Embed Size (px)
Citation preview

GENERAL AND COMPARATIVE ENDOCRINOLOGY 71, 29-35 (1988)
Nature of Moulting Control in Amphibians: Effects of Cortisol Implants in Toads Bufo bufo
C.BARKERJP)RGENSEN
Zoophysiological Laboratory A, August Krogh Institute, Universitetsparken 13, DK-2100 Copenhagen @, Denmark
Accepted February 2, 1988
Implantation of cortisol pellets in a dorsal lymph sac reduced the frequency with which toads replaced the strata cornea of the epidermis. Except in a few cases, however, the replaced strata cornea were not shed as in a normal moult. The mean interval between replacements of strata cornea rose from about 9-10 days in control toads to about 18 days in intact toads bearing implants. Removal of the cortisol pellets approximately 1 month after their insertion was followed by moulting on the fourth (or fifth) day. After the first moult, the frequency of replacement of strata cornea was initially high, but returned gradually to the normal level, probably concurrently with increasing levels of endogenous adrenocortical hormones. In toads with extirpated or ectopically transplanted pars distalis of the hypo- physis, implanted cortisol pellets increased the interval between replacements of strata cornea from some few days to about 14-15 days, not significantly different from the intact toads, but no sheddings occurred. It is concluded that the moulting rhythm in toads, and other amphibians, is basically a rhythm in epidermal differentiation, expressed in the fre- quency with which the strata cornea are being replaced. The duration of the intervals between replacements varies with the level of circulating hormones, and replacement pro- ceeds in the absence of a corresponding rhythm in secretion of the hormones. Shedding of a replaced stratum comeum depends upon the integrated function of a complex of pro- cesses, susceptible to disruption. Such disruption presumably prevailed at the pharmaco- logical levels of circulating hormone in the cortisol-implanted toads. Q 1988 Academic PKSS, IK.
Amphibians moult more or less regularly. A moult encompasses a complex of pro- cesses of different nature. It is initiated by an epidermal differentiation that results in the replacement of the old stratum corneum by a new stratum corneum, mostly one cell layer thick. The replaced stratum corneum is subsequently shed through a complex process, including both physiological and behavioral components. The physiological component consists of a copious secretion of mucus beneath the replaced stratum cor- neum, now a slough. The mucus serves as a lubricant during the shedding of the slough, which is carried out by a complicated be- havioral act. After shedding, which typical- ly ends with the slough being eaten, follows
an intermoult period of a duration that var- ies between species and with the tempera- ture (see Larsen, 1976).
Regular, rhythmical moulting in amphib- ians depends upon endocrine activity, but the nature of the hormonal involvement in moulting is not finally elucidated. In toads, Bufo spp., extirpation or ectopic transplan- tation of the pars distalis of the hypophysis reduces the intervals between replacement of strata cornea, e.g., from about 9 days to a few days, but the replaced strata cornea are not shed and pile up to produce an ab- normal keratinized layer covering the epi- dermis (Houssay, 1949; Jorgensen and Larsen, 1961; Budtz, 1979). Shedding can be produced by treating the operated toads
29 0016~6480/88 $1.50 Copyright 0 1988 by Academic Press, Inc. All rights of reproduction in any form reserved

30 C. BARKER JORGENSEN
with small doses of ACTH or an adrenocor- MATERIALS AND METHODS tical hormone, corticosterone being partic- ularly effective (Jorgensen and Larsen, 1964; Stefano and Donoso, 1964). Howev- er, moulting could not be normalized even by daily hormonal treatment of hypophy- sectomized toads; the frequency of forma- tion of new strata cornea remained abnor- mally high (Jorgensen and Larsen, 1964; Budtz, 1977).
The lacking normalization of moulting by daily injections of ACTH or an adrenocor- tical hormone might be due to the short half-life of the circulating hormones (Btichmann et al., 1972). It was therefore investigated whether a constant level of an adrenocortical hormone might normalize moulting. Such a constant level was ob- tained by implanting pellets of cortisol un- der the skin of toads in which the pars dis- talis had been extirpated or transplanted to an eye muscle. The ectopically trans- planted pars distalis in B. b&o continues to secrete prolactin, but little or no ACTH, TSH, or GH (Jorgensen, 1970, 1974; Mc- Keown, 1972).
