Hawley, J. A. (2004) Disassociation ofmuscle triglyceride content and insulin sensitivity after exercise training in patients withType 2 diabetes. Diabetologia 47, 23–30
2 Powell, D. J., Turban, S., Gray, A., Hajduch, E. and Hundal, H. S. (2004) Intracellularceramide synthesis and protein kinase Cζ activation play an essential role inpalmitate-induced insulin resistance in rat L6 skeletal muscle cells. Biochem. J. 382,619–629
3 Dimopoulos, N., Watson, M., Sakamoto, K. and Hundal, H. S. (2006) Differential effects ofpalmitate and palmitoleate on insulin action and glucose utilization in rat L6 skeletalmuscle cells. Biochem. J. 399, 473–481
4 Hajduch, E., Balendran, A., Batty, I. H., Litherland, G. J., Blair, A. S., Downes, C. P. andHundal, H. S. (2001) Ceramide impairs the insulin-dependent membrane recruitment ofprotein kinase B leading to a loss in downstream signalling in L6 skeletal muscle cells.Diabetologia 44, 173–183
5 Chavez, J. A. and Summers, S. A. (2003) Characterizing the effects of saturated fatty acidson insulin signaling and ceramide and diacylglycerol accumulation in 3T3-L1 adipocytesand C2C12 myotubes. Arch. Biochem. Biophys. 419, 101–109
6 Turinsky, J., O’Sullivan, D. M. and Bayly, B. P. (1990) 1,2-Diacylglycerol and ceramidelevels in insulin-resistant tissues of the rat in vivo. J. Biol. Chem. 265,16880–16885
7 Dobrzyn, A. and Gorski, J. (2002) Ceramides and sphingomyelins in skeletal muscles ofthe rat: content and composition. Effect of prolonged exercise. Am. J. Physiol.Endocrinol. Metab. 282, E277–E285
8 Straczkowski, M., Kowalska, I., Nikolajuk, A., Dzienis-Straczkowska, S., Kinalska, I.,Baranowski, M., Zendzian-Piotrowska, M., Brzezinska, Z. and Gorski, J. (2004)Relationship between insulin sensitivity and sphingomyelin signaling pathway in humanskeletal muscle. Diabetes 53, 1215–1221
9 Powell, D. J., Hajduch, E., Kular, G. and Hundal, H. S. (2003) Ceramide disables3-phosphoinositide binding to the pleckstrin homology domain of protein kinase B(PKB)/Akt by a PKCζ -dependent mechanism. Mol. Cell. Biol. 23, 7794–7808
10 Summers, S. A., Garza, L. A., Zhou, H. and Birnbaum, M. J. (1998) Regulation ofinsulin-stimulated glucose transporter GLUT4 translocation and Akt kinase activity byceramide. Mol. Cell. Biol. 18, 5457–5464
11 Bourbon, N. A., Sandirasegarane, L. and Kester, M. (2002) Ceramide-induced inhibitionof Akt is mediated through protein kinase Cζ : implications for growth arrest.J. Biol. Chem. 277, 3286–3292
12 Salinas, M., Lopez-Valdaliso, R., Martin, D., Alvarez, A. and Cuadrado, A. (2000)Inhibition of PKB/Akt1 by C2-ceramide involves activation of ceramide- activated proteinphosphatase in PC12 cells. Mol. Cell. Neurosci. 15, 156–169
13 Mathias, S., Pena, L. A. and Kolesnick, R. N. (1998) Signal transduction of stress viaceramide. Biochem. J. 335, 465–480
14 Doornbos, R. P., Theelen, M., van der Hoeven, P. C., van Blitterswijk, W. J., Verkleij, A. J.and van Bergen en Henegouwen PM, A. J. (1999) Protein kinase Cζ is a negativeregulator of protein kinase B activity. J. Biol. Chem. 274, 8589–8596
15 Konishi, H., Kuroda, S. and Kikkawa, U. (1994) The pleckstrin homology domain of RACprotein kinase associates with the regulatory domain of protein kinase Cζ .Biochem. Biophys. Res. Commun. 205, 1770–1775
16 Mao, M., Fang, X., Lu, Y., Lapushin, R., Bast, J. R. and Mills, G. B. (2000) Inhibition ofgrowth-factor-induced phosphorylation and activation of protein kinase B/Akt by atypicalprotein kinase C in breast cancer cells. Biochem. J. 352, 475–482
17 Hodgkinson, C. P., Sale, E. M. and Sale, G. J. (2002) Characterization of PDK2 activityagainst protein kinase Bγ . Biochemistry 41, 10351–10359
18 Holland, W. L., Brozinick, J. T., Wang, L. P., Hawkins, E. D., Sargent, K. M., Liu, Y.,Narra, K., Hoehn, K. L., Knotts, T. A. et al. (2007) Inhibition of ceramide synthesisameliorates glucocorticoid-, saturated-fat-, and obesity-induced insulin resistance.Cell Metab. 5, 167–179
