Embed Size (px)
Citation preview

YEAST VOL. 10 1075-1082 (1994)
nmt2 of Fission Yeast: A Second Thiamine-Repressible Gene Co-ordinately Regulated with nmtl ANDREA G. 0. MANETTI, MARC0 ROSETTO AND KINSEY G. MAUNDRELL*t
Department of Evolutionary Biology, University of Siena, Via Mattioli 4, 53100 Siena, Italy *Glaxo Institute, 14 Chemin des Aulx, 1228 Plan-les-Ouates, Geneva, Switzerland
Received 4 January 1994; accepted 27 March 1994
We previously described a screen for thiamine-repressible genes in Schizosaccharomyces pombe and reported on one such gene, nmtl, required for thiamine biosynthesis. Here we describe a second gene, nmt2, recovered in the same screen. Disruption of nmt2 also resulted in thiamine auxotrophy, indicating a role for the nmt2 gene product in thiamine biosynthesis. Both genes are highly transcribed in minimal medium and repressed in medium containing thiamine, and nuclear ‘run-on’ experiments c o n k that expression in both cases is controlled by the rate of transcription initiation. The virtually identical kinetics of induction and repression suggest that the two genes are co-ordinately regulated. Sequence comparison of the two promoters reveals a canonical TATA box, downstream of which is a perfectly conserved 11 bp element. Transcript mapping experiments show that transcription initiation of both genes is centred on this element.
KEY WORDS - Schizosaccharomyces pombe; thiamine; transcription; inducible promoter.
INTRODUCTION The general features of promoter structure and transcriptional control have been conserved throughout eukaryotic evolution from yeast to mammals (reviewed in Guarente and Bermingham- McDonogh, 1992). Extensive studies among yeasts have mostly focused on the budding yeast Sac- charomyces cerevisiae and have been greatly facili- tated by identification of several sets of genes under common control (Struhl, 1987a). These studies have revealed cis-acting ad trans-acting elements which are important for transcriptional control and which in some cases have been found to have surprisingly similar counterparts in mammalian cells (Struhl, 1987b; Vogt et al., 1987; Wiederrecht et al., 1987).
In the case of the fission yeast, Schizosaccharo- myces pombe, several lines of evidence suggest that the mechanisms involved in transcriptional regu- lation may be even more closely related to those in higher eukaryotes (Russell and Nurse, 1986; Jones et al., 1988; Prabhala et al., 1992). However, the dearth of well-characterized transcriptionally regu- lated genes in this organism has limited progress
?Corresponding author.
0 1994 by John Wiley & Sons Ltd CCC 0749-503X/94/08107548
on the characterization of the factors involved. We have previously reported the isolation of thiamine- sensitive genes by differential screening of a S. pombe genomic library using radiolabelled cDNA probes derived from cells grown in the presence o r absence of thiamine (Maundrell, 1990). Two genes strongly repressed by thiamine were identi- fied and designated nmtl and nmt2. The properties of nmtl were described previously (Maundrell, 1990). In the present report we describe in more detail the properties of nmt2 and show that its transcriptional regulation exactly parallels that of nmtl.
MATERIALS AND METHODS
Strains and media All S. pombe strains used were derived from the
wild-type 972-h- strain and are described in the text. Media and transformation are as described in Moreno et al. (1991). The bacterial strain for routine work was JMlOlTR (supE, thi-, srl::tnlO, recA A(1ac-proAB), [F, traD36, proAB, IacZZAM 151) and standard media were used throughout.

1076 A. G. 0. MANETTI ET AL..
data revealed two complete open reading frames transcribed in opposite directions. Northern blots probed with each end of the clone (data not shown) indicated that the left-hand end of the clone as shown in Figure la encodes the thiamine- repressible transcript. The nucleotide sequence of the left-hand HindIII-EcoRV is shown in Figure lb. An uninterrupted open reading frame of 903 nucleotides potentially encodes a protein of 32 kDa.
Screening the data base for proteins of similar sequence revealed a high homology (7 1% overall identity) with the stress protein, sti35, from Fusarium (Choi et al., 1990; see Discussion). The structure of the second open reading frame, referred to as avn2 will be described elsewhere.
