Embed Size (px)
Citation preview

Insect Biochem., i973, 3, 263-273. [Scientechnica (Publishers) Ltd.] 263
N A D P H O X I D A T I O N BY M I C R O S O M A L P R E P A R A T I O N S
O F GYPSY M O T H L A R V A L T I S S U E S
SAMI A H M A D AND A N D R E W J. F O R G A S H
Department of Entomology and Economic Zoology, Rutgers - -The State University, New Brunswick, New Jersey o89o3, U.S.A.
(Received 8 September, 1972)
A B S T R A C T
N A D P H oxidation by microsomal preparations from several tissues of fifth-instar larvae of gypsy moth, Porthetria dispar Linn. was studied. The distribution of this activity was as follows: 53"9, 27 .8 , 9"6, 4'4, and 4"3 per cent in midgut, fat body, hindgut, Malpighian tubules, and foregut respectively. Relatively specific activities of these microsomal preparations were xoo, 81, 77, 61, and 55 for hindgut, Malpighian tubules, midgut, foregut, and fat body. N A D P H oxidation was rapid for the first eo minutes of the reaction and ex- hibited a Km of 1"75 x io -b M NADPH. Cytochrome C greatly enhanced N A D P H oxidation without affecting the Kin. Carbon monoxide inhibited microsomal oxidation of N A D P H and the inhibition was partially reversed by cytochrome C. This suggested the possibili ty of the presence of cytochrome P,50. S K F 525-A inhibited N A D P H oxidation and the locus of inhibition appeared to be cytochrome P,50. Sulphhydryl inhibitors, p-chloromercuri- benzoate, p-chloromercuriphenyl sulphonate and Cu 2+, inhibited N A D P H oxidation and the inhibition was not reversed by cytochrome C. This indicated the involvement of NADPH-cytochrome C-reductase in the electron transport chain in gypsy moth.
MiXED-function oxidases play a major role in the metabolism of xenobiotics in mammals (Gillette, Conney, Cosmides, Estabrook, Fours, and Mannering, t969) and insects (Hodgson and Plapp, I97O ). These mixed-function oxidases are present in microsomes which are primarily composed of fragments of endoplasmic reticulum (Siekevitz, I963). This system is known to require NADPH and molecular oxygen for activity. Whereas NADPH is known to interact with NADPH-cytochrome C-reductase (E.C.x.6.2.3), molecular oxygen is utilized by cytochrome P450, in hydroxylation of xenobiotics (Brodie, Gillette, and LaDu, x958; Gillette and others, I969; and Kamin and Masters, I968 ). Since its first isolation by Horecker (i95o) NADPH-cytochrome C-reductase has been purified and characterized (Williams and Kamin, 1962; Phillips and Langdon, 196z; Nishibayashi, Omura, and Sato, I963; Shuster, i964; Masters, Bilimoria, Kamin, and Gibson, 1965; Masters, Kamin, Gibson, and Williams, I965; and Iyanagi and Yamazaki, i969).
In insects, Lang (x959, I96z ) reported NADPH-cytochrome C-reductase for the first time in mosquitoes. Recently a number of excellent reports have appeared on this enzyme system in the housefly (Folsom and Hodgson, i97o; Wilson and Hodgson, I97Ia, b). It has been shown that the insect enzyme is a flavoprotein and its interaction with NADPH, cytochrome C, and certain specific inhibitors is also essentially the same as that of the mammalian enzyme.

264 AHMAD AND FORGASH Insect Biochem.
In this paper NADPH-oxidation in gypsy moth Porthetria dispar Linn. is reported. Gypsy moth is a serious pest of forest trees in several north-eastern states of the U.S.A., and nothing is known concerning the electron transport chain in the microsomes of this insect species. Therefore this report on NADPH oxidation and NADPH-cytochrome C- reductase in gypsy moth larvae should be of interest to comparative biochemists and toxicologists in their study of microsomal metabolism of pesticides in insects in general and gypsy moth in particular.
MATERIALS AND M E T H O D S
CHEMICALS
Pyridine, CuCI~, MgC12, KCN, and KC1 were obtained from Fisher Scientific Company, Fairlawn, N.J. Aniline was supplied by J. T. Baker Co., Phillipsburg, N.J., and 2-diethyl- amino-ethyl-2-diphenyl valerate (SKF 525-A) was supplied by Smith, Kline & French, Phila- delphia, Pa. Cytochrome C, reduced form of Triphosphopyridine Nucleotide (NADPH), and p-chloromercuribenzoic acid and p-chloromercuriphenyl sulphonic acid were obtained from Sigma Chemical Co., St. Louis, Mo. All the reagents were of the highest purity commercially available.
INSECT MATERIAL Third-instar larvae of gypsy moth were supplied by the New Jersey Department of Agriculture,
Beneficial Insect Laboratory, Trenton, N.J. The larvae were held at 27 ° C. and 55 per cent R.H. Non-sexed, fifth-instar larvae from this stock were used for tissue preparations.
