Embed Size (px)
Citation preview

This article was downloaded by: [Queensland University of Technology]On: 23 November 2014, At: 09:43Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Biological Rhythm ResearchPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/nbrr20
Neurosecretory cells in the optic lobesof the brain and activity rhythms in Lycosa tarentula (Araneae: Lycosidae)Jacqueline Kovoor a , Arturo Muñoz-cuevas a & Joaquin Ortega-Escobar ba Zoologie-Arthropodes , Museum National d'Histoire Naturelle , 61rue de Buffon, 75231, Paris, Franceb Departamento de Psicología Biológica , Universidad Autónomade Madrid , 28049, Madrid, EspañaPublished online: 03 Feb 2007.
To cite this article: Jacqueline Kovoor , Arturo Muñoz-cuevas & Joaquin Ortega-Escobar (2005)Neurosecretory cells in the optic lobes of the brain and activity rhythms in Lycosa tarentula (Araneae: Lycosidae), Biological Rhythm Research, 36:3, 237-253, DOI: 10.1080/09291010500051659
To link to this article: http://dx.doi.org/10.1080/09291010500051659
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &

Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

Neurosecretory cells in the optic lobes of the brain andactivity rhythms in Lycosa tarentula (Araneae: Lycosidae)
JACQUELINE KOVOOR1, ARTURO MUNOZ-CUEVAS1,
& JOAQUIN ORTEGA-ESCOBAR2
1Museum National d’Histoire Naturelle, Zoologie-Arthropodes, 61 rue de Buffon, 75231 Paris, France,
and 2Departamento de Psicologıa Biologica, Universidad Autonoma de Madrid, 28049-Madrid, Espana
AbstractSeveral paired groups of neurosecretory cells (NS) were identified in the dorsal cortical neurons of theoptic lobes of the brain of Lycosa tarentula (Araneae). Two large bottle-shaped cells (NS A1, A2) and acluster of ca. 20 smaller cells (NS B) were found between the lamina and medulla of the anterior medianeyes (AM). The forward oriented bundles of NS B axons run alongside large fibres linked to the synapticzones of the indirect eyes. In front of the arcuate body, an islet of about 10 fusiform cells (NS C1) sendsshort axons close to the internal cortical border. Other large cells (NS C2, C3) are found from themedulla of the AM to the anterior border of the central body. Their long axons end deeply in the brainneuropil. NS B and C1 function synchronously. The secretory cycles of NS A1 and A2 seem to be inopposition. The activity of these three types of NS depends on the phase of the day. Anatomicalrelationships of NS A, B and C1 with visual afferent/efferent fibres via synaptic buttons indicate a role ofthese cells in the modulation of circadian rhythms of visual and locomotor activity. On the other hand,NS C2 and C3, the functioning of which is not synchronous, might be involved in the modulation orcontrol of the elementary movements of L. tarentula when active or at rest.
Keywords: neurosecretion, optic lobes of the brain, histology, biological rhythms, Araneae, spiders
Introduction
According to recent studies, spontaneous locomotor activity and cyclic structural modifica-
tions of retinal rhabdoms of the chelicerate Lycosa tarentula (Araneae: Lycosidae) show a
circadian rhythmicity (Kovoor et al. 1995, 1999). It is also known that neurotransmitters and
neuromodulators such as serotonin, octopamine, other catecholamines and melatonin are
detectable in the brain of arachnids, and especially spiders (Fowler & Goodnight 1974; Meyer
et al. 1984; Carricaburu & Munoz-Cuevas 1987, 1998; Munoz-Cuevas & Carricaburu 1990,
2000; Schmid & Duncker 1993; Schmid et al. 1990; Seyfarth et al. 1990, 1993). The sites of
the secretion of neuromodulators or neurotransmitters in the brain cells have nevertheless
been rarely localized. A group of neurosecretory cells located in the suboesophageal ganglion,
close to the circum-oesophageal connectives, was indeed interpreted as the circadian clock of
Correspondence: J. Ortega-Escobar, Departamento de Psicologıa Biologica, Universidad Autonoma de Madrid, 28049-Madrid,
Spain. Tel. + 34-91-4975188. Fax: + 34-91-4975215. E-mail: [email protected]
Biological Rhythm Research
July 2005; 36(3): 237 – 253
ISSN 0929-1016 print/ISSN 1744-4179 online # 2005 Taylor & Francis Group Ltd
DOI: 10.1080/09291010500051659
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