The strain carrying an nmt2 null allele is a thiamine auxotroph
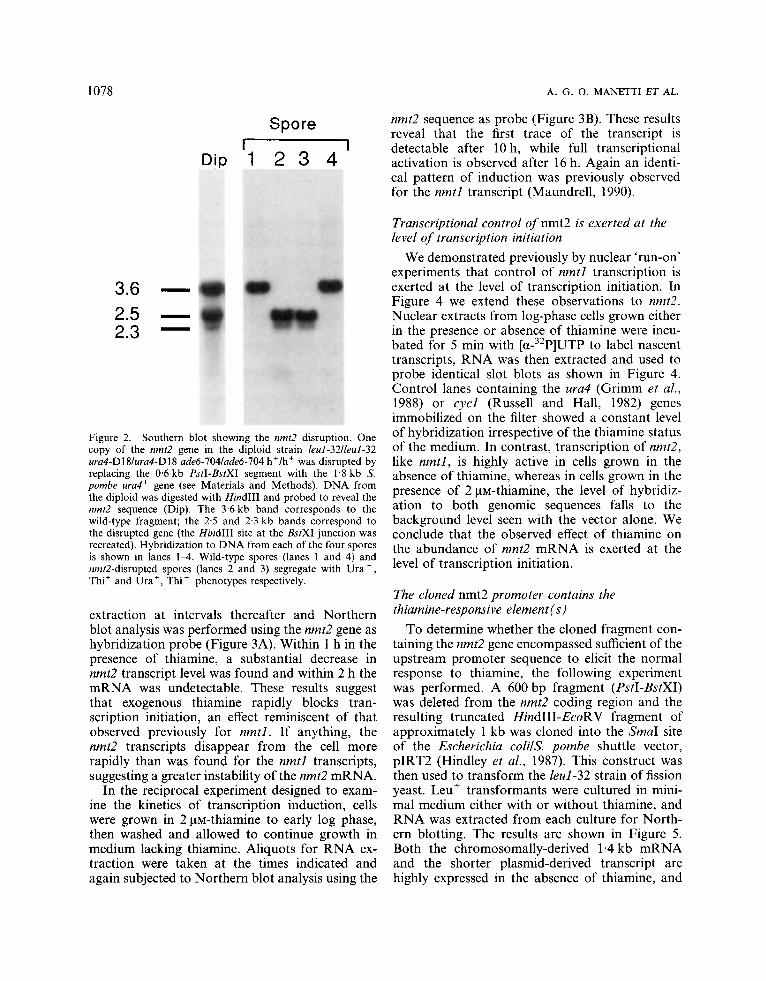
An nmt2 disruptant strain was constructed fol- lowing the procedure described by Grimm et al. (1988). The S. pombe ura4+ gene was inserted into the nmt2 coding sequence as described in Materials and Methods, fortuitously recreating the HindIII site at the BstXI junction. The disrupted gene was then isolated following digestion with PvuII, which cuts either side of the pUCll9 polylinker, and the resulting 5.1 kb fragment was used to transform the diploid S. pombe strain leul-32lleuI-32 ura4- D18Iura4-Dl8 add-704laded-704 h+/h+. DNA was prepared from a number of stable Ura+ transformants and analysed by Southern blotting after digestion with HindIII. Because of the re- created HindIII site, homologous integration into one chromosome of the diploid results in three hybridizing bands when the entire 3.6 kb fragment is used as probe: the undisrupted 3.6 kb band, a weakly hybridizing 2.3 kb band and a strongly hybridizing 2.5 kb band. This predicted pattern of hybridization was obtained in four of 11 stable integrants analysed (Figure 2, Dip). h+Ih9' diploid revertant strains identified by exposure to I, vapour were sporulated and tetrads dissected. All four spores were viable on complete medium and the ura4+ marker segregated 2:2 in each case. DNA made from each of the four spores was digested with HindIII and analysed by Southern blotting (Figure 2, lanes 14) . In all cases, a clear correlation was found between the Ura+ pheno- type and disruption of the wild-type nmt2 gene. Cells derived from each of the four spores were then cultured in medium lacking thiamine. The two nmt2-disrupted strains were not capable of
Analytica I procedures Procedures used for DNA cloning were as de-
scribed in Sambrook et al. (1989). RNA isolation and Northern blotting was described previously (Maundrell, 1990). The 5' end of the nmt2 mRNA was mapped by primer extension using a syn- thetic oligonucleotide 5' ACTGGCAAGTCAGT CTTGAA 3' complementary to the coding se- quence between ositions +40 to +59. The oligo- nucleotide was P2P-labelled with polynucleotide kinase and annealed and extended with reverse transcriptase as described elsewhere (Grimm et al., 1988).
For DNA sequence determination, restriction fragments were subcloned into pUC119 and pro- gressively resected with exonuclease I11 using the procedure described by Henikoff (1984). Certain regions were sequenced using specific oligonucleo- tide primers. DNA sequence data were handled using the UWGCG software (Devereux et al., 1984).
Transcriptional 'run-on' experiments were per- formed exactly as described previously (Maundrell, 1990).
To create the nmt2 gene disruption strain, the procedure of Grimm et al. (1988) was used. A 0.6 kb segment of the nmt2 coding sequence be- tween the PstI and BstXI sites was excised and replaced with the 1.8 kb fragment encoding the ura4+ gene. In this construct the HindIII site adjacent to the BstXI site was recreated. The disrupted gene was isolated as a linear fragment by digestion with PvuII, which cuts either side of the pUCll9 polylinker, and used to transform the diploid strain leu1 -321leul-32 ura4-Dl81ura4- D 18 ade6-704lade6-704 h+/h+ as described. Tetrad analyses were performed using standard pro- cedures (Gutz et al., 1974).