The experiment particularly aimed at re- vealing the nature of the action of the adre- nocortical hormones, whether rhythmic moulting depends upon corresponding cy- cles in the rate of secretion of ACTH and thus of adrenocortical hormones. It was shown previously that injection of single doses of ACTH or an adrenocortical hor- mone in intact toads did not elicit a moult at any time during the moulting cycle. The ac- tual moult thus did not seem to depend upon an increased rate of secretion of ACTH. It was concluded that the adreno- cortical hormones probably were permis- sive and the rhythm inherent in the skin, but the possibility remained that a moult may be induced by a temporary fall, or dis- continuation, and subsequent rise in ACTH secretion (Jorgensen and Larsen, 1964; Larsen, 1976). To investigate this possibil- ity, cortisol pellets were also implanted subcutaneously in a group of intact toads.
Experiments were carried out during the months of April through June on toads collected in the autumn and kept during the winter in simulated hibernation in a refrigerator at about 2”. Of 54 toads used 3 were females and 51 were males, which weighed about 30 g.
The toads were kept at 20” in natural daylight. They were placed in 36 x 55 x 22-cm plastic boxes, 5-10 toads in each box. The bottoms of the boxes were covered with filter paper, and the toads had access to water in a dish and to shelter under halved flower pots. They were offered mealworms for a 24-hr period once a week.
Extirpation and ectopic transplantation of pars dis- talis of the hypophysis were performed as described by van Dongen et al. (1966). Constant administration of adrenocortical hormone was obtained by inserting about 50 mg cortisol (hydrocortisone) pellets in a dor- sal lymph sac. The pellets were formed from anhy- drous powder in a steel press at a pressure of 6000 kg cm-‘. The preweighed pellets were weighed again at their removal from the lymph sac in order to determine the daily mean rate of absorption of the hormone.
The duration of intermoult periods was determined by noticing days of disappearance of lipstick smeared on the head, as described by Bendsen (1956). The mean duration of intervals between successive re- placements of strata cornea was determined by count- ing the number of strata cornea on biopsies of the epi- dermis cut at 1 pm, according to the technique de- scribed by Budtz (1979).
RESULTS
A4oulting in Controls
The duration of intermoult periods was recorded in a group of 10 male toads over a period of 2 months. The frequency distribu- tion of the moults show a pronounced peak at 9-10 days (Fig. 1). The occasional longer intervals presumably reflect instances where a replaced stratum corneum was not
uIlL&-& L 12 M 28 Mouiting mtervals, days
FIG. 1. Bufo bufo. Frequency distribution of moult- ing intervals in control toads.

MOULTING IN TOADS 31
immediately shed. The registered moults may in these cases consist of two or more strata cornea (Jorgensen and Larsen, 1961). In the present material the lbday inter- moult period may represent two and the 22- and 27-day periods three replaced strata cornea. The 50 recorded intermoult periods may therefore include six skipped shed- dings, or 12% of all replaced strata cornea. Intermoult periods 214 days were there- fore excluded from the calculation of the duration of the periods passing between successive replacements of strata cornea in the untreated, control toads. This duration was 8.7 -+ 1.6 (SD) days (Table 1).
Effects of Implanted Cortisol Pellets
Pars distalis extirpated or transplanted. In a pilot experiment, including three fe- male and two male toads that had cortisol pellets subcutaneously implanted concur- rently with the extirpation of the pars dis- talis, the epidermis maintained its normal appearance even after 2 weeks, but no moults were observed.
At the end of the 2 weeks the pellets were removed, and weighing showed that 721 ? 48 (SD) pg cortisol had been absorbed per day. Within 3 days of removal of the pellets the toads exhibited the typical acute sign of hypophysectomy in the form of loosened strata cornea.
On the fourth day, the cortisol pellets were reinserted, and within 6-7 hr all 5 toads had moulted. The toads were kept un- der observation for 2 months further, dur- ing which period they remained in good health and did not moult. During this sec- ond period the daily absorption of cortisol from the pellets amounted to 297 t 78 kg.
These doses of cortisol are high com- pared with the threshold dose of about 5 ug needed to produce a moult in a hypophy- sectomized toad (Jorgensen and Larsen, 1964). High, slowly declining levels of an adrenocortical hormone may thus prevent the replacement of strata cornea in rapid succession that results from extirpation of the pars distalis, but the normalization of epidermal function did not extend to the restoration of the shedding processes and thus of complete moulting.
