Embed Size (px)
Citation preview

Mutation Research, 178 (1987) 11-20 11 Elsevier
MTR 04323
N A D P H as rate-limiting factor for microsomal metabol i sm
An alternative and economic NADPH-genera t ing system for microsomal mono-oxygenase in in vitro genotoxicity studies *
M. Paolini, P. Hrelia 1, C. Corsi 2, G. Bronzetti 2, G.L. Biagi and G. Cantelli-Forti Istituto di Farmacologia dell'Universit?~ di Bologna, Via Irnerio, 48, 40126 Bologna, 1 Istituto di Istologia ed Embriologia Generale,
Via Belmeloro, 8, 40126 Bologna, and 2 lstituto di Mutagenesi e Differenziamento CNR, Via Svezia, 10, 56100 Pisa (Italy)
(Received 14 May 1986) (Revision received 5 November 1986)
(Accepted 7 November 1986)
Keywords: Microsomal metabolism; NADPH; Mono-oxygenase; Deethylase; Demethylase.
Summa~
The effect of NADPH supply on enzymatic activity and its stability were investigated with respect to the mono-oxygenase activities of 7-ethoxyresorufin O-deethylase (ERD), dinemorphan N-demethylase (DND), aminopyrine N-demethylase (APD), 7-ethoxycoumarin O-deethylase (ECD) and p-nitroanisole O-demethylase (p-NAD) under incubation conditions for the liver microsomal assay (LMA). Experiments with $9 liver fractions of mouse (induced with Na-phenobarbital and/3-naphthoflavone) and rat (induced with Aroclor 1254) were set out at different pre-incubation times with and without exogenous isocitrate dehydrogenase (IC-DH) in the LMA. Such LMA mixtures contain Mn 2+, NADP +, DL-isocitrate (IC) and endogenous IC-DH as NADPH-generating machinery.
No changes in mono-oxygenase stability and lipid peroxidation (LP) were observed in the presence of exogenous IC-DH. The metabolizing capability at the considered times was the maximal one, as shown by no stability changes after the direct addition of IC-DH to the enzymatic assays.
Exogenous IC-DH in the incubation for LMA did not alter the mitotic crossing-over and the mitotic gene conversion of dimethylnitrosamine (DMNA) and AR2MNFN (a nitroimidazo[2,1-b]thiazole) in the tester D7 strain of Saccharomyces cerevisiae.
It was concluded that endogenous IC-DH seems to be sufficient to provide a saturating level of NADPH for mono-oxygenase activities during incubations for LMA without additional external NADPH-generating enzyme activity.
* This work was presented at the 17th Annual Meeting of the Environmental Mutagen Society, 9-13 April 1986, Bal- timore, MD (U.S.A.).
Correspondence: Prof. Giorgio Cantelli-Forti, Istituto di Farmacologia, Universith degli Studi, Via Irnerio, 48, 40126 Bologna (Italy).
Abbreviations. APD, aminopyrine N-demethylase; AC, Aroclor 1254; AR2MNFN, a nitroimidazo[2,1-b]thiazole derivative;
AHH, aryl hydrocarbon hydroxylase; DMSO, dimethyl sulphoxide; DND, dinemorphan N-demethylase; DMNA, di- methylnitrosamine; ERD, 7-ethoxyresorufin O-deethylase; ECD, 7-ethoxycoumarin O-deethylase; G-6-P, glucose 6-phos- phate; G6P-DH, glucose-6-phosphate dehydrogenase; IC, Dt- isocitrate; IC-DH, isocitrate dehydrogenase, LMA, liver micro- somal assay; LP, lipid peroxidation; MDA, malonyl dial- dehyde; PB, sodium phenobarbital; B-NF, /3-naphthoflavone; TBA, 2-thiobarbituric acid; TCA, trichloroacetic acid.
0027-5107/87/$03.50 © 1987 Elsevier Science Publishers B.V. (Biomedical Division)

12
A suitable concentration of the active meta- bolic intermediate(s) must be produced for a suffi- cient period of time to induce mutagenicity with a potentially active compound in in vitro mutagenic- ity test systems. In this regard, the balance be- tween the activating microsomal mono-oxygenases and detoxifying mechanisms (including their rela- tive stabilities during incubations for the LMA), in addition to the spontaneous decay of the active metabolic intermediate, unspecific binding, cellu- lar permeability and DNA repair (see, for exam- ple: Harm, 1980; Peter and Bolt, 1981; Pacifici et al., 1981), should be the main factors influencing the genetic response. However, previous studies in our laboratory (Cantelli-Forti et al., 1984) have shown that the decline of some demethylation and deethylation reactions, in livers from mice and rats, is due to reduced availability of NADPH for mixed function oxidation. It was supported by the lower genetic effect of DMNA and AR2MNFN on Saccharomyces cerevisiae in the absence of exogenous G6P-DH. Studies by Miller and With- lock (1982) provided direct evidence that NADPH is a rate-limiting factor in the oxidation of benzo[a]pyrene in murine hepatoma cells. More recently, Sadowski et al. (1985) found similar re- sults in permeabilized whole cell systems. Thur- man and Scholz (1969) demonstrated that dif- ferences in the rate of aminopyrine oxidation and of p-nitroanisole demethylation (Thurman et al., 1977), were due to different rates of NADPH generation. Although induction by fl-naphthofla- vone or 3-methylcholanthrene increases the NADPH-generating capacity (Kauffman et al., 1980; Nebert et al., 1981) and in this way the pyridine nucleotide pool size, such induction is able to sustain high rates of mono-oxygenation but only for about 40 rain (Conway et al., 1983). Reduced NADPH may become the rate-limiting factor when high rates of mixed function oxida- tion occur in the liver of phenobarbital-treated rats (Thurman et al., 1977). The NADPH supply has also been noted to be a regulating factor of drug metabolism in intact hepatocytes (Thurman and Kauffman, 1979). The role of reducing equiv- alents, provided from the pentose cycle pathway, has been studied (Belinski et al., 1985).