RESULTS
Isolation and sequence analysis of nmt2 The nmt2 gene was isolated on a 3.6 kb HindIII
fragment (Figure la) following a differential screening protocol designed to detect genes either induced or repressed by thiamine (Maundreil, 1990). This sequence was subcloned into pUC119 in both orientations and the nucleotide sequence was determined on both strands following progres- sive resection with exonuclease I11 (Henikoff, 1984). Some regions were sequenced using custom synthesized oligonucleotide primers. The sequence

SECOND GENE FOR THIAMINE BIOSYNTHESIS 1077
nmt2
H PP B E C C P N H
1 A A G T T A A A A A A T C A G A T C C A C T A C T A A T T T G C A A C C G A A A C T ~ T A T G T ~ C C ~ T T T ~ T A C T A C ~ G T T A T G A A C C C ~ ~ ~ ~ G T c)
101 T T G T T T G T A G T C A W V U L A A T G A A A C T C G A A A C A T C O T A A G
201 C G G T T C T G A ; L T A T T A T T C T T M G T T T ~ T ~ C C C C ~ A P A T A V V T P Q T A F K T D L P V E K T A
301 C T C A T A A C A C T G T A G T C A A A A C T G A A A T G G G T O C T T T A T C E N T V V K S E M G A L S K A Y P T Y S L D E S F S F A P I R E S
4 0 1 CACTGTMGCGTGCTATGTCGTCGTATTTCAG~CTT~T~TATGTGMTCC~CATTGTTATTGTT~~TCCTTCTGCA~CTTACT T V S R A M T R R Y F S D L D K Y A E S D I V I V G A G S A G L T
501 G W L G T T A C T A C A T T G G M C T C O C C O C C C T C A T T T G A A ~ T T ~ T A T C A T C ~ T ~ T G T T ~ T C ~ T G G T G G T ~ T T ~ T T G G T C C T C A G T T T A A Y Y I G T R R P D L K I A I I E A S V A P G G G A W L G G Q L F
601 T T A G T G C A A T C C T T G T T C G T ~ G C A G T G A T C T T T T C T T ~ T ~ T T C C T G T T C C T T ~ ~ T ~ ~ T G A T T A T G T T G T C C T ~ C A T G C G S A M V V R K P A D L F L N E I G V P Y E D E G D Y V V V K E A A
701 T C T T T T C A C W U ~ C A C T G T T A T C C C G T A C T T T C G C A C T T
801 G A T G G T A A ~ C G C A T T O C C C C T G T T G T T A ~ T T C G ; 901 A T T T C C T T G T T T C T G T A C T G G T C A T G A T ~ T C C A T T C G
L F T S T V H A R T L A L P N V K L F N A T A V E D L I V K E G K
D G K Q R I A G V V T N W T L V S L N E G L Q S C M D P N T I N A E
L V V S A T G E D G P F G A F C V K R L A S A Q L V S N L E I D ~ R
1001 T C C T T T A G A T A T G A A C C G T G T G A ~ C C T T A T T G T A A A G P L D U N R A E D L I V K G T R E V F P G ~ I V G G ~ E L S E F D
1101 G G T G C A A C C G T A T O O C A C C T C C C T ~ T A T G A T G T T
IZOI TGAAAAGTACTTGTAAAGA;LAATAGTGTAC=AGCAACCTTTTGATATTTCTACATTTGTTTTAGTCATT~TTTTACCATTACTA~C~TTTTT~~T~ 1301 CAATTTACTTATACT~~~TT~TACATTTACTT~CAGAAT~CCTGTTTTCTCGTAGTTTTGGTTA~T~T~GTTTTATGCTTTTCA~
1501 A A T A T A A T A T T T T A T A T T G G M C T M & . A A T T A A A T A Figure 1. Structure of the nmt2 gene. (a) Restriction map of the 3.6 kb genomic fragment which encompasses the nmf2 gene. This region of the fission yeast genome includes two convergent coding sequences. Northern blot analysis revealed that the left-hand 1.5 kb HindIII-EcoRV fragment encodes the thiamine-repressible nmf2 transcript (data not shown). The other gene, referred to as avn2, will be described elsewhere. Restriction sites referred to in the text are indicated: B, BstXI; C, Clal; E, EcoRV; H, HindIII; N, NdeI; P, Pstl. (b) Nucleotide sequence of the HindIII-EcoRV fragment containing nmf2. The nucleotide sequence was determined on both strands using a combination of exonuclease 111 digests and specific oligonucleotide primers. All ambiguities in the sequence were resolved. The arrow indicates the site of transcription initiation which occurs 24 bp downstream of a perfect TATA box. Transcription initiation is located within an 11 bp element (boxed) also present at the corresponding site in the nmtl promoter (see Figure 6).
