Embed Size (px)
Citation preview

H. Schroer u. E. Zimmermann Molekularbiologische Aspekte der Blutgerinnungsvorgang e I. Einfuhrung
Der wahrend der Phylogenese als Neuer- werbung in Erscheinung tretende geschlos- sene Blutkreislauf verlangt eine Kompen- sation des entstehenden Stromungsdruckes der bewegten Fliissigkeit. Unter diesem Aspekt darf das Kreislaufsystem als die mechanische Kopplung einer leistungsstar- ken Pumpe mit einem geschlossenen, me- chanisch-flexiblen Schlauchsystem gesehen werden. Die teils pulsierend, teils konti- nuierlich stromende Fliissigkeit ist mit sich anderndem Gefafidurchmesser wechseln- den laminaren Scherkrafien ausgesetzt, die ihrerseits von der Stromungsgeschwindig- keit und der Flexibilitat der Gefafiwan- dung abhangig sind. Die biologisch not- wendige Sicherung der Funktion eines ge- schlossenen Kreislaufs schliei3t ein, dai3 Fehlstellen, die aufgrund nachlassender Flexibilitat oder aufierer mechanischer Ein- wirkungen entstanden sind, durch ein in der zirkulierenden Fliissigkeit existieren- des System ,,repariert" werden konnen. Der fur eine solche Notfallsituation zur Verfugung stehende "repair mechanism" umfafit neben den hamostatischen Funk- tionen der Blutplattchen alle enzymati- schen Vorgange zur Umwandlung des ei- gentlichen Gerinnungssubstrats Fibrinogen in Fibrin, einschliei3lich der Bereitstellung des dazu notwendigen Enzyms Thrombin. Die Vervollstandigung des biologisch so wichtigen Regelkreises zur Aufrechterhal- tung eines geshlossenen Kreislaufes bedarf eines weiteren Mechanismus, um entstan- dene Gerinnungsprodukte abzubauen und die enzymatische Aktivitat aller beteilig- ten Faktoren auf das Normalmafi zu redu- zieren. Dieses System zur Kompensation des Gerinnungspotentials auf den physio- logischen Normalzustand wird durch den Begriff ,,Fibrinolyse" umschrieben.
Es lafit sich anschaulich vorstellen, wie das Zirkulieren von Transport- und Aus- tauschfliissigkeit im Gefai3system unter Zuhilfenahme von korpereigenen Sub- stanzen aufrecht erhalten und gegen mog- liche Storungen abgesichert wird. Fur alle dam notwendigen Faktoren mufite ein ge- steuerter Synthese-Mechanismus geschaf- fen werden, der die Bereitstellung von korpuskularen Bestandteilen, inaktiven Vorstufen sowie hochmolekularen und
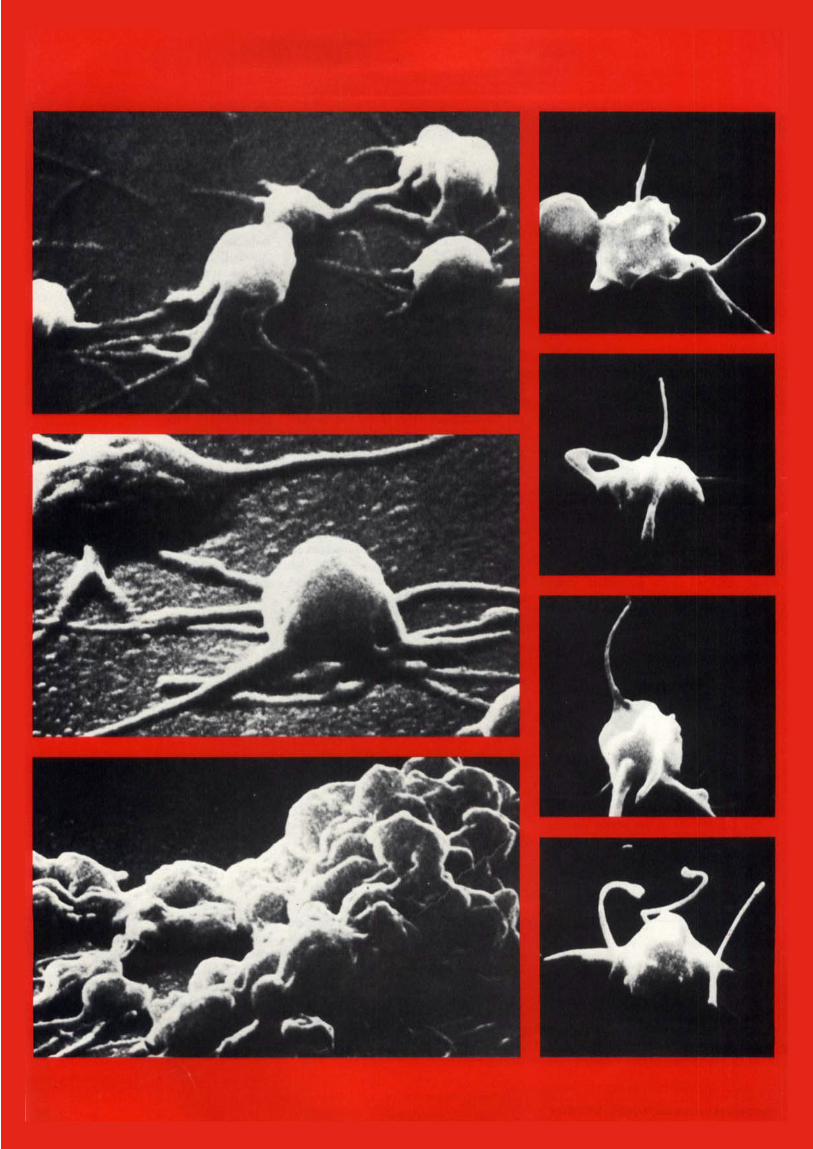
Elektronenmikroskopische Aufnahme von polymerisierbaren Gerinnungssubstanzen fixierten und dehydrierten Blutplattchen gewahrleistet. Die genetische Information (nach Murphey). zur Bildung des Fibrinogens mufite in das
Genom des hoher entwickelten Organis- mus aufgenommen werden. Weiterhin war es erforderlich, Enzyme zu entwickeln, die das losliche Fibrinogen in sein unlosliches Polymerisationsprodukt, namlich Fibrin, umwandeln konnen. Soweit es sich dabei um Proteine handelt, miissen auch fur alle diese beteiligten Aktivatoren und Inhibi- toren des Prothrombin-Thrombin-Systems bestimmte Gen-Loci vorhanden sein. Mit anderen Worten: Der Blutkreislauf ist nicht nur streng physikalischen Gesetzen unterworfen, sondern die Erhaltung seiner Funktion erfordert das Zusammenwirken von genetischen Informationen und be- darfsgerechter Synthese von Enzymen und polymerisierbaren korpereigenen Substan- Zen. Fur den zu erwartenden physiologi- schen und pathophysiologischen Notfall miissen inaktive Enzymvorstufen in aus- reichenden Mengen im Blut bestandig zir- kulieren und in raumlichem Kontakt zu der Substanz stehen, die mit ihrer Hilfe zum Polymerisationsprodukt Fibrin um- gesetzt werden kann.