19 Hanada, K. (2003) Serine palmitoyltransferase, a key enzyme of sphingolipid metabolism.Biochim. Biophys. Acta 1632, 16–30
20 Hornemann, T., Richard, S., Rutti, M. F., Wei, Y. and von Eckardstein, A. (2006) Cloningand initial characterization of a new subunit for mammalian serine-palmitoyltransferase.J. Biol. Chem. 281, 37275–37281
21 Hajduch, E., Alessi, D. R., Hemmings, B. A. and Hundal, H. S. (1998) Constitutiveactivation of protein kinase Bα (PKBα) by membrane targeting promotes glucose andSystem A amino acid transport, protein synthesis and GSK3 inactivation in L6 musclecells. Diabetes 47, 1006–1013
22 Bradford, M. M. (1976) A rapid and sensitive method for the quantitation of microgramquantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72,248–254
23 Holleran, W. M., Williams, M. L., Gao, W. N. and Elias, P. M. (1990) Serine-palmitoyltransferase activity in cultured human keratinocytes. J. Lipid Res. 31, 1655–1661
24 Haasch, D., Berg, C., Clampit, J. E., Pederson, T., Frost, L., Kroeger, P. and Rondinone,C. M. (2006) PKCθ is a key player in the development of insulin resistance.Biochem. Biophys. Res. Commun. 343, 361–368
25 Li, Y., Soos, T. J., Li, X., Wu, J., Degennaro, M., Sun, X., Littman, D. R., Birnbaum, M. J.and Polakiewicz, R. D. (2004) Protein kinase Cζ inhibits insulin signaling byphosphorylating IRS1 at Ser1101. J. Biol. Chem. 279, 45304–45307
26 Standaert, M. L., Galloway, L., Karnam, P., Bandyopadhyay, G., Moscat, J. and Farese,R. V. (1997) Protein kinase C-ζ as a downstream effector of phosphatidylinositol 3-kinaseduring insulin stimulation in rat adipocytes: potential role in glucose transport.J. Biol. Chem. 272, 30075–30082
27 Zheng, W., Kollmeyer, J., Symolon, H., Momin, A., Munter, E., Wang, E., Kelly, S.,Allegood, J. C., Liu, Y., Peng, Q. et al. (2006) Ceramides and other bioactive sphingolipidbackbones in health and disease: lipidomic analysis, metabolism and roles in membranestructure, dynamics, signaling and autophagy. Biochim. Biophys. Acta 1758,1864–1884
c© The Authors Journal compilation c© 2009 Biochemical Society

SPT and insulin resistance 801
28 Paumen, M. B., Ishida, Y., Muramatsu, M., Yamamoto, M. and Honjo, T. (1997) Inhibitionof carnitine palmitoyltransferase I augments sphingolipid synthesis and palmitate-induced apoptosis. J. Biol. Chem. 272, 3324–3329
29 Shimabukuro, M., Zhou, Y. T., Levi, M. and Unger, R. H. (1998) Fatty acid-induced β-cellapoptosis: a link between obesity and diabetes. Proc. Natl. Acad. Sci. U.S.A. 95,2498–2502
30 Summers, S. A. (2006) Ceramides in insulin resistance and lipotoxicity. Prog. Lipid Res.45, 42–72
31 Gual, P., Le Marchand-Brustel, Y. and Tanti, J. F. (2005) Positive and negative regulationof insulin signaling through IRS-1 phosphorylation. Biochimie 87, 99–109
32 Qu, X., Seale, J. P. and Donnelly, R. (1999) Tissue and isoform-selective activation ofprotein kinase C in insulin-resistant obese Zucker rats: effects of feeding. J. Endocrinol.162, 207–214
33 Griffin, M. E., Marcucci, M. J., Cline, G. W., Bell, K., Barucci, N., Lee, D., Goodyear, L. J.,Kraegen, E. W., White, M. F. and Shulman, G. I. (1999) Free fatty acid-induced insulinresistance is associated with activation of protein kinase Cθ and alterations in the insulinsignaling cascade. Diabetes 48, 1270–1274
34 Itani, S. I., Zhou, Q., Pories, W. J., Macdonald, K. G. and Dohm, G. L. (2000) Involvementof protein kinase C in human skeletal muscle insulin resistance and obesity. Diabetes 49,1353–1358
35 Gao, Z., Hwang, D., Bataille, F., Lefevre, M., York, D., Quon, M. J. and Ye, J. (2002) Serinephosphorylation of insulin receptor substrate 1 by inhibitor κB kinase complex.J. Biol. Chem. 277, 48115–48121
36 Gao, Z., Zhang, X., Zuberi, A., Hwang, D., Quon, M. J., Lefevre, M. and Ye, J. (2004)Inhibition of insulin sensitivity by free fatty acids requires activation of multiple serinekinases in 3T3-L1 adipocytes. Mol. Endocrinol. 18, 2024–2034