visual sensitivity in a scorpion, Androctonus australis (Buthidae) (Fleissner & Schliwa 1977;
Fleissner & Heinrichs 1982; Fleissner & Fleissner 1985). Obviously similar cells were
described in Limulus polyphemus (Xiphosures) by Fahrenbach (1981) and Barlow et al.
(1989).
The anatomy and secretory cycle of several groups of neurosecretory cells found in the
optic lobes of L. tarentula are described below. The involvement of these cells in the
modulation or control of visual as well as spontaneous locomotor activity rhythms is suggested
and discussed.
Material and methods
L. tarentula was reared by J. O.-E. in Madrid to provide a sufficient number of adult females in
good condition.
Illumination rhythm
Selected specimens had been submitted to illumination cycles of 12 hours (L 8.00 h –D
20.00 h) at 1900 lx. The spontaneous locomotor activity rhythm of these animals were
registered during 15 days before they were treated for histological study. All treated animals
showed a ‘‘normal’’ locomotor activity rhythm, i.e. they were active at least for three hours
from the beginning of the dark phase, at about 20.00 h, as has been shown previously (Ortega-
Escobar, 2002).
Histology
Twelve adult females were studied. Sets of three of these animal prosomas were fixed in
Bouin’s aqueous fluid at the following times: 06.00 h, 12.00 h, 18.00 h and 24.00 h,
dehydrated, and embedded in paraffin. Serial sections, 7 mm thick, in the three orthogonal
planes, were treated by general histological stainings and histochemical reactions suited for
studying the visual system and the secretory cycle of neurosecretory cells in the optic lobes of
the brain. Structural conditions of photoreceptor rhabdoms were verified in all 12 animals.
Neurosecretory cells were characterized using aldehyde fuchsin and alcian blue on potassium
permanganate preoxidized sections, Mann-Dominici’s and Solcia et al.’s (1968) stainings,
and the p-dimethylaminobenzaldehyde reaction (Glenner & Lillie 1957). Technical
procedures are detailed in Gabe’s (1968) and Lillie and Fullmer’s (1976) handbooks.
Sections of male specimens previously studied were also treated. Furthermore, four females
which had been submitted to a 24 hour illumination were fixed at 06.00 h (two individuals) or
10.00 h (two individuals) and then treated similarly to the other 12 females.
Results
Anatomy
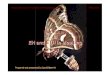
Three groups of neurosecretory cells have been identified at different levels of the optic lobes
(Figure 1). The most anterior group is localized at about 300 mm from the sagittal plane of the
prosoma and borders the fibre tract linking the first and second synaptic zones of the anterior
median retinae (AM S1 and AM S2). It comprises two symmetrical clusters, about 150 mm in
diameter, of some 20 odd particular cells in two categories (A and B) surrounded by globuli
(Figure 2).
238 J. Kovoor et al.
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

On each side, two bottle-shaped A cells are 110 mm long; their nucleus (30 x 20 mm)
is found in their basal part which is 50 mm broad at the most. One of these two cells
Figure 1. Parasagittal section of an optic lobe of the brain till the arcuate body (AB) showing several sites of
neurosecretory cell groups. Masson-Goldner trichrome, green filter. AM S2: second synaptic zone of an anterior
median retina. Scale bar: 100 mm.
Figure 2. The most anterior group of neurosecretory cells (NS A1, A2, B) in a transversal section of the prosoma. Part
of an optic lobe, lateral region is on the right-hand side. Periodic acid-Schiff-Groat’s hematoxylin-methyl blue, green
filter. Scale bar: 50 mm.
Neurosecretory cells in the brain of Lycosa tarentula 239
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

(NS A1) lies along the dorso-lateral border of the optic lobe, its proximal region is
inserted between the fibre tract succeeding to AM S1 and the cluster of NS B cells
and globuli (Figure 5). The basal part of NS A2 is more internally applied on the
ventral face of the fibre tract; its neck is elongated amongst the globuli, toward the
neuropil (Figure 2).
The bunch of NS B cells is made up of closely packed fusiform somata, which send their
long straight axons as a tight bundle oriented forwards. These fibres run alongside efferent or
afferent fibres linked to the indirect eye synaptic zones, in the axial region of the optic lobes
(Figures 3 and 4). Synaptic buttons are clearly observed between the NS B processes and the
visual interneurons (Figure 4).
Cortical cell layers comprising other neurosecretory cells lie between the second
synaptic zone of the anterior median retinae (AM S2) and the arcuate body (AB). Just
next to the anterior lobe of the arcuate body, an oval cluster of about 10 cells (NS
C1) is enclosed in the internal half of cortical cells (Figure 6). These NS C1 are
fusiform, 72 – 75 mm long, with a nucleus 9 mm in diameter in females and 55 mm long
with a nucleus 12 – 13 mm in diameter in males. Their very thin processes meet
afferent/efferent fibres linked to AM S2, in the neuropil bordering the cortical cells
(Figure 7).
Large-sized neurosecretory cells (NS C2, NS C3) are more or less aligned or sparse
alongside various other types of neurons of the optic lobe dorsal cortex which precedes the
arcuate body. The round distal part of NS C3 contains the nucleus (diameter:14 mm) and lies
Figure 3. Cluster of NS B cells and axons. Horizontal section. Ferric ferricyanide-fast nuclear red, orange filter. Scale
bar: 25 mm.
240 J. Kovoor et al.
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