Our results and the above-mentioned consider- ation indicate that studies for in vivo and in vitro
drug metabolism may be altered by the steady-state levels of NADPH.
It is our design to employ a more convenient generation and utilization of NADPH for S9-frac- tion-mediated genotoxicity. Short-term tests with metabolic activation generally utilize Mg 2+, glu- cose 6-phosphate (G-6-P), NADP + and endoge- nous G6P-DH for the NADPH-generating ma- chinery. In this work the use of endogenous NADPH-linked IC-DH, instead of G6P-DH, was examined to provide reducing equivalents needed for mixed function oxidase activity. We explored the use of an alternative NADPH-generating sys- tem which requires only the adding of Mn 2+, NADP + and DL-isocitrate to the $9 fraction but not IC-DH for maximal mono-oxygenase activity. This is due to the fact that in rodent livers the activity of IC-DH is about 11-14 times higher than that of G6P-DH (Veech, 1978; Veech et al., 1972; Yasumi et al., 1980; Cantelli-Forti et al., 1984; Kosenko and Kaminsky, 1985).
The stability of ERD, DND, APD, ECD and p-NAD activities, with and without IC-DH in the exact incubation mixtures for the LMA, was studied. The stability of endogenous IC-DH and lipid peroxidation was also considered.
Finally, to determine whether the biological response is affected by the presence of exogenous IC-DH in the incubation mixtures for LMA, we performed a mutagenicity assay with strain D7 of Saccharomyces cerevisiae using dimethylnitrosa- mine (DMNA) and AR2MNFN as premutagens. Genetic results were compared to those observed with and without G6P-DH. Possible implications of the improvement in LMA economy and sensi- tivity are discussed.
Materials and methods
The animal inductions, the $9 preparations and the determinations of APD, p-NAD, ECD and protein, were reported in a previous paper (Cantelli-Forti et al., 1984). DND and ERD activ- ities were assayed according to Gervasi et al. (1984) and Klotz et al. (1984), respectively. The enzyme stability was determined in the conditions of LMA at time 0, after 1 and 2 of pre-incubation in the presence of NADP +, oL-isocitrate and MnC1 z in Na+/K+-phosphate buffer (0.01 M, pH
13
7.4), so that the respective final concentrations were 1, 5 and 1.2 mM, with and without IC-DH (final concentration 0.80 U/ml) . The isocitrate dehydrogenase was assayed using the procedure reported by Boehringer-Mannheim GmBH (1975) and lipid peroxidation according to Buege and Aust (1978).
Mutagenesis test The yeast cells in stationary phase (about 6 x
10 s cells) were incubated in the LMA in the pres- ence of various concentrations of DMNA (final concentrat ion 50, 100 and 200 mM) and AR2MNFN (final concentration 0.5, 5 X 10-2 and 5 × 10 3 /~M). The experiments were performed with and without exogenous G6P-DH (with the correspondent LMA mixture, Cantelli-Forti et al., 1984) or IC-DH (final concentrations, respec- tively, 0.7 and 0.8 U/ml) . The mixtures were incubated for 2 h at 37 ° C. Then they were plated on selective media to evaluate trp + convertants and ade- recombinants expressed as number of cells/105 and 103 survivors, respectively, and on complete medium for survivor counts (Zimmer- mann et al., 1975).
Statistics Results were submitted to statistical analysis
using the Wilcoxon's ' rank method' as reported by Box and Hunter (1978).
?
;.J
?