Die Betrachtung von Einzelelementen die- ses physiologischen Vorganges kann be- stenfalls zur Charakterisierung von ,, einem" Enzym in ,,einer" Teilreaktion der multipotenten Gerinnung dienen. Vielleicht erklfrt sich gerade aus diesen Beschrankungen das Auftreten von wider- spriichlichen Theorien oder alteingefahre- nen Lehrmeinungen, die sich beim heuti- gen Wissensstand eher hemmend auf die Gesamtentwicklung auswirken.
Die vorliegende Arbeit stellt einen Ver- such dar, die herkommlichen physikalisch- chemischen und physiologischen Vorstel- lungen mit modernen naturwissenschaftli- chen Erkenntnissen zu einem sinnvollen Ganzen zu verbinden.
11. Fibrinogenmetabolismus und Fibrin- polymerisat ion
Die molekularbiologische Betrachtung des Gerinnungsvorganges sei zunachst unmit- telbar auf das eigentliche ,,gerinnende" Substrat, das Fibrinogen, bezogen. Dies erscheint - unter Beriicksichtigung der vorliegenden experimentellen Fakten - logisch gerechtfertigt, denn unser Wissen iiber die Aktivierungsvorgange zur BiI- dung des Thrombins basiert weitgehend auf indirekten Schlufifolgerungen, die aus Beobachtungen zur Reaktionskinetik ein-
Pharmazie in unserer Zeitl2. Jahrg. 19731Nr. 3 79

zelner Faktoren oder Wirkungsgruppen hergeleitet sind. Wiirde man die Aussage der einschlagigen Literatur zu Rate ziehen, so ware auch von der Didaktik der Be- trachtung her das Voranstellen des Fibri- nogens empfehlenswert, da in den meisten Fallen die vereinfachende Schluflfolgerung aufgenommen wurde, die Umwandlung des Fibrinogens zu Fibrin sei hinreichend erforscht und beschrieben. Die folgenden Ausfuhrungen werden jedoch zeigen, dai3 gerade diese Endreaktion weitaus kompli- zierter verlauft als bisher angenommen werden konnte.
Die Bezeichnung ,,Fibrinogen" zur Cha- rakterisierung eines loslichen Proteins, das nach Eingriff des Thrombins zu unlosli- chem und polymerisiertem Fibrin wird, 1ai3t vermuten, dai3 wir es zumindest bei dem gerinnungsfahigen Stoff des Blutes mit einem einheitlichen und einwandfrei charakterisierbaren Molekul zu tun haben. Die Untersuchungen von Fibrinogenpra- paraten vershiedenster Herstellungsver- fahren mit sfulenchromatographischen oder elektrophoretischen Verfahrenstech- niken zeigen indessen, dai3 ,,das" Fibrino- gen ein heterogenes Gemisch von wahr- scheinlich gleichen, aber in unterschiedli- chen Mengenverhaltnissen assoziierten Un- tereinheiten (Monomeren?) darstellt. Die einzelnen Praparationsverfahren beruhen auf der Loslichkeitsdiff erenz dieses Prote- inkomplexes in Gemischen von Wasser rnit organischen Losungsmitteln und der er- niedrigten Loslichkeit nach Zugabe von Salzen oder niedermolekularen Substan- Zen mit salzahnlichem Charakter. Allen diesen Praparaten ist gemeinsam, dai3 sie sich in verschiedene Unterfraktionen auf- teilen lassen und im allgemeinen ein mitt- leres Molgew. von 340.000 bis zu 360.000 besitzen (Abbildung la, b, c).
Die funktionelle Bedeutung dieser ,,Fibri- nogenkomplexe" ist nicht allein in ihrer Substratfunktion fur das Thrombin zu se- hen. Bei Zentrifugationsversuchen in Sac- charose-Dichtegradienten sedimentiert ein schwererer Anteil des Fibrinogens rnit ei- nem mittleren Molgew. von 1-2 Millio- nen, der zwischen 3 und 5 % der Gesamt- menge ausmacht. Die Existenz solcher Polymeren rechtfertigt die Annahme, daf3 das Fibrinogen aufgrund seiner Assozia- tionsfahigkeit von Untereinheiten zu hochmolekularen Komplexen in den wand- nahen laminaren Stromungsschichten des
Gefai3systems die Funktion der GefBi3- wandabdichtung durch zirkulierende zwei-' dimensionale Vernetzungsprodukte aus- uben kann (Abbildung 2).
Ein sehr starkes experimentelles Argument fur die Existenz von Fibrinogenkomple- xen erbrachte die Analyse der N-termina- len, cysteinreichen Peptide durch B. Blom- back, der die Verkniipfung von Fibrino- gen-Peptidketten durch den Nachweis von intramolekular vernetzten Endpeptiden be- statigt (personl. Mitteilung).
Diese Befunde werfen folgerichtig die Frage nach der dreidimensionalen Struktur des Fibrinogenmolekiils auf, fur die mehrere Modellvorstellungen entworfen wurden. Das Fibrinogen ist ein globulinahnliches Glycoproteid rnit einem Kohlenhydratan- teil von ungefahr 3 %, das im allgemeinen als ein Dimer angesehen wird. Obwohl Ca- pet-Anronini und Gunand [l] in einer neueren Arbeit darauf hinweisen, dai3 der Dimeren-Komplex bei 37" instabil ist und in seine Monomeren zu dissoziieren vermag, bestehen noch erhebliche Zweifel an dem Typus der Monomeren - Verkniipfung.
