Embed Size (px)
DESCRIPTION
farmacogenetica
Citation preview

The Pharmacogenomics Journal (2002) 2, 30–42 2002 Nature Publishing Group All rights reserved 1470-269X/02 $25.00
www.nature.com/tpj
Received: 6 June 2001
Revised: 4 September 2001
Accepted: 7 September 2001
REVIEW
Pharmacogenetics of the arylamineN-acetyltransferases
NJ Butcher1,2
S Boukouvala3
E Sim3
RF Minchin1,2
1Laboratory for Cancer Medicine, WesternAustralian Institute for Medical Research, RoyalPerth Hospital, Perth, Western Australia;2Department of Pharmacology, University ofWestern Australia, Nedlands, WesternAustralia; 3Department of Pharmacology,University of Oxford, Oxford, UK
Correspondence:Dr NJ Butcher, Laboratory for CancerMedicine, Level 6, Medical ResearchFoundation Building, Royal Perth Hospital,Perth, Western Australia 6000Tel: +61 8 9224 0338Fax: +61 8 9224 0322E-mail: nbutcher�receptor.pharm.uwa.edu.au
ABSTRACTThe arylamine N-acetyltransferases (NATs) are involved in the metabolism ofa variety of different compounds that we are exposed to on a daily basis.Many drugs and chemicals found in the environment, such as those in ciga-rette smoke, car exhaust fumes and in foodstuffs, can be either detoxifiedby NATs and eliminated from the body or bioactivated to metabolites thathave the potential to cause toxicity and/or cancer. NATs have been impli-cated in some adverse drug reactions and as risk factors for several differenttypes of cancers. As a result, the levels of NATs in the body have importantconsequences with regard to an individual’s susceptibility to certain drug-induced toxicities and cancers. This review focuses on recent advances in themolecular genetics of the human NATs.The Pharmacogenomics Journal (2002) 2, 30–42. DOI: 10.1038/sj/tpj/6500053
Keywords: N-acetyltransferase; NAT; acetylation; arylamine; polymorphism
IntroductionThe arylamine N-acetyltransferases (NATs) are found in nearly all species frombacteria to humans. They catalyse the acetyltransfer from acetylcoenzyme A toan aromatic amine, heterocyclic amine or hydrazine compound. In humans,acetylation is a major route of biotransformation for many arylamine and hydra-zine drugs, as well as for a number of known carcinogens present in the diet,cigarette smoke and the environment.1–4 The reaction pathway is catalysed bytwo cytoplasmic acetyltransferases (NAT; EC 2.3.1.5), N-acetyltransferase Type I(NAT1) and N-acetyltransferase Type II (NAT2). The genes encoding both pro-teins were first isolated in 1989 by Grant et al who showed that each consistsof an intronless open reading frame of 870 base pairs.5 The two genes are 87%homologous and are located at 8p22,6–8 a chromosomal region commonlydeleted in human cancers.9–13 Ohsako and Deguchi isolated the transcript foreach gene from a human liver cDNA library in 1990.14 Sequencing of NAT1 andNAT2 revealed a number of allelic variants that affect activity of both genes invivo. This work provided a genetic understanding for the long known functionalpolymorphism in NAT2 activity15 and, more recently, in NAT1 activity.16–18 Gen-etic variation modulates the acetylator status of individuals and therefore mayimpact upon their predisposition to toxicity and disease. This review focuses onthe genomics of the arylamine N-acetyltransferases and the impact of geneticvariation of the enzymes on drug response in humans.
THE ARYLAMINE N-ACETYLTRANSFERASE GENE FAMILYTo date, 22 NAT-like genes have been identified in 14 different prokaryotic andeukaryotic species, although it is likely that additional genes will be discoveredas more genomes become sequenced. The genes invariably have an intronlesscoding sequence that encodes for a protein between 254 and 332 amino acids

Pharmacogenetics of the arylamine N-acetyltransferasesNJ Butcher et al
31
in length (Figure 1). Figure 1 shows a multiple sequencealignment for 22 NAT proteins. The highest conservedregions occur at the amino terminus, whereas the carboxylterminus shows very little conservation between species.Consistent with the recently published crystal structure forthe Salmonella typhimurium NAT,19 all NATs possess a con-served cysteine, histidine and aspartate that have beenimplicated to form a catalytic triad. Inhibitor studies20 andsite-directed mutagenesis studies21 have confirmed that thecysteine (Cys68 in the human proteins) is crucial for NATactivity.
A phylogenetic tree for the NAT proteins is shown inFigure 2. It indicates the separate clustering of the prokary-otic and eukaryotic sequences, with the exception of A. med-iterranei NAT which is distant from both groups. Thisenzyme is part of the rifamycin B synthesis pathway andcatalyses the final amide bond formation reaction.22 TheNAT1 and NAT2 sequences for rat, mouse and hamster clus-ter together suggesting that the two proteins are encoded bygenes that were present before the divergence of the threerodent species. By contrast, the two human proteins aremore closely related to each other, perhaps because thegenes duplicated later in evolution. The general relationshipof the amino acid sequences is similar to that recently pub-lished for a limited number of NAT nucleotide sequences.23
STRUCTURAL CHARACTERISTICS OF THE NATPROTEINSThe first crystal structure of an arylamine N-acetyltransferasewas recently published.19 Although the NAT was of bacterialorigin, it revealed a number of surprising features that pro-vided novel structural and functional information about theenzyme.19 Specifically, a cysteine-histidine-aspartate cata-lytic triad was identified in the N-terminus of the protein.Based on structural analysis, the protein has been dividedinto three domains. The first consists of a helical bundle,located from amino acid 1 to approximately 90 (based onnumbering in Figure 1), which forms one side of a cleft inwhich the cysteine involved in acetyl transfer resides. AllNATs are highly homologous in this region. The seconddomain consists of residues from approximately 90 to 210and is located on the other side of the cleft. It mostly con-sists of �-sheet structures. The last domain at the carboxylterminus is a combination of �-sheets and �-helices, andthis region shows the greatest diversity between species. Thestructural features surrounding the triad are similar to thecysteine protease superfamily of proteins which includes thetransglutaminases, cathepsins and caspases. While theseproteins traditionally catalyse the hydrolysis of amide sub-strates, the NATs and the transglutaminases catalyse an acyl-transfer that results in amide bond formation. This is alsothe case for the NAT homolog found in A. mediterranei.22 Todate, the crystal structure of the human NATs has not beenresolved although the homology with NAT from bacteriasuggests similar features will be present.
www.nature.com/tpj
N-ACETYLTRANSFERASE NOMENCLATUREThe first attempt to devise a consensus nomenclature for theNATs was published in 1995 and included genes from sixspecies and a total of 39 alleles.15 Since then, many newalleles have been identified and the nomenclature for thesewas updated in 2000.24 The Human Gene NomenclatureCommittee has agreed that the symbol NAT be assigned tothe arylamine N-acetyltransferase genes. Currently, NATalso is used for unrelated genes such as yeast protein N-ter-minal acetyltransferase, human noradrenalin transporter,eukaryotic translation initiation factor, translation repressorprotein and death associated protein 5.
The classification of the eukaryotic genes into NAT1 andNAT2 subfamilies has been done largely on a historical basisof the research area and is a consensus nomenclature.24
While this has been appropriate for most members of thegenes for NAT, it has raised some confusion with the mouse,hamster and rat Nat2 genes, which encode for proteins withsubstrate specificity similar to human NAT1 (acetylate p-aminobenzoic acid;25–28). At this stage, mouse is the onlyspecies to have three genes for NAT, pseudogenes excepted.Classification of different alleles into different clusters isbased on the most significant nucleotide substitutionpresent.24
The prokaryotic genes are sufficiently dissimilar to theeukaryotic genes for NAT to preclude subclassification.Consequently, the prokaryotic genes are referred to as Natonly. An international committee has been established tooversee the nomenclature of the N-acetyltransferases. Thecommittee is responsible for nomenclature updates andassignment of new alleles. This has been found to be essen-tial to ensure consistence of allelic names in the literature.A Web site that provides information about the naming ofexisting and new alleles for all species can be found athttp://www.louisville.edu/medschool/pharmacology/NAT.html.
REACTION MECHANISM AND SUBSTRATESPECIFICITYThe arylamine N-acetyltransferases differ from the manyother acetyl coenzyme A-dependent transferases present incells because of their ping-pong bi bi reaction mech-anism.29,30 The reaction takes place in two separate steps.Initially, acetyl coenzyme A binds to the enzyme and theacetyl moiety is transferred from the cofactor to a cysteine(Cys68 for the human isoforms) of the protein. Coenzyme Ais then released. The second step involves the binding ofsubstrate to the acetylated enzyme following which the ace-tyl moiety is transferred to the substrate. Finally, the acetyl-ated product is released from the enzyme. The first step ofthe reaction can proceed in the absence of arylamine sub-strate.31 It may be that the enzyme exists in an acetylatedstate in the cell when no substrate is present, although thishas not been shown to date, and the stability of the acetyl-ated intermediate should be assessed.
There is no clear structural motif that determines substratespecificity for the different isoforms of NAT. In general, p-aminobenzoic acid (PABA), p-aminobenzoyl glutamate and

Pharmacogenetics of the arylamine N-acetyltransferasesNJ Butcher et al
32
Figure1ClustalXmultiplesequencealignmentof22NATproteinsfrom14differentspecies.Aconservationscoreforeachcolumnisshownbythehistogram.Theshaded
columnsrepresentthecatalytictriadwhichisconservedinallNATs.Thethreedomains,identifiedfromthecrystalstructureofNATfrom
S.ty
phim
uriu
m,areshownas
shadedbarsbelowthehistogram.
The Pharmacogenomics Journal

Pharmacogenetics of the arylamine N-acetyltransferasesNJ Butcher et al
33
Figure 2 Phylogenetic tree for the NATs determined from the
amino acid alignments shown in Figure 1. The dendrogram shows
the relative distance between each sequence on the horizontal axis.