ISOLATION OF TISSUES
The larvae were dissected in ice-chilled, l ' i 5 per cent (w/v) KCI solution and their Malpighian tubules, alimentary canals, and fat bodies were removed. In some experiments the canals were divided into three portions. The first segment, henceforth referred to as foregut, consisted of the oesophageal duct and the anterior stomach (also considered to be oesophageal in origin--Forbush and Fernald, 1896). The remaining alimentary canal was divided into two portions at the junction of the Malpighian tubules. The anterior part, consisting of an extensive stomach and a portion of intestine, is presently referred to as 'midgut ', whereas the third segment composed of posterior intestinal segment and rectum, is designated 'h indgut ' .
Undigested food and excreta were expelled from gut fragments by means of a rolling motion of the finger.
PREPARATION OF MICROSOMES Tissues from 5o larvae were washed twice with O.l M phosphate buffer (O'l M KH2PO4
adjusted to pH 7"4 by o-i M NaOH) containing o'25 M sucrose. The tissues were homogenized in io ml. of this buffer for 2 minutes at 12oo r.p.m., with a Teflon R pestle fitted to a Craftsman ¼-in. drill (Roebuck & Co. ; Simpsons-Sears Ltd.). An ice jacket was used to keep the homogenate cool. The homogenates were centrifuged for 15 minutes at 2 ° C. and at IO,OOO g in a Lourdes Model LRA refrigerated centrifuge. The sediments consisting of cellular debd, nuclei, and mitochondria were discarded. The supernatants were again centrifuged for 6o minutes at 2-5 ° C. and at lOO,OOO g in the type 4 o rotor of a Spinco Model L preparative ultracentrifuge. Each microsomal pellet thus obtained was resuspended in 5 ml. of O.l M phosphate buffer adjusted to pH 7"4.
The protein concentration of each fraction was determined by the method of Lowry, Rosebrough, Farr, and Randall (1951) as modified by Miller (1959).
INCUBATION MIXTURES The standard incubation mixture contained the microsomal fraction and NADPH in a total
volume of 2"5 ml. of o ' i M phosphate buffer adjusted to pH 7"4. In the experiments on the effects of enzyme concentration the enzyme was varied between o'o5 and o.8 mg. of microsomal protein. In all other experiments, enzyme concentration equivalent to o'6 rag. protein was
I973, 3 MICROSOMAL NADPH OXIDATION IN GYPSY MOTH 265
used. NADPH concentration was 0"33 pmoles per 2"5 ml. In experiments on the effect of substrate concentration, the levels of NADPH were varied between o'175 and o'5 gmoles. In some experi- ments o.i pmole of freshly prepared cytochrome C was added to the incubation mixture.
All incubations were carried out at 3 °0 C. with constant shaking, and the reaction rate was measured by recording absorbance at 34o nm. in a Beckman DU spectrophotometer.
INHIBITION STUDIES Compounds insoluble in water were prepared according to the method of Folsom and Hodgson
(197o). The effect of carbon monoxide was determined by first adding o'3 pmoles of NADPH to a cuvette containing a microsomal suspension (=o.5 mg. protein) and phosphate buffer to a final volume of 2"5 ml. Carbon monoxide was bubbled through the cuvette for 5 mintues before
.075 e m
o x
.1- a. ,05 a .< Z ¢) o . 0 2 5 E
O
dkl
m ~ o ® g g g m • i . , .o ,.o _ m m "o
. ~ = . o 2 "o c ~ • o . . . .
Tissues
- . 7 5
0
e. o m
i t m
O s _
5 cc
0} E
2 5
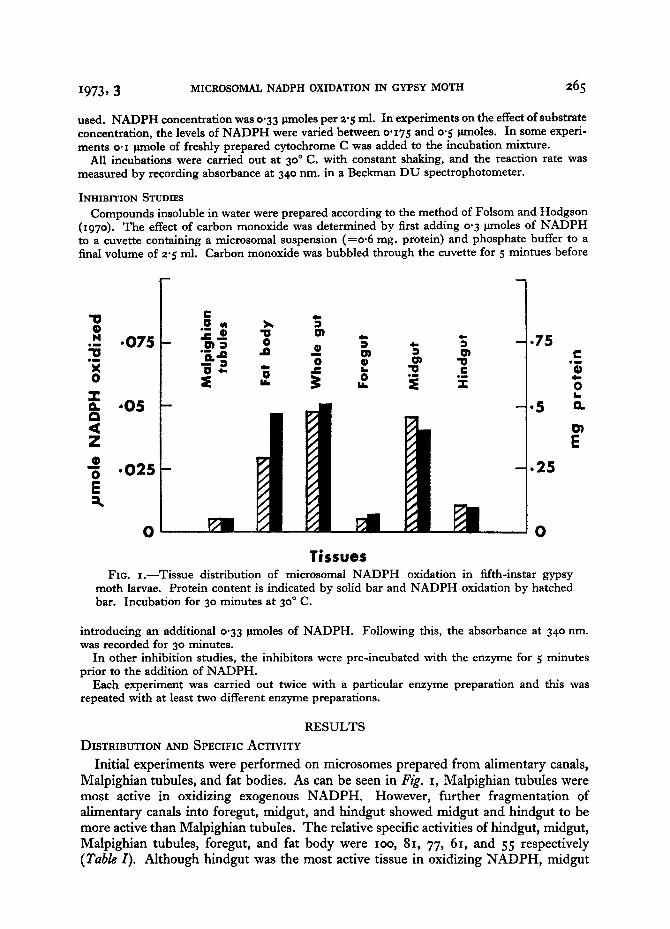
Fxo. i.--Tissue distribution of microsomal NADPH oxidation in fifth-instar gypsy moth larvae. Protein content is indicated by solid bar and NADPH oxidation by hatched bar. Incubation for 30 minutes at 3o ° C.
introducing an additional 0"33 pmoles of NADPH. Following this, the absorbance at 34 °nm. was recorded for 3o minutes.