A subsequent experiment included 9 male toads with the pars distalis extirpated and 20 males toads with the pars distalis transplanted to an eye muscle. The experi- ment confirmed the previous result, with no clear change in appearance of the epidermis for up to 2 to 3 weeks. However, three moults were recorded, two in the group of toads with extirpated pars distalis and one in the group with transplanted pars distalis, all moults occurring 9-10 days after opera- tion and implantation of the cortisol pellets.
TABLE I EFFECTOF SUBCUTANEOUS IMPLANTATION OF CORTISOL PELLETS ON REPLACEMENTOF STRATUM
CORNEUM IN TOADS. Bufo b&o
Groups
Normal controls” Cortisol treated:”
Intact toads Pars distalis extirpated Pars distalis transplanted
.________-
N
9
10 9
20
n
46
22 22 49
Interval between replacements (days + SD)
8.7 2 1.6
17.7 + 8.5 14.5 k 4.2 14.3 + 4.8
Cortisol absorption (pg day-‘)
490 t 60 405 i_ 110 390 + 110
Note. N, number of toads in the group; n, number of recorded replacements of strata cornea. u Calculated from the known intervals between successive replacements. b Calculated from the individual mean intervals between replacements, estimated from the number of strata
cornea counted at the end of the experimental period.
32 C. BARKER JaRGENSEN
Thirty-two days after the operation skin biopsies were taken, and the number of strata cornea was counted in order to cal- culate the mean duration of the intervals between replacements in the individual toads. Table 1 shows that the duration was the same in both groups, viz. 14.5 + 4.2 (SD) days in the toads with extirpated pars distalis and 14.3 2 4.8 in the other group. During the experimental period about 400 p,g cortisol had on an average been ab- sorbed per toad per day.
Intact toads. In an experiment including 10 male toads with implanted cortisol pel- lets none were observed to moult, and after 34 weeks slight, abnormal keratinization of the epidermis could be registered. After 34 days skin biopsies were taken and the pellets were removed. The mean duration of intervals between replacement of strata cornea was 17.7 ? 8.5 days, and the daily absorption of cortisol amounted to about 500 kg (Table 1).
Presumably, 1 month of exposure to mas- sive doses of circulating cortisol depresses the secretion of ACTH from the pars dista- lis and thus the endogenous secretion of ad- renocortical hormones. It was therefore of particular interest to follow what happened to moulting in the intact toads after removal of the cortisol pellets. No moults were reg- istered on the first 3 days, but on Day 5 all out of eight toads had moulted. (In one toad the pellet broke and was not completely re- covered, and another toad died for un- known reasons on the day after removal of the pellet.) Because the toads were not in- spected on Day 4, the day of moulting can- not be precisely stated, but the interval be- tween removal of the pellets and the first moult probably lasted 4 days in most or all of the toads.
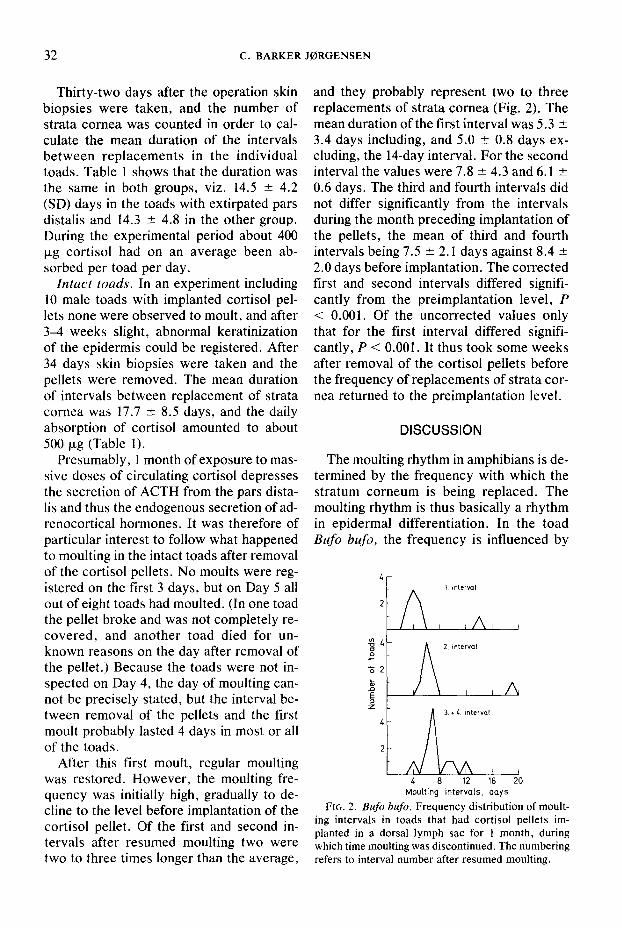
After this first moult, regular moulting was restored. However, the moulting fre- quency was initially high, gradually to de- cline to the level before implantation of the cortisol pellet. Of the first and second in- tervals after resumed moulting two were two to three times longer than the average,
and they probably represent two to three replacements of strata cornea (Fig. 2). The mean duration of the first interval was 5.3 & 3.4 days including, and 5.0 + 0.8 days ex- cluding, the 1Cday interval. For the second interval the values were 7.8 + 4.3 and 6.1 -+ 0.6 days. The third and fourth intervals did not differ significantly from the intervals during the month preceding implantation of the pellets, the mean of third and fourth intervals being 7.5 + 2.1 days against 8.4 + 2.0 days before implantation. The corrected tirst and second intervals differed signifi- cantly from the preimplantation level, P < 0.001. Of the uncorrected values only that for the first interval differed signifi- cantly, P < 0.001. It thus took some weeks after removal of the cortisol pellets before the frequency of replacements of strata cor- nea returned to the preimplantation level.