E
o
2 0
1 5
1 0
0 5
0
2 0 -
7 • ,-4 1 . 5
T.o
X
? "~ 0.5
X
0 e" 0
Time of p r e - i n c u b a t i o n (hours)
, , '=
Time of pre-incubation (hours)
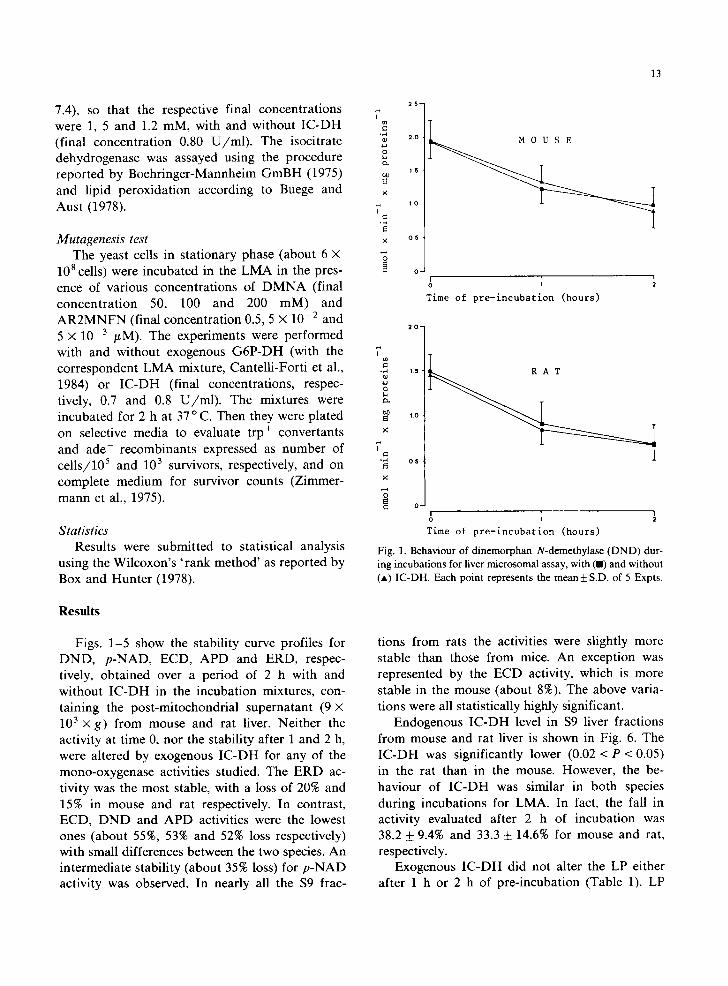
Fig. 1. Behaviour of dinemorphan N-demethylase (DND) dur- ing incubations for liver microsomal assay, with (I) and without (*) IC-DH. Each point represents the mean + S.D. of 5 Expts.
Results
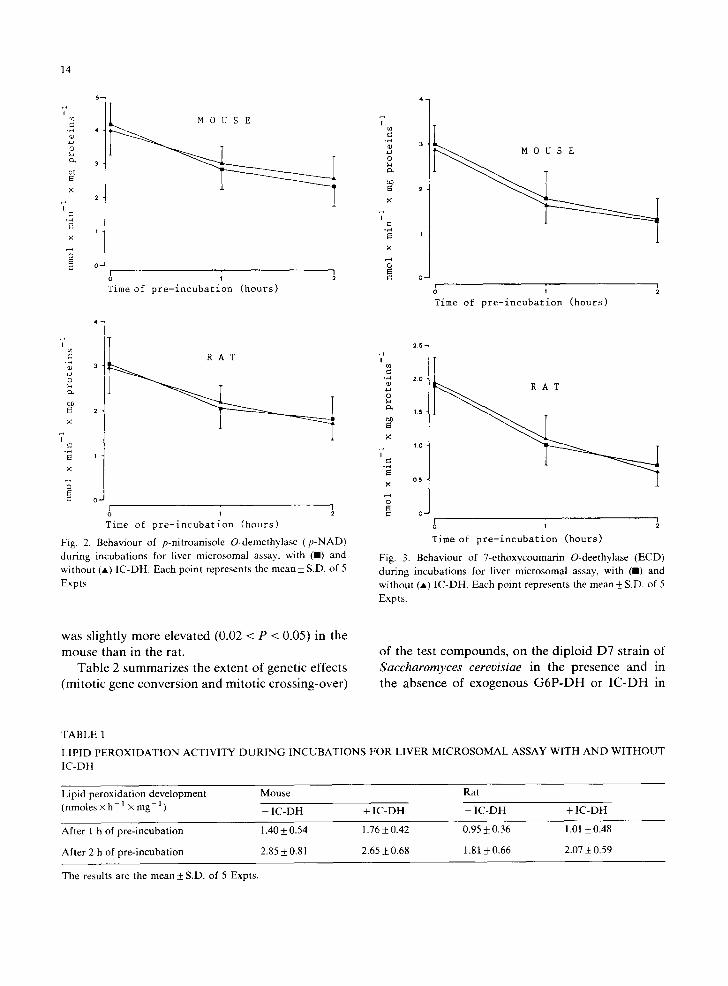
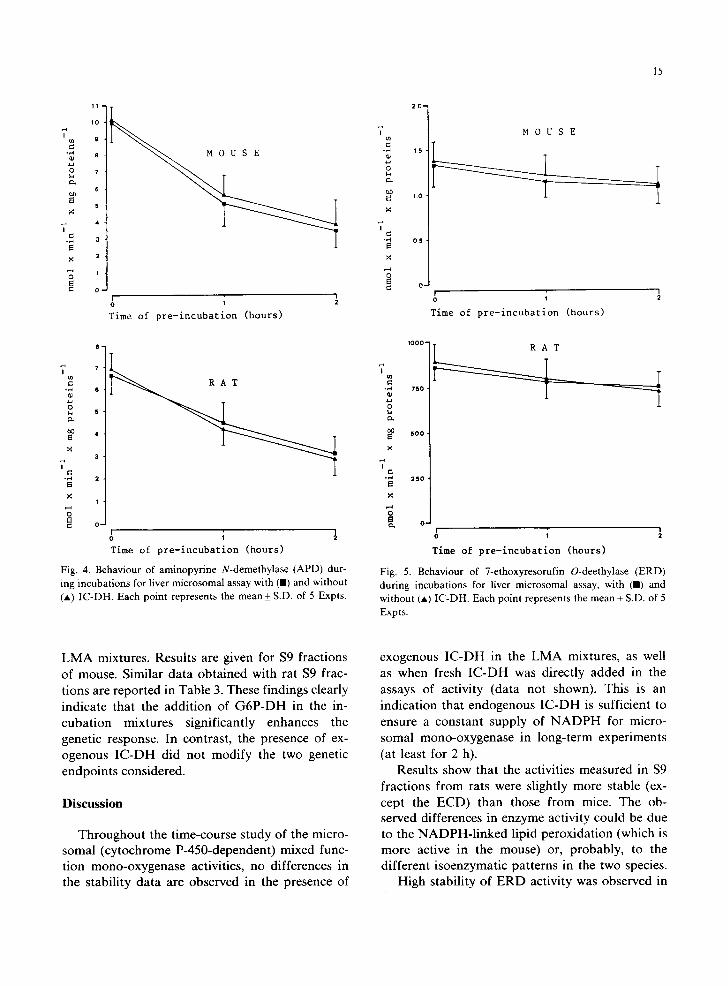
Figs. 1-5 show the stability curve profiles for DND, p-NAD, ECD, APD and ERD, respec- tively, obtained over a period of 2 h with and without IC-DH in the incubation mixtures, con- taining the post-mitochondrial supernatant (9 × 103x g) from mouse and rat liver. Neither the activity at time 0, nor the stability after 1 and 2 h, were altered by exogenous IC-DH for any of the mono-oxygenase activities studied. The ERD ac- tivity was the most stable, with a loss of 20% and 15% in mouse and rat respectively. In contrast, ECD, DND and APD activities were the lowest ones (about 55%, 53% and 52% loss respectively) with small differences between the two species. An intermediate stability (about 35% loss) for p-NAD activity was observed. In nearly all the $9 frac-