All sequences were acquired from GenBank including the chicken
NATX (accession number J03737) which, to date, has not been
assigned a name.
p-aminosalicylic acid (PAS) are considered specific substratesfor human NAT1 (or mouse NAT2). These substrates can becharacterised by the presence of relatively small hydrophilicsubstitutions in the para position of the aromatic ring. Bycontrast, sulfamethazine, procainamide and dapsone areacetylated primarily by human NAT2. Some compoundssuch as 2-aminofluorene are excellent substrates for bothhuman NAT1 and NAT2.3
The prokaryotic NATs acetylate PABA poorly or not at alland their range of substrates, which includes isoniazid, sug-gests that they are functionally similar to the human NAT2isoform.32 However, the prokaryotic NATs are also capableof acetylating the human NAT1 specific substrate 5-amino-salicylic acid,33 indicating that prokaryotic NATs should notbe considered functionally equivalent to either NAT1 orNAT2 in humans. In some bacterial strains such as H. pylori,
www.nature.com/tpj
NAT activity has been reported although there is no evi-dence of a NAT-like gene in their genome. The very high Km
values reported for substrates like PABA and 2-aminofluor-ene indicate that the acetylation reaction is very inefficientand is probably catalysed by a protein unrelated to theNATs.34
One prokaryotic NAT homolog deserves further attention.The Rif gene cluster in A. mediterranei that encodes the pro-teins necessary for the synthesis of rifamycin B includes thegene RifF for rifamycin amide synthase. The amino acidsimilarity of RifF to the other prokaryotic NATs is approxi-mately 45% (Figure 1) and the enzyme possesses the con-served catalytic triad in its amine terminus. RifF catalyses anamide bond formation during the latter stages of rifamycinsynthesis (Figure 3).22 Other NAT-like proteins may becomeevident as more genomes are sequenced and it is possiblethat the NAT protein has evolved in different species toundertake quite different cellular tasks.32
GENE LOCALISATION, STRUCTURE AND EXPRESSIONTwo NAT isoenzymes have been identified in humans,namely NAT1 and NAT2, which are the products of distinctgenetic loci, designated NAT1 and NAT2, respectively.6 Arelated pseudogene, NATP1, has also been identified,6 whichcontains multiple frameshift and nonsense mutations. Thetwo functional NAT genes share an 87% nucleotide identity,which translates to an 81% homology at the amino acidlevel. While the entire transcript of NAT1 is derived from asingle exon, that of NAT2 is derived from the protein enco-ding exon together with a second noncoding exon of 100 bplocated about 8 kb upstream of the translation start site.6,35
Human NAT1 and NAT2, as well as NATP1, have been local-ised to the short arm of chromosome 8,6,36 more specificallyin region 8p22.8 The NAT loci are separated by only 170–360 kb and are in the orientation NAT1 → NATP1 → NAT2,with NAT1 being on the centromeric side of marker D8S261and NAT2 coinciding with marker D8S21.36 Both NAT1 andNAT2 genes display pronounced allelic variation, with 26different human NAT1 and 29 different human NAT2 allelesidentified to date.24
A similar situation is present in the mouse where threefunctional genes, designated Nat1, Nat2 and Nat3, have
Figure 3 RifF, an NAT-like protein from Amycolatopsis mediterranei,catalyses an amide bond formation during the latter stages of rifa-
mycin synthesis.22 Although RifF is similar to the other prokaryotic
NATs, it has evolved to function specifically in the synthesis of rifa-
mycin B.

Pharmacogenetics of the arylamine N-acetyltransferasesNJ Butcher et al
34
been identified to encode for NAT isoenzymes.37,38 MouseNAT2 shows similar substrate specificity with human NAT1,while mouse NAT1 is capable of metabolising the humanNAT2-specific substrate isoniazid.39 To date, no specific sub-strate has been identified for NAT3, although the encodinglocus appears to be functional.38,40 The localisation and gen-omic organisation of the mouse Nat genes is similar to theirhuman counterparts. The three genes are clustered togetherin a 130-kb genomic region on mouse chromosome 8, cyto-genetic band B3.1-B3.341 and within a genetic distance ofabout 31 cM from the centromere.42 This chromosomalregion is syntenic with the region harbouring the genes forhuman NAT on chromosome 8p. Polymorphism has beendetected only in the Nat2 gene of both A/J37 and A/HeJ43
inbred mice, in the form of a missense A → T mutation atposition 296 of the open reading frame, causing the slowacetylator phenotype. It is of interest that mouse Nat2 alsopossesses a short non-coding exon, located about 6 kbupstream of the intronless open reading frame.41 This raisesthe possibility that mouse Nat2 may be the geneticorthologue of the human NAT2 gene, although mouseNAT2 and human NAT1 proteins appear to be func-tionally equivalent.
Information about the localisation and genomic organis-ation of the Nat genes in other eukaryotic species has beenlimited. All genes have an intronless open reading frame andpolymorphism in NAT activity has been described in strainsof rabbits,44 hamsters45 and rats,46 all of which possess twoNat genes. Upstream non-coding exons have also beendescribed for the rabbit Nat genes.47 Cats and other felidsonly have one gene for NAT,48 while dogs lack NAT activity,due to absence of Nat genes in their genome.49
HUMAN NATsHuman NAT2 Alleles
Since the human NAT2 locus was established as the site ofthe classical acetylation polymorphism,50,51 the study ofNAT2 allelic variation has been an area of intense investi-gation. To date, 29 different NAT2 alleles have been detectedin human populations (Table 1; reviewed in 15,24,52). Each ofthe variant alleles is comprised of between one and fournucleotide substitutions, of which 13 have been identified,located in the protein encoding region of the gene. Nine ofthese lead to a change in the encoded amino acid (C190T,G191A, T341C, A434C, G499A, G590A, A803G, A845C, and G857A),while the remaining four are silent (T111C, C282T, C481T,and C759T).
Several studies have been performed that show clear corre-lations between NAT2 genotype and phenotype.53–55 Earlygenotyping studies screened for the presence of the C481T(M1), the G590A (M2), the G857A (M3) and sometimes theG191A (M4) nucleotide changes, all of which were shown tocause a slow acetylation phenotype. Moreover, there was agene-dosage effect. Individuals who were homozygous forNAT2 polymorphisms had a slow acetylator phenotype,individuals heterozygous for NAT2 polymorphisms had anintermediate acetylator phenotype, and individuals wholacked NAT2 polymorphisms had a rapid acetylator pheno-
The Pharmacogenomics Journal
Table 1 Human NAT2 alleles (adapted from References 52
and 56)
Allele Phenotype Nucleotide Amino acidchange(s) change(s)
NAT2*4 Rapid None NoneNAT2*5A Slow T341C, C481T Ile114 → ThrNAT2*5B Slow T341C, C481T, A803G Ile114 → Thr,
Lys268 → ArgNAT2*5C Slow T341C, A803G Ile114 → Thr,
Lys268 → ArgNAT2*5D Slow T341C Ile114 → ThrNAT2*5E Slow T341C, G590A Ile114 → Thr,
Arg197 → GlnNAT2*5F Slow T341C, C481T, T759T, Ile114 → Thr,
A803G Lys268 → ArgNAT2*6A Slow C282T, G590A Arg197 → GlnNAT2*6B Slow G590A Arg197 → GlnNAT2*6C Slow C282T, G590A, A803G Arg197 → Gln,
Lys268 → ArgNAT2*6D Slow T111C, C282T, G590A Arg197 → GlnNAT2*7A Slow G857A Lys286 → GluNAT2*7B Slow C282T, G857A Lys286 → GluNAT2*10 Unknown G499A Glu167 → LysNAT2*11 Unknown C481T NoneNAT2*12A Rapid A803G Lys268 → ArgNAT2*12B Rapid C282T, A803G Lys268 → ArgNAT2*12C Rapid C481T, A803G Lys268 → ArgNAT2*13 Rapid C282T NoneNAT2*14A Slow G191A Arg64 → GlnNAT2*14B Slow G191A, C282T Arg64 → GlnNAT2*14C Slow G191A, T341C, C481T, Arg64 → Gln,
A803G Ile114 → Thr,Lys268 → Arg
NAT2*14D Slow G191A, C282T, G590A Arg64 → Gln,Arg197 → Gln
NAT2*14E Slow G191A, A803G Arg64 → Gln,Lys268 → Arg
NAT2*14F Slow G191A, T341C, A803G Arg64 → Gln,Arg114 → Thr,Lys268 → Arg
NAT2*14G Slow G191A, C282T, A803G Arg64 → Gln,Lys268 → Arg
NAT2*17 Slow A434C Gln145 → ProNAT2*18 Unknown A845C Lys282 → ThrNAT2*19 Slow C190T Arg64 → Trp
type. It should be noted that the method of detection of theabove polymorphisms only identifies a subset of the variantalleles found in human populations, and there is potentialfor the misclassification of genotype and deduced pheno-types (reviewed in 56).
Initial studies in liver tissue suggested that the slow acetyl-ator phenotype associated with the presence of certainnucleotide substitutions in the protein encoding region ofthe NAT2 gene was due to a marked decrease in NAT2 pro-tein content, while NAT2 mRNA levels remainedunchanged.57 Several studies have since investigated themechanism by which nucleotide substitutions in the NAT2gene affect acetylation capacity by the use of recombinant

Pharmacogenetics of the arylamine N-acetyltransferasesNJ Butcher et al
35
expression systems.58–62 Hein and coworkers63 performed acomprehensive study that assessed the acetylation capacityof 16 different NAT2 alleles in a bacterial expression system.Of the seven specific NAT2 substitutions that they exam-ined, the T341C, G590A, G857A, and G191A substitutions pro-duced recombinant NAT2 allozymes with reduced acety-lation capacities, while the C481T, C282T, and A803Gsubstitutions produced recombinant NAT2 allozymes withacetylation capacities similar to the wild-type NAT2 4 pro-tein. As a result, NAT2 alleles that contain any of the specificsubstitutions that produced recombinant NAT2 allozymeswith reduced acetylation capacities are associated with aslow acetylator phenotype, and include the NAT2 5, NAT26, NAT2 7, NAT2 14, and NAT2 17 clusters (see Table 1).