In other inhibition studies, the inhibitors were pre-incubated with the enzyme for 5 minutes prior to the addition of NADPH.
Each experiment was carried out twice with a particular enzyme preparation and this was repeated with at least two different enzyme preparations.
RESULTS
DISTRIBUTION AND SPECIFIC ACTIVITY
Initial experiments were performed on microsomes prepared from alimentary canals, Malpighian tubules, and fat bodies. As can be seen in Fig. i, Malpighian tubules were most active in oxidizing exogenous N A D P H . However, further fragmentation of alimentary canals into foregut, midgut, and hindgut showed midgut and hindgut to be more active than Malpighian tubules. The relative specific activities of hindgut, midgut, Malpighian tubules, foregut, and fat body were ioo, 8x, 77, 6I, and 55 respectively (Table I). Although hindgut was the most active tissue in oxidizing N A D P H , midgut
266 AHMAD AND FORGASH Insect Biochem.
was found to be the major site of oxidation because of the relative abundance of micro- somal protein (single larval protein yield; hindgut microsomes o.o 9 mg., and midgut microsomes o. 4 mg.). The distribution of activity in various tissues was as follows:
Table I . - - N A D P H OXIDATION IN GYPSY MOTH LARVAE; SPECIFICITY AND DISTRIBUTION IN VARIOUS TISSUES
Tissue
Fat body Foregut Malpighian tubules Midgut Hindgut Total tissues
SPECIFIC ACTIVITY *
RELATIVE SP. ACTIVITY
0"042 0.046 0"058 0-068 0"075
55 6x 77 8i
I 0 0
PERCENTAGE OF TOTAL ACTIVITY t
27"8 4"3 4"4
53"9 9"6
I O O ' O
* ~tmoles NADPH oxidized/o'6 mg. protein/3o minutes at 3 °0 C. t NADPH oxidation/tissue as percentage of total NADPH oxidation/larva.
"~ . 0 6 -
• 0 5 -
..o . o 4 - "U O N
e m
• ~ - . 0 3 X 0
Z B_ . 0 2 a < Z Q .01
m
0 E :~ 0 i
0
_1 I I J
10 2 0 3 0 4 0
"U
N o m "U
, e m
X O
2: L t~ < Z
O m
O E
a .
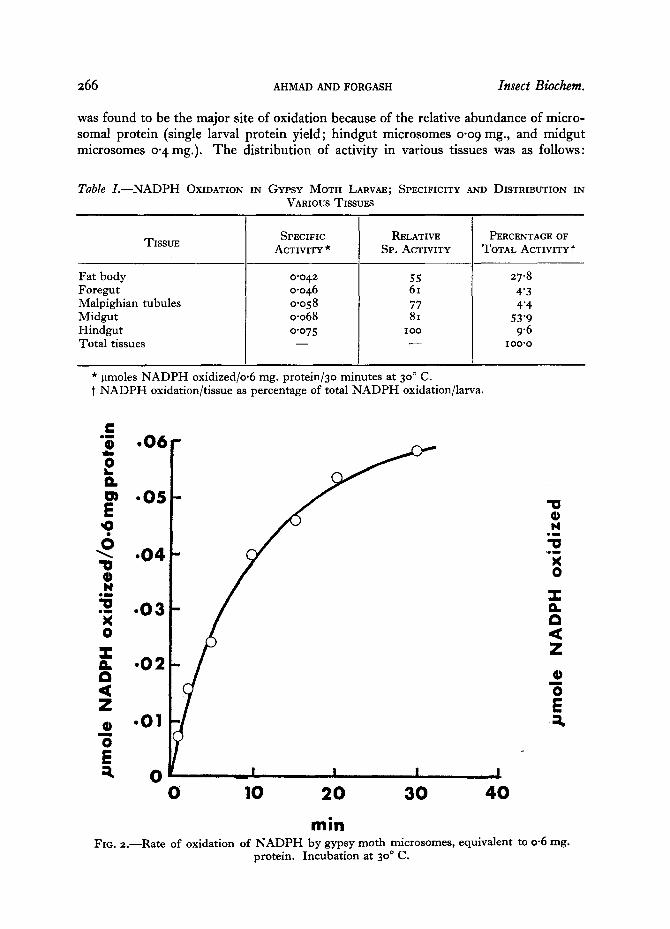
r a i n FIG. 2.--Rate of oxidation of NADPH by gypsy moth microsomes, equivalent to 0.6 rag.
protein. Incubation at 30 ° C.