Trotzdem sollte der Monomeren-Dissozia- tion schon allein im Hinblick auf ihre mogliche physiologische Bedeutung mehr Beachtung geschenkt werden; unter den Zirkulationsbedingungen der hoher ent- wickelten Sauger rnit einer Korpertempe- ratur von eben 37" wurde namlich der ge- samte Anteil des Fibrinogens in Form von Monomeren mit einem mittleren Moleku- largewicht von 180.000 vorliegen. Durch Disulfidgruppen konnten dann verschie- den groi3e Assoziate einer Untereinheit, namlich des vorhandenen Monomers, fur verschiedene Bedarfsfalle im Rahmen der Kreislauffunktionen gebildet werden.
Verlassen wir diese hypothetische Betrach- tung und wenden wir uns dem mehrfach erharteten Befund zu, dai3 das Fibrinogen- Monomer aus drei Peptidketten (a, j3, y ) besteht, die vor allem im N-terminalen Be- reich uber Disulfidbrucken miteinander verbunden sind. Bei einem durchschnittli- chen Molekulargewicht des Fibrinogens von 340.000 miissen sie aus ungeffhr 900 Aminosauren (pro Peptidkette) gebildet sein (Abbildung 3).
Der in Abbildung 3 wiedergegebene, stark schematisierte Aufbau des Fibrinogenmo- lekuls entspricht naturlich nicht seiner tat- sachlich existierenden dreidimensionalen Struktur. Diese ist weitgehend unbekannt und Iai3t fur Modellvorstellungen einen breiten Raum.
Das in neuester Zeit von Koppel[2] entwik- kelte Modell beruht auf der Annahme, dai3 die im Fibrinogen-Dimer existenten sechs Peptidketten einen Pentagondodekaeder in der Weise bilden, dai3 jede Peptidkette im Zickzack den Umfang des Raummodells umlaufl. In den Kantenbereichen sollen je zwei umlaufende helicale Peptidketten par- allel zueinander angeordnet sein, wahrend die Ecken von drei in diesen Bereichen li- nearen Polypeptiden gebildet werden. Ver- gleichen wir dieses Modell mit der Beobach- tung, dai3 Fibrinogen bei Temperaturen um 37" in Monomere dissoziiert, so werden wir auf erhebliche Interpretationsschwierigkei- ten stoaen, wahrend die Bildung von Asso- ziaten aus gleichen Untereinheiten zur Bil- dung von linearen oder zweidimensionalen Molekulaggregaten danach formal moglich ware. Einwande gegen dieses Modell resul- tieren aus den Analysendaten Blombacks, der eindeutig nachweisen konnte, dai3 35 o/o der SS-Brucken im N-terminalen Bereich des Molekuls anzutreffen sind, wahrend
80 Pharrnazie in unserer ZeitJ2. Jahrg. 19731Nr. 3

Links : Abb. 1. Elektropherogramme von Fibri- nogenpraparaten in Polyacrylamid. Ver- suchsbedingungen: Acrylamidkonzentra- tion 5 yo, 0,1 M Phosphatpuffer, p H 8,3; Feldstarke, Stromstarke und Laufdauer in allen drei Rohrchen identisch. a) Fibrino- gen nach Ware et al. [6]; b) Fibrinogen nach Blombick [7]; c) Fibrinogen nach Mosesson und Sherry [8].
Rechts: Abb. 2. Sedimentation von Fibrinogen (Fraktion Cohn I) in linearen Saccharose- gradienten. Versuchsbedingungen: 0,l M Natriumphosphat, p H 7,2, rnit 5 - 40 O h
Saccharose. Zentrifugation: 25 000 RPM, Rotortyp SW, 6 x 16,5 ml; MSE, London, England. Laufdauer: 40 Stunden.
Abb. 3. Schematische Darstellung der Bil- dung des Fibrinogen-Dimers.
Abb. 4. Schematische Darstellung der Fi- brinogen-Umwandlung zu intermedifr auftretenden Fibrinmonomeren (die Di- sulfidbrucken zwischen den Ketten sind hier in der Zeichnung weggelassen).
I L
t 0'05
2 i- x Y;
10 20 30 Rohrchen-Nr. -
a -Kette MG: 600.000
SH SH + p-Kette
+
y-Kette w SH
1 Monomer MG: 180.000 *
"-$ 1
I I I I s-s s-s s-s s-s ~.
Dimer I--- Ekhoo- s-s s-s 340.000
Teilpolyrnerisation I
das Dodekaedermodell mit einer Disulfid- briicke pro Kante rechnet, um das Molekiil stabilisieren zu konnen. Aui3erdem setzt das Model1 voraus, dai3 die einzelnen Polypep- tidketten zu 100 Yo in der a-Helixkonfigu- ration vorliegen, wahrend Untersuchungeii der optischen Rotationsdispersion nur auf einen Anteil von 30 % schlieBen lassen.
Die Betrachtung der Funktion des Fibrino- gens als Substrat des Thrombins fiihrt uns wieder zuriick in den Bereich des physiolo- gischen Geschehens; die Mechanismen nam- lich, die zur Aktivierung des Thrombins fiihren, enden mit der thrombinbedingten Auslosung der Fibrinpolymerisation und der Bildung von unloslichen Gerinnseln oder Thromben, deren Ausmai3 und Loka- lisation allein iiber den Grad der damit verbundenen Zirkulationsbeeintrachtigung entscheiden. Die Fibrinogenkonversion wird durch die proteolytische Aktivitat des Thrombins ausgelost, die zur Abtrennung der FibrinopeptideA und B aus den entspre- chenden Polypeptidketten in der ,,Vor- wartsphase" der Gerinnung - d. h. wah- rend des fibrinoplastischen Gerinnungsab- laufes - fiihrt. Die Freisetzung solcher kurzkettiger artspezifischer Peptide aus 14- 21 Aminosauren geht der Bildung von ho- her molekularen Fibrinmonomeren mit ei- nem Molgew. von 4.95 106 und einer Lange von 4000 A voraus. Wenn wir dem Fibrinogen ein mittleres Molgew. von un- gefahr 340.000 zuschreiben und eine line- are Anordnung beider Monomere in einer Achse annehmen, dann kann die gesamte Lange des dimeren Molekiils nur einige Hundert A betragen. Molekulargewichte um 5 Millionen, wie im Falle des Fibrinpo- lymers berichtet, konnen dann nur auf eine intermediar auftretende lineare Polymeri- sation um die A- und B-Peptide verkurzter Fibrinogenmonomerer zuriickzufiihren sein (Abbildung 4).