on the cortical basal lamina. Its shape is that of an ampulla, the very long neck (15 mm wide)
of which is inserted between other cortical cells. NS C2 and NS C3 differ by their total lengths
only, which are 80 or 100 – 120 mm, respectively. Long processes of these cells reach the
midbrain neuropil.
Figure 4. Axons of NS B cells contact wide interneurons through synaptic buttons. Transverse section, plumbic
hematoxylin, green filter. Scale bar: 25 mm.
Figure 5. NS A1 cell covering the basal part of NS B cells. Transverse section, one-step trichrome, orange filter. Scale
bar: 25 mm.
Neurosecretory cells in the brain of Lycosa tarentula 241
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

Secretory cycle of neurosecretory cells
Cytological features of NS A1 and NS A2, especially seen when sections are stained by
aldehyde fuchsin or alcian blue after potassium permanganate oxidization, suggest a non-
synchronous functioning, or even opposite secretory cycles of these two cells. In NS A1,
Figure 6. NS C1 cells (arrow) close to the arcuate body (AB). Periodic acid-Schiff-Groat’s hematoxyline-methyl blue,
green filter. C2, C3 :NS C2, NS C3. Scale bar: 50 mm.
Figure 7. The cluster of NS C1 and its connexion (arrow) to fibres linked to the AM retina second synaptic zone.
Parasagittal section. Periodic acid-Schiff-Groat’s hematoxylin, green filter. Scale bar: 25 mm.
242 J. Kovoor et al.
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

synthesis of the secretory products takes place during the diurnal and partly nocturnal phases
(Figure 8); at 24.00 h, secretion accumulates in the apical cytoplasm of the cell body (Figure
10); at 06.00 h it is released (Figure 11). Signs of synthesis appear only at 18.00 h in NS A2
cytoplasm (Figure 8). While secretory products do not figure in the cell at any time, the
cytoplasm appears most often as a loose network (Figure 10).
All NS B cells function synchronously. Synthesis of neurosecretory products occurs mostly
during the illuminated phase; at 18.00 h, secretion is present in the apical half of the cells,
above the nucleus. At 24.00 h, NS B cells are still full of secretion (Figures 9 and 10); at
06.00, aldehyde fuchsin staining, rather weak, indicates that a neurosecretory product has
been released at least partly (Figure 11).
Cytological variations throughout the day are seen in NS C1 similar to those of NS B.
Neurosecretion accumulates during the light phase; cells are full at 18.00 h (Figures 6 and 7)
and the product reaches their apical end at 24.00 h, just before release (Figures 12 and 13).
Neurosecretion release seems to be the signal of the diurnal restoration of the rhabdoms of the
median anterior retinae which can be observed several hours later. Synthetic activity of NS C1
occurs continuously the whole day; nevertheless, it seems to be at a minimum at 06.00 h,
since the diameter of rhabdoms in the anterior median retinae is also the smallest.
Largest neurosecretory cells in the optic lobes, NS C2 and NS C3, the processes of which
deeply plunge in the brain neuropil, show diverse cytological features whatever the phase of
the day. At the four experimental times, some NS C2 or NS C3 are full of secretion while
others show numerous ergastoplasmic lamellae and a large nucleolus or seem to have released
their secretory product (Figure 14).
In the particular case of the animals submitted to 24 hours of continuous illumination, and
fixed at 06.00 h or 10.00 h, NS A1 and NS A2 had not released their secretory products. On
the contrary, the cytoplasm of NS A1 shows characteristics of a synthetic activity: numerous
ergastoplasmic lamellae and a large nucleolus in the nucleus. NS A2 also showed a large
nucleolus while the cytoplasm contained dense masses surrounded by lamellae (Figure 15).
On the other hand, NS C2 and NS C3 were filled with secretion (Figures 16 and 17). These
animals had been completely motionless during their lengthened illumination phase.
Figure 8. NS A1, A2 and B cells at 18.00 h. Cytological aspect of secretion elaboration in all cells (in black). Mann-
Dominici, orange filter. Scale bar: 50 mm.
Neurosecretory cells in the brain of Lycosa tarentula 243
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