The molecular mechanisms responsible for the pro-duction of the slow acetylator phenotypes are not wellunderstood at present. Some base changes appeared to causea slow acetylation phenotype by producing an unstable pro-tein. NAT2 allozymes encoded by alleles with base substi-tutions at positions 191, 590, or 857 were found to be sig-nificantly more unstable in bacterial expression systemsthan the wild-type protein.52,61,64 However, in these studiesthe amount of immunodetectable NAT2 protein was not dif-ferent upon expression of the variant and wild-type alleles.This is in contrast to the earlier observations by Grant andcoworkers57 who showed that liver NAT2 content was mark-edly reduced in slow acetylators, suggesting that the arti-ficial environment of bacterial expression systems may notaccurately reflect what occurs in mammalian cells withregard to protein degradation.
Recently, a study by Leff and coworkers62 characterisedseveral different human NAT2 alleles in a yeast expressionsystem. They found that three novel alleles, namely NAT2*5D (T341C), NAT2*14G (G191A, C282T, and A803G), andNAT2*6D (C111T, C282T, and G590A), expressed proteins thathad N- and O-acetylation capacities similar to the expressedprotein of the commonly occurring slow NAT2*5B allele,and significantly less than that of the wild-type NAT2*4allele. The expression of NAT2 5B and NAT2 5D was foundto be significantly lower than that of the wild-type protein,suggesting that the base substitution at position 341, whichis common to the NAT2*5 cluster, is sufficient for reductionin NAT2 protein expression. This was not found to be thecase for NAT2 6D and NAT2 14G, which were expressed atlevels comparable to wild-type. By contrast, NAT2 6D andNAT2 14G were found to be significantly less stable thanwild-type.
The frequency of the slow acetylator phenotype variesconsiderably among ethnic groups,65 and this is due to thediffering frequencies of the polymorphisms that correspondto the slow acetylator alleles. In Caucasian and Africanpopulations, the frequency of the slow acetylation pheno-type varies between 40 and 70%, while that of Asian popu-lations, such as Japanese, Chinese, Korean, and Thai, rangefrom 10 to 30% (reviewed in 66). Caucasian and Africanpopulations have high frequencies of NAT2*5 alleles(�28%) and low frequencies of NAT2*7 alleles (�5%), whileAsian populations have low incidences of NAT2*5 alleles
www.nature.com/tpj
(�7%) and higher incidences of NAT2*7 alleles (�10%).Also, NAT2*14 alleles are almost absent from Caucasian andAsian populations (�1%), but are present in African popu-lations at comparably higher frequencies (�8%).
Human NAT1 Alleles
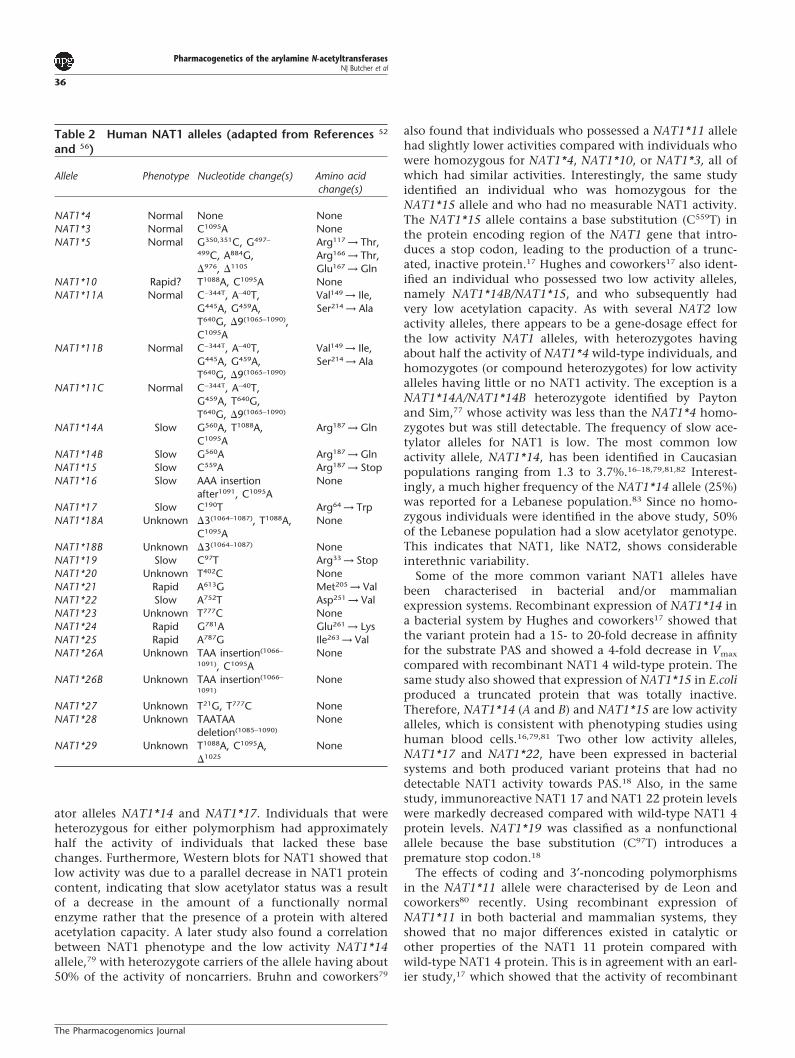
Historically, NAT1 was thought to be genetically invariantor monomorphic in nature. However, wide inter-individualvariability in NAT1 activity towards PABA or PAS51,67–72 wassuggestive of a genetic polymorphism, but NAT1 activitieswere generally unimodally distributed. It wasn’t until 1993when Vatsis and Weber73 first reported the existence of sev-eral allelic variations at the NAT1 locus that interest in theNAT1 gene was aroused, marking the beginning of a system-atic survey of NAT1 genotypes. To date, 26 different NAT1alleles have been detected in human populations (Table 2;reviewed in 15,24,52), however, only a small number havebeen shown to alter phenotype in vivo. Hughes and cowork-ers17 used PAS as a probe drug to phenotype a populationfor NAT1 activity. By measuring urinary metabolite ratios,they were able to detect individuals with marked impair-ments of NAT1 function. However, there was only a moder-ate correlation between phenotypes determined by in vivoand in vitro methods, and the authors themselves suggestthat less than 50% of the phenotypic variation observed invivo was related to variation in NAT1 function. It appearsthat the measurement of NAT1 activity of blood cells is themost reliable method of phenotyping for NAT1. While littleis known about the relative expression of NAT1 in varioushuman tissues, studies in the rabbit model suggest thatNAT1 activity is comparable in most tissues, including bloodcells.17 Therefore, it is reasonable to assume that NAT1activity of blood cells is a good surrogate of systemic activityin humans.
The first report of a correlation between NAT1 genotypeand phenotype was by Bell and coworkers in 1995.74 Theyshowed that the NAT1*10 allele was associated with activitytwo-fold higher than that of the wild-type allele, NAT1*4,in bladder and colon tissue samples.74 In the bladder, higherlevels of DNA adducts were detected in NAT1*10 hetero-zygotes compared with NAT1*4 homozygotes.75,76 TheNAT1*10 allele also has been associated with a marginallyelevated activity in liver samples4 and erythrocytes.77 NAT1*10 has no mutations in the protein encoding region of thegene, but contains two nucleotide substitutions (T1088Aand C1095A) in its 3′-untranslated region. The T1088A basechange alters the consensus polyadenylation signal(AATAAA → AAAAAA) leading to the suggestion thatincreased activity may be due to enhanced mRNA stab-ility.74,78 However, several recent studies,16–18,79,80 do notsupport the idea that the NAT1*10 allele is associated withelevated NAT1 activity. As a result, the functional signifi-cance of this allele remains unclear at present.
A population study16 showed a distribution of NAT1activity that was clearly bimodal in nature, with 8% of theindividuals being slow acetylators. Moreover, the abovestudy was one of the first to report a correlation betweenNAT1 genotype and phenotype involving the slow acetyl-
Pharmacogenetics of the arylamine N-acetyltransferasesNJ Butcher et al
36
Table 2 Human NAT1 alleles (adapted from References 52
and 56)
Allele Phenotype Nucleotide change(s) Amino acidchange(s)
NAT1*4 Normal None NoneNAT1*3 Normal C1095A NoneNAT1*5 Normal G350,351C, G497– Arg117 → Thr,
499C, A884G, Arg166 → Thr,�976, �1105 Glu167 → Gln
NAT1*10 Rapid? T1088A, C1095A NoneNAT1*11A Normal C−344T, A−40T, Val149 → Ile,
G445A, G459A, Ser214 → AlaT640G, �9(1065–1090),C1095A
NAT1*11B Normal C−344T, A−40T, Val149 → Ile,G445A, G459A, Ser214 → AlaT640G, �9(1065–1090)
NAT1*11C Normal C−344T, A−40T,G459A, T640G,T640G, �9(1065–1090)
NAT1*14A Slow G560A, T1088A, Arg187 → GlnC1095A
NAT1*14B Slow G560A Arg187 → GlnNAT1*15 Slow C559A Arg187 → StopNAT1*16 Slow AAA insertion None
after1091, C1095ANAT1*17 Slow C190T Arg64 → TrpNAT1*18A Unknown �3(1064–1087), T1088A, None
C1095ANAT1*18B Unknown �3(1064–1087) NoneNAT1*19 Slow C97T Arg33 → StopNAT1*20 Unknown T402C NoneNAT1*21 Rapid A613G Met205 → ValNAT1*22 Slow A752T Asp251 → ValNAT1*23 Unknown T777C NoneNAT1*24 Rapid G781A Glu261 → LysNAT1*25 Rapid A787G Ile263 → ValNAT1*26A Unknown TAA insertion(1066– None
1091), C1095ANAT1*26B Unknown TAA insertion(1066– None
1091)
NAT1*27 Unknown T21G, T777C NoneNAT1*28 Unknown TAATAA None
deletion(1085–1090)
NAT1*29 Unknown T1088A, C1095A, None�1025
ator alleles NAT1*14 and NAT1*17. Individuals that wereheterozygous for either polymorphism had approximatelyhalf the activity of individuals that lacked these basechanges. Furthermore, Western blots for NAT1 showed thatlow activity was due to a parallel decrease in NAT1 proteincontent, indicating that slow acetylator status was a resultof a decrease in the amount of a functionally normalenzyme rather that the presence of a protein with alteredacetylation capacity. A later study also found a correlationbetween NAT1 phenotype and the low activity NAT1*14allele,79 with heterozygote carriers of the allele having about50% of the activity of noncarriers. Bruhn and coworkers79
The Pharmacogenomics Journal
also found that individuals who possessed a NAT1*11 allelehad slightly lower activities compared with individuals whowere homozygous for NAT1*4, NAT1*10, or NAT1*3, all ofwhich had similar activities. Interestingly, the same studyidentified an individual who was homozygous for theNAT1*15 allele and who had no measurable NAT1 activity.The NAT1*15 allele contains a base substitution (C559T) inthe protein encoding region of the NAT1 gene that intro-duces a stop codon, leading to the production of a trunc-ated, inactive protein.17 Hughes and coworkers17 also ident-ified an individual who possessed two low activity alleles,namely NAT1*14B/NAT1*15, and who subsequently hadvery low acetylation capacity. As with several NAT2 lowactivity alleles, there appears to be a gene-dosage effect forthe low activity NAT1 alleles, with heterozygotes havingabout half the activity of NAT1*4 wild-type individuals, andhomozygotes (or compound heterozygotes) for low activityalleles having little or no NAT1 activity. The exception is aNAT1*14A/NAT1*14B heterozygote identified by Paytonand Sim,77 whose activity was less than the NAT1*4 homo-zygotes but was still detectable. The frequency of slow ace-tylator alleles for NAT1 is low. The most common lowactivity allele, NAT1*14, has been identified in Caucasianpopulations ranging from 1.3 to 3.7%.16–18,79,81,82 Interest-ingly, a much higher frequency of the NAT1*14 allele (25%)was reported for a Lebanese population.83 Since no homo-zygous individuals were identified in the above study, 50%of the Lebanese population had a slow acetylator genotype.This indicates that NAT1, like NAT2, shows considerableinterethnic variability.