tions from rats the activities were slightly more stable than those from mice. An exception was represented by the ECD activity, which is more stable in the mouse (about 8%). The above varia- tions were all statistically highly significant.
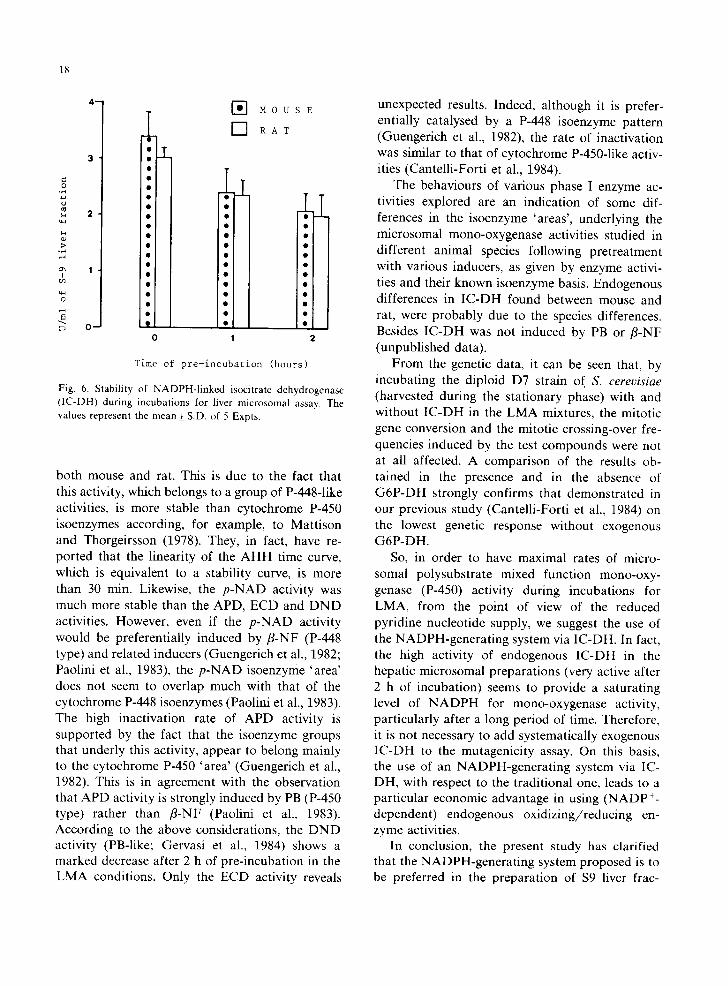
Endogenous IC-DH level in $9 liver fractions from mouse and rat liver is shown in Fig. 6. The IC-DH was significantly lower (0.02 < P < 0.05) in the rat than in the mouse. However, the be- haviour of IC-DH was similar in both species during incubations for LMA. In fact, the fall in activity evaluated after 2 h of incubation was 38.2 _+ 9.4% and 33.3 _+ 14.6% for mouse and rat, respectively.
Exogenous IC-DH did not alter the LP either after 1 h or 2 h of pre-incubation (Table 1). LP
14
?
o
×
T
o E
5--
3
2
l , =,
Time of pre-incubation (hours)
?
o
x
?
x
o E
Time of pre-incubation (hours)
Fig. 2. Behaviour of p-nitroanisole O-demethylase (p -NAD) during incubations for liver microsomal assay, with (m) and without (A) IC-DH. Each point represents the mean_+ S.D. of 5 Expts,
?
o
X
×
T
o
Time of pre-incubation (hours)
I
o
~0 E
×
?