Discussion
Among the neurosecretory cells which have been described above in the optic lobes of L.
tarentula, only NS C2 and NS C3 may be assimilated to the aboral neurosecretory cells
decribed by Kuhne (1959: ‘‘aborale protecerebrale Zellgruppe’’) in Pardosa lugubris
(Lycosidae), Tegenaria derhami and Araneus cornutus, and later by Meier (1967) in thomisids
and dysderids. The presence of these cells is thus considered to be a common feature of all
spiders. Other groups of neurosecretory cells, NS A, B and C1, were not mentioned by these
two authors in their inventory of the whole central nervous system of spiders. The localization,
Figure 10. NS A1, A2 and B cells at 24.00 h. Note the opposite aspects of NS A cells. Phosphotungstic hematoxylin,
green filter. Scale bar: 50 mm.
Figure 9. NS B cells at 24.00 h.. Secretion is accumulated in the supranuclear region of the cells (arrow heads).
Mann-Dominici, orange filter. Scale bar: 20 mm.
244 J. Kovoor et al.
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

and anatomical functional characteristics of the NS cells described here, with reports
concerning other arthropods, suggest that neurosecretion in the optic lobes is involved in the
control of circadian rhythms of locomotor and visual activity in Lycosa tarentula.
Insects
In numerous insects, especially in cockroaches and crickets and also in Drosophila, it has been
demonstrated that secretions of neurosecretory cells found in the optic lobes of the brain are
Figure 12. NS C1 cells at 23.00 h. Oxidization-aldehyde fuchsin-one-step trichrome, green filter. Scale bar: 50 mm.
Figure 11. NS A1 and B at 06.00 h. Secretion (in black) is present in the neck of NS A (arrow), and near the apex of
NS B cells (arrow heads). Oxidization-aldehyde fuchsin-one-step trichrome, green filter. Scale bar: 25 mm.
Neurosecretory cells in the brain of Lycosa tarentula 245
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

involved in the regulation of locomotor and/or visual activity (see review article by Helfrich-
Forster et al. 1998).
Chelicerates
The circadian rhythmicity of retinal sensitivity has been studied in the chelicerate Limulus
polyphemus (Barlow et al. 1977; Chamberlain & Barlow 1979). Lateral eyes are more sensitive
during the night than during the day. In these eyes, the activity of the efferent innervation is
Figure 13. Detail of Figure 12. Trails (black) of secretion leave the cells which contact fibres linked to the second
synaptic zone of an anterior median retina. Scale bar: 25 mm.
Figure 14. NS C3 cells at 18.00 h. Different cytological aspects of these cells. Oxidization-aldehyde fuchsin-one-step
trichrome, green filter. Scale bar: 50 mm.
246 J. Kovoor et al.
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

controlled by a group of neurons of the cheliceral ganglion (Calman & Battelle 1991). But it
has not yet been ascertained that these neurons solely constitute the circadian clock (Calman
& Battelle, 1991).
The sensitivity of the median eyes of the scorpion Androctonus australis shows a
circadian rhythm. Circadian impulses are transmitted to the optic nerve via
neurosecretory fibres the cell bodies of which are situated in the tritocerebrum
(Heinrichs & Fleissner 1987). As in Limulus, these neurons may be the circadian clock
or a part of it only.
Figure 15. NS A1, A2 and B cells at 06.00 h., after a 24 hours illumination. Plumbic hematoxylin, green filter. Scale
bar: 25 mm.
Figure 16. NS C3 cells at 06.00 h., after a 24 hours illumination. Oxidization-aldehyde fuchsin-trichrome, green
filter. Scale bar: 50 mm.
Neurosecretory cells in the brain of Lycosa tarentula 247
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

In spiders Argiope amoena and A. bruennichii, Yamashita and Tateda (1981) found that
there is an efferent input to the retina, generated by an endogenous circadian clock, that
enhances the sensitivity of the retina. More recently, Yamashita (2002) has studied the
circadian oscillation of sensitivity in the anterior median eye of several species (the garden
spider Araneus ventricosus, the jumping spider Menemerus confusus, and the orb-weaving
spiders Argiope amoena and A. bruennichii). While Araneus and Argiope show a circadian
oscillation of sensitivity, Menemerus do not show any.
In several spiders, fibres have been found in the retina that show neurosecretory
morphological characteristics. For example, Uehara et al. (1993) found efferent fibres in
the retina of the liphistiid spider Neptathela kimurai; these fibres contained both small clear
vesicles and large electron dense granules, and branched repeatedly with varicosities
providing sinaptoid contacts with the receptor cells. In a wolf spider, Lycosa erythrognata or
L. thorelli, Melamed and Trujillo-Cenoz (1966) noted the presence of ‘‘clear fibres’’ that
contained numerous dark granules and clusters of small vesicles. They observed specialized
areas of contact between these fibres and the photoreceptor cells.
In the spider Lycosa tarentula, the situation is more complex than in other chelicerates
previously studied. The former is a burrowing species, both day and night active; visually
guided behaviour and locomotor activity are synchronized by the visual system (Ortega-
Escobar, 2002). On the other hand, each pair of eyes of L. tarentula exhibits its own functional
characteristics: the rhythm of rhabdom renewal is proper to each retina type (Kovoor et al.
1995, 1999). Optic masses corresponding to the anterior median retinae are isolated from
those of the three other pairs of indirect retinae (anterior lateral AL, posterior lateral PL and
posterior median PM) in a dorsal superficial region of the optic lobes and linked to the arcuate
body.
In the animals studied, the rhabdom diameter in the anterior median retinae were at a
minimum at about 06.00 h and a maximum at 18.00 h. Variations of the diameter, as well as
the surface, of rhabdoms were rather small as compared to those in the indirect retinae;
nevertheless, we assume that the secretory product of NS C1, clustered in the vicinity of the
arcuate body, is involved in the modulation of the circadian modifications of AM rhabdoms.
The secretion of these cells would pass along the fibres linked to the synaptic zones of AM. Its
Figure 17. Detail of Figure 16. Scale bar: 25 mm.
248 J. Kovoor et al.
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