Some of the more common variant NAT1 alleles havebeen characterised in bacterial and/or mammalianexpression systems. Recombinant expression of NAT1*14 ina bacterial system by Hughes and coworkers17 showed thatthe variant protein had a 15- to 20-fold decrease in affinityfor the substrate PAS and showed a 4-fold decrease in Vmax
compared with recombinant NAT1 4 wild-type protein. Thesame study also showed that expression of NAT1*15 in E.coliproduced a truncated protein that was totally inactive.Therefore, NAT1*14 (A and B) and NAT1*15 are low activityalleles, which is consistent with phenotyping studies usinghuman blood cells.16,79,81 Two other low activity alleles,NAT1*17 and NAT1*22, have been expressed in bacterialsystems and both produced variant proteins that had nodetectable NAT1 activity towards PAS.18 Also, in the samestudy, immunoreactive NAT1 17 and NAT1 22 protein levelswere markedly decreased compared with wild-type NAT1 4protein levels. NAT1*19 was classified as a nonfunctionalallele because the base substitution (C97T) introduces apremature stop codon.18
The effects of coding and 3′-noncoding polymorphismsin the NAT1*11 allele were characterised by de Leon andcoworkers80 recently. Using recombinant expression ofNAT1*11 in both bacterial and mammalian systems, theyshowed that no major differences existed in catalytic orother properties of the NAT1 11 protein compared withwild-type NAT1 4 protein. This is in agreement with an earl-ier study,17 which showed that the activity of recombinant

Pharmacogenetics of the arylamine N-acetyltransferasesNJ Butcher et al
37
NAT1 4 and NAT1 11 were similar, but is in contrast toanother study which reported a slightly reduced activity ofblood cells from individuals who carried the NAT1*11allele.79 de Leon and coworkers,80 using a mammalianexpression system, have recently shown that the NAT1*16phenotype is caused by polymorphism in the 3′-untrans-lated region that leads to a decrease in protein expression.NAT1*16 has a triple adenosine insertion on the 3′ side ofthe polyadenylation signal (AATAAA) which significantlyalters the secondary structure of the pre-mRNA, leading toa 2-fold reduction in the amount of NAT1 16 protein andactivity, compared with NAT1 4 and NAT1 10.
Some NAT1 alleles also may produce proteins with activi-ties that are higher than that of the wild-type protein NAT14. Recombinant expression of NAT1*21, NAT1*24, andNAT1*25 in bacterial systems produced allozymes withactivities 2- to 3-fold higher than NAT1 4.18 However, thelevels of immunoreactive protein expressed by NAT1*4,NAT1*21, NAT1*24, and NAT1*25 were equivalent.18 Thefunctional significance of the other NAT1 variants remainsunclear at present.
NAT AND DISEASEThe association between acetylator status and the risk ofvarious diseases has been extensively reported, and reviewedin detail.84–87 Altered risk with either the slow or rapidphenotype has been observed for bladder, colon and breastcancer, systemic lupus erythematosis, diabetes, Gilbert’s dis-ease, Parkinson’s disease and Alzheimer’s disease. Theseassociations imply a role for environmental factors that aremetabolised by the NATs, in particular NAT2, in each dis-order. However, identifying those factors has remained elus-ive. Humans are exposed to many toxic NAT substratesincluding the food-derived heterocyclics present in the dietas well as arylamines such as 4-aminobiphenyl and �-naph-thylamine present in tobacco smoke.88–91 Moreover, occu-pational exposure to arylamine carcinogens such as benzi-dine has also been reported.92,93
Because of the role of acetylation in the metabolic acti-vation and detoxification of arylamine and heterocyclic car-cinogens, acetylator status and cancer risk has been widelyinvestigated. Unlike the relatively rare but highly penetrantgenes involved in familial cancers, those genes responsiblefor metabolic polymorphisms have low penetrance andcause only a moderate increase in cancer risk. Nevertheless,their widespread occurrence in the general population sug-gests they are a significant contributor to individual risk. In1979, Lower et al94 first demonstrated an associationbetween the slow acetylator phenotype and bladder cancer.This work was followed by many independent studies. How-ever, few diseases have consistently demonstrated a relation-ship between phenotype and risk. For example, several stud-ies have implicated the rapid phenotype as an increased riskfactor for colon cancer,95–97 whereas others have beenunable to confirm this finding.98–100 Geographical differ-ences, ethnicity, lack of study power, dietary differences anddifferences in other risk factors between study groups havebeen suggested as reasons for variable results from inde-
www.nature.com/tpj
pendent laboratories. Recent reports suggesting that NATactivity may be altered by environmental factors and sub-strate-dependent down-regulation101 may also explain whyinconsistent associations have been seen.
When acetylator phenotype has been linked to carcino-gen exposure, more consistent results have been reported.For example, the rapid phenotype has emerged as a strongrisk factor for colorectal cancer in those individuals whohave a higher exposure to the food-derived heterocyclicamines.99,100,102 These observations provide strong circum-stantial evidence that the heterocyclic amines have animportant role in colorectal cancer, and extensive animalstudies support this.103–106 They also illustrate the impor-tance of establishing associations between genetic polymor-phisms and disease risk. From such studies, it should bepossible to pursue the causative agent(s) of the disease whereno obvious candidate agent is evident.
Recently, the NAT2 acetylator phenotype has been linkedto increased risk associated with neurodegenerative diseasessuch as Parkinson’s disease and Alzheimer’s disease.107–109
For late-onset Alzheimer’s disease, an odds ratio of 3.0 (95%CI 1.3–7.3) has been reported for the rapid phenotype innon-apoE epsilon carriers.109 By contrast, the slow pheno-type appears to increase risk of Parkinson’s disease with anodds ratio of 3.58 (95% CI 1.96–6.56).107 Although theseresults need to be confirmed with larger epidemiologicalstudies, they point to environmental factors that are sub-strates for the NATs having a role in the onset of these dis-eases. Alternatively, the different alleles for NAT2 that pro-duce the rapid or slow phenotype may co-segregate withunrelated genes that are the causative agent for the differentneurodegenerative diseases.