1973, 3 MICROSOMAL NADPH OXIDATION IN GYPSY MOTH 267
midgut 53"9 per cent, fat body 27"8 per cent, hindgut 9-6 per cent, Malpighian tubules 4"4 per cent, and foregut 4"3 per cent (Table I).
ENZYME KINETICS Owing to the relative abundance of microsomal proteins in the midgut, enzyme
kinetics and effects of inhibitors were studied using microsomal preparations of this tissue. Rate of oxidation of NADPH by this enzyme preparation is illustrated in Fig. 2. The rate was rapid for the first 2o minutes of the incubation. Studies on effects of
"0 e 14
e m m
"0 o m m
x 0
3: a_ a < Z e
m
E
. 0 7 5
. 0 5
. 0 2 5
0 0
r m
I I I
• 25 .5 .75 1.O
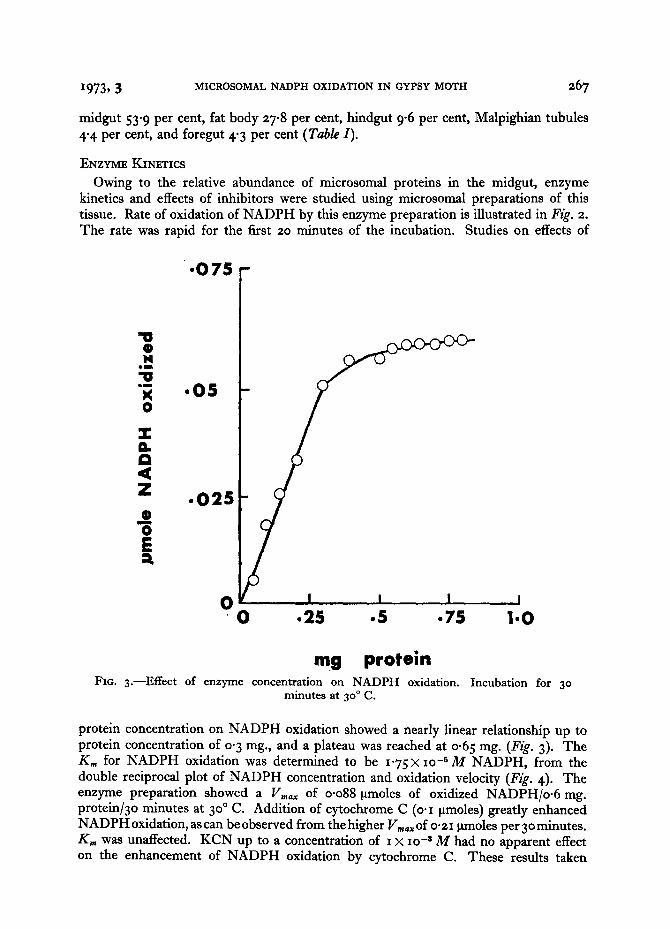
mg pro te in FIG. 3 . - -Ef fec t of enzyme concen t ra t ion on N A D P H oxidat ion.
m i nu t e s at 3 °o C. I n c u b a t i o n for 30
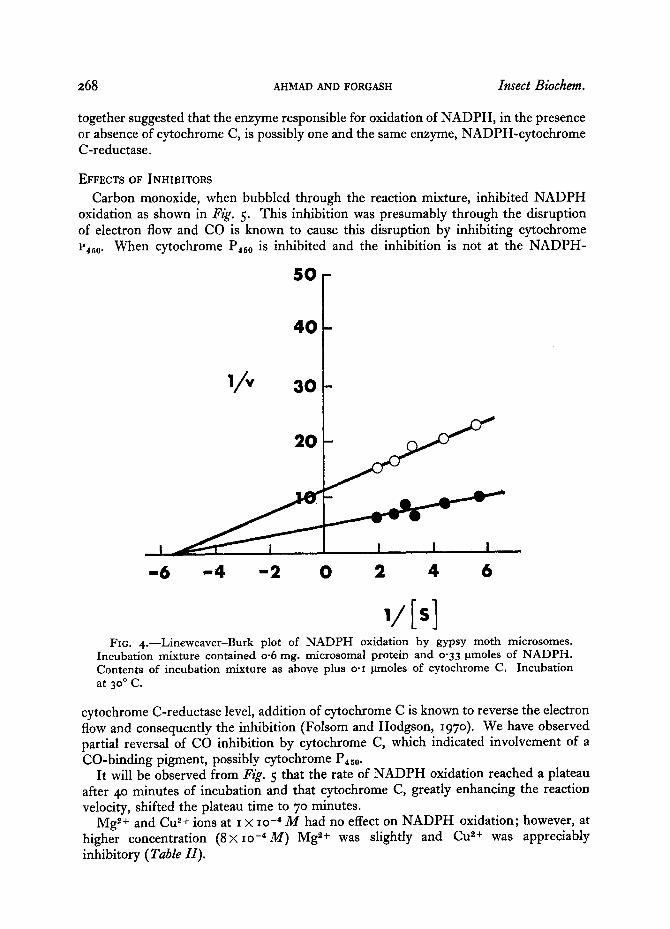
protein concentration on NADPH oxidation showed a nearly linear relationship up to protein concentration of 0. 3 ms., and a plateau was reached at o.65 rag. (Fig. 3). The K,~ for NADPH oxidation was determined to be 1-75 × lO -5 M NADPH, from the double reciprocal plot of NADPH concentration and oxidation velocity (Fig. 4). The enzyme preparation showed a Vmo~ of o.o88 ~tmoles of oxidized NADPH/o.6 ms. protein/3o minutes at 3 °o C. Addition of cytochrome C (o.I ~tmoles) greatly enhanced NADPH oxidation, as can be observed from the higher Vm°~ of 0.21 ttmoles per 3 ° minutes. K,, was unaffected. KCN up to a concentration of I × io -8 M had no apparent effect on the enhancement of NADPH oxidation by cytochrome C. These results taken