G A N R M G P T S V V *
1401 GTAGTTTTGTACAATATAATTGGCTAACGARATATCA~CAGTTTAGAGT~CTGT~~~~TCTTGGTTATCTTCT~TTTCACA~TCGTT
sustained growth in the absence of thiamine, although as was found previously for the nmtl gene, residual growth carried on for several doub- lings, presumably until the high internal pools of thiamine became exhausted (Schweingruber et al., 1991; Tommasino and Maundrell, 1991).
nmt 1 and nmt2 are co-ordinately regulated As with nmtl described previously (Maundrell,
1990), nmt2 is abundantly transcribed in minimal
medium and is dramatically repressed when cells are grown in medium containing thiamine. In order to make a more detailed comparison of the pattern of transcription of these two genes, the kinetics of induction and repression of nmt2 in response to thiamine were studied. To study transcriptional repression, cells were grown in minimal medium until early log phase, at which point thiamine was added to a final concentration of 2 p ~ . Aliquots of cells were taken for RNA
1078 A. G. 0. MANETTI ET AL.
3.6 - 2.5 - 2.3 -
Figure 2. Southern blot showing the nmtZ disruption. One copy of the nmtZ gene in the diploid strain leul-32/leul-32 ura4-D18/ura4-D18 ade6-704/ade6-704 hf/h+ was disrupted by replacing the 0.6 kb PstI-BstXI segment with the 1.8 kb S. pombe u r d gene (see Materials and Methods). DNA from the diploid was digested with HindIII and probed to reveal the nmtZ sequence (Dip). The 3.6 kb band corresponds to the wild-type fragment; the 2.5 and 2.3 kb bands correspond to the disrupted gene (the HindIII site at the BstXI junction was recreated). Hybridization to DNA from each of the four spores is shown in lanes 1 4 . Wild-type spores (lanes 1 and 4) and nmt2-disrupted spores (lanes 2 and 3) segregate with Ura - , Thi+ and Uraf , Thi - phenotypes respectively.
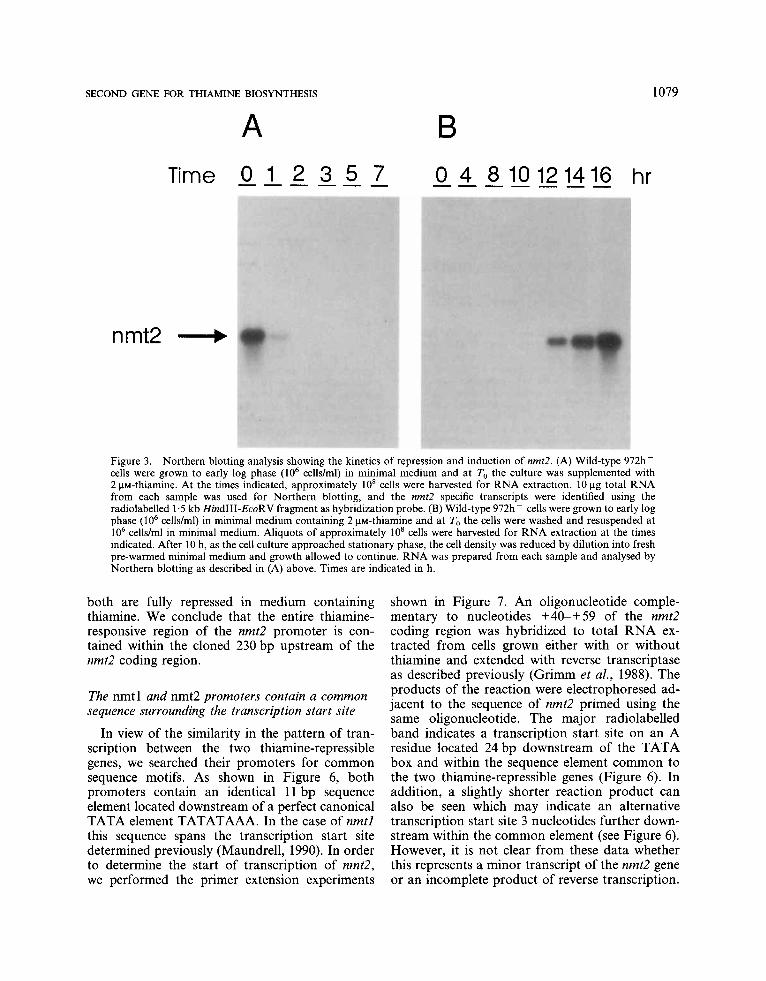
extraction at intervals thereafter and Northern blot analysis was performed using the nmt2 gene as hybridization probe (Figure 3A). Within 1 h in the presence of thiamine, a substantial decrease in nmt2 transcript level was found and within 2 h the mRNA was undetectable. These results suggest that exogenous thiamine rapidly blocks tran- scription initiation, an effect reminiscent of that observed previously for nmtl. If anything, the nmt2 transcripts disappear from the cell more rapidly than was found for the nmtl transcripts, suggesting a greater instability of the nmt2 mRNA.
In the reciprocal experiment designed to exam- ine the kinetics of transcription induction, cells were grown in 2yM-thiamine to early log phase, then washed and allowed to continue growth in medium lacking thiamine. Aliquots for RNA ex- traction were taken at the times indicated and again subjected to Northern blot analysis using the
nmt2 sequence as probe (Figure 3B). These results reveal that the first trace of the transcript is detectable after 10 h, while full transcriptional activation is observed after 16 h. Again an identi- cal pattern of induction was previously observed for the nmtl transcript (Maundrell, 1990).