Time of pre-incubation (hours)
Fig. 3. Behaviour of 7-ethoxycoumarin O-deethylase (ECD) during incubations for liver microsomal assay, with ( I ) and without (A) IC-DH. Each point represents the mean _-4- S.D. of 5 Expts.
was slightly more elevated (0.02 < P < 0.05) in the mouse than in the rat.
Table 2 summarizes the extent of genetic effects (mitotic gene conversion and mitotic crossing-over)
of the test compounds, on the diploid D7 strain of Saccharomyces cerevisiae in the presence and in the absence of exogenous G6P-DH or IC-DH in
TABLE 1
LIPID PEROXIDATION ACTIVITY D U R I N G INCUBATIONS FOR LIVER MICROSOMAL ASSAY WITH A N D W I T H O U T
IC-DH
Lipid peroxidation development Mouse Rat
(nmoles × h - ~ x mg 1 ) - IC-DH + IC-DH - IC-DH + IC-DH
After 1 h of pre-incubation 1.40 +_ 0.54 1.76 _+ 0.42 0.95 +- 0.36 1.01 +- 0.48
After 2 h of pre-incubation 2.85 + 0.81 2.65 +- 0.68 1.81 :t- 0.66 2.07 _+ 0.59
The results are the mean+_S.D, of 5 Expts.
1 5
?
oJ
;<
7
x
o F.
1 1 -
10
9
8
7
6
5
4
3
2
I
0
Time of pre-incubation (hours)
8-
7
oJ
~ 4
N 3
7
X 1
o ~ o
Time of pre-incubation (hours)
Fig . 4. Behaviour of aminopyrine N-demethylase (APD) dur- ing incubations for liver microsomal assay with (11) and without ( * ) IC-DH. Each point represents the mean+_S.D, of 5 Expts .
7
E
7 e~ .,-4
r~
20-
10
M O U S E
T
o ,
Time of pre-incubation (hours)
1000"
7 • ,-I 750 O~
e~
N 5oo x
,.-i i " ~ 250
X
[ o.
Time of p r e - i n c u b a t i o n (hours)
Fig . 5. Behaviour of 7-ethoxyresorufin O-deethylase ( E R D )
during incubations for liver microsomal assay, with (U) and without (A) I C - D H . Each point represents the mean + S.D. o f 5
Expts.
LMA mixtures. Results are given for $9 fractions of mouse. Similar data obtained with rat $9 frac- tions are reported in Table 3. These findings clearly indicate that the addition of G6P-DH in the in- cubation mixtures significantly enhances the genetic response. In contrast, the presence of ex- ogenous IC-DH did not modify the two genetic endpoints considered.
Discussion
Throughout the time-course study of the micro- somal (cytochrome P-450-dependent) mixed func- tion mono-oxygenase activities, no differences in the stability data are observed in the presence of
exogenous IC-DH in the LMA mixtures, as well as when fresh IC-DH was directly added in the assays of activity (data not shown). This is an indication that endogenous IC-DH is sufficient to ensure a constant supply of N A D P H for micro- somal mono-oxygenase in long-term experiments (at least for 2 h).
Results show that the activities measured in $9 fractions from rats were slightly more stable (ex- cept the ECD) than those from mice. The ob- served differences in enzyme activity could be due to the NADPH-l inked lipid peroxidation (which is more active in the mouse) or, probably, to the different isoenzymatic patterns in the two species.
High stability of ERD activity was observed in

TA
BL
E 2
IND
UC
TIO
N
OF
G
EN
ET
IC
EF
FE
CT
S
ON
Sa
ccha
rom
yces
ce
revi
siae
D
7 S
TR
AIN
B
Y
DM
NA
A
ND
A
R2
MN
FN
IN
T
HE
S
US
PE
NS
ION
T
ES
T
WIT
H
RA
T
$9
FR
AC
TIO
NS
+G
6P
-DH
O
R I
C-D
H a
Co
mp
ou
nd
C
onc.
S
urvi
val
(%)
ade
reco
mb
inan
ts/1
03
sur
vivo
rs
trp
+ co
nv
erta
nts
/10
5 s
urvi
vors
(mM
) -G
6P
- +
G6
P-
-IC
- +
IC-
-G6
P-
+G
6P
- -I
C-
+IC
- G
6P-
+G
6P
- IC
- +
IC-
DH
D
H
DH
D
H
DH
D
H
DH
D
H
DH
D
H
DH
D
H
Con
trol
10
0 10
0 10
0 10
0 1.
10
1.17
1.
25
1.19
1.
18
0.95
1.
31
1.02
(2
716)
(2
563)
(2
392)
(2
515)
(3
) (3
) (3
) (3
) (3
20)
(243
) (3
13)
(256
)
DM
NA
50
82
83
81
78
1.
35
1.88
2.
06
2.04
1.
57
1.13
2.
70
1.93
(2
227)
(2
127)
(1
938)
(1
962)
(3
) (4
) (4
) (4
) (3
50)
(240
) (5
23)
(378
) 10
0 80
72
77
67
1.
84
2.71
2.
71
2.97
2.
31
2.91
3.