DISCUSSION
The moulting rhythm in amphibians is de- termined by the frequency with which the stratum corneum is being replaced. The moulting rhythm is thus basically a rhythm in epidermal differentiation. In the toad B&o bufo, the frequency is influenced by
Moulting intervals, days
FIG. 2. Bufo b&o. Frequency distribution of moult- ing intervals in toads that had cortisol pellets im- planted in a dorsal lymph sac for 1 month, during which time moulting was discontinued. The numbering refers to interval number after resumed moulting.

MOULTING IN TOADS 33
the level of circulating adrenocortical hormone(s). When hormone is lacking, as in toads with the pars distalis of the hy- pophysis extirpated or ectopically trans- planted, the frequency of replacement of strata cornea is greatly enhanced (Jorgensen and Larsen, 1960, 1961, 1964; Budtz 1979, 1985b). The investigations re- ported in this paper show that pharmaco- logical levels of an adrenocortical hormone, cortisol, increased the duration of the inter- vals between replacements from about 9 days to about 18 days in intact toads and to about 14-15 days in toads with the pars distalis extirpated or transplanted. The dif- ference between the groups of intact and of operated toads was not significant. These findings agree with those of Budtz (1988).
A replaced stratum corneum is mostly shed immediately, but occasionally shed- ding may be postponed until a subsequent moult. In the absence of circulating adreno- cortical hormone the replaced strata cornea are not shed. The high levels of circulating hormone produced by the subcutaneous implantation of cortisol pellets also discon- tinued shedding, but in three instances, in toads with the pars distalis extirpated or ec- topically transplanted, moults were regis- tered after the insertion of the pellets, prob- ably in connection with the first replace- ments of strata cornea that occurred at the elevated hormone level. Shedding of a re- placed stratum corneum is thus not in prin- ciple incompatible with pharmacological levels of an adrenocortical hormone in the blood.
Removal of the cortisol pellets about 1 month after their insertion was followed, in the intact toads, by moulting >3 and ~5 days later, most or all of the toads probably moulting on the fourth day. After the first moult, the frequency of replacement of strata cornea was initially high, but re- turned gradually to the normal level. This sequence of events may be interpreted as follows: The massive and prolonged treat- ment with cortisol effectively depressed the secretion of ACTH. After removal of the
source, cortisol is rapidly eliminated from the organism. In man the half-life of cortisol is about 100 min (Dixon et al., 1967). The half-life of cortisol in the toad is not known, but the half-life of corticosterone is in the order of 1 hr (Buchmann et al., 1972). Rapid elimination also of cortisol is supported by the finding that the toads with extirpated pars distalis responded to cortisol with moulting 3 days after removal of the corti- sol pellets, indicating that cortisol had been absent from the organism for about 18 hr or more (Jorgensen and Larsen, 1964). Corti- sol was thus eliminated within 2 days of removal of the pellets. Moulting in the in- tact toads about 4 days after removal of the cortisol pellets therefore indicates that the hypothalamic-corticotropic complex was active again l-2 days after elimination of the exogenous adrenocortical hormone. The resumed secretion of ACTH caused the shedding of the strata cornea that had been replaced during the hormone-deficient period, intervening between the elimination of the exogenous and reactivation of the en- dogenous source of hormone.