NAT and Drug Response
The genetic polymorphism in N-acetyltransferase activitywas first discovered in patients treated with isoniazid fortuberculosis.110 This drug is primarily excreted followingacetylation catalysed by NAT2. Since then, many thera-peutic agents have been shown to be polymorphically ace-tylated in humans. These include hydralazine, procaina-mide, sulphamethazine, endralazine, a number ofsulphonamides, nitrazepam and dapsone. However, theincidence of failed or less effective clinical response as aconsequence of acetylation polyphorphism is uncommon.This is because most drugs that are metabolised by the NATshave a wide therapeutic window or because acetylation is aminor metabolic pathway. An exception is hydralazine.Early studies showed that the antihypertensive activity ofhydralazine was less in rapid acetylators and that a 40%higher dose was necessary for a similar therapeutic effectcompared with slow acetylators.111 This difference appearedto be due to a change in the bioavailability of the drugwhich decreased from 33% in slow acetylators to less than10% in rapid acetylators.112
A more common consequence of the polymorphic acety-lation of therapeutic agents is an increase in the frequencyand severity of side-effects associated with either the rapidor slow phenotype (Table 3). These adverse effects often

Pharmacogenetics of the arylamine N-acetyltransferasesNJ Butcher et al
38
Table 3 Effect of acetylator status on drug response and
toxicity
Drug Phenotype Effect Reference
Dapsone Slow Neurotoxicity 118,126
Sulphamethoxazole Slow Hypersensitivity 127,128
Hydralazine Slow Systemic lupus 129,130
erythematosusRapid Decreased therapeutic 111,112
effectIsoniazid Slow Interaction with phenytoin 131
Slow Interaction with rifampicin 132
Cotrimoxazole Slow Various adverse reactions 133
Sulphasalazine Slow Various toxicities 114
Slow Hepatotoxicity 134
Slow Nausea/vomiting 135
Amonafide Rapid Leukopenia 120,121
Procainamide Slow Systemic lupus 136–138
erythematosusPhenelzine Rapid Decreased therapeutic 139,140
effectp-Phenylenediamine Slow Contact dermatitis 141
arise as the result of a shift in the metabolic pathwaysresponsible for the activation and detoxification of the drug.This is best illustrated by the amine-containing sulphona-mides, such as sulphamethoxazole, that undergo hydroxyl-ation to a reactive N-hydroxy metabolite capable of coval-ently binding to macromolecules and giving rise toidiosyncratic adverse reactions.113 These drugs can also beacetylated by NAT2 to non-reactive N-acetyl metabolites. Inslow acetylators, a higher proportion of the drug is N-hydroxylated and consequently, these individuals are at agreater risk of sulphonamide-induced toxicity.114–116 How-ever, as pointed out by Spielberg, the incidence of severeadverse side effects to sulphonamides is much less than theincidence of the slow acetylator phenotype suggesting thatother factors predispose individuals to idiosyncraticadverse reactions.113
Risk of developing side effects, such as neurotoxicity orhaemolytic anemia, to dapsone therapy is very similar tothat described for the sulphonamides.117 The most severeincidence of toxicity occurred in individuals with a slow ace-tylator phenotype who are rapid hydroxylators, which isconsistent with the role each pathway has in the activationand detoxification of the drug.118
While slow acetylators are at a greater risk of toxicity fromsulphonamides and dapsone, other therapeutic agents exhi-bit increased incidence of adverse reactions in rapid acetyla-tors. Amonafide is a novel arylamine that has previouslybeen used in clinical trials for the treatment of various can-cers. It undergoes N-acetylation to an active metabolite thatcontributes to systemic toxicity. Several studies have shownthat myelosuppression is greater in rapid acetylators (whiteblood cell nadirs of 500 �l−1) compared to slow acetylators(white blood cell nadirs of 3400 �l−1) following a standarddose of 300 mg m−2 daily for 5 days.119–121. This has led todifferent recommended doses for the two groups.120,122
The Pharmacogenomics Journal
The recent discovery and cloning of prokaryotic NATs hasraised the possibility that bacterial metabolism of drugs andother xenobiotics can contribute to their therapeutic andtoxicological efficacy in vivo.33,123 Payton and associatesshowed that M. smegmatis transformed with the M. tubercu-losis NAT gene has a 3-fold higher resistance to isoniazid dueto an increase in acetylation of the drug.123 These obser-vations suggested that the level of NAT expression in targetbacteria may be an important therapeutic modifier for anti-biotics that are extensively acetylated. In addition, polymor-phisms in the bacterial NAT genes could lead to differenttherapeutic responses.
Okumura et al124 found that the acetylated metabolites ofa range of arylamines such as p-aminobenzoic acid, 4-ami-nobiphenyl and 1-aminopyrene were excreted in the urineand feces of dogs that lack N-acetyltransferase activity.49
They showed that the intestinal microflora were responsiblefor the formation of the acetyl derivatives. Similarly, themicroflora in the intestine of rats contribute to the acety-lation of 2-nitrofluorene and the formation of DNA adductsin liver, kidney, lung and heart following oral adminis-tration.125 Taken together, these studies suggest that bac-terial NAT has a role in the activation and detoxification ofxenobiotics in the host organism and may play animportant role in the metabolism of anti-inflammatorydrugs, such as 5-aminosalicylic acid.33
CONCLUDING REMARKSAlthough considerable allelic variation exists for both NAT1and NAT2, our understanding of the molecular mechanismsand functional significance of many of these alleles, parti-cularly for NAT1, is still limited. The majority of functionalstudies to date have been performed in bacterial expressionsystems, and the results of such studies may not necessarilyaccurately reflect what occurs in vivo, due to differences indegradation/processing pathways between bacterial andmammalian systems. Discrepancies between the two sys-tems have been reported with regard to NAT protein contentand slow acetylator alleles, and characterisation using mam-malian systems may provide a better understanding of themolecular mechanisms leading to different NAT pheno-types.
Much of the research in the area of NATs has involvedidentifying relationships between allele frequencies and dis-ease, particularly different forms of cancers. Although sev-eral studies have reported associations between differentNAT alleles and various cancers, other studies have failed todo so. While these inconsistencies may be due to severalfactors, such as differences in exposure to arylamine carcino-gens, it may well be that genotype does not necessarily accu-rately reflect phenotype. For example, although the regu-lation of NAT1 expression certainly has a geneticcomponent, this only accounts for part of the observedvariability in NAT1 activities. We have shown that signifi-cant variation in NAT1 activity can be observed within asingle phenotype and within the same individuals measuredon different occasions, suggesting that a considerable partof the variation in activity is environmentally based. In sup-

Pharmacogenetics of the arylamine N-acetyltransferasesNJ Butcher et al
39
port of this, we have recently provided evidence for sub-strate-dependent regulation of NAT1,101 and identified aminimum promoter sequence for the human NAT1 thatconsists of an AP-1-like motif flanked on either side by aTCATT sequence (manuscript in preparation). Transienttransfection assays showed that both the AP-1 motif and the3′-TCATT sequence were essential for basal promoteractivity, while the 5′-TCATT sequence appeared to act as anattenuator of phorbol 12-myristate 13-acetate induction.Moreover, antibody supershift assays suggested that c-jun,Oct-1, and YY1 transcription factors form complexes withthe NAT1 minimum promoter. Therefore, environmentalfactors that alter the expression of transcription factors, suchas those mentioned above also may modulate the basalexpression of NAT1 in vivo. There also is the possibility thatother promoter sequences and binding motifs exist furtherupstream from the minimum basal promoter sequence thatcould modulate NAT1 activity under certain conditions.
DUALITY OF INTERESTNone declared.
ABBREVIATIONSNAT1 arylamine N-acetyltransferase 1NAT2 arylamine N-acetyltranferase 2PAS p-aminosalicylic acidPABA p-aminobenzoic acid
REFERENCES1 Hein DW, Doll MA, Rustan TD, Gray K, Feng Y, Ferguson RJ et al. Meta-
bolic activation and deactivation of arylamine carcinogens by recombi-nant human NAT1 and polymorphic NAT2 acetyltransferases. Carcino-genesis 1993; 14: 1633–1638.
2 Kato R. Metabolic activation of mutagenic heterocyclic aromaticamines from protein pyrolysates. Crit Rev Toxicol 1986; 16: 307–348.
3 Minchin RF, Reeves PT, Teitel CH, McManus ME, Mojarrabi B, Ilett KFet al. N- and O-acetylation of aromatic and heterocyclic amine carcino-gens by human monomorphic and polymorphic acetyltransferasesexpressed in COS-1 cells. Biochem Biophys Res Commun 1992; 185:839–844.
4 Zenser TV, Lakshmi VM, Rustan TD, Doll MA, Deitz AC, Davis BB et al.Human N-acetylation of benzidine: role of NAT1 and NAT2. CancerRes 1996; 56: 3941–3947.
5 Grant DM, Blum M, Demierre A, Meyer UA. Nucleotide sequence ofan intronless gene for a human arylamine N-acetyltransferase relatedto polymorphic drug acetylation. Nucl Acids Res 1989; 17: 3978.
6 Blum M, Grant DM, McBride W, Heim M, Meyer UA. Human arylamineN-acetyltransferase genes: isolation, chromosomal localization, andfunctional expression. DNA & Cell Biol 1990; 9: 193–203.
7 Franke S, Klawitz I, Schnakenberg E, Rommel B, Van de Ven W, Buller-diek J et al. Isolation and mapping of a cosmid clone containing thehuman NAT2 gene. Biochem Biophys Res Communi 1994; 199: 52–55.
8 Hickman D, Risch A, Buckle V, Spurr NK, Jeremiah SJ, McCarthy A etal. Chromosomal localization of human genes for arylamine N-acetyl-transferase. Biochem J 1994; 297: 441–445.
9 Bova GS, Carter BS, Bussemakers MJ, Emi M, Fujiwara Y, Kyprianou Net al. Homozygous deletion and frequent allelic loss of chromosome8p22 loci in human prostate cancer. Cancer Res 1993; 53: 3869–3873.
10 Maitra A, Tavassoli FA, Albores-Saavedra J, Behrens C, Wistuba, II, Bry-ant D et al. Molecular abnormalities associated with secretory carci-nomas of the breast. Hum Pathol 1999; 30: 1435–1440.
11 Lutchman M, Pack S, Kim AC, Azim A, Emmert-Buck M, van Huffel Cet al. Loss of heterozygosity on 8p in prostate cancer implicates a role
www.nature.com/tpj
for dematin in tumor progression. Cancer Genet Cytogenet 1999; 115:65–69.
12 Brown MR, Chuaqui R, Vocke CD, Berchuck A, Middleton LP, Emmert-Buck MR et al. Allelic loss on chromosome arm 8p: analysis of sporadicepithelial ovarian tumors. Gynecol Oncol 1999; 74: 98–102.
13 Wistuba, II, Behrens C, Virmani AK, Milchgrub S, Syed S, Lam S et al.Allelic losses at chromosome 8p21–23 are early and frequent eventsin the pathogenesis of lung cancer. Cancer Res 1999; 59: 1973–1979.
14 Ohsako S, Deguchi T. Cloning and expression of cDNAs for polymor-phic and monomorphic arylamine N-acetyltransferases from humanliver. J Biol Chem 1990; 265: 4630–4634.
15 Vatsis KP, Weber WW, Bell DA, Dupret JM, Evans DA, Grant DM etal. Nomenclature for N-acetyltransferases. Pharmacogenetics 1995; 5:1–17.
16 Butcher NJ, Ilett KF, Minchin RF. Functional polymorphism of thehuman arylamine N-acetyltransferase type 1 gene caused by C190Tand G560A mutations. Pharmacogenetics 1998; 8: 67–72.
17 Hughes NC, Janezic SA, McQueen KL, Jewett MA, Castranio T, Bell DAet al. Identification and characterization of variant alleles of humanacetyltransferase NAT1 with defective function using p-aminosalicylateas an in-vivo and in-vitro probe. Pharmacogenetics 1998; 8: 55–66.