Die Freisetzung des Fibrinopeptids A verlauR bei diesem Vorgang mit groaerer Geschwindigkeit als die Freisetzung des Fibrinopeptids B, wahrend die Gesamtre- aktion zur Bildung des Fibrinmonomers eine Reaktion 1. Ordnung ist. Sie erst er- moglicht die sich anschliei3ende Aggrega- tion zum raumlich vernetzten Fibringe- rinnsel. Mit Hilfe eines fibrinstabilisieren- den Enzyms werden anschliegend im Sinne einer Transpeptidierung spezifische Ami- nogruppen des Lysins mit Asparagin- und Glutaminresten verkniipfl und das be-
Pharmazie in unserer Zeit/2. Iahrg. 19731Nr. 3 81

kannte unlosliche Fibringerinnsel kom- pakt gefaltet und stabilisiert. Alle in die- sen Vorgang einbegriff enen Zwischenreak- tionen sind bisher nichtuntersucht worden, weil die Analyse solcher komplexer Reak- tionsketten mit einer Vielzahl von diffe- renten Gleichgewichtskonstanten erhebliche experimentelle Schwierigkeiten bereitet.
111. Prothrombin-Thrombin-Transforma- tion
In den vorangegangenen Abschnitten war nur vom aktiven Thrombin und seiner Wirksamkeit fur die FibrinogenIFibrin- Umwandlung die Rede. Die Vielzahl von Einzelfaktoren, die fur die intermedi'ire Bildung dieses Schlusselenzyms der Gerin- nung notwendig sind, blieb bisher aus Grunden der besseren Obersicht uner- wahnt.
Die Entwicklung spezieller Funktionssche- mata zur Einordnung aller bisher bekann- ten Gerinnungsfaktoren in unterschiedli- che Folge- oder Stufenreaktionen, die letztlich zur Freisetzung des Thrombins fuhren, vermag die Unsicherheiten nicht zu mildern, die bei der Frage nach der Steuerung und Regulation eines solchen komplizierten Systems auftauchen mussen. Beschranken wir uns auf eigenstandige Faktoren mit Proteincharakter, so ergibt sich no& mehr Anhalt fur die anfangs er- wahnte Notwendigkeit zur Fixierung die- ser Informationen im genetischen Material und der damit verbundenen funktionsge- rechten Steuerung von Transskription und Translation, d. h. der m-RNA-Synthese und der ribosomalen Ubertragung dieser Informationen in die entsprechenden Pro- teinstrukturen. Soweit es sich um schein- bar unspezifische, korpereigene Substanzen (inaktive Faktoren) verschiedener Zellen oder Zellpartikel handelt, mug ihr Eingriff in den Ablauf des Gerinnungsprozesses als notwendig, sogar im Sinne von Beschleu- nigung oder Hemmung, nicht aber als en- zymspezifisch im Sinne eines streng kau- salen Reaktionszusammenhanges gesehen werden. Wir werden auf diesen Gedanken- gang bei der Betrachtung der Prothrom- bin-Thrombin-Umwandlung noch mehr- mals zurudrkommen mussen, um vereinfa- chende Deutungen auf molekularbiologi- scher Basis fur dieses Geschehen zu finden.
Obgleich gereinigte Prothrombinprapara- te ein einheitliches Verhalten sowohi wah-
rend der Sedimentation im Schwerefeld als auch in der freien Elektrophorese zeigen, enthalten diese Proteinkomplexe verschie- dene an der Thrombinbildung beteiligte Komponenten, die im wesentlichen den unmittelbaren Enzymvorstuf en desThrom- bins und des aktiven Faktors X (bzw. des Autoprothrombins C in der Bezeichnungs- weise von Seegers) entsprechen. Das Mol- gew. dieses Komplexes liegt um 68.000 (Rind). Durch eine anschliefiende Reinigung an DEAE-Cellulose konnen zwar verschie- dene Aktivitaten ,,abgetrennt" werden, aber das genannte Molgew. des Prothrom-
bins wird davon nur in einem geringen Um- fange beeinflufit. Aus diesen Sedimenta- tionsergebnissen lassen sich die molekula- ren Dimensionen errechnen. Der Prothrom- binkomplex hat die Form eines Rotations- ellipsoids mit einem Achsenverhaltnis von 119 A/34 A (a/b). Bei einem Kohlenhydrat- anteil von 11,2 % enthalt dieses Protein Alanin in N terminaler Position und acht Disulfidbrucken im Innern des Mole- kuls.
Der erste Teilschritt zur Thrombinbildung fuhrt zur Dissoziation des Prothrombinkom-
Prothrombin MG: 68.000
Prethrombinkomdex abnetrennres
MG: 15.000 I Spaltung der Polypeptid-
MG: 52.000
kette - Prethrombin
Thrombin MG: 33.000
d Spalcung der verbleibenden
Untereinhei t
Abb. 5. Schematische Darstellung der Um- wandlung von Prothrombin in Thrombin.
Rechts : Abb. 7. Schematische Darstellung der Um- wandlung des Prothrombinkomplexes un- ter Beteiligung des abgetrennten Frag- mentes (Autoprothrombin 111).
Abb. 6. Spaltung des Prothrombin-Kom- plexes in die inaktiven Vorstufen Pre- thrombin und Autoprothrombin 111. Au- toprothrombin C als Folgeprodukt des Autoprothrombins I11 steuert die Umset- zung zum Thrombin (nach Seegers).
6 Prothrombin
I I 1 Prethrombin Inhibitor Autoprothrombin 111
Ca2 + -1onen Gewebsextrakt Plattchen-Cofaktor I I Ca2 +-Ionen
Acceleratorglobulin Lipide
Autoprothrombin C + Peptid (e) ?