13
2.52
(2
173
) (1
845
) (1
842
) (1
685
) (4
) (5
) (5
) (5
) (5
02)
(537
) (5
76)
(424
) 20
0 71
61
72
61
3.
11
6.40
6.
39
6.52
3.
50
3.87
5.
18
4.56
(1
928
) (1
563
) (1
722
) (1
534
) (6
) (1
0)
(11)
(1
0)
(675
) (6
05)
(892
) (6
99)
AR
2M
NF
N
5 x
10
- 3
92
91
100
95
1.20
2.
57
2.92
2.
93
1.12
1.
07
1.45
1.
31
(249
9)
(233
2)
(239
9)
(238
9)
(3)
(6)
(7)
(7)
(280
) (2
50)
(347
) (3
13)
5 x
10 -
2 90
87
99
90
2.
05
4.04
4.
22
3.97
1.
27
1.38
1.
59
1.71
(2
444)
(2
230)
(2
368)
(2
263)
(5
).
(9)
(10)
(9
) (3
10)
(308
) (3
76)
(387
) 0.
5 89
78
89
87
2.
90
5.50
6.
11
5.94
1.
61
2.13
1.
65
2.12
(2
418)
(1
999)
(2
129)
(2
188)
(7
) (1
1)
(13)
(1
3)
(389
) (4
26)
(351
) (4
64)
a In
par
enth
eses
the
col
onie
s co
unte
d.
The
res
ults
are
the
mea
n of
3
4 in
depe
nden
t E
xpts
.

TA
BL
E 3
IND
UC
TIO
N
OF
GE
NE
TIC
E
FF
EC
TS
O
N S
acch
arom
yces
ce
revi
siae
D7
ST
RA
IN
BY
DM
NA
A
ND
AR
2M
NF
N
IN T
HE
S
US
PE
NS
ION
T
ES
T W
ITH
M
OU
SE
$9
F
RA
CT
ION
S
_+ G
6P
-DH
OR
IC
-DH
a
Co
mp
ou
nd
C
onc.
S
urvi
val
(%)
ade-
re
com
bin
ants
/10
3 s
urvi
vors
(mM
) -
C6P
- +
G6P
- -
IC-
+ IC
- -
G6P
- +
G6P
- D
H
DH
D
H
DH
D
H
DH
trp
+ co
nv
erta
nts
/10
5 s
urvi
vors
- IC
- -
IC-
- G
6P-
+ G
6P-
- IC
- +
IC-
DH
D
H
DH
D
H
DH
D
H
Con
trol
m
ouse
DM
NA
AR
2M
NF
N
- 10
0 10
0 10
0 10
0 0.
75
0.49
0.
46
(266
7)
(202
7)
(216
6)
(224
5)
(2)
(1)
(1)
50
100
98
99
75
0.74
1.
51
0.93
(2
671
) (1
986)
(2
144)
(1
683
) (2
) (3
) (2
) 10
0 83
87
78
68
1.
36
1.70
1.
78
(2 2
14)
(1 7
63)
(1 6
89)
(1 5
26)
(3)
(3)
(3)
200
71
70
66
61
2.11
2.
82
2.80
(1
894
) (1
419)
(1
629
) (1
369
) (4
) (4
) (4
)
5 ×
10
- 3
97
91
100
100
0.77
1.
63
0.92
(2
587)
(1
845
) (2
178)
(2
248)
(2
) (3
) (2
) 5
× 10
z
85
88
89
90
0.88
1.
68
1.56
(2
267)
(1
784)
(1
927)
(2
020)
(2
) (3
) (3
) 0.
5 78
80
75
85
2.
40
3.08
2.
57
(208
0)
(1 6
22)
(1 6
24)
(1 9
08)
(5)
(5)
(4)
0.44
0.
25
0.16
0.
19
0.21
(1
) (6
7)
(32)
(4
1)
(47)
1.19
0.
31
0.25
0.
31
1.71
(2
) (8
3)
(50)
(6
6)
(141
) 1.
31
0.37
0.
42
0.87
1.
31
(2)
(82)
(7
4)
(147
) (1
99)
2.92
0.
71
0.67
1.
78
1.89
(4
) (1
34)
(95)
(2
54)
(258
)
0.89
0.
37
0.30
0.
15
0.28
(2
) (9
6)
(55)
(3
2)
(62)
1.
98
0.40
0.
50
0.54
0.
48
(4)
(91)
(8
9)
(104
) (1
99)
2.62
0.
54
0.68
0.
71
0.62
(5
) (1
12)
(110
) (1
10)
(258
)
a In
par
enth
eses
the
col
onie
s co
unte
d.
The
res
ults
are
the
mea
n o
f 3
-4 i
ndep
ende
nt E
xpts
.
18
o
2
? 1
0
4 1
t
o~ o!