18 Lin HJ, Probst-Hensch NM, Hughes NC, Sakamoto GT, Louie AD, KauIH et al. Variants of N-acetyltransferase NAT1 and a case-control studyof colorectal adenomas. Pharmacogenetics 1998; 8: 269–281.
19 Sinclair JC, Sandy J, Delgoda R, Sim E, Noble ME. Structure of arylam-ine N-acetyltransferase reveals a catalytic triad. Nat Struct Biol 2000;7: 560–564.
20 Andres HH, Klem AJ, Schopfer LM, Harrison JK, Weber WW. On theactive site of liver acetyl-CoA. Arylamine N-acetyltransferase from rapidacetylator rabbits (III/J). J Biol Chem 1988; 263: 7521–7527.
21 Dupret JM, Grant DM. Site-directed mutagenesis of recombinanthuman arylamine N-acetyltransferase expressed in Escherichia coli. Evi-dence for direct involvement of Cys68 in the catalytic mechanism ofpolymorphic human NAT2. J Biol Chem 1992; 267: 7381–7385.
22 Floss HG, Yu TW. Lessons from the rifamycin biosynthetic gene cluster.Curr Opin Chem Biol 1999; 3: 592–597.
23 Upton A Johnson N Sandy J, Sim E. Arylamine N-acetyltransferases ofmice, men and microorganisms. Trends Pharmacol Sci 2001; 22:140–146.
24 Hein DW, Grant DM, Sim E. Update on consensus arylamine N-acetyl-transferase gene nomenclature. Pharmacogenetics 2000; 10: 291–292.
25 Hein DW, Doll MA, Rustan TD, Gray K, Ferguson RJ, Feng Y. Construc-tion of Syrian hamster lines congenic at the polymorphic acetyl-transferase locus (NAT2): acetylator genotype-dependent N- and O-acetylation of arylamine carcinogens. Toxicol Appl Pharmacol 1994;124: 16–24.
26 de Leon JH, Martell KJ, Vatsis KP, Weber WW. Slow acetylation in miceis caused by a labile and catalytically impaired mutant N-acetyltransfer-ase (NAT2 9). Drug Metab Dispos 1995; 23: 1354–1361.
27 Ware JA, Svensson CK. Longitudinal distribution of arylamine N-acetyl-transferases in the intestine of the hamster, mouse, and rat. Evidencefor multiplicity of N-acetyltransferases in the intestine. Biochem Pharm-acol 1996; 52: 1613–1620.
28 Drobitch RK, Divakaruni P, Svensson CK. Effect of streptolysin-O-on rathepatic acetyl coenzyme-A: arylamine N-acetyltransferase and cyto-chrome P-450 2B 1/2 activities ex vivo. Immunopharmacol Immunotox-icol 1998; 20: 159–171.
29 Yamamura E, Sayama M, Kakikawa M, Mori M, Taketo A, Kodaira K.Purification and biochemical properties of an N-hydroxyarylamine O-acetyltransferase from Escherichia coli. Biochim Biophys Acta 2000;1475: 10–16.
30 Kattner E, Dubbels R, Schloot W. Acetylation of procainamide andisoniazid by a rat liver-N-acetyl-transferase. Eur J Drug Metab Pharmaco-kinet 1981; 6: 81–84.
31 Riddle B, Jencks WP. Acetyl-coenzyme A: arylamine N-acetyltransfer-ase. Role of the acetyl-enzyme intermediate and the effects of substitu-ents on the rate. J Biol Chem 1971; 246: 3250–3258.
32 Payton M, Mushtaq A, Yu TW, Wu LJ, Sinclair J, Sim E. Eubacterialarylamine N-acetyltransferases-identification and comparison of 18members of the protein family with conserved active site cysteine, his-tidine and aspartate residues. Microbiology 2001; 147: 1137–1147.
33 Delomenie C, Fouix S, Longuemaux S, Brahimi N, Bizet C, Picard B et

Pharmacogenetics of the arylamine N-acetyltransferasesNJ Butcher et al
40
al. Identification and functional characterisation of arylamine N-acetyl-transferase in eubacteria: evidence for highly selective acetylation of5-aminosalicylic acid. J Bacteriol 2001; 183: 3417–3427.
34 Chung JG, Wang HH, Tsou MF, Hsieh SE, Lo HH, Yen YS et al. Evidencefor arylamine N-acetyltransferase activity in the bacterium Helicobacterpylori. Toxicol Lett 1997; 91: 63–71.
35 Ebisawa T, Deguchi T. Structure and restriction fragment length poly-morphism of genes for human liver arylamine N-acetyltransferases.Biochem Biophys Res Commun 1991; 177: 1252–1257.
36 Matas N, Thygesen P, Stacey M, Risch A, Sim E. Mapping AAC1, AAC2and AACP, the genes for arylamine N-acetyltransferases, carcinogenmetabolising enzymes on human chromosome 8p22, a region fre-quently deleted in tumours. Cytogenet Cell Genet 1997; 77: 290–295.
37 Martell KJ, Vatsis KP, Weber WW. Molecular genetic basis of rapid andslow acetylation in mice. Mol Pharmacol 1991; 40: 218–227.
38 Kelly SL, Sim E. Arylamine N-acetyltransferase in Balb/c mice: identifi-cation of a novel mouse isoenzyme by cloning and expression in vitro.Biochem J 1994; 302: 347–353.
39 Martell KJ, Levy GN, Weber WW. Cloned mouse N-acetyltransferases:enzymatic properties of expressed Nat-1 and Nat-2 gene products.Mol Pharmacol 1992; 42: 265–272.
40 Estrada-Rodgers L, Levy GN, Weber WW. Substrate selectivity of mouseN-acetyltransferases 1, 2, and 3 expressed in COS-1 cells. Drug MetabDispos 1998; 26: 502–505.
41 Fakis G, Boukouvala S, Buckle V, Payton M, Denning C, Sim E. Chro-mosome mapping of the genes for murine arylamine N-acetyltransfer-ases (NATs), enzymes involved in the metabolism of carcinogens:identification of a novel upstream noncoding exon for murine Nat2.Cytogenet Cell Genet 2000; 90: 134–138.
42 Mattano SS, Erickson RP, Nesbitt MN, Weber WW. Linkage of Nat andEs-1 in the mouse and development of strains congenic for N-acetyl-transferase. J Hered 1988; 79: 430–433.
43 Fretland AJ, Doll MA, Gray K, Feng Y, Hein DW. Cloning, sequencing,and recombinant expression of NAT1, NAT2, and NAT3 derived fromthe C3H/HeJ (rapid) and A/HeJ (slow) acetylator inbred mouse: func-tional characterization of the activation and deactivation of aromaticamine carcinogens. Toxicol Appl Pharmacol 1997; 142: 360–366.
44 Blum M, Grant DM, Demierre A, Meyer UA. N-acetylation pharmaco-genetics: a gene deletion causes absence of arylamine N-acetyltransfer-ase in liver of slow acetylator rabbits. Proc Natl Acad Sci USA 1989; 86:9554–9557.
45 Ferguson RJ, Doll MA, Rustan TD, Hein DW. Cloning, expression, andfunctional characterization of rapid and slow acetylator polymorphicN-acetyl-transferase encoding genes of the Syrian hamster. Pharmaco-genetics 1996; 6: 55–66.
46 Hein DW, Rustan TD, Bucher KD, Martin WJ, Furman EJ. Acetylatorphenotype-dependent and -independent expression of arylamine N-acetyltransferase isozymes in rapid and slow acetylator inbred rat liver.Drug Metab Dispos 1991; 19: 933–937.
47 Blum M, Grant DM, Demierre A, Meyer UA. Nucleotide sequence ofa full-length cDNA for arylamine N-acetyltransferase from rabbit liver.Nucl Acids Res 1989; 17: 3589.
48 Trepanier LA, Cribb AE, Spielberg SP, Ray K. Deficiency of cytosolicarylamine N-acetylation in the domestic cat and wild felids caused bythe presence of a single NAT1-like gene. Pharmacogenetics 1998; 8:169–179.
49 Trepanier LA, Ray K, Winand NJ, Spielberg SP, Cribb AE. Cytosolic aryl-amine N-acetyltransferase (NAT) deficiency in the dog and other can-ids due to an absence of NAT genes. Biochem Pharmacol 1997; 54:73–80.
50 Blum M, Demierre A, Grant DM, Heim M, Meyer UA. Molecular mech-anism of slow acetylation of drugs and carcinogens in humans. ProcNatl Acad Sci USA 1991; 88: 5237–5241.
51 Grant DM, Blum M, Beer M, Meyer UA. Monomorphic and polymor-phic human arylamine N-acetyltransferases: a comparison of liver iso-zymes and expressed products of two cloned genes. Mol Pharmacol1991; 39: 184–191.
52 Grant DM, Hughes NC, Janezic SA, Goodfellow GH, Chen HJ, GaedigkA et al. Human acetyltransferase polymorphisms. Mutat Res 1997; 376:61–70.
53 Bell DA, Taylor JA, Butler MA, Stephens EA, Wiest J, Brubaker LH et al.Genotype/phenotype discordance for human arylamine N-acetyl-
The Pharmacogenomics Journal
transferase (NAT2) reveals a new slow-acetylator allele common inAfrican-Americans. Carcinogenesis 1993; 14: 1689–1692.
54 Cascorbi I, Drakoulis N, Brockmoller J, Maurer A, Sperling K, Roots I.Arylamine N-acetyltransferase (NAT2) mutations and their allelic link-age in unrelated Caucasian individuals: correlation with phenotypicactivity. Am J Hum Genet 1995; 57: 581–592.
55 Hickman D, Risch A, Camilleri JP, Sim E. Genotyping human polymor-phic arylamine N-acetyltransferase: identification of new slow allotypicvariants. Pharmacogenetics 1992; 2: 217–226.
56 Hein DW, Doll MA, Fretland AJ, Leff MA, Webb SJ, Xiao GH et al. Mol-ecular genetics and epidemiology of the NAT1 and NAT2 acetylationpolymorphisms. Cancer Epidemiol, Biomarkers & Prevention 2000; 9:29–42.