37 Standaert, M. L., Bandyopadhyay, G., Perez, L., Price, D., Galloway, L., Poklepovic, A.,Sajan, M. P., Cenni, V., Sirri, A., Moscat, J. et al. (1999) Insulin activates protein kinasesCζ and Cλ by an autophosphorylation-dependent mechanism and stimulates theirtranslocation to GLUT4 vesicles and other membrane fractions in rat adipocytes.J. Biol. Chem. 274, 25308–25316
38 Caruso, M., Miele, C., Oriente, F., Maitan, A., Bifulco, G., Andreozzi, F., Condorelli, G.,Formisano, P. and Beguinot, F. (1999) In L6 skeletal muscle cells, glucose inducescytosolic translocation of protein kinase Cα and trans-activates the insulin receptorkinase. J. Biol. Chem. 274, 28637–28644
39 Horovitz-Fried, M., Brutman-Barazani, T., Kesten, D. and Sampson, S. R. (2008) Insulinincreases nuclear protein kinase Cδ in L6 skeletal muscle cells. Endocrinology 149,1718–1727
40 Cipok, M., ga-Mizrachi, S., Bak, A., Feurstein, T., Steinhart, R., Brodie, C. and Sampson,S. R. (2006) Protein kinase Cα regulates insulin receptor signaling in skeletal muscle.Biochem. Biophys. Res. Commun. 345, 817–824
41 Nawaratne, R., Gray, A., Jorgensen, C. H., Downes, C. P., Siddle, K. and Sethi, J. K.(2006) Regulation of insulin receptor substrate 1 pleckstrin homology domain by proteinkinase C: role of serine 24 phosphorylation. Mol. Endocrinol. 20, 1838–1852
42 Senn, J. J. (2006) Toll-like receptor-2 is essential for the developmentof palmitate-induced insulin resistance in myotubes. J. Biol. Chem. 281,26865–26875
43 Shi, H., Kokoeva, M. V., Inouye, K., Tzameli, I., Yin, H. and Flier, J. S. (2006) TLR4 linksinnate immunity and fatty acid-induced insulin resistance. J. Clin. Invest. 116,3015–3025
44 Tsukumo, D. M., Carvalho-Filho, M. A., Carvalheira, J. B., Prada, P. O., Hirabara, S. M.,Schenka, A. A., Araujo, E. P., Vassallo, J., Curi, R., Velloso, L. A. and Saad, M. J. (2007)Loss-of-function mutation in Toll-like receptor 4 prevents diet-induced obesity andinsulin resistance. Diabetes 56, 1986–1998
45 Radin, M. S., Sinha, S., Bhatt, B. A., Dedousis, N. and O’Doherty, R. M. (2008) Inhibitionor deletion of the lipopolysaccharide receptor Toll-like receptor-4 confers partialprotection against lipid-induced insulin resistance in rodent skeletal muscle. Diabetologia51, 336–346
46 Sinha, S., Perdomo, G., Brown, N. F. and O’Doherty, R. M. (2004) Fatty acid-inducedinsulin resistance in L6 myotubes is prevented by inhibition of activation and nuclearlocalization of nuclear factor κB. J. Biol. Chem. 279, 41294–41301
47 Hirabara, S. M., Silveira, L. R., Abdulkader, F., Carvalho, C. R., Procopio, J. and Curi, R.(2007) Time-dependent effects of fatty acids on skeletal muscle metabolism.J. Cell. Physiol. 210, 7–15
48 Stratford, S., Dewald, D. B. and Summers, S. A. (2001) Ceramide dissociates3′-phosphoinositide production from pleckstrin homology domain translocation.Biochem. J. 354, 359–368
49 Pickersgill, L., Litherland, G. J., Greenberg, A. S., Walker, M. and Yeaman, S. J. (2007)Key role for ceramides in mediating insulin resistance in human muscle cells.J. Biol. Chem. 282, 12583–12589
Received 5 June 2008/30 September 2008; accepted 16 October 2008Published as BJ Immediate Publication 16 October 2008, doi:10.1042/BJ20081149
c© The Authors Journal compilation c© 2009 Biochemical Society
Biochem. J. (2009) 417, 791–801 (Printed in Great Britain) doi:10.1042/BJ20081149
SUPPLEMENTARY ONLINE DATAModulating serine palmitoyl transferase (SPT) expression and activityunveils a crucial role in lipid-induced insulin resistance in rat skeletalmuscle cellsMaria L. WATSON*, Matthew COGHLAN† and Harinder S. HUNDAL*1
*Division of Molecular Physiology, Sir James Black Centre, College of Life Sciences, University of Dundee, Dundee, DD1 5EH, Scotland, U.K., and †AstraZeneca, CVGI, Alderley Park,Macclesfield, Cheshire SK10 4TG, U.K.