Peptid ( e )
Thrombin 1- +
82 Pharmazie in unserer Zeit12. Jahrg. 19731Nr. 3

plexes in Prethrombin mit einem Molgew. von annahernd 52.000-53.000 und ein niedermolekulares Fragment von 15.000. Das inaktive Prethrombin enthalt immer noch 20-40% der Kohlenhydrate der Aus- gangsverbindung, aber die Zahl der N-ter- minalen Aminosauren hat sich auf zwei er- hoht: Lysin und Threonin. Die notwendige Schluflfolgerung aus dieser Beobachtung ist, dai3 Prethrombin aus zwei Untereinheiten oder Polypeptidketten bestehen sollte und dai3 folgerichtig ein Teil des Prothrombin- komplexes wahrend dieses Schrittes abge- trennt und das Restmolekul gespalten wur-
1 Prethrombinkomdex
1 Prethrombin
Autopro- Dab B I D thrombin 111
Autopro- I D thrombF- I D
de. Erst die nachfolgenden Schritte lassen aus dem Prethrombin das aktive Thrombin mit einem mittleren Molgew. von 32.000 bis 33.000 entstehen, das nun in die Gerin- nungsvorgange einzugreifen vermag. Seine molekularen Dimensionen weisen auf ein gestrecktes Molekul mit einer Lange von 84 A und einer Breite von 30 A hin. Seine N-terminalen Aminosauren sind Isoleucin und Threonin. Es handelt sich eben- falls um ein Molekul aus zwei Untereinhei- ten, wobei das Auftreten von Isoleucin an- stelle von Lysin ebenso wie im Falle des Prethrombins die Annahme erlaubt, dai3 eine Polypeptidkette verkurzt oder total abgebaut wurde (Abbildung 5).
Die geschilderte Verkleinerung des Aus- gangsproduktes ,,Prothrombinkomplex" im Verlauf der Thrombinbildung ist eine stark vereinfachte Darstellung des naturlichen Geschehens und beruht vor allem auf den gemessenen Molekulargewichten sowie auf dem AuRreten neuer, endstandiger Amino- sauren. Es erhebt sich die Frage, wie diese mehrstufigen Prozesse gesteuert und gere- gelt werden und was mit den abgetrennten Fragmenten des Prothrombinkomplexes geschieht. Von Seegers et al. wurde ein Ge- rinnungsschema vorgeschlagen, das weitge- hend den experimentellen Beobachtungen gerecht zu werden versucht (Abbildung 6 ) .
Wie wir aus dem vorstehend skizzierten Reaktionsablauf ersehen konnen, scheint das niedermolekulare Bruchstuck der Pro- thrombin/Prethrombin-Umwandlung einen eigenstandigen Faktor (Autoprothrombin I11 oder inaktiver Faktor X) zu bilden, der im nachfolgenden Schritt als Autopro- thrombin C (aktiver Faktor X) die Um- wandlung des Prethrombins zu aktivem Thrombin katalysiert. Das Autoprothrom- bin 111 mu8 ebenfalls eine Schlussel- position in den physiologischen Gerinnungs- vorgangen innehaben und verdient aus die- sem Grunde, naher untersucht zu werden.
Das Molgew. des Autoprothrombins I11 (87.000 nach Esnouf und Williams, 52.631 nach Seegers et ul.) 1& darauf schlieflen, dai3 diese Substanz um ein Mehrfaches gro- i3er ist als das abgetrennte Fragment der Prothrombin / Prethrombin- Umwandlung.
Mehr noch als diese Angaben zwingen uns die unterschiedlichen Analysendaten iiber die endstandigen Aminosauren dam, neue Konzeptionen zu entwickeln.
Tabelle 1. N-terminale AminosZuren des Faktors X (Autoprothrombin 111)
Alanin, Glycin nach Esnouf und Williams [3], Glycin, Serin nach Hogenauer [4], Glycin, Alanin und Serin nach Murano [5]
Wenn wir annehmen, dai3 es sich im Falle des Autoprothrombins I11 um ein Molekul rnit einem Molgew. von maximal 87.000 handelt, ist es unwahrscheinlich, dai3 das komplette Molekul Bestandteil des Pro- thrombins isr. Entweder waren die Prapa- rate des Prothrombins lediglich mit diesem Faktor verunreinigt oder es mui3 - falls das Modell von Seegers zutrifft - ange- nommen werden, dai3 das abgetrennte Frag- ment assoziieren kann, um das aktive Auto- prothrombin III zu bilden (Abbildung 7).
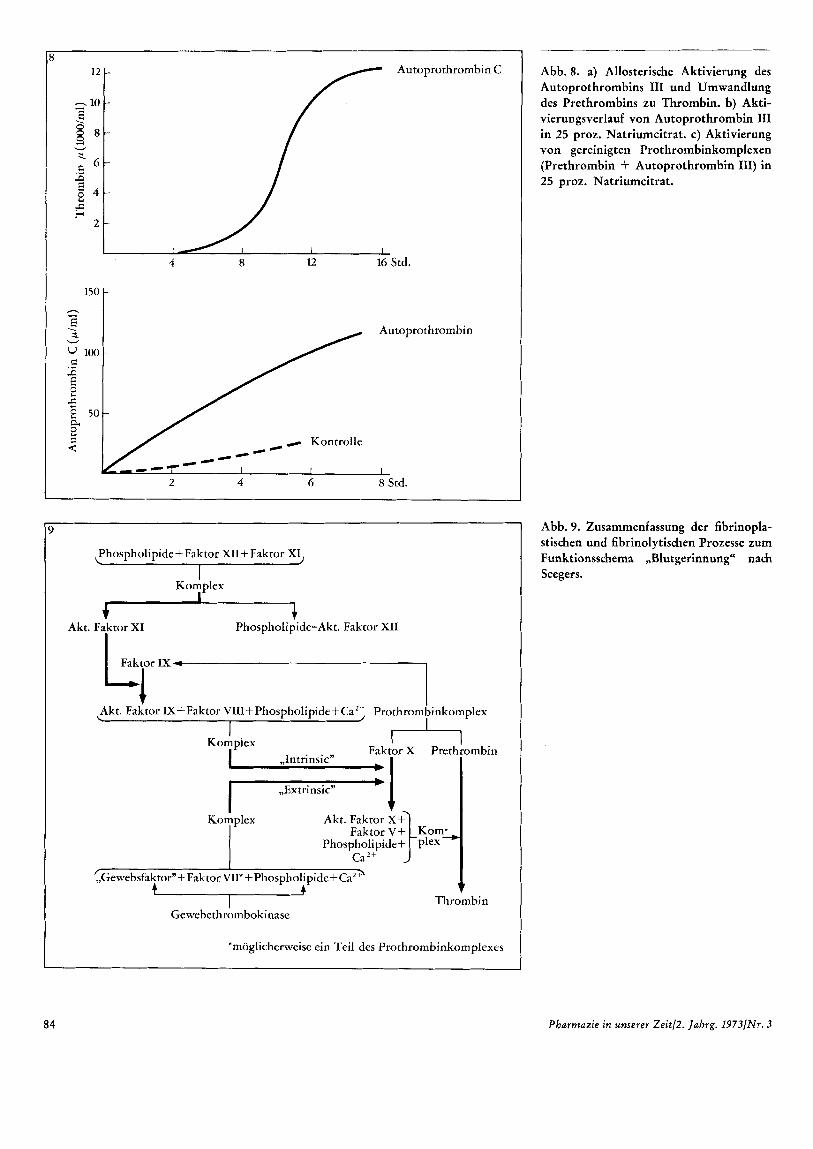