Transcriptional control of nmt2 is exerted at the level of transcription initiation
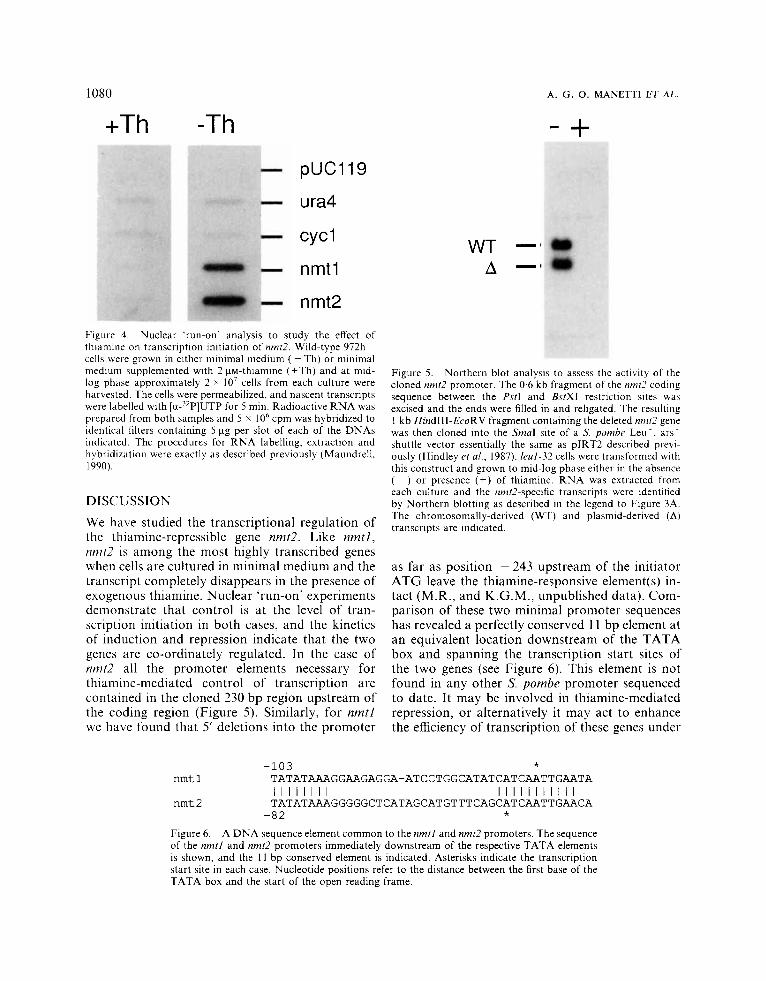
We demonstrated previously by nuclear ‘run-on’ experiments that control of nmtl transcription is exerted at the level of transcription initiation. In Figure 4 we extend these observations to nmt2. Nuclear extracts from log-phase cells grown either in the presence or absence of thiamine were incu- bated for 5 min with [CL-~~PIUTP to label nascent transcripts, RNA was then extracted and used to probe identical slot blots as shown in Figure 4. Control lanes containing the ural (Grimm et al., 1988) or cycl (Russell and Hall, 1982) genes immobilized on the filter showed a constant level of hybridization irrespective of the thiamine status of the medium. In contrast, transcription of nmt2, like nmtl, is highly active in cells grown in the absence of thiamine, whereas in cells grown in the presence of 2 yM-thiamine, the level of hybridiz- ation to both genomic sequences falls to the background level seen with the vector alone. We conclude that the observed effect of thiamine on the abundance of nmt2 mRNA is exerted at the level of transcription initiation.
The cloned nmt2 promoter contains the thiamine-responsive element ( s )
To determine whether the cloned fragment con- taining the nmt2 gene encompassed sufficient of the upstream promoter sequence to elicit the normal response to thiamine, the following experiment was performed. A 600 bp fragment (PstI-BstXI) was deleted from the nmt2 coding region and the resulting truncated HindIII-EcoRV fragment of approximately 1 kb was cloned into the SmaI site of the Escherichia colilS. pombe shuttle vector, pIRT2 (Hindley et al., 1987). This construct was then used to transform the leu1 -32 strain of fission yeast. Leu+ transformants were cultured in mini- mal medium either with or without thiamine, and RNA was extracted from each culture for North- ern blotting. The results are shown in Figure 5. Both the chromosomally-derived 1.4 kb mRNA and the shorter plasmid-derived transcript are highly expressed in the absence of thiamine, and
SECOND GENE FOR THIAMINE BIOSYNTHESIS
A 1079
B Time 0 1 2 3 5 z
nmt2 111,
------- 0 4 8 10 12 1416 hr
Figure 3. Northern blotting analysis showing the kinetics of repression and induction of nmt2. (A) Wild-type 972h - cells were grown to early log phase (lo6 cells/ml) in minimal medium and at To the culture was supplemented with 2 pM-thiamine. At the times indicated, approximately 10' cells were harvested for RNA extraction. 10 pg total RNA from each sample was used for Northern blotting, and the nmt2 specific transcripts were identified using the radiolabelled 1.5 kb HindIII-EcoRV fragment as hybridization probe. (B) Wild-type 972h- cells were grown to early log phase (lo6 cells/ml) in minimal medium containing 2 pM-thiamine and at To the cells were washed and resuspended at lo6 cells/ml in minimal medium. Aliquots of approximately 10' cells were harvested for RNA extraction at the times indicated. After 10 h, as the cell culture approached stationary phase, the cell density was reduced by dilution into fresh pre-warmed minimal medium and growth allowed to continue. RNA was prepared from each sample and analysed by Northern blotting as described in (A) above. Times are indicated in h.
both are fully repressed in medium containing thiamine. We conclude that the entire thiamine- responsive region of the nmt2 promoter is con- tained within the cloned 230 bp upstream of the nmt2 coding region.