release would be the signal for initiating the restoration of the rhabdoms which can be
observed several hours later. Connexions of NS C1 axons have been detected with fibres of
the AM tract only. Anterior median eyes are not at all involved in the control of entrainment
of locomotor rhythm (Ortega-Escobar, 2002). It can thus be concluded that NS C1 are
unlikely to be modulators or oscillators of this rhythm.
On the other hand it can be suggested, from the observation of numerous synaptic
button-like between NS B axons and interneurons joining the synaptic zones of the
indirect retinae (cf. Figure 5), that NS B could act as a modulator or even oscillator of the
rhythmic changes of the rhabdoms in AL, PL, and PM retinae, and also of locomotor
rhythm. It was demonstrated by Ortega-Escobar (2002) that when anterior lateral eyes
only are left uncovered, spontaneous locomotor rhythm of L. tarentula tallies with an
advance or delay of the illumination cycle. The release of the NS B secretory product
between 24.00 h and 06.00 h would have the effect of stimulating rhabdom restoration in
AL retinae, for example, which takes place some hours later, while locomotor activity
would be abolished.
A correlation between a particular phase of the secretory cycle of NS A, C2, and C3, with
the initiation of spontaneous locomotor activity, which occurs at about 20.00 h in L. tarentula,
is difficult to establish. Nevertheless, data obtained from several females submitted to
24 hours of continuous illumination, and fixed in the morning, has led us to suggest the
following. In the two NS A of these animals, the cytoplasm appeared to be ‘‘blocked’’ in
synthetic phase of the secretory cycle, while the release of their products should have occurred
before fixation. In the same animals, all NS C2 and C3 were full of secretion. These animals
had been totally motionless all through the 24 hours. It is thus quite possible that the secretory
products of NS A and NS C2 and C3 are involved in the regulation of certain aspects of the
spontaneous locomotor activity in L. tarentula. NS A might represent a part of the circadian
clock for locomotor activity; NS C2 and C3 which send their processes deeply into the central
neuropil of the brain might establish connexions with neuromotor fibres or interneurons
linked to the suboesophageal ganglion, especially to locomotor appendage ganglions.
Gronenberg (1990), analysing the functional organization of sensory interneurons in the
nervous system of Cupiennius salei (Araneae, Ctenidae), described ‘‘ascending’’ interneurons
from the longitudinal tracts of the suboesophageal ganglion to the brain, and other
‘‘descending’’ interneurons from the posterior part of the corpora pedunculata (dorsal
anterior to the arcuate body) to median longitudinal tracts of the suboesophageal ganglion.
Although no precise study has been made on these type of interneurons and their
arborisations, anatomical data already collected (Kovoor et al. in press) in the optic lobes
suggest that interneuron organization of L. tarentula is close to that of C. salei. That
strengthens our hypothesis.
Most probably the various groups of neurosecretory cells in the optic lobes of L. tarentula
have distinct functions: their secretory products control or modulate only a part of the visual
activity (NS C1), or operate at the same time on visual and spontaneous locomotor activities
(NS B), or even modulate only locomotor activity (NS A, NS C2 and C3).
Neurotransmitters, neuromodulators
The identification of neurotransmitters or neuromodulators produced by neurosecretory cells
described above can be only speculative, based on biochemical and physiological studies
made on arthropods, among which are some chelicerates. The main candidates which may be
proposed are serotonin (5-hydroxytryptamine), octopamine, dopamine, adrenaline and
noradrenaline, and also melatonin.
Neurosecretory cells in the brain of Lycosa tarentula 249
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