Der Schlussel fur die Losung dieses Pro- blems scheint in Seegers' eigenen Experi- menten zu liegen: Er unternahm den etwas ungewohnlich erscheinenden Versuch, die Umwandlung von inaktivem Prethrombin in aktives Thrombin in 25 proz. Natriumci- trat zu untersuchen: Prethrombin allein ge- winnt unter diesen Bedingungen nie irgend- welche thrombingleichen Aktivitaten. Wird aber dem Ansatz Autoprothrombin C zu- gesetzt, also das schon aktive Produkt aus Autoprothrombin 111, steigt die Thrombin- aktivitat linear mit der Inkubationszeit an. Der notwendige Zusatzversuch war die Un- tersuchung der Autoprothrombin III-Um- wandlung zum aktiven Autoprothrombin C unter gleichen Versuchsbedingungen. Es konnte zweifelsfrei demonstriert werden, dai3 Autoprothrombin I11 in 25 proz. Na- triumcitrat eine Aktivierung erfahrt, die groi3e Ahnlichkeit mit der Aktivierungsre- aktion von allosterischen Enzymen erken- nen 1ai3t. Aufgrund dieses fundamentalen Experiments kann unsere Vorstellung der PrethrombiniThrombin-Umwandlung auf eine mogliche allosterische Aktivierung des Autoprothrombins 111 zuruckgefuhrt wer- den (Abbildung 8).
Die allosterische Beeinflussung der Gerin- nungsmechanismen scheint allgemeinerer Natur zu sein, wenn man vor allem an die Beteiligung unspezifischer Phospholipide denkt, die mit den verschiedensten Fakto- ren Multienzymkomplexe zu bilden vermo- gen. Solche quartaren Proteinstrukturen
Pharmazie in unserer Zeitl2. Jahrg. 1973/Nr. 3 83
4 8 12 16 Std.
150
2 4 6 8 Std.
Phospholipide+Faktor XII+ Faktor XI
Komplex I I
4 I
4 Akt. Faktor XI Phospholipide-Akt. Faktor XI1
t I Akt. Faktor IX+ Faktor VIII + Phospholipide + Ca2+ Prothrombinkomplex
I A I I
Faktor X Prethrombin 4 I ,,Intrinsic”
Komplex Akt. Faktor X+7 Fakt
Phospholi
1 LGewebsfaktor”+ Faktor VII* +Phospho1ipide+Ca2’ & A
7 Gewebethrombokinase
Thrombin
*moglicherweise ein Teil des Prothrombinkomplexes
Abb. 8. a) Allosterische Aktivierung des Autoprothrombins 111 und Umwandlung des Prethrombins zu Thrombin. b) Akti- vierungsverlauf von Autoprothrombin 111 in 25 proz. Natriumcitrat. c) Aktivierung von gereinigten Prothrombinkomplexen (Prethrombin + Autoprothrombin 111) in 25 proz. Natriumcitrat.
Abb. 9. Zusammenfassung der fibrinopla- stischen und fibrinolytischen Prozesse zum Funktionsschema ,,Blutgerinnung“ nach Seegers.
84 Pharmazie in inserer Zei t l2 . Jahrg. 19731Nr. 3

sind weiter verbreitet, als man bisher ver- mutet hat, und man darf annehmen, da8 die meisten Proteine mit Molekulargewich- ten uber 50.000 eher aus Untereinheiten aufgebaut sind als aus einer einzigen Poly- peptidkette.
Der Aufbau der enzymatischen Proteine aus Untereinheiten hat den ungeheuren Vorteil, dai3 ihre Aktivitat durch die Bindung klei- nerer Molekule (Substrat oder Produkt ei- ner Enzymreaktion) auf das physiologische Optimum einreguliert werden kann. Ana- loge molekulare Wechselwirkungen diirften auftreten, wenn enzymatisch aktive Fakto- ren (IX, X), inaktive Faktoren (V, VII, VIII) und Phospholipide funktionelle Komplexe bilden:
Faktor X+Faktor V+Phospholipide-+ aktivierter Faktor X - Komplex
Faktor IXf Faktor VIII +Phospholipide-t aktivierter.Faktor IX - Komplex
Der Beitrag der inaktiven Faktoren zu die- sen Komplexen sollte nicht so sehr in einer coenzymlhnlichen Funktion, sondern eher unter dem Gesichtspunkt einer Oberflachen- aktivierung gesehen werden. Coenzyme sind Substanzen mit prosthetischen Grup- pen, denen eine eindeutige biochemische Funktion zugeordnet werden kann, - diese Eigenschaft ist den inaktiven Faktoren nicht zuzuschreiben. Betrachten wir dagegen das gesamte Gefaflsystem mit dem darin stro- menden Blut als ein funktionelles Compart- ment mit normalerweise in biologischen Sy- stemen nicht anzutreff enden Dimensionen, so ist eine strukturelle Organisation aller Gerinnungsfaktoren an stromenden Parti- keln wie den Thrombocyten logisch und zweckmai3ig.