[] []
MOUSE
RAT
1
2
Time of pre-incubation (hours)
Fig. 6. Stability of NADPH-l inked isocitrate dehydrogenase (IC-DH) during incubations for liver microsomal assay. The values represent the mean + S.D. of 5 Expts.
both mouse and rat. This is due to the fact that this activity, which belongs to a group of P-448-1ike activities, is more stable than cytochrome P-450 isoenzymes according, for example, to Mattison and Thorgeirsson (1978). They, in fact, have re- ported that the linearity of the AHH time curve, which is equivalent to a stability curve, is more than 30 min. Likewise, the p-NAD activity was much more stable than the APD, ECD and DND activities. However, even if the p-NAD activity would be preferentially induced by fl-NF (P-448 type) and related inducers (Guengerich et al., 1982; Paolini et al., 1983), the p-NAD isoenzyme 'area' does not seem to overlap much with that of the cytochrome P-448 isoenzymes (Paolini et al., 1983). The high inactivation rate of APD activity is supported by the fact that the isoenzyme groups that underly this activity, appear to belong mainly to the cytochrome P-450 'area' (Guengerich et al., 1982). This is in agreement with the observation that APD activity is strongly induced by PB (P-450 type) rather than fl-NF (Paolini et al., 1983). According to the above considerations, the DND activity (PB-like; Gervasi et al., 1984) shows a marked decrease after 2 h of pre-incubation in the LMA conditions. Only the ECD activity reveals
unexpected results. Indeed, although it is prefer- entially catalysed by a P-448 isoenzyme pattern (Guengerich et al., 1982), the rate of inactivation was similar to that of cytochrome P-450-1ike activ- ities (Cantelli-Forti et al., 1984).
The behaviours of various phase I enzyme ac- tivities explored are an indication of some dif- ferences in the isoenzyme 'areas', underlying the microsomal mono-oxygenase activities studied in different animal species following pretreatment with various inducers, as given by enzyme activi- ties and their known isoenzyme basis. Endogenous differences in IC-DH found between mouse and rat, were probably due to the species differences. Besides IC-DH was not induced by PB or /3-NF (unpublished data).
From the genetic data, it can be seen that, by incubating the diploid D7 strain of S. cereuisiae
(harvested during the stationary phase) with and without IC-DH in the LMA mixtures, the mitotic gene conversion and the mitotic crossing-over fre- quencies induced by the test compounds were not at all affected. A comparison of the results ob- tained in the presence and in the absence of G6P-DH strongly confirms that demonstrated in our previous study (Cantelli-Forti et al., 1984) on the lowest genetic response without exogenous G6P-DH.
So, in order to have maximal rates of micro- somal polysubstrate mixed function mono-oxy- genase (P-450) activity during incubations for LMA, from the point of view of the reduced pyridine nucleotide supply, we suggest the use of the NADPH-generating system via IC-DH. In fact, the high activity of endogenous IC-DH in the hepatic microsomal preparations (very active after 2 h of incubation) seems to provide a saturating level of NADPH for mono-oxygenase activity, particularly after a long period of time. Therefore, it is not necessary to add systematically exogenous IC-DH to the mutagenicity assay. On this basis, the use of an NADPH-generating system via IC- DH, with respect to the traditional one, leads to a particular economic advantage in using (NADP +- dependent) endogenous oxidizing/reducing en- zyme activities.
In conclusion, the present study has clarified that the NADPH-generating system proposed is to be preferred in the preparation of $9 liver frac-

t i o n s f r o m r o d e n t s fo r in v i t r o g e n o t o x i c i t y t e s t i n g
w i t h m e t a b o l i c a c t i v a t i o n of a n u n k n o w n x e n o -
b io t i c .
Acknowledgements
W e are g r a t e f u l to Prof . W i l l i a m W. Au , D iv i -
s i o n of E n v i r o n m e n t a l T o x i c o l o g y of t h e D e p a r t -
m e n t of P r e v e n t i v e M e d i c i n e a n d C o m m u n i t y
H e a l t h , M e d i c a l B r a n c h , G a l v e s t o n , T X (U.S .A. ) ,
fo r r e v i s i n g t he m a n u s c r i p t .
T h i s w o r k was s u p p o r t e d j o i n t l y b y C N R ( N a -
t i o n a l R e s e a r c h C o u n c i l o f I t a ly ) t a r g e t p r o j e c t s
' P r e v e n t i v e a n d R e h a b i l i t a t i v e M e d i c i n e ' ( C o n -
t r a c t N o . 85 .00486 .56) a n d ' O n c o l o g y ' ( C o n t r a c t
N o . 85 .02070.44) .
References
Belinski, S.A., L.A. Reinke, R. Scholz, F.C. Kauffman and R.C. Thurman (1985) Rates of pentose cycle flux in per- fused rat liver, Evaluation of the role of reducing equiv- alents from the pentose cycle for mixed function oxidation, Mol. Pharmacol., 28, 371-376.
Boehringer-Mannheim (1975) Isocitric dehydrogenase, in: Bio- chemical Information, Vol. II, Boehringer Mannheim GmbH, Mannheim, p. 101.
Box, G.E.P., and W.G. Hunter (1978) Statistic for Experi- ments, Wiley, New York, pp. 80-82.
Buege, J.A., and S.D. Aust (1978) Microsomal lipid peroxida- tion, Methods Enzymol., 52, 302-310.
Cantelli-Forti, G., M. Paolini, P. Hrelia, C. Corsi, G.L. Biagi and G. Bronzetti (1984) NADPH-generating system: in- fluence on microsomal mono-oxygenase stability during incubations for the liver-microsomal assay with rat and mouse S-9 fractions, Mutation, Res., 129, 197-291.