268 AHMAD AND FORGASH Insect Biochem.
together suggested that the enzyme responsible for oxidation of NADPH, in the presence or absence of cytochrome C, is possibly one and the same enzyme, NADPH-cytochrome C-reductase.
EFFECTS OF INHIBITORS
Carbon monoxide, when bubbled through the reaction mixture, inhibited NADPH oxidation as shown in Fig. 5. This inhibition was presumably through the disruption of electron flow and CO is known to cause this disruption by inhibiting cytochrome P~50. When cytochrome P45o is inhibited and the inhibition is not at the NADPH-
5 0 -
4 0
l/v ao
20 i J - 6 - 4 - 2 0 2 4 6
,/Is] FIG. 4.--Lineweaver-Burk plot of NADPH oxidation by gypsy moth microsomes.
Incubation mixture contained o-6 mg. microsomal protein and o'33 ~tmoles of NADPH. Contents of incubation mixture as above plus o.x ~tmoles of cytochrome C. Incubation at 3 °0 C.
cytochrome C-reductase level, addition of cytochrome C is known to reverse the electron flow and consequently the inhibition (Folsom and Hodgson, I97o ). We have observed partial reversal of CO inhibition by cytochrome C, which indicated involvement of a CO-binding pigment, possibly cytochrome P450.
It will be observed from Fig. 5 that the rate of NADPH oxidation reached a plateau after 4 ° minutes of incubation and that cytochrome C, greatly enhancing the reaction velocity, shifted the plateau time to 7 ° minutes.
Mg 2÷ and Cu ~+ ions at I × Io -4 M had no effect on NADPH oxidation; however, at higher concentration ( 8 × I o - 4 M ) Mg ~+ was slightly and Cu ~+ was appreciably inhibitory (Table II).
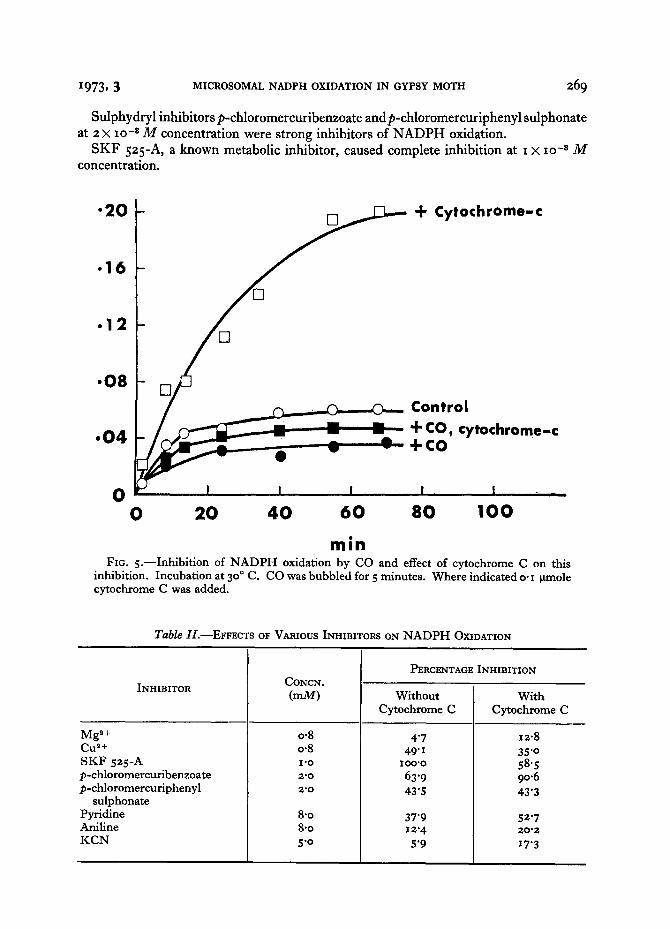
1973, 3 MICROSOMAL NADPH OXIDATION IN GYPSY MOTH 269
Sulphydryl inhibitorsp-chloromercuribenzoate andp-chloromercuriphenyl sulphonate at 2 × lO -3 M concentration were strong inhibitors of NADPH oxidation.
SKF 525-A , a known metabolic inhibitor, caused complete inhibition at I × lO -8 M
concen t ra t ion .