57 Grant DM, Morike K, Eichelbaum M, Meyer UA. Acetylation pharmaco-genetics. The slow acetylator phenotype is caused by decreased orabsent arylamine N-acetyltransferase in human liver. J Clin Invest 1990;85: 968–972.
58 Dupret JM, Goodfellow GH, Janezic SA, Grant DM. Structure-functionstudies of human arylamine N-acetyltransferases NAT1 and NAT2.Functional analysis of recombinant NAT1/NAT2 chimeras expressed inEscherichia coli. J Biol Chem 1994; 269: 26830–26835.
59 Hein DW, Rustan TD, Ferguson RJ, Doll MA, Gray K. Metabolic acti-vation of aromatic and heterocyclic N-hydroxyarylamines by wild-typeand mutant recombinant human NAT1 and NAT2 acetyltransferases.Arch Toxicol 1994; 68: 129–133.
60 Hickman D, Palamanda JR, Unadkat JD, Sim E. Enzyme kinetic proper-ties of human recombinant arylamine N-acetyltransferase 2 allotypicvariants expressed in Escherichia coli. Biochem Pharmacol 1995; 50:697–703.
61 Delomenie C, Goodfellow GH, Krishnamoorthy R, Grant DM, DupretJM. Study of the role of the highly conserved residues Arg9 andArg64 in the catalytic function of human N-acetyltransferases NAT1and NAT2 by site-directed mutagenesis. Biochem J 1997; 323:207–215.
62 Leff MA, Fretland AJ, Doll MA, Hein DW. Novel human N-acetyl-transferase 2 alleles that differ in mechanism for slow acetylator pheno-type. J Biol Chem 1999; 274: 34519–34522.
63 Hein DW, Doll MA, Rustan TD, Ferguson RJ. Metabolic activation ofN-hydroxyarylamines and N-hydroxyarylamides by 16 recombinanthuman NAT2 allozymes: effects of 7 specific NAT2 nucleic acid substi-tutions. Cancer Res 1995; 55: 3531–3536.
64 Hein DW, Ferguson RJ, Doll MA, Rustan TD, Gray K. Molecular geneticsof human polymorphic N-acetyltransferase: enzymatic analysis of 15recombinant wild-type, mutant, and chimeric NAT2 allozymes. HumMol Genet 1994; 3: 729–734.
65 Evans DA. N-acetyltransferase. Pharmacol Therapeut 1989; 42: 157–234.
66 Meyer UA, Zanger UM. Molecular mechanisms of genetic polymor-phisms of drug metabolism. Annu Rev Pharmacol Toxicol 1997; 37:269–296.
67 Glowinski IB, Radtke HE, Weber WW. Genetic variation in N-acetylationof carcinogenic arylamines by human and rabbit liver. Mol Pharmacol1978; 14: 940–949.
68 McQueen CA, Weber WW. Characterization of human lymphocyte N-acetyltransferase and its relationship to the isoniazid acetylator poly-morphism. Biochem Genet 1980; 18: 889–904.
69 Cribb AE, Grant DM, Miller MA, Spielberg SP. Expression of monomor-phic arylamine N-acetyltransferase (NAT1) in human leukocytes. JPharmacol Exp Therapeut 1991; 259: 1241–1246.
70 Kilbane AJ, Petroff T, Weber WW. Kinetics of acetyl CoA: arylamine N-acetyltransferase from rapid and slow acetylator human liver. DrugMetab Dispos 1991; 19: 503–507.
71 Ward A, Hickman D, Gordon JW, Sim E. Arylamine N-acetyltransferasein human red blood cells. Biochem Pharmacol 1992; 44: 1099–1104.
72 Weber WW, Vatsis KP. Individual variability in p-aminobenzoic acid N-acetylation by human N-acetyltransferase (NAT1) of peripheral blood.Pharmacogenetics 1993; 3: 209–212.
73 Vatsis KP, Weber WW. Structural heterogeneity of Caucasian N-acetyl-transferase at the NAT1 gene locus. Arch Biochem Biophys 1993; 301:71–76.
74 Bell DA, Badawi AF, Lang NP, Ilett KF, Kadlubar FF, Hirvonen A. Poly-morphism in the N-acetyltransferase 1 (NAT1) polyadenylation signal:

Pharmacogenetics of the arylamine N-acetyltransferasesNJ Butcher et al
41
association of NAT1*10 allele with higher N-acetylation activity inbladder and colon tissue. Cancer Res 1995; 55: 5226–5229.
75 Badawi AF, Hirvonen A, Bell DA, Lang NP, Kadlubar FF. Role of aro-matic amine acetyltransferases, NAT1 and NAT2, in carcinogen-DNAadduct formation in the human urinary bladder. Cancer Res 1995; 55:5230–5237.
76 Kadlubar FF, Badawi AF. Genetic susceptibility and carcinogen-DNAadduct formation in human urinary bladder carcinogenesis. Toxicol Lett1995; 82–83: 627–632.
77 Payton MA, Sim E. Genotyping human arylamine N-acetyltransferasetype 1 (NAT1): the identification of two novel allelic variants. BiochemPharmacol 1998; 55: 361–366.
78 Bell DA, Stephens EA, Castranio T, Umbach DM, Watson M, DeakinM et al. Polyadenylation polymorphism in the acetyltransferase 1 gene(NAT1) increases risk of colorectal cancer. Cancer Res 1995; 55:3537–3542.
79 Bruhn C, Brockmoller J, Cascorbi I, Roots I, Borchert HH. Correlationbetween genotype and phenotype of the human arylamine N-acetyl-transferase type 1 (NAT1). Biochem Pharmacol 1999; 58: 1759–1764.
80 de Leon JH, Vatsis KP, Weber WW. Characterization of naturally occur-ring and recombinant human N-acetyltransferase variants encoded byNAT1. Mol Pharmacol 2000; 58: 288–299.
81 Bouchardy C, Mitrunen K, Wikman H, Husgafvel-Pursiainen K, DayerP, Benhamou S et al. N-acetyltransferase NAT1 and NAT2 genotypesand lung cancer risk. Pharmacogenetics 1998; 8: 291–298.
82 Lo-Guidice JM, Allorge D, Chevalier D, Debuysere H, Fazio F, Lafitte LJet al. Molecular analysis of the N-acetyltransferase 1 gene (NAT1*)using polymerase chain reaction-restriction fragment-single strandconformation polymorphism assay. Pharmacogenetics 2000; 10: 293–300.
83 Dhaini HR, Levy GN. Arylamine N-acetyltransferase 1 (NAT1) geno-types in a Lebanese population. Pharmacogenetics 2000; 10: 79–83.
84 Clark DW. Genetically determined variability in acetylation and oxi-dation. Therapeutic implications. Drugs 1985; 29: 342–375.
85 Evans DA. Survey of the human acetylator polymorphism in spon-taneous disorders. J Med Genet 1984; 21: 243–253.
86 Evans DA. Genetic Factors in Drug Therapy. Cambridge University Press:Cambridge, 1993.
87 Weber WW. Pharmacogenetics. Oxford University Press: New York,1997.
88 Besarati Nia A, Van Straaten HW, Kleinjans JC, Van Schooten FJ. Immu-noperoxidase detection of 4-aminobiphenyl- and polycyclic aromatichydrocarbons-DNA adducts in induced sputum of smokers and non-smokers. Mutat Res 2000; 468: 125–135.
89 Nagao M, Wakabayashi K, Ushijima T, Toyota M, Totsuka Y, SugimuraT. Human exposure to carcinogenic heterocyclic amines and theirmutational fingerprints in experimental animals. Environ Health Per-spect 1996; 104: 497–501.
90 Smith CJ, Livingston SD, Doolittle DJ. An international literature surveyof ‘IARC Group I carcinogens’ reported in mainstream cigarette smoke.Food & Chem Toxicol 1997; 35: 1107–1130.
91 Sram RJ, Binkova B. Molecular epidemiology studies on occupationaland environmental exposure to mutagens and carcinogens, 1997–1999. Environ Health Perspect 2000; 108: 57–70.
92 Mao Y, Hu J, Ugnat AM, White K. Non-Hodgkin’s lymphoma and occu-pational exposure to chemicals in Canada. Canadian Cancer RegistriesEpidemiology Research Group. Ann Oncol 2000; 11: 69–73.
93 Zhou Q, Talaska G, Jaeger M, Bhatnagar VK, Hayes RB, Zenzer TV etal. Benzidine-DNA adduct levels in human peripheral white blood cellssignificantly correlate with levels in exfoliated urothelial cells. MutatRes 1997; 393: 199–205.
94 Lower GM, Jr, Nilsson T, Nelson CE, Wolf H, Gamsky TE, Bryan GT.N-acetyltransferase phenotype and risk in urinary bladder cancer:approaches in molecular epidemiology. Preliminary results in Swedenand Denmark. Environ Health Perspect 1979; 29: 71–79.
95 Wohlleb JC, Hunter CF, Blass B, Kadlubar FF, Chu DZ, Lang NP. Aro-matic amine acetyltransferase as a marker for colorectal cancer:environmental and demographic associations. Int J Cancer 1990; 46:22–30.
96 Ilett KF, David BM, Detchon P, Castleden WM, Kwa R. Acetylationphenotype in colorectal carcinoma. Cancer Res 1987; 47: 1466–1469.
97 Lang NP, Chu DZ, Hunter CF, Kendall DC, Flammang TJ, Kadlubar FF.
www.nature.com/tpj
Role of aromatic amine acetyltransferase in human colorectal cancer.Arch Surg 1986; 121: 1259–1261.
98 Brockton N, Little J, Sharp L, Cotton SC. N-acetyltransferase polymor-phisms and colorectal cancer: a HuGE review. Am J Epidemiol 2000;151: 846–861.
99 Chen J, Stampfer MJ, Hough HL, Garcia-Closas M, Willett WC, Henne-kens CH et al. A prospective study of N-acetyltransferase genotype,red meat intake, and risk of colorectal cancer. Cancer Res 1998; 58:3307–3311.
100 Welfare MR, Cooper J, Bassendine MF, Daly AK. Relationship betweenacetylator status, smoking, and diet and colorectal cancer risk in thenorth-east of England. Carcinogenesis 1997; 18: 1351–1354.