Conway, J.G., F.C. Kauffman and R.G. Thurman (1983) Genetic regulation of NADPH supply in perfused mouse liver: role of the Ah locus during induction by 3-methyl- cholanthrene, J. Biol. Chem., 258, 3825-3831.
Gervasi, P.G., D. Benedetti, L. Citti, M. Del Monte and G. Turchi (1984) Dinemorphan N-demethylation by mouse liver microsomes, Experientia, 40, 180-182.
Guengerich, F.P., G.A. Dannas, S. Tracy Wright, M.V. Martin and S.M. Kaminsky (1982) Purification and characteriza- tion of fiver microsomal cytochromes P-450, Electro- phoretic, spectral, catalytic and immunochemical properties and inducibility of eight isoenzymes isolated from rat treated with phenobarbital or fl-naphthoflavone, Biochem- istry, 21, 6019-6030.
Harm, W. (1980) Biological Effects of Ultraviolet Radiation, Cambridge University Press, Ch. 8.
Kauffman, F.C., R.K. Evans, L.A. Reike, S.A. Belinski, C. Ballow and R.C. Thurman (1980) Effects of 3-methyl- cholanthrene on oxidized nicotinamide-adenine dinucleo-
19
tide phosphate-dependent dehydrogenase and selected metabolites in perfused rat liver, Biochem. Pharmacol., 26, 697-700.
Klotz, A.V., J.J. Stageman and C. Walsh (1984) An alternative 7-ethoxyresorufin O-deethylase activity assay: a continuous visible spectrophotometric method for measurement of cy- tochrome P-450 mono-oxygenase activity, Anal. Biochem., 140, 138-145.
Kosenko, E.A., and Y.G. Kaminsky (1985) A comparison between effects of chronic ethanol consumption, ethanol withdrawal and fasting in ethanol-fed rats on the free cytosolic NADP+/NADPH ratio and NADPH-regener- ating enzyme activities in the liver, Int. J. Biochem., 17, 895-902.
Mattison, D.R., and S. Thorgeirsson (1978) Gonad aryl hydro- carbon hydroxylase in rats and mice, Cancer Res., 38, 1368-1373.
Miller, A.G., and J.P. Withlock (1982)Efficient metabolism of benzo[a]pyrene at nanomolar concentrations by intact murine hepatoma cells, Cancer Res., 42, 4473-4478.
Nebert, D.W., and H.V. Gelboin (1968) Substrate-inducible microsomal aryl hydroxylase in mammalian cell culture, I. Assay and properties of induced enzyme, J. Biol. Chem., 243, 6242-6249.
Nebert, D.W., H.J. Eisen, M. Hegishi, M.A. Lang, L.M. Hjel- meland and A.B. Okey (1981) Genetic mechanisms control- ling the induction of polysubstrate mono-oxygenase (P-450) activities, Annu. Rev. Pharmacol. Toxicol., 21,431-462.
Pacifici, G.M., A.R. Boobis, M.J. Brodie, M.E. McManus and D.S. Davies (1981) Tissue and species differences in en- zymes of epoxide metabolism, Xenobiotica, 11, 73-79.
Paolini, M., C. Bauer, C. Corsi, R. Del Carratore, R. Nieri and G. Bronzetti (1983) Stability of microsomal mono-oxygenase during incubations for the liver microsomal assay with $9 fractions of mouse liver under various inductions, Mutation Res., 110, 221-230.
Peter, H., and H.M. Bolt (1981) Irreversible protein binding of aerylonitrile, Xenobiotica, 11, 51-56.
Sadowski, I.J., J.A. Wright and L.G. Israels (1985) A per- meabilized cell system for studying regulation of aryl hy- drocarbon hydroxylase: NADPH as rate limiting factor in benzo[a]pyrene metabolism, Int. J. Biochem., 17, 1023- 1025.
Thurman, R.G., and F.C. Kauffman (1979) Factors regulating drug metabolism in intact hepatocytes, Pharmacol. Rev., 31,229-252.
Thurman, R.G., and R. Scholz (1969) Control of mixed func- tion oxygenase in perfused rat liver, Pharmacologist, 11, 260.
Thurman, R.G., D.P. Marazzo, L.S. Jogenes and F.C. Kauff- man (1977) The continuous kinetic determination of p- nitroanisole O-demethylase in hemoglobin-free perfused rat liver, J. Pharmacol. Exp. Ther., 201,499-506.
Veech, R.L. (1978) Regulation of coenzyme potential by near equilibrium reactions, in: P.A. Srebe and R.W. Estabrook (Eds.), Microenvironments and Metabolic Compartimenta- tion, Academic Press, New York, pp. 17-60.
Veech, R.L., K. Guynn and D. Veloso (1972) The time course in the effects of ethanol on the redox and phosphorylation states of rat liver, Biochem. J., 127, 387-397.

20
Yasumi, M., T. Minaga, K. Nakamura, A. Kizu and H. Izichi (1980) Inhibition of cardiac NADP-linked isocitrate dehy- drogenase by adriamycin, Biochem. Biophys. Res. Com- mun., 93, 631-636.
Zimmermann, F.K., R. Kern and H.S. Rosenberg (1975) A yeast strain for simultaneous detection of induced mitotic crossing-over, mitotic gene conversion and reverse muta- tion, Mutation Res., 28, 381-388.





