.20
.16
.12
.08
. 0 4
[] + Cytochrome-c
Control + CO, cytochrome-c +CO
0 ~ I I I I I
0 20 4 0 6 0 80 ! 0 0
min FIG. 5 . - - Inhibi t ion of N A D P H oxidation by CO and effect of cytochrome C on this
inhibition. Incubation at 3 o° C. CO was bubbled for 5 minutes. Where ind ica tedo . i Ixmole cytochrome C was added.
Table//.--EFFECTS OF VARIOUS INHIBITORS ON N A D P H OXIDATION
INHIBITOR
Mg2 + C a 2 +
S K F 525-A p-chloromereuribenzoate p-chloromereuriphenyl
sulphonate Pyridine Aniline K C N
CONCN. (mM)
0.8 o.8 I'0
2"0
2"0
8"0 8"0 5"0
PERCENTAGE INHIBITION
Without Cytochrome C
4"7 49"1
I00"0
63 "9 43"5
37"9 x2"4 5"9
With Cytochrome C
x2"8 35'0 58"5 90"6 43"3
52"7 20"2
x7"3

27o AHMAD AND FORGASH Insect Biochem.
Pyridine at 8x l O - 3 M caused 37'9 per cent inhibition, whereas aniline at the same concentration showed only I2"4 per cent inhibition and no inhibition at x × io -4 M concentration.
x × lO -3 M KCN showed no inhibition. However, at higher concentration (5x lO -3 M) slight inhibition (5"9 per cent) was apparent.
EFFECTS OF CYTOCHROME C ON INHIBITION
Cytochrome C appreciably reversed inhibition by SKF 525-A (Table 11) indicating cytochrome P450 at the end of the microsomal electron transport chain to be the site of SKF 525-A inhibition, rather than NADPH-cytochrome C-reductase. Some reversal of inhibition by Cu 2+ was also observed.
Cytochrome C did not reverse the inhibition by other compounds and in some cases the inhibition was enhanced.
DISCUSSION In mammals the liver is known to have a higher level of microsomal oxidation than
any other tissue. In insects there is no analogous organ, but the gut has been shown to have the highest activity in sixth-instar Prodenia eridania larvae (Krieger and Wilkinson, 1969), in Heliothis zea and Anthaerea polyphemus (Khan, 1969) , and in the housefly (Tsukamoto and Casida, 1967). The fat body is highest in Calliphora erythrocephala larvae (Price and Kuhr, 1969) and the Malpighian tubules in the cockroach, Periplaneta americana (Nakatsugawa and Dahm, i962 ).
Using NADPH oxidation as evidence for mixed-function oxidases, it has been shown in the present study that this enzyme system is present in the gut, fat body, and Mal- pighian tubules of fifth-instar larvae of gypsy moth. On the basis of observed specific rates, hindgut appeared to be the most active tissue, whereas, on the basis of total activity, midgut appeared to be the principal site of mixed-function oxidase activity.
The present study on gypsy moth NADPH-cytochrome C-reductase and earlier reports of the occurrence of this enzyme in mosquitoes (Lang, i959, 1961 ) and in the housefly (Folsom and Hodgson, i97o; and Wilson and Hodgson, 197ia , b) broadens our knowledge of this enzyme in insects and strengthens our assumption that this enzyme may be of universal occurrence in insects as in mammals. The observed Km value of 1.75 × lO -5 M NADPH for gypsy moth enzyme is very close to 1.8 × lO -5 M NADPH for housefly (Folsom and Hodgson, 197o ) and 2. 5 × lO -5 M for rat liver (Ernster and Orrenius, 1965). Thus this enzyme not only appears to be universally present but it exhibits similar kinetics.
Oxidation of NADPH to NADP+ and subsequent reduction of hemeprotein (Fe a+-- Protein) are mediated by NADPH-cytochrome C-reductase, in the electron transport chain (Kamin and Masters, 1968 ). Apart from its natural substrate in the electron transport chain, NADPH-cytochrome C-reductase is capable of reducing several dyes, ferricyanides, and inorganic ions and derives its name through its ability of rapidly reducing cytochrome C. It is therefore conceivable that addition of an exogeneous electron acceptor such as cytochrome C should enhance the rate of electron flow from NADPH. This appeared to be so with gypsy moth enzyme preparation, and under our experimental conditions, NADPH oxidation upon addition of cytochrome C was enhanced by a factor of 2. 4 (Vmax ratio of NADPH oxidation, with and without the addition of cytochrome C).

1973, 3 MICROSOMAL NADPH OXIDATION IN GYPSY MOTH 27I
NADPH oxidation was inhibited by CO in a way similar to that reported for the housefly. According to Folsom and Hodgson (197o) CO inhibition indicates electron flow via a CO-binding pigment, presumably cytochrome P450. Our data on the reversal of CO inhibition by an exogeneous electron acceptor, cytochrome C, indicate that this acceptor received electrons prior to cytochrome P45o in the electron transport chain in the gypsy moth.