Serotonin
According to Muszynska-Pytel and Cymborowsky (1978a, 1978b), a correlation does exist
between serotonin concentration in the central body, rich in neurosecretory elements, a
secretion of neurosecretory cells of the pars intercerebralis, and the locomotor activity of
crickets (Acheta domestica) during the day. Control of the locomotor rhythm of crickets
depends on the opposite activities of two neuromodulators. The pars intercerebralis product
has not been identified.
In chelicerates, serotonin was rarely detected. The cyclic production of secrotonin related
to locomotor activity was demonstrated in Leiobunum longipes (Opiliones) by Fowler and
Goodnight (1966, 1974). Injections of serotonin in the dorsal vessel of other opilions
(Discocyrtus dilatatus, Gonyleptidae) cause a modulation of the amplitude of electroretino-
grams which is greater in the day-time than at night (Munoz-Cuevas & Carricaburu, 1990).
The production site of the neuromodulator is unknown. However, several groups of
immunoreactive, serotonin-like cells were identified in the suboesophageal ganglion and
cheliceral neuromers, but not in the optic lobes, of a spider, Cupiennius salei (Ctenidae)
(Seyfarth et al., 1990). Besides, a histochemical demonstration of serotonin, by the p-
dimethylaminobenzaldehyde reaction, failed in the brain of L. tarentula.
Octopamine
It was well established in the chelicerate Limulus that the circadian rhythm of sensitivity of the
lateral retinae was mainly controlled by octopamine (Battelle 1980, 1990: Battelle & Evans
1984; Evans et al. 1983; Calman & Battelle 1991; Lee & Wise 1991) transiting via efferent
fibres of brain cells surrounded by ganglionic cells of the medullar group (Chamberlain &
Barlow 1980). Similarly in the scorpion Androctonus australis, octopamine released by two
symmetrical groups of cells and their efferents localized in the brain is also the main
neurotransmitter governing the retinal sensitivity (Fleissner & Fleissner 1985).
Octopamine was not found in the brain of spiders as a neurotransmitter controlling the
visual or locomotor systems. However, Seyfarth et al. (1993) have localized octopamine, by
immunocytochemistry, in a series of neurosecretory elements of the suboesophageal and
cheliceral ganglions of Cupiennius salei. The arborizations of these neurons may diffuse
octopamine towards the arcuate body which does not produce this neurotransmitter by itself.
The rhythm of secretion of octopamine has not been studied in C. salei.
Catecholamines
According to the studies of Meyer et al. (1984), adrenaline, noradrenaline and dopamine are
present in the central nervous system of spiders. The quantities of these neurotransmitters are
higher in vagrant spiders such as Lycosidae and Salticidae the behaviour of which is visually
guided, than in Theraphosidae, or sedentary web spiders such as Agelenidae and Araneidae.
Exogenous dopamine and noradrenaline, and also modafinil (R), result in variations of the
electroretinograms of L. tarentula depending on the type of retina recorded and the phase of
activity (Carricaburu & Munoz-Cuevas 1998; Munoz-Cuevas & Carricaburu 2000).
Melatonin
Melatonin has been found almost throughout the animal kingdom. Its role is important as a
neurotransmitter controlling circadian rhythms adjusted to alternative light and darkness
250 J. Kovoor et al.
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