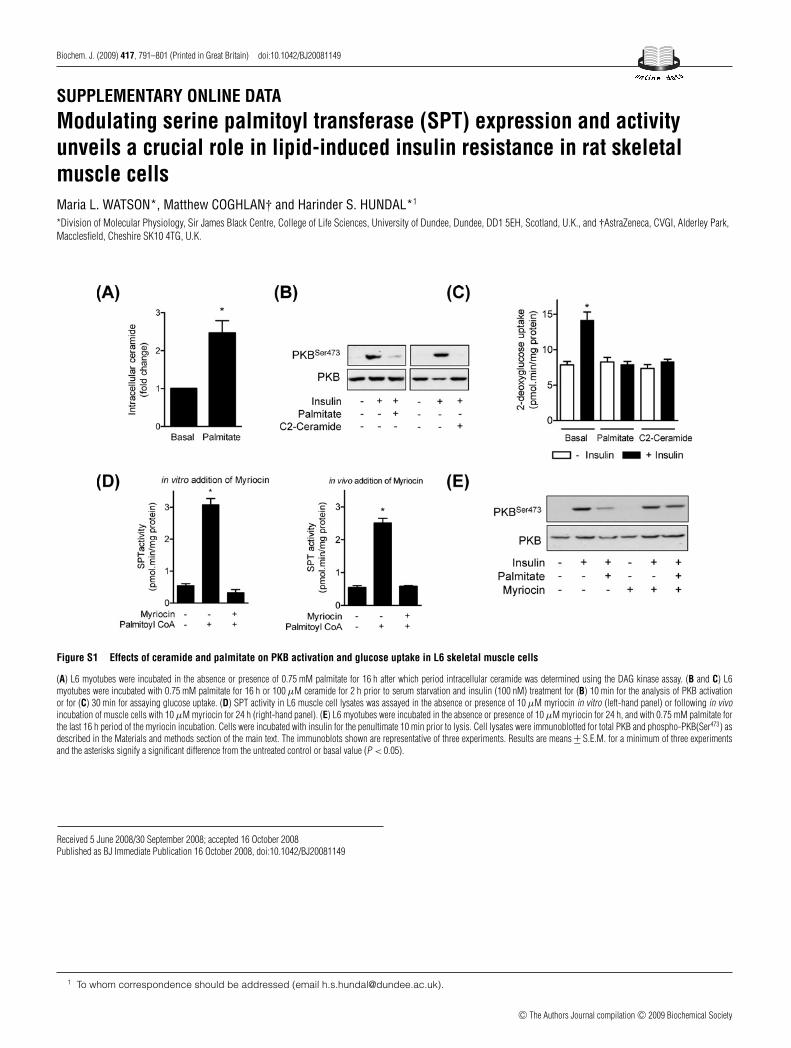
Figure S1 Effects of ceramide and palmitate on PKB activation and glucose uptake in L6 skeletal muscle cells
(A) L6 myotubes were incubated in the absence or presence of 0.75 mM palmitate for 16 h after which period intracellular ceramide was determined using the DAG kinase assay. (B and C) L6myotubes were incubated with 0.75 mM palmitate for 16 h or 100 μM ceramide for 2 h prior to serum starvation and insulin (100 nM) treatment for (B) 10 min for the analysis of PKB activationor for (C) 30 min for assaying glucose uptake. (D) SPT activity in L6 muscle cell lysates was assayed in the absence or presence of 10 μM myriocin in vitro (left-hand panel) or following in vivoincubation of muscle cells with 10 μM myriocin for 24 h (right-hand panel). (E) L6 myotubes were incubated in the absence or presence of 10 μM myriocin for 24 h, and with 0.75 mM palmitate forthe last 16 h period of the myriocin incubation. Cells were incubated with insulin for the penultimate 10 min prior to lysis. Cell lysates were immunoblotted for total PKB and phospho-PKB(Ser473) asdescribed in the Materials and methods section of the main text. The immunoblots shown are representative of three experiments. Results are means +− S.E.M. for a minimum of three experimentsand the asterisks signify a significant difference from the untreated control or basal value (P < 0.05).
Received 5 June 2008/30 September 2008; accepted 16 October 2008Published as BJ Immediate Publication 16 October 2008, doi:10.1042/BJ20081149
1 To whom correspondence should be addressed (email [email protected]).
c© The Authors Journal compilation c© 2009 Biochemical Society





