The inhibition of NADPH oxidation by p-chloromercuribenzoate and p-chloro- mercuriphenyl sulphonate indicates that NADPH-cytochrome C-reductase is sensitive to - -SH inhibition. This agrees with previous work on mammals (Masters, Kamin, and others, 1965). Furthermore, the inhibition by p-chloromercuribenzoate and p-chloromercuriphenyl sulphonate was not reversed by exogeneous cytochrome C and, therefore, these inhibitors appear to act at the NADPH-cytochrome C-reductase level. Our results and conclusions are in agreement with the work of Folsom and Hodgson (197o) on housefly, with the exception of our data on Cu 2+ inhibition. Cu ~+ is known to be an --SH inhibitor (Masters, Kamin, and others, 1965) and is therefore expected to inhibit NADPH-cytochrome C-reductase. We have observed Cu 2+ inhibition of NADPH oxidation by gypsy moth enzyme, and this inhibition was partially reversed by the addition of cytochrome C. Thus it would appear that Cu 2+ is acting at the cyto- chrome P450 level, rather than the NADPH-cytochrome C-reductase level. On the other hand, it is possible that Cu 2+ inhibits both NADPH-cytochrome C-reductase and cytochrome P45o, in which case cytochrome C will cause some reversal of Cu 2+ inhibition. In this context, it is interesting to note that in mammals sulphydryl inhibitors have been shown to be effective inhibitors of NADPH-cytochrome C-reductase (Masters, Kamin, and others, 1965) and to accelerate the breakdown of cytochrome P,5o to cytochrome P4~o (Cooper, Narasimhula, Rosenthal, and Estabrook, i965; Hansen and Hodgson, 1971 ). It is therefore conceivable that besides inhibiting of NADPH-cytochrome C-reductase, Cu 2+ ions at high concentration convert some of cytochrome P45o to the inactive cytochrome P420--and the reversal of inhibition that we observed with cyto- chrome C apparently corresponds to inactivation of some cytochrome P45o.
SKF 525-A, a known inhibitor of microsomal oxidations (Brodie and others, 1958 ) caused total inhibition of NADPH oxidation at I × IO -s M concentration. This inhibi- tion was substantially reversed by cytochrome C, indicating the locus of inhibition to be at cytochrome P450, which is known to interact with xenobiotics.
Our data (Table 11) indicate that inhibition by p-chloromercuribenzoate was intensified to some degree by the addition of cytochrome C. This increase in inhibition is not readily explainable, except as a procedural error.
In brief, our studies on NADPH oxidation have shown the gut to be the principal site of an active microsomal electron transport chain in gypsy moth larvae. Using certain specific inhibitors and substrates some information has been gained on the nature of NADPH-cytochrome C-reductase and indications are that a CO-binding pigment, presumably cytochrome P450, is present as a component of the electron transport chain.
ACKNOV~LEDGEMENTS
Paper of the Journal Series, New Jersey Agricultural Experiment Station, Rutgers--The State University, New Brunswick, New Jersey. This work was supported by McIntire-Stennis funds. The authors wish to thank Mr. W. M. Metterhouse and Mr. R. Chianese of the New

272 AHMAD AND FORGASH Insect Biochem.
Jersey Department of Agriculture, Beneficial Insect Laboratory, Trenton, New Jersey, for the supply of gypsy moth larvae.
REFERENCES
BRODIE, D. B., GILLETTE, J. R., and LADu, B. N. (I958), 'Enzymatic metabolism of drugs and other foreign compounds', ,4. Rev. Bioehem., 27, 427-454.
COOPER, D. Y., NARASIMHULA, S., ROSENTHAL, O., and ESTABROOK, R. W. (I965) , in Oxygenases and Related Redox Systems (ed. King, Mason, and Morrison), pp. 838-863. New York: Wiley.
ERNSTER, L., and Ommslus, S. (I965), ' Substrate-induced synthesis of the hydroxylating enzyme system of liver microsomes', Fedn Proc. Fedn Am. Socs exp. Biol., 24, 119o-1199.
FOLSOM, M. D., and HODGSON, E. (I97O), 'Biochemical characteristics of insect microsomes: NADPH oxidation by intact microsomes from the housefly, Musca domestica', Comp. Biochem. Physiol., 37, 3Ol-31°.
FORBUSH, E. H., and FERNALD, C. H. (I896), The Gypsy Moth Porthetria dispar (Linn.). Boston: Wright & Potter.
GILLETTE, J. R., CONNEY, A. H., COSMIDES, G. J., ESTABROOK, R. W., FOUTS, J. R., and MANNERINa, G. J. (eds.) (I969), Microsomes and Drug Oxidations. New York: Academic Press.
HANSEN, L. G., and HODGSON, E. (I97I), 'Inhibition of microsomal oxidases from the housefly', Pesticide Biochem. Physiol., I, lO9-119.
HODOSON, E., and PLAPP, F. W. (I97O), 'Biochemical characteristics of insect microsomes', J. agric. Fd Chem., 18, lO48-1o55.
HOm~CKER, B. L. (195o), 'Triphosphopyridine nucleotide-cytochrome-e-reductase in liver', J. biol. Chem., 183, 593-60o.