The nmtl and nmt2 promoters contain a common sequence surrounding the transcription start site
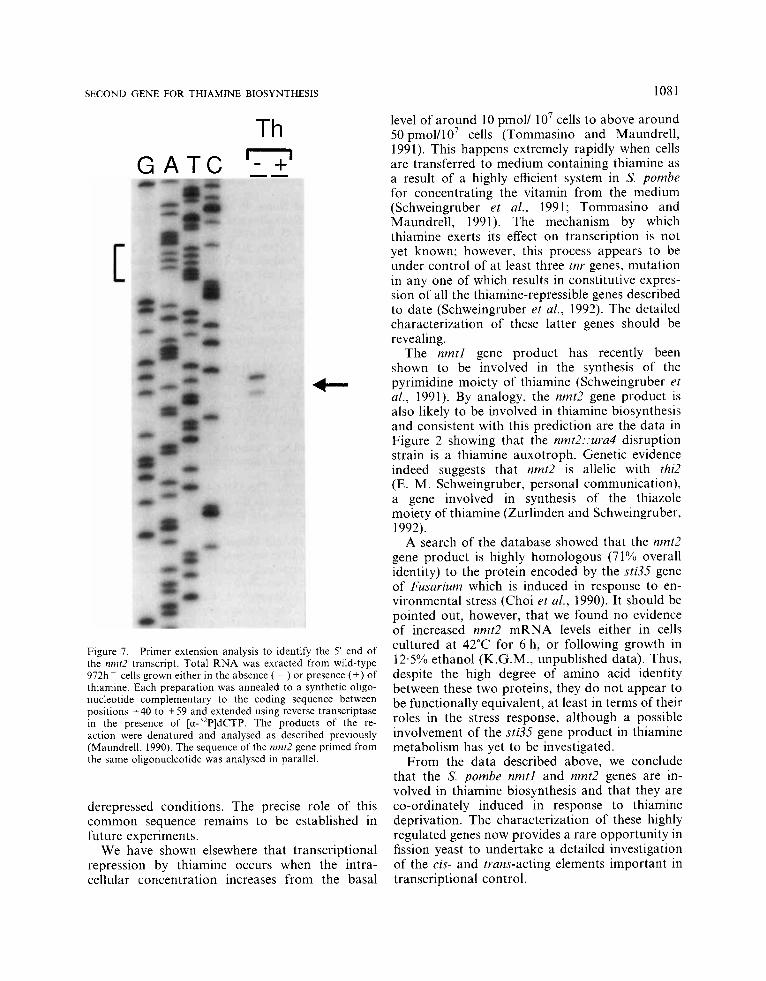
In view of the similarity in the pattern of tran- scription between the two thiamine-repressible genes, we searched their promoters for common sequence motifs. As shown in Figure 6, both promoters contain an identical 11 bp sequence element located downstream of a perfect canonical TATA element TATATAAA. In the case of nmtl this sequence spans the transcription start site determined previously (Maundrell, 1990). In order to determine the start of transcription of nmt2, we performed the primer extension experiments
shown in Figure 7. An oligonucleotide comple- mentary to nucleotides +40-+59 of the nmt2 coding region was hybridized to total RNA ex- tracted from cells grown either with or without thiamine and extended with reverse transcriptase as described previously (Grimm et al., 1988). The products of the reaction were electrophoresed ad- jacent to the sequence of nmt2 primed using the same oligonucleotide. The major radiolabelled band indicates a transcription start site on an A residue located 24 bp downstream of the TATA box and within the sequence element common to the two thiamine-repressible genes (Figure 6). In addition, a slightly shorter reaction product can also be seen which may indicate an alternative transcription start site 3 nucleotides further down- stream within the common element (see Figure 6). However, it is not clear from these data whether this represents a minor transcript of the nmt2 gene or an incomplete product of reverse transcription.
1080
+Th -Th
pUC119
u ra4
cycl
nmtl
A. G . 0. MANETTI ET AL.
- +
WT - A -
nmt2
Figure 4. Nuclear ‘run-on’ analysis to study the effect of thiamine on transcription initiation of nmtZ. Wild-type 972h ~
cells were grown in either minimal medium ( - Th) or minimal medium supplemented with 2 pM-thiamine (+Th) and at mid- log phase approximately 2 x lo7 cells from each culture were harvested. The cells were permeabilized, and nascent transcripts were labelled with [a-’*P]UTP for 5 min. Radioactive R N A was prepared from both samples and 5 x 10‘ cpm was hybridized to identical filters containing 5 pg per slot of each of the DNAs indicated. The procedures for RNA labelling, extraction and hybridization were exactly as described previously (Maundrell, 1990).