Ihnen wiirde damit die Rolle spezifischer ,,Carrier" zufallen, ihrer Oberflache die ei- nes ,,strukturellen Organisators". Damit lassen sich die Thrombocyten hinsichtlich ihrer Hamostasefunktion mit den intrazel- lularen Mitochondrien oder dem endothe- lialen Reticulum vergleichen, also Struktu- ren, von denen die raumlich orientierte Bindung verschiedenster enzymatischer Pro- teine an Membranen bekannt ist. Die Inte- gration der Enzyme in biologische Grenz- flachen diff eriert natiirlich von Zelltypus zu Zelltypus. Die jeweils angewandten Pra- parationsmethoden fiihren daher notwen- digerweise zu schweren Eingriffen in die
Struktur und Funktion der isolierten En- zyme. Unter Beriicksichtigung dieser Tat- sache sind die beschriebenen Phospholipid- Enzym-Komplexe der aktiven Faktoren des Gerinnungssystems als Teilelemente eines Ganzen zu bewerten, die immer no&, ih- rem Ursprung gemai3, komplex und multi- potent sind. Bei Fortfuhrung dieses Gedan- kenganges kommt man zur Vorstellung von einer membrangebundenen Organisation aller aktiven Faktoren der Gerinnung als dem biologischen Grundzustand, fur den es bis heute zwar keine schliissigen Beweise gibt, der aber durchaus im Bereich des Mog- lichen liegt, da analoge Systeme intrazellu- lar existent sind.
IV. Regulation und Feedback-Mechanis- men der Gerinnung
Die ,,Vorwartsreaktion" des Gerinnungs- systems umfa8t alle Einzelreaktionen, die zur Bereitstellung des Thrombins notwen- dig sind. Die in den vorangegangenen Ab- schnitten erwahnten Teilschritte werden dabei zu einem sinnvollen Ganzen zusam- mengefa8t (Abbildung 9).
Die auslosende Reaktion zum Start des Reaktionscyclus ist im intravasalen System die Freisetzung von Phosphatiden aus Thrombocyten oder Erythrocyten, die als niedermolekulare Substanzen im Sinne ei- ner allosterischen Regulation die Um- wandlung von Autoprothrombin 111 (Fak- tor X) zum aktiven Komplex Autopro- thrombin C ,,triggernu, wobei die kleinste Menge entstehenden Thrombins gleichzei- tig steuernd und beschleunigend in diesen Mechanismus eingreift. Da der Prothrom- binkomplex allein durch dieses Enzym zum Prethrombin, dem essentiellen Vorlau- fer des Thrombins, umgesetzt werden kann, mui3 auch fur diesen Schritt ein Feedback- mechanismus existieren, fur den Schroer et al. [lo] einen Regelkreis postuliert ha- ben, in welchem ein Inhibitor aus dem Pro- thrombinkomplex die RegelgroSe darstellt.
Neben diesem relativ langsam reagierenden "intrinsic pathway", der auf das Compart- ment des Kreislaufsystems beschrankt ist, existiert eine enge funktionelle Koppelung zum umgebenden Gewebe ("extrinsic path- way"): Traumatische Storungen in diesem Bereich fuhren momentan zur Freisetzung eines Gewebefaktors, der mit dem Faktor VII und den oberflachenaktiven Phospholi- piden einen enzymgleichen Komplex bildet.
Die Regulation des Gesamtgeschehens ist nicht nur auf eine allosterische Beeinflus- sung der enzymatischen Aktivitaten be- schrankt, sondern wird im gleichen Ma8e durch Inhibitoren im Sinne einer ,,Grobein- stellung" gesteuert. Angriffspunkte fur die Inhibitoren sind prinzipiell alle enzymati- schen Zwischenstufen oder das Endprodukt der Prothrombinumwandlung selbst, das Thrombin. Als spezifische physiologische Inhibitoren wurden der Anti-Faktor XI, Anti-Faktor VIII, das Antithrombin I11 und das ebenfalls als Thronibininhibitor wir- kende ae-Makroglobulin beschrieben. Eine zusatzliche Beeinflussung des Gerinnungs- ablaufes ist durch den oben erwahnten In- hibitor aus dem Prothrombinkomplex so- wie durch die Aktivitat des Thrombins selbst gegeben, das den zur Bildung des Au- toprothrombin-C-Komplexes notwendigen Faktor V nicht nur zu aktivieren, sondern auch nachfolgend proteolytisch zu spalten vermag.
Das Einspielen des Gesamtsystems ,,Kreis- lauf" auf den physiologischen Normal- zustand nach Ablauf einer Gerinnung ist vor allem an die Inaktivierung des Thrombins, an den raschen Abbau des Fi- brinpolymerisates und die Synthese ver- brauchter Gerinnungsfaktoren gebunden. Die auflergewohnlich hohe Turnover-Rate vieier Gerinnungsfaktoren wird allgemein als Folge eines standigen Verbrauchs im Sinne einer ,,latenten intravasalen Ge- rinnung" gedeutet. Dieser physiologi- sche ProzeS einer permanenten Gerin- nung kleinsten Ausma8es im stromenden Blut setzt eine relative Oberbetonung der ,,Ruckwartsreaktion" (Fibrinolyse) im Ge- samtsystem voraus. Der Organismus ist so zur bestandigen Neusynthese aller Fakto- ren gezwungen, da das dynamische Gleich- gewicht durch den Faktorenverbrauch nach der Seite der Fibrinolyse verschoben ist. Dementsprechend ist die Konzentration an inaktiven Vorstufen proteolytischer Enzy- me im Plasma und anderen Korperflussig- keiten notwendigerweise hoch. Eine funk- tionell sehr sinnvolle Koppelung zwischen ,,Vorwarts"- und ,,Ruckward'-Reaktion ist dadurch gegeben, da8 die fibrinolytische Substanz in ihrer inaktiven Vorstufe, dem Plasminogen oder Profibrinolysin, eine ho- he Affinitat sowohl zum Fibrinogen als auch zum Fibrin hat. Das entstehende Fibrin- polymerisat enthalt allein aus diesem Grun- de 20-30 % des im Plasma vorhandenen Plasminogens, so dai3 die spezifischen Akti-
Pharmazie in unserer Zeitl2. Jahrg. 19731Nr. 3 85

vatoren zur Bildung des proteolytisch akti- ven Plasmins oder Fibrinolysins schon an diesen Komplex gebunden sind und mit dem entstehenden Thrombus immobilisiert werden konnen. Das Postulieren der Exi- stenz eines Plasminogen-Aktivators ist um- stritten, da Hinweise dafur vorliegen, dai3 das aktive Plasmin an einen Inhibitor ge- bunden sein kann, dessen Dissoziation das proteolytische Enzym freigibt. Erschwerend fur die Klarung der Frage, ob ein Aktiva- tor oder ein Inhibitor eingreifen mu& war der Nachweis eines spezifischen Inhibitors, der in hohen Konzentrationen im Plasma vorkommt, des Antiplasmins. Allein die Af- finitatsdiff erenz der beiden Inhibitoren zum Plasmin entscheidet uber die Geschwindig- keit der Bildung inaktiver Komplexe der einen oder anderen Art. In Gegenwart von Fibrin dissoziieren allerdings die Plasmin- Inhibitor-Komplexe, da ihre Dissoziations- konstanten groi3er sind als die des Fibrin- Plasmin-Komplexes. Das gebundene Plas- min spaltet proteolytisch das Fibrin zu nie- deren Peptiden und lost die vernetzte Fi- brinstruktur auf.