IYANAOI, T., and YAMAZAKI, I. (I969), 'One-electron-transfer reactions in biochemical systems. III. One-electron reduction of quinones by microsomal flavin enzymes', Bioehim. biophys. Acta, I729 37o-381.
KAMIN, H., and MASTERS, B. S. S. (I968), 'Electron transport in microsomes', in Enzymatic Oxidations of Toxicants (ed. Hodgson), pp. 5-26. Raleigh: North Carolina State University.
KHAN, M. A. Q. (1969), ' DDT-Dehydrochlorinase and aldrin-epoxidase activity in corn earworm and polyphemus moth larvae, and house fly adults',J, econ. Ent., 62, 723-725.
KRIEGER, R., and WILKINSON, C. F. (1969), 'Microsomal mixed-function oxidases in insects. I. Localization and properties of an enzyme system effecting aldrin epoxidation in larvae of the southern armyworm (Prodenia eridania)', Biochem. Pharmac., 18, 14o3-1415.
LANC, C. A. (I959), 'Cytochrome-c-reductase activities during development', Expl Cell Res., 179 516-518.
LANG, C. A. (I96I), 'TPNH-cytochrome-c-reductase in the mosquito', Fedn Proc. Fedn Am. Socs exp. Biol., 2o9 47.
LOWRY, O. H., ROSEBROUGH, N. J., FARR, A. L., and RANDALL, R. J. (195 I), 'Protein measurement with the Folin phenol reagent', J. biol. Chem., I939 265-275.
MASTERS, B. S. S., BILIMORIA, M. H., KAMIN, H., and GIBSON, Q. H. (I965), 'The mechanism of I- and 2-electron transfers catalyzed by reduced triphosphopyridine nucleotide-eytochrome- c-reductase', J. biol. Chem., 24o, 4081-4o88.
MASTERS, B. S. S., KAMIN, H., GIBSON, Q. H., and WILLIAMS, C. H., jun. (I965), ' Studies on the mechanism of microsomal triphosphopyridine nucleotide-cytochrome c reductase', J. biol. Chem., 2409 921-931.
MILLER, G. L. (1959), 'Protein determination for large number of samples', Analyt. Chem., 319 964.
NAKATSUGAWA, T., and DAHM, P. A. (I962), 'Activation of guthion by tissue preparations from the American cockroach', J. econ. Ent., 559 594-599.
NISHIBAYASHI, H., OMURA, T., and SATO, R. (I963), 'A flavoprotein oxidizing NADPH isolated from liver microsomes', Biochim. biophys. Acta, 67, 520-522.
PHILLIPS, A. H., and LANGDON, R. G. (1962), 'Hepatic triphosphopyridine nucleotide-cytochrome c reductase: Isolation, characterization, and kinetic studies', J. biol. Chem., 2379 2652-266o.
PRICE, G. M., and KuI-IR, R. J. (1969) , 'The metabolism of the insecticide carbaryl (l-naphthyl N-methylcarbamate) by fat body of the blowfly larva Calliphora erythrocephala', Bioehem. J., 1129 133-138.

I973, 3 MICROSOMAL NADPH OXIDATION IN GYPSY MOTH 273
SHUSTER, L. (x964) , 'Metabolism of drugs and toxic substances', .4. Rev. Biochem., 33, 57x-596. St~KEVtTZ, P. (x963), 'Protoplasm: Endoplasmic reticulum and microsomes and their properties',
.4. Rev. Physiol., 25, x5-4 o. TSUKAMOTO, M., and CASIDA, J. E. (x967) , 'Metabolism of methylcarbamate insecticides by the
NADPH z-requiring enzyme system from house flies', Nature, Lond., 213, 49-5 x. WXLLXAMS, C. H., jun., and KAMIN, H. (x962), 'Microsomal triphosphopyridine nucleotide-
cytochrome c reductase: isolation, characterization and kinetic studies', 3. biol. Chem., 237, 587-595.
WmSON, T. G., and HODCSON, E. (x97xa), 'Microsomal NADPH-cytochrome c reductase from the housefly, Musca domestlca, solubilization and purification', Insect Biochem., Is x9-26.
WXLSON, T. G., and HODGSON, E. (x97xb), 'Microsomal NADPH-cytochrome c reductase from the housefly, Musca domestica: Properties of the purified enzyme', Insect Biochem., x, x 7 x-x 8o.
Key Wordlndex: CO inhibition, cytochrome C, cytochrome P4s0, E.C.x.6.2.3, electron transport chain, gypsy moth, insect microsomes, microsomal oxidations, mixed-function oxidases, NADPH oxidation, NADPH-cytochrome C-reductase, Porthetria dispar Linn., SKF 525-A, sulphydryl inhibitors.

![[1-3] Microsomal Lipid... · Chem.-Biol. Interactions, 50 (1984) 361-366 Elsevier Scientific Publishers Ireland Ltd. Short Communication 361 MICROSOMAL LIPID PEROXIDATION AND OXIDATIVE](https://img.pdfslide.net/doc/110x75/6089787ce01a1042bc238926/1-3-microsomal-lipid-chem-biol-interactions-50-1984-361-366-elsevier.jpg)



