(Vivien-Roels & Pevet 1986). Melatonin was demonstrated for the first time in the eyes and
brain of a spider (L. tarentula) in 1998 (unpublished results; dosages of melatonin by
radioimmunoassays were carried out in the Laboratory of Dr. B. Vivien-Roels, Laboratoire de
Neurobiologie des Rhytmes, Universite Louis Pasteur, Strasbourg (France) whom we warmly
thank for her cooperation). Nocturnal concentrations were higher than diurnal concentra-
tions. Injected in the dorsal vessel of the spider, melatonin causes an increase of nocturnal
responses (ERG) and has little effect on diurnal responses of indirect retinae the rhythmic
activity of which is more diurnal than nocturnal (unpublished results).The reverse is observed
when dopamine or noradrenaline are tested: catecholamines and melatonin are thus active in
opposing phases on the indirect eyes of L. tarentula. In the anterior lateral eyes of the orb-
weaving spiders, Argiope amoena and A. bruennichii, Yamashita and Arita (1995) could change
the ERG waveform by exogenous application of octopamine during the day. In contrast,
exogenous application during night-time had little or no effect.
Melatonin and catecholamines represent good candidates to be neuromodulators or
neurotransmitters of locomotor and/or visual activities of L. tarentula. We suggest that NS A
and NS B cells could rhythmically synthesize and diffuse them into the brain.
Immunocytochemical tests should be carried out, especially in the sites described above, to
answer the still open questions concerning circadian clocks and activity rhythm modulation in
spiders.
References
Barlow Jr RB, Bolanowski SJ, Brachman ML. 1977. Efferent optic fibers mediate circadian rhythms in the Limulus
eye. Science 197:86 – 89.
Barlow Jr RB, Chamberlain SC, Lehman HK. 1989. Circadian rhythms in the invertebrate retina. In: Stavenga DG,
Hardie RC, editors. Facets of Vision. Berlin: Springer-Verlag, p 235 – 256.
Battelle B-A. 1980. Neurotransmitter candidates in the visual system of Limulus polyphemus :Synthesis and
distribution of octopamine. Vision Res 20:911 – 922.
Battelle B-A. 1990. Regulation of retinal functions by octopaminergic efferent neurons in Limulus. In: Osborne N,
Chader J, editors. Prog. Retin. Res. 10. Berlin: Springer-Verlag. p 333 – 355.
Battelle B-A, Evans JA. 1984 Octopamine release from centrifugal fibers of the Limulus peripheral visual system. J
Neurochem 42:71 – 79.
Calman BG, Battelle BA. 1991. Central origin of the efferent neurons projecting to the eyes of Limulus polyphemus.
Visual Neurosci 6:481 – 495.
Carricaburu P, Munoz-Cuevas A. 1987. La modulation du rythme visuel circadien par l’Octopamine chez les
scorpions et l’adaptation a la vie souterraine. C R Acad Sci Serie III Sci Vie 305:285 – 288.
Carricaburu P, Munoz-Cuevas A. 1998. Action du Modafinil sur l’electroretinogramme de Lycosa tarentula en
relation avec le rythme d’activite visuelle (Araignees, Lycosidae). C R Soc Biol 192:1163– 1173.
Chamberlain SC, Barlow RB Jr. 1979. Light and efferent activity control rhabdom turnover in Limulus
photoreceptors. Science 206:361 – 363.
Chamberlain SC, Barlow RB. 1980. Neuroanatomy of the visual efferents in the horse shoe crab (Limulus
polyphemus). J Comp Neurol 192:387 – 400.
Evans JA, Battelle B-A, Chamberlain SC. 1983. Autoradiographic localization of newly synthetized octopamine to
retinal efferents and the Limulus visual system. J Comp Neurol 219:369 – 383.
Fahrenbach WH. 1981. The morphology of the Limulus visual system. VII. Innervation of photoreceptors by
neurosecretory efferents. Cell Tissue Res 216:655 – 659.
Fleissner G, Fleissner G. 1985. Neurobiology of a circadian clock in the visual system of scorpions. In: Barth FG,
editor. Neurobiology of Arachnids. Berlin: Springer-Verlag. p 351 – 375.
Fleissner G, Heinrichs S. 1982. Neurosecretory cells in the circadian-clock system of the scorpion, Androctonus
australis. Cell Tissue Res 224:233 – 238.
Fleissner G, Schliwa M. 1977. Neurosecretory fibres in the median eyes of the scorpion; Androctonus australis L. Cell
Tissue Res 178:189 – 198.
Fowler DJ, Goodnight CJ. 1966. The cyclic production of 5-hydroxytryptamine in the opilionid. Am Zool 6:187 –
193.
Neurosecretory cells in the brain of Lycosa tarentula 251
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