The initially high and gradually decreas- ing frequency of replacement of strata cor- nea that followed the first spontaneous moult probably reflects gradually increas- ing levels of circulating adrenocortical hormone(s), indicating a slow recovery to normal activity of the hypothalamic- corticotropic-adrenocortical complex.
The basic pattern of cell proliferation and differentiation, including rhythmic replace- ment of stratum corneum, is similar in all amphibians (see Larsen, 1976). The mor- phological components of the moulting pat- tern are thus evolutionarily conservative. in contrast to the plasticity in the endocrine components that secure rhythmical re- placements of strata cornea. In urodeles. this rhythm depends upon thyroxine. the frequency of replacements, and thus moult- ing. increasing with increasing levels of thy- roxine (Jorgensen and Larsen, 1963; Os- born, 1936, 1963, 1964a, b).
This difference between urodeles and

34 C. BARKER JQRGENSEN
toads (and perhaps other anurans) in the endocrine control of the moulting rhythm may be quantitative rather than qualitative. Exogenous thyroxine can also increase the moulting frequency in toads, but moulting continues more or less normally in the ab- sence of circulating thyroid hormones (Jorgensen et al., 1965; Rosenkilde, 1982). It is not known whether adrenocortical hor- mones can reduce the frequency of moult- ing in urodeles.
Both in urodeles and toads the duration of the intervals between replacements of strata cornea thus varies with the level of circulating hormones, and replacements proceed in the absence of a corresponding rhythm in secretion of these hormones. The question of whether, in toads, the actual moult necessitates upon a temporarily dis- continued secretion of ACTH can therefore be answered in the negative.
Shedding of a replaced stratum corneum constitutes a complex of processes. As mentioned, shedding may fail altogether, and in rare instances the complex of shed- ding processes may proceed out of context with the replacement of a stratum corneum, or the components of the shedding complex may proceed disintegrated (Jorgensen and Larsen, 1964). Presumably, the formation of cocoons, observed in several groups of amphibians from arid habitats, depends upon such disintegration of the shedding complex. Cocoons are formed through omission of the shedding behavior, which results in the covering of the body with a layer of strata cornea interspaced with mu- coid material (Ruibal and Hillman, 1981; McClanahan et al., 1983). Cocoons are pro- duced during aestivation and greatly reduce the evaporative water loss.
Normal moulting thus depends upon the integrated function of a hierarchical sys- tem, composed of elements ranging in na- ture from morphological to physiological and to behavioral. Obviously, a functional system as complex as moulting is vulnera- ble. Presumably, the most robust element is
the rhythmical replacements of strata cor- nea. This process depends upon the pres- ence of hormones which act as metabolic factors. Also the successful integration of replacement and shedding of a stratum cor- neum may be hormonally dependent, but it remains to be elucidated exactly how and where hormones act.
ACKNOWLEDGMENTS
The critical reading by Dr. Paul Budtz and Dr. Lis Olesen Larsen greatly improved the manuscript. The work was supported by a grant from the Carlsberg Foundation.
REFERENCES
Bendsen, J. (1956). Shedding of the skin in the com- mon toad. Bufo bufo. Vidsk Medd. Dan. Nat. Hist. Foren. 118, 21 l-225.
Btichmann, N. B., Spies, I., and Vijayakumar, S. (1972). Hypophysial corticotropic function and its hypothalamic control in Bufo bufo (L.) evaluated by the plasma concentration of corticosterone. Gen. Comp. Endocrinol. 18, 306-314.
Budtz, P. E. (1977). Aspects of moulting in anurans and its control. Symp. 2001. Sot. London 39,317- 334.
Budtz, P. E. (1979). Epidermal structure and dynam- ics of the toad, B&o b&o, deprived of the pars distalis of the pituitary gland. J. Zool. 189, 57-92.
Budtz, P. E. (1985a). Epidermal tissue homeostasis. I. Cell pool size, cell birth rate and cell loss by moulting in the intact toad, Bufo bufo. Cell Tissue Kinet. 18, 521-532.