101 Butcher NJ, Ilett KF, Minchin RF. Substrate-dependent regulation ofhuman arylamine N-acetyltransferase-1 in cultured cells. Mol Pharma-col 2000; 57: 468–473.
102 Lang NP, Butler MA, Massengill J, Lawson M, Stotts RC, Hauer-JensenM et al. Rapid metabolic phenotypes for acetyltransferase and cyto-chrome P4501A2 and putative exposure to food-borne heterocyclicamines increase the risk for colorectal cancer or polyps. Cancer Epide-miol, Biomarkers & Prevention 1994; 3: 675–682.
103 Kaderlik KR, Minchin RF, Mulder GJ, Ilett KF, Daugaard-Jenson M,Teitel CH et al. Metabolic activation pathway for the formation ofDNA adducts of the carcinogen 2-amino-1-methyl-6-phenylimid-azo[4,5-b]pyridine (PhIP) in rat extrahepatic tissues. Carcinogenesis1994; 15: 1703–1709.
104 Kadlubar F, Kaderlik RK, Mulder GJ, Lin D, Butler MA, Teitel CH et al.Metabolic activation and DNA adduct detection of PhIP in dogs, rats,and humans in relation to urinary bladder and colon carcinogenesis.Princess Takamatsu Symp 1995; 23: 207–213.
105 Kristiansen E, Meyer O, Thorup I. The ability of two cooked foodmutagens to induce aberrant crypt foci in mice. Eur J Cancer Prev1997; 6: 53–57.
106 Purewal M, Fretland AJ, Schut HA, Hein DW, Wargovich MJ. Associ-ation between acetylator genotype and 2-amino-1-methyl-6-phenyl-imidazo[4,5-b]pyridine (PhIP) DNA adduct formation in colon andprostate of inbred Fischer 344 and Wistar Kyoto rats. Cancer Lett2000; 149: 53–60.
107 Bandmann O, Vaughan JR, Holmans P, Marsden CD, Wood NW.Detailed genotyping demonstrates association between the slow ace-tylator genotype for N-acetyltransferase 2 (NAT2) and familial Parkin-son’s disease. Mov Disord 2000; 15: 30–35.
108 Tan EK, Khajavi M, Thornby JI, Nagamitsu S, Jankovic J, Ashizawa T.Variability and validity of polymorphism association studies in Parkin-son’s disease. Neurology 2000; 55: 533–538.
109 Ogawa M. [Biochemical, molecular genetic and ecogenetic studiesof polymorphic arylamine N-acetyltransferase (NAT2) in the brain].Fukuoka Igaku Zasshi – Fukuoka Acta Medica 1999; 90: 118–131.
110 Bonicke R, Reif W. Enzymatische inaktivierung von isonicotinsaurehy-drazid im menschlichen und tierischen organismus. Arch Exp PatholPharmacol 1953; 220: 321–333.
111 Jounela AJ, Pasanen M, Mattila MJ. Acetylator phenotype and theantihypertensive response to hydralazine. Acta Medica Scand 1975;197: 303–306.
112 Shepherd AM, Ludden TM, McNay JL, Lin MS. Hydralazine kineticsafter single and repeated oral doses. Clin Pharmacol Therapeut 1980;28: 804–811.
113 Spielberg SP. N-acetyltransferases: pharmacogenetics and clinicalconsequences of polymorphic drug metabolism. J PharmacokinetBiopharmaceut 1996; 24: 509–519.
114 Das KM, Dubin R. Clinical pharmacokinetics of sulphasalazine. ClinPharmacokinet 1976; 1: 406–425.
115 Shear NH, Spielberg SP, Grant DM, Tang BK, Kalow W. Differencesin metabolism of sulfonamides predisposing to idiosyncratic toxicity.Ann Intern Med 1986; 105: 179–184.
116 Cribb AE, Nuss CE, Alberts DW, Lamphere DB, Grant DM, GrossmanSJ et al. Covalent binding of sulfamethoxazole reactive metabolitesto human and rat liver subcellular fractions assessed by immuno-chemical detection. Chem Res Toxicol 1996; 9: 500–507.
117 May DG, Porter JA, Uetrecht JP, Wilkinson GR, Branch RA. The contri-bution of N-hydroxylation and acetylation to dapsone pharmacoki-netics in normal subjects. Clin Pharmacol Therapeut 1990; 48: 619–627.

Pharmacogenetics of the arylamine N-acetyltransferasesNJ Butcher et al
42
118 Bluhm RE, Adedoyin A, McCarver DG, Branch RA. Development ofdapsone toxicity in patients with inflammatory dermatoses: activityof acetylation and hydroxylation of dapsone as risk factors. Clin Phar-macol Therapeut 1999; 65: 598–605.
119 Ratain MJ, Mick R, Berezin F, Janisch L, Schilsky RL, Williams SF et al.Paradoxical relationship between acetylator phenotype and amon-afide toxicity. Clin Pharmacol Therapeut 1991; 50: 573–579.
120 Ratain MJ, Mick R, Berezin F, Janisch L, Schilsky RL, Vogelzang NJ etal. Phase I study of amonafide dosing based on acetylator phenotype.Cancer Res 1993; 53: 2304–2308.
121 Ratain MJ, Rosner G, Allen SL, Costanza M, Van Echo DA, HendersonIC et al. Population pharmacodynamic study of amonafide: a Cancerand Leukemia Group B study. J Clin Oncol 1995; 13: 741–747.
122 Ratain MJ, Mick R, Janisch L, Berezin F, Schilsky RL, Vogelzang NJ etal. Individualized dosing of amonafide based on a pharmacodynamicmodel incorporating acetylator phenotype and gender. Pharmaco-genetics 1996; 6: 93–101.
123 Payton M, Auty R, Delgoda R, Everett M, Sim E. Cloning and charac-terization of arylamine N-acetyltransferase genes from Mycobacteriumsmegmatis and Mycobacterium tuberculosis: increased expressionresults in isoniazid resistance. J Bacteriol 1999; 181: 1343–1347.
124 Okumura F, Ueda O, Kitamura S, Tatsumi K. N-acetylation and N-formylation of carcinogenic arylamines and related compounds indogs. Carcinogenesis 1995; 16: 71–76.
125 Moller L, Zeisig M, Midtvedt T, Gustafsson JA. Intestinal microfloraenhances formation of DNA adducts following administration of 2-NFand 2-AAF [published erratum appears in Carcinogenesis 1994 Dec;15(12): 2969]. Carcinogenesis 1994; 15: 857–861.
126 Guo R, Thormann W, Lauterberg B. Relationship between high inci-dence of adverse dapsone reactions and slow acetylate phenotypeor low plasma/lymphocyte glutathione level. Chin Med J 1996; 109:933–936.
127 Zielinska E, Niewiarowski W, Bodalski J, Rebowski G, Skretkowicz J,Mianowska K et al. Genotyping of the arylamine N-acetyltransferasepolymorphism in the prediction of idiosyncratic reactions to tri-methoprim-sulfamethoxazole in infants. Pharmacy World & Science1998; 20: 123–130.
128 Cribb AE, Nakamura H, Grant DM, Miller MA, Spielberg SP. Role ofpolymorphic and monomorphic human arylamine N-acetyltransfer-ases in determining sulfamethoxazole metabolism. Biochem Pharma-col 1993; 45: 1277–1282.
The Pharmacogenomics Journal
129 Lemke LE, McQueen CA. Acetylation and its role in the mutagenicityof the antihypertensive agent hydralazine. Drug Metab Disposition1995; 23: 559–565.
130 Strandberg I, Boman G, Hassler L, Sjoqvist F. Acetylator phenotypein patients with hydralazine-induced lupoid syndrome. Acta MedicaScand 1976; 200: 367–371.
131 Walubo A, Aboo A. Phenytoin toxicity due to concomitant antitu-berculosis therapy. S Afr Med J 1995; 85: 1175–1176.
132 Ohno M, Yamaguchi I, Yamamoto I, Fukuda T, Yokota S, Maekura Ret al. Slow N-acetyltransferase 2 genotype affects the incidence ofisoniazid and rifampicin-induced hepatotoxicity. Int J TuberculosisLung Dis 2000; 4: 256–261.
133 Zielinska E, Niewiarowski W, Bodalski J. The arylamine N-acetyl-transferase (NAT2) polymorphism and the risk of adverse reactions toco-trimoxazole in children. Eur J Clin Pharmacol 1998; 54: 779–785.
134 Kitas GD, Farr M, Waterhouse L, Bacon PA. Influence of acetylatorstatus on sulphasalazine efficacy and toxicity in patients with rheuma-toid arthritis. Scand J Rheumatol 1992; 21: 220–225.
135 Pullar T, Hunter JA, Capell HA. Effect of acetylator phenotype on effi-cacy and toxicity of sulphasalazine in rheumatoid arthritis. Ann RheumDis 1985; 44: 831–837.
136 Mongey AB, Sim E, Risch A, Hess E. Acetylation status is associatedwith serological changes but not clinically significant disease inpatients receiving procainamide. J Rheumatol 1999; 26: 1721–1726.
137 Tan EM, Rubin RL. Autoallergic reactions induced by procainamide.J Allergy Clin Immunol 1984; 74: 631–634.
138 Reidenberg MM, Drayer DE. Procainamide, N-acetylprocainamide,antinuclear antibody and systemic lupus erythematosus. Angiology1986; 37: 968–971.
139 Paykel ES, West PS, Rowan PR, Parker RR. Influence of acetylatorphenotype on antidepressant effects of phenelzine. Br J Psychiatry1982; 141: 243–248.
140 Johnstone EC. The relationship between acetylator status and inhi-bition of monoamine oxidase, excretion of free drug and antidepress-ant response in depressed patients on phenelzine. Psychopharmacolo-gia 1976; 46: 289–294.
141 Kawakubo Y, Nakamori M, Schopf E, Ohkido M. Acetylator pheno-type in patients with p-phenylenediamine allergy. Dermatology 1997;195: 43–45.





