DISCUSSION We have studied the transcriptional regulation of the thiamine-repressible gene nmt2. Like nmtl, nmt2 is among the most highly transcribed genes when cells are cultured in minimal medium and the transcript completely disappears in the presence of exogenous thiamine. Nuclear ‘run-on’ experiments demonstrate that control is at the level of tran- scription initiation in both cases, and the kinetics of induction and repression indicate that the two genes are co-ordinately regulated. In the case of nmt2 all the promoter elements necessary for thiamine-mediated control of transcription are contained in the cloned 230 bp region upstream of the coding region (Figure 5) . Similarly, for nmtl we have found that 5’ deletions into the promoter
-103
Figure 5. Northern blot analysis to assess the activity of the cloned nmt2 promoter. The 0.6 kb fragment of the nmtl? coding sequence between the PstI and BstXI restriction sites was excised and the ends were filled in and religated. The resulting 1 kb HindIII-EcoRV fragment containing the deleted nmr2 gene was then cloned into the SmaI site of‘ a S. pomhe Leu’, ars’ shuttle vector essentially the same as pIRT2 described previ- ously (Hindley et al., 1987). leul-32 cells were transformed with this construct and grown to mid-log phase either in the absence ( - ) or presence (+) of thiamine. R N A was extracted from each culture and the nmt2-specific transcripts were identified by Northern blotting as described in the legend to Figure 3A. The chromosomally-derived (WT) and plasmid-derived (A) transcripts are indicated.
as far as position ~ 243 upstream of the initiator ATG leave the thiamine-responsive element( s) in- tact (M.R., and K.G.M., unpublished data). Com- parison of these two minimal promoter sequences has revealed a perfectly conserved 11 bp element at an equivalent location downstream of the TATA box and spanning the transcription start sites of the two genes (see Figure 6). This element is not found in any other S. pombe promoter sequenced to date. It may be involved in thiamine-mediated repression, or alternatively it may act to enhance the efficiency of transcription of these genes under
* nmt 1 TATATAAAGGAAGAGGA-ATCCTGGCATATCATCAATTGAATA
nmt 2 TATATAAAGGGGGCTCATAGCATGTTTCAGCATCAATTGAACA I I I I I I I I I I I I I I I I I I I
- 8 2 * Figure 6 . A DNA sequence element common to the nmtl and nmt2 promoters. The sequence of the nmtl and nmt2 promoters immediately downstream of the respective TATA elements is shown, and the 1 1 bp conserved element is indicated. Asterisks indicate the transcription start site in each case. Nucleotide positions refer to the distance between the first base of the TATA box and the start of the open reading frame.
SECOND GENE FOR THIAMINE BIOSYNTHESIS
Th G A T C --
1081
level of around 10 pmoll lo7 cells to above around 50 pmol/107 cells (Tommasino and Maundrell, 1991). This happens extremely rapidly when cells are transferred to medium containing thiamine as a result of a highly efficient system in S. pombe for concentrating the vitamin from the medium (Schweingruber et al., 1991; Tommasino and Maundrell, 1991). The mechanism by which thiamine exerts its effect on transcription is not yet known; however, this process appears to be under control of at least three tnr genes, mutation in any one of which results in constitutive expres- sion of all the thiamine-repressible genes described to date (Schweingruber et al., 1992). The detailed characterization of these latter genes should be revealing.
The nmtl gene product has recently been shown to be involved in the synthesis of the pyrimidine moiety of thiamine (Schweingruber et al., 1991). By analogy, the nmt2 gene product is also likely to be involved in thiamine biosynthesis and consistent with this prediction are the data in Figure 2 showing that the nmt2::ura4 disruption strain is a thiamine auxotroph. Genetic evidence indeed suggests that nmt2 is allelic with thi2 (E. M. Schweingruber, personal communication), a gene involved in synthesis of the thiazole moiety of thiamine (Zurlinden and Schweingruber, 1992).
A search of the database showed that the nmt2 gene product is highly homologous (7 1 O/U overall identity) to the protein encoded by the sti35 gene of Fusarium which is induced in response to en- vironmental stress (Choi et al., 1990). It should be pointed out, however, that we found no evidence of increased nmt2 mRNA levels either in cells
4-
cultured at 42°C for 6 h, or following growth in 12'5% ethanol (K.G.M., unpublished data). despite the high degree of amino acid identity between these two proteins, they do not appear to be functionally equivalent, at least in terms of their roles in the Stress response, although a possible involvement of the sti35 gene product in thiamine metabolism has yet to be investigated.
From the data described above, we conclude that the S. pombe nmtl and nmt2 genes are in- volved in thiamine biosynthesis and that they are co-ordinately induced in response to thiamine deprivation. The characterization of these highly regulated genes now provides a rare opportunity in fission yeast to undertake a detailed investigation of the cis- and trans-acting elements important in transcriptional control.
Figure 7. Primer extension analysis to identify the 5' end of the nmt2 transcript. Total RNA was exracted from wild-type 97231 ~ cells grown either in the absence ( - ) or presence (+) of thiamine. Each preparation was annealed to a synthetic oligo- nucleotide complementary to the coding sequence between positions +40 to +59 and extended using reverse transcriptase in the presence of [ a - 3 2 P ] d ~ ~ p . The products of the re- action were denatured and analysed as described previously (Maundrell, 1990). The sequence of the nmt2 gene primed from the same oligonucleotide was analysed in parallel.
derepressed conditions. The precise role of this common sequence remains to be established in future experiments.