Fowler DJ, Goodnight CJ. 1974. Physiological populations of the arachnid Leiobunum longipes (Opilione-
s : Phalangiidae). Syst Zool 23:219 – 225.
Gabe M. 1968. Techniques Histologiques. Paris: Masson editeur.
Glenner GG, Lillie RD. 1957. The histochemical demonstration of indole derivatives by the post-coupled p-
dimethylaminobenzilidene reaction. J Histochem Cytochem 5:279 – 297.
Gronenberg W. 1990. The organization of plurisegmental mechanosensitive interneurons in the central nervous
system of the wandering spider Cupiennius salei. Cell Tissue Res 260:49 – 61.
Heinrichs S, Fleissner G. 1987. Neuronal components of the circadian clock in the scorpion Androctonus australis:
Central origin of the efferent neurosecretory elements projecting to the median eyes. Cell Tissue Res 250:277 –
285.
Helfrich-Forster C, Stengl M, Homberg U. 1998. Organization of the circadian system in insects. Chronobiol Int
15:567 – 594.
Kovoor J, Munoz-Cuevas A, Ortega-Escobar J. 1995. Diel morphological changes in the photoreceptors of Lycosa
tarentula (Araneae, Lycosidae). Biol Rhythm Res 26:272 – 291.
Kovoor J, Munoz-Cuevas A, Ortega-Escobar J. 1999. Circadian structural changes in the retina of Lycosa tarentula
(Araneae, Lycosidae). Biol Rhythm Res 30:407 – 423.
Kovoor J, Munoz-Cuevas A, Ortega-Escobar J. In press. The visual system of Lycosa tarentula (Araneae: Lycosidae).
Microscopic anatomy of the protocerebral optic centres. The Italian Journal of Zoology.
Kuhne H. 1959. Die neurosekretorischen Zellen und der retrocerebrale Komplex von spinnen. Zool Jb Anat 78:528 –
599.
Lee HM, Wyse GA. 1991. Immunocytochemical localization of octopamine in the central nervous system of Limulus
polyphemus : a light and electron microscopic study. J Comp Neurol 307:683 – 694.
Lillie RD, Fullmer HM. 1976. Histopathology Technic and Practical Histochemistry. 4th edition. NewYork:
MacGraw-Hill Book Company.
Meier F. 1967. Beitrage zur Kenntnis der postembryonalen Entwicklung der Spinnen (Araneida, Labidognatha),
unter besonderer Berucksichtigung der Histogenese des Zentralnervensystems. Rev Suisse Zool 74:1 – 127.
Melamed J, Trujillo-Cenoz O. 1966. The fine structure of the visual system of Lycosa (Araneae: Lycosidae). Part I.
Retina and optic nerve. Z Zellforsch 74:12 – 31.
Meyer W, Schlesinger C, Poehling HM, Ruge W. 1984. Comparative quantitative aspects of putative
neurotransmitters in the central nervous system of spiders (Arachnida : Araneida). Comp Biochem Physiol
78C:357 – 362.
Munoz-Cuevas A, Carricaburu P. 1990. Modulation in electroretinograms during 24 hours by serotonin of
Discocyrtus dilatatus (Chelicerata, Opiliones, Gonyleptidae). Acta Zool Fenn 190:287 – 291.
Munoz-Cuevas A, Carricaburu A. 2000. Electroretinographic modulation by dopamine and noradrenaline in the
spider Lycosa tarentula (Araneae: Lycosidae). Ekologia (Bratislava) 19:171 – 180.
Muszynska-Pytel M, Cymborowski B. 1978a. The role of serotonin in regulation of the circadian rhythms of
locomotor activity in the cricket (Acheta domesticus L.). I. Circadian variations in serotonin concentration in the
brain and hemolymph. Comp Biochem Physiol 59C:13 – 15.
Muszynska-Pytel M, Cymborowski B. 1978b The role of serotonin in regulation of the circadian rhythms of
locomotor activity in the cricket (Acheta domesticus L.) II. Distribution of serotonin and variations in different
brain structures. Comp Biochem Physiol 59C:17 – 20.
Ortega-Escobar J. 2002. Circadian rhythms of locomotor activity in Lycosa tarentula and the pathways of ocular
entrainment. Biol Rhythm Res 33:561 – 576.
Schmid A, Duncker M. 1993. Histamine immunoractivity in the central nervous system of the spider Cupiennius salei.
Cell Tissue Res 273:533 – 545.
Schmid A, Duncker M, Sporhase-Eichmann U. 1990. Verteilung FMRFamid-und GABA-artiger Immunreaktivitat
im ZNS der Jagdspinne Cupiennius salei. Verh Dts Zool Ges 83:639 – 640.
Seyfarth E-A, Hammer K, Grunert U. 1990. Serotonin-like immunoreactivity in the CNS of spiders. Verh Dts Zool
Ges 83:640.
Seyfarth E-A, Hammer K, Sporhase-Eichmann U, Horner M, Vullings HGB. 1993. Octopamine immunoreactive
neurons in the fused central nervous system of spiders. Brain Res 611:197 – 206.
Solcia E, Vassalo G, Capella C. 1968. Selective staining of endocrine cells by basic dyes after acid hydrolysis. Stain
Technol 43:257 – 263.
Uehara A, Uehara K, Ogawa K. 1993. Efferent fibers and daily rhabdomal changes in the anteromedial eye of the
liphistiid spider, Heptathela kimuraii. Cell Tissue Res 272:517 – 522.
Vivien-Roels B, Pevet P. 1986. Is melatonin an evolutionary conservative molecule involved in the transduction of
photoperiodic information in all living organisms? In: Reiter IRJ, Karasek M, editors. Advances in Pineal
Research. John Libbey and Co. Ltd., p 61 – 68.
252 J. Kovoor et al.
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014

Yamashita S. 2002. Efferent innervation of photoreceptors in spiders. Microsc Res Techn 58:356 – 364.
Yamashita S, Arita F. 1995. Efferent control in the anterior lateral eyes of orb-weaving spiders. Zool Sci 12:385 – 389.
Yamashita S, Tateda H. 1981. Efferent neural control in the eyes of orb-weaving spiders. J Comp Physiol A 143:477 –
483.
Neurosecretory cells in the brain of Lycosa tarentula 253
Dow
nloa
ded
by [
Que
ensl
and
Uni
vers
ity o
f T
echn
olog
y] a
t 09:
43 2
3 N
ovem
ber
2014