Budtz, P. E. (1985b). Epidermal tissue homeostasis. II. Cell pool size, cell birth rate and cell loss in toads deprived of the pars distalis of the pituitary gland. Cell Tissue Kinet. 18, 533-542.
Budtz, P. E. (1988). Epidermal tissue homeostasis. III. Effect of hydrocortisone on cell pool size, cell birth rate and cell loss in normal toads and in toads deprived of the pars distahs of the pituitary. Cell Tis. Kinet., in press.
Dixon, P. F., Booth, M., and Butler, J. (1967). The corticosteroids. In “Hormones in Blood” (C. H. Gray and A. L. Bacharach, Eds.), pp. 306-390. Academic Press, New York/London.
Houssay, B. A. (1949). Hypophyseal functions in the toad Bufo arenaram Hensel. Q. Rev. Biol. 24, I- 27.
Jorgensen, C. B. (1970). Hypothalamic control of hy- pophyseal function in anurans. In “The Hypothalamus” (L. Martini, M. Motta, and F.

MOULTING IN TOADS 35
Franschini, Eds.), pp. 649-661. Academic Press, New York.
Jorgensen, C. B. (1974). Integrative functions of the brain. In “Physiology of the Amphibia” (B. Lofts, Ed.), pp. l-51. Academic Press, New York.
Jorgensen, C. B., and Larsen, L. 0. (1960). Hormonal control of moulting in amphibians. Nature (Lon- don) 185, 244-245.
Jorgensen, C. B., and Larsen, L. 0. (1961). Molting and its hormonal control in toads. Gen. Comp. Endocrinol 1, 145-153.
Jorgensen, C. B., and Larsen, L. 0. (1963). Neuro- adenohypophysial relationships. Symp. Zool. Sot. London 9, 59-82.
Jorgensen, C. B., and Larsen, L. 0. (1964). Further observations on molting and its hormonal control in Bufo bufo (L.). Gen. Comp. Endocrinol. 4,389- 400.
Jorgensen, C. B., Larsen, L. O., and Rosenkilde, P. (1965). Hormonal dependency of molting in am- phibians: Effect of radiothyroidectomy in the toad Bufo bufo (L.). Gen. Camp. Endocrinol. 5, 248- 251.
Larsen, L. 0. (1976). Physiology of molting. In “Physiology of the Amphibia” (B. Lofts, Ed.), Vol. III, pp. 53-100. Academic Press, New York.
McClanahan, L. L., Ruibal, R., and Shoemaker, V. H. (1983). Rate of cocoon formation and its physiological correlates in a ceratophryd frog. Physiol. Zool. 56, 430-435.
McKeown, B. A. (1972). Prolactin and growth hor- mone concentrations in the plasma of the toad Bufo bufo (L.) following ectopic transplantation of the pars distalis. Gen. Comp. Endocrinol. 19, 167-174.
Osborn, C. M. (1936). The inhibition of molting in urodeles following thyroidectomy or hypophysec- tomy. Anat. Rec. 66, 257-270.
Osborn, C. M. (1963). Endocrine factors in molting in urodeles. Anat. Rec. 145, 341-342. [Abstract]
Osborn, C. M. (1964a). Further studies on molting in urodeles. Anat. Rec. 148, 393. [Abstract]
Osbom, C. M. (1964b). The inhibition of molting in Triturus viridescens by a single injection of I”’ Amer. Zool. 4. [Abstract 1711
Rosenkilde, P. (1982). The role of thyroid hormone in adult amphibians. In “Phylogenetic Aspects of Thyroid Hormones” (S. Suzuki, Ed.), pp. 91-106. Center for Academic Publications Japan, Tokyo.
Ruibal, R., and Hillman, S. (1981). Cocoon structure and function in the burrowing hylid frog. Pterno- hylu fodiens. J. Herpetol. 15, 403-408.
Stefano, F. J. E., and Donoso, A. 0. (1964). Hy- pophyso-adrenal regulation of moulting in the toad. Gen. Comp. Endocrinol. 4, 473-480.
Van Dongen, W. I.. Jorgensen, C. B., Larsen, L. 0.. Rosenkilde, P.. and van Dongen, P. G. W. J. (1966). Function and cytology of the normal and autotransplanted pars distalis of the hypophysis in the toad Bulo bulo (L). Gen. Comp. Endocrinol. 6, 491-518.





