We have shown elsewhere that transcriptional repression by thiamine occurs when the intra- cellular concentration increases from the basal

1082 A. G . 0. MANETTI ET AL.
ACKNOWLEDGEMENTS We would like to thank Dr Rino Rappuoli of the IRIS Research Centre for support during the in- itial phase of this work, Dr M. E. Schweingruber for communicating unpublished data, Elisabeth Schmid for expert technical assistance and Magali Leeman-Husler and Christopher Hebert for art- work and photography.
REFERENCES Choi, G. H., Marek, E. T., Schardl, C. L., Richey,
M. G., Chang, S. and Smith, D. A. (1990). sti35, a stress-responsive gene in Fusarium spp. J. Bacteriol. 172,45224528.
Devereux, J., Haeberli, P. and Smithies, 0. (1984). A comprehensive set of sequence analysis programs for the Vax. Nucleic Acids Res. 12, 387-395.
Grimm, C., Kohli, J., Murray, J. and Maundrell, K. (1988). Genetic engineering of Schizosaccharomyces pombe: a system for gene disruption and replacement using the ural gene as a selectable marker. Mol. Gen. Genet. 215, 81-86.
Guarente, L. and Bermingham-McDonogh, 0. (1992). Conservation and evolution of transcriptional mech- anisms in eukaryotes. Trends Genet. 8, 27-32.
Gutz, H., Heslot, H., Leupold, U. and Loprienno, N. (1974). In King, R. C. (Ed.), Schizosaccharomyces pombe Handbook of Genetics. Plenum Publishing Co., New York, pp. 1395-1446.
Henikoff, S. (1984). Unidirectional digestion with exo- nuclease 111 creates targeted breakpoints for DNA sequencing. Gene 28, 351-359.
Hindley, J., Phear, G., Stein, M. and Beach, D. (1987). sucl+ encodes a predicted 13-kilodalton protein that is essential for cell viability and is directly involved in the division cycle of Schizosaccaromyces pombe. Mol. Cell. Biol. 7, 504-51 1.
Jones, R., H., Moreno, S., Nurse, P. and Jones, N. C. (1988). Expression of the SV40 promoter in fission yeast: identification and characterization of an APO-1 like factor. Cell 53, 569-667.
Maundrell, K. (1990). nmtl of fission yeast: a highly expressed gene completely repressed by thiamine. J. Biol. Chem. 265, 10857-10864.
Moreno, S., Klar, A. and Nurse, P. (1991). Molecular genetic analysis of fission yeast, Schizosaccharomyces pombe. Methods Enzymol. 194, 795-823.
Prabhala, G., Rosenberg, G. H. and Kaufer, N. F. (1992). Architectural features of pre-mRNA introns in the fission yeast Schizosaccharomyces pombe. Yeast 8,
Russell, P. and Hall, B. (1982). Structures of Schizosac- charomyces pombe cytochrome c gene, Mol. Cell. Biol. 2, 106116.
Russell, P. and Nurse, P. (1986). Schizosaccharomyces pombe and Saccharomyces cerevisiae: a look at yeasts divided. Cell 45, 781-782.
Sambrook, J., Fritsch, E. F. and Maniatis, T. (1989). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor University Press, Cold Spring Harbor, New York.
Schweingruber, A. M., Dlugonski, J., Edenharter, E. and Schweingruber, M. E. (1991). Thiamine in Schizosaccharomyces pombe: dephosphorylation, intra- cellular pool, biosynthesis and transport. Curr. Genet.
Schweingruber, A.-M., Fankhauser, H., Dlugonski, J., Steinmann-Loss, C. and Schweingruber, M. E. (1992). Isolation and characterization of regulatory mu- tants from Schizosaccharomyces pombe involved in thiamine-regulated gene expression. Genetics 130, 4 4 5 4 9 .
Struhl, K. (1987a). Promoters, activator proteins, and the mechanism of transcriptional activation in yeast. Cell 49, 295-297.
Struhl, K. (1987b). The DNA-binding domains of the jun oncoprotein and the yeast GCN4 transcriptional activator protein are functionally homologous. Cell
Tommasino, M. and Maundrell, K. (1991). Uptake of thiamine by Schizosaccharomyces pombe and its effect as a transcriptional regulator of thiamine-sensitive genes. Curr. Genet. 20, 63-66.
Wiederrecht, G., Shuey, D. J., Kibbe, W. A. and Parker, C. S. (1987). The Saccharomyces and Drosophila heat shock transcription factors are identical in size and DNA-binding properties. Cell 48, 507-51 5.
Zurlinden, A. and Schweingruber, M. E. (1992). Cloning and regulation of Schizosaccharomyces pombe thi2, a gene involved in thiamine biosynthesis. Gene 117, 141-143.
17 1-1 82.
19, 249-254.
50, 841-846.





