Betrachtet man dieses Geschehen am eigent- lichen Ort seiner Bedeutung, der termina- len Strombahn, so lai3t sich experimentell nachweisen, dai3 Plasminogen-Aktivatoren aus dem vascularen Endothel austreten und so zur Lyse von Fibringerinnseln beitragen. Solche lytischen Prozesse sind auch fur den Abbau des bei der latenten Gerinnung ent- stehenden, sich an der Gefai3wand abla- gernden fibrinahnlichen Materials unent- behrlich. Die Mechanismen zur Bereitstel- lung des Plasminogen-Aktivators und alle notwendigen Prozesse zur Umwandlung des Plasminogens zu Plasmin sind weitge- hend unbekannt. Unter physiologischen Normalbedingungen wird ein Teil des ak- tiven Plasmins durch Antiplasmin inakti- viert, wahrend ein kleinerer Prozentsatz im Gefaasystem bestandig zirkuliert. Der An- stieg des Plasminogen-Aktivators steigert die Umsatzrate von Plasminogen in Plas- min, das gleichzeitig durch die genannten Inhibitoren nach der Fibrinolyse wieder in- aktiviert wird.
V. Zusammenfassung und Ausblidz
Die geschilderten Zusammenhange der Ge- rinnungsprozesse, die wir in fibrinoplastische und fibrinolytische Reaktionen unterteilen konnen, sind ein Ausdruck der phylogeneti- schen Hoherentwicklung der Vertebraten.
Sie sind auf das physiologische Compart- ment des Kreislaufs beschrankt, stehen aber in standiger Wechselwirkung mit dem Ge- samtorganismus. Die Synthese der gerin- nungsaktiven Faktoren findet aui3erhalb dieses Systems uberwiegend in der Leber statt, so dai3 ihr Antransport und der Ab- transport inaktivierter Faktoren gewahr- leistet sein mug. Alle ablaufenden Reak- tionen zur Bildung des Fibrins und damit die Sicherung der Kreislauffunktion sind aufeinander abgestimmt und werden durch eine Reihe aktiver Faktoren gesteuert. Die von uns vorgeschlagene Konzeption der allosterischen Regelung von Einzelschritten beruht auf einer molekularbiologischen Be- wertung der Tendenz der beteiligten Fak- toren, verschiedene funktionelle Komplexe zu bilden. Das Einbeziehen der Thrombo- cyten fur den Transport solcher Komplexe ist naheliegend, da die meisten - wenn nicht alle - an der Gerinnung beteiligten Faktoren adsorptiv an ihre Oberflache ge- bunden sind. Die weiterfuhrende Hypo- these, dai3 die oberflachengebundenen Fak- toren funktionell geordnet und die bis- her untersuchten aktiven Komplexe Teil- elemente dieser Ordnungsstruktur sind, ist ein Analogieschlui3 zu bereits beschrie- benen biologischen Funktionseinheiten wie den Mitochondrien oder dem reticuloendo- thelialen System. Wie bei allen Hypothesen hat auch unsere Vorstellung den Nachteil, dai3 ihre Richtigkeit erst durch weitere Ex- perimente bewiesen werden mui3. Sie scheint uns aber geeignet, als Denkmodell der Gerinnungsforschung neue Impulse zu vermitteln.
Literatur
[l] F. Capet-Antonini u. S. Gunand: C. R. Acad. Sci., Paris 265, 2093 (1967).
[2] G. Koppel, in ,,Biochemie und Akti- vierung des Prothrombins; Kontaktakti- vierung, biologisch gesehen; Niereninsuf- fizienz und Hamostase". F. K. Schattauer Verlag Stuttgart-New York, 1968, S. 129.
[3] M. P. Esnouf u. W. J. Williams: Bio- chem. J. 84, 62 (1962).
[4] E. Hogenauer, K. Lechner u. E. Deutsch: Thromb. Diath. Haemorrhag. 19, 304 (1968).
[5] G. Murano, Ph. D.-Dissertation, Way- ne State University, Detroit, USA.
[6] A. G. Ware, M. Guest u. W. H. Seegers: Arch. Biochem. 13, 231 (1947).
[7] B. Blomback u. M. B. Blomback: Arkiv Kemi 10, 415 (1956).
[8] M. W. Mosesson u. S. Sherry: Bioche- mistry 5, 2829 (1966).
[9] W. H. Seegers, L. McCoy, E. Marciniak u. G. Murano: in "Disseminated Intravas- cular Coagulation", F. K. Schattauer Ver- lag Stuttgart-New York, 1969, S. 239.
[lo] H. Schroer, D. Groben u. G. Hauck: 13th Internat. Congr. of Hematology - Abstract Vol. Munich 1970, p. 15.
Prof. Dr. med. Heinz Schroer, Direktor des Physiolog. Inst. der Univ. Munster, geb. 1922 in Duisburg-Beeck. Promotion 1949. Habilitation fur Physiologie 1958 Univ. Wurzburg, dort 1964 apl. Professor. Seit 1971 0. Professor an der Univ. Mun- ster. Arbeitsgebiete: Physiologie der Blut- gerinnung und der Mikrozirkulation.
Privatdozent Dr. rer. nat. Eberhard Zim- mermann, geb. 1930 in Grimma. Promo- tion 1959 am Inst. fur Vegetative Physio- logie der Univ. Frankfurt. Habilitation fur Physiologische Chemie 1968 Univ. Wurzburg. Seit 1972 Dozent am Physio- log. Inst. der Univ. Munster. Arbeitsge- biete: Zellularer RNS-Stoffwechsel, Visko- sitat, Chromosomale Informationen, Phy- siologie und Biochemie der Blutgerinnung.
86 Pharmazie in Unserer Zeitl2. Jahrg. 1973/Nr. 3





























