Embed Size (px)
Citation preview

Cancer Metastasis Reviews 3, 5-24 (1984) © 1984, Martinus Nijhoff Publishers, Boston. Printed in the Netherlands
Nonmetastatic tumor cells acquire metastatic properties following somatic hybridization with normal cells
P. De Baetselier, 1 E. Roos , 2 L. Brys, 1 L. Remels, 1 M. Gobert, 1 D. Dekegel, ~ S. Segal 3 and M. Feldman 3 1 Dienst Algemene Biologie, Instituut voor Moleculaire Biologie, Vrije Universiteit Brussel, Belgium; 2 Division of Cell Biology, The Netherlands Cancer Institute, Amsterdam, The Netherlands; and 3 Department of Cell Biology, The Weizmann Institute of Science, Rehovot, Israel
Keywords: somatic hybridization, metastasis, tumor progression
Summary
Somatic cell hybridization between nonmetastatic tumor cells and normal cells of the lymphoreticular system results in hybrid cells manifesting metastatic properties of defined target organ specificity. Thus, fusion of the nonmetastatic BALB/c originated NSI plasmacytoma with C57BL B lymphocytes resulted in hybridomas, each of which were metastatic. Of 10 hybridomas, 7 generated metastases in the spleen and liver, whereas 3 generated liver metastases. The generation of liver metastases by hybridomas which homed to both spleen and liver, but not by those which homed to the liver only, was controlled by the spleen. The acquisition of metastatic properties via somatic cell fusion seems to represent a general principle, in which the normal partner determines the target organ specificity for the metastatic growth. Thus, fusion of SP2/O myeloma cells with syngeneic B lymphocytes also resulted in a hybrid cell metastasizing to the spleen and liver, yet a somatic hybrid between NSI and a macrophage or dendritic-like cell metastasized to the lung. Cell surface molecules encoded by the genome of the normal partner was demonstrated to control the target organ specificity: antibodies against MHC-encoded antigens of the normal B cell partner prevented the generation of metastases by hybridomas metastasizing to the spleen and liver, but not by those metastasizing to the liver only. This is in accordance with the function of MHC molecules on lymphocytes in controlling their homing to lymphoid organs. Hybridomas of T cell lymphomas also manifested metastatic properties. Analysis of the cell surface Thy-1 antigens of a hybridoma (DCH10), produced via somatic fusion between BW5145 lymphoma and a putative macrophage cell indicated that cells of liver metastases (DCH10-Li) generated by the hybrid cells might have undergone further somatic cell fusion in vivo with host (T?) cells. These cells have acquired new metastatic properties, generating metastases in spleen, liver and kidneys. In fact, even the inoculation of the parental BW lymphoma cells resulted in a case of liver metastasis (BW-Li). Such BW-Li cells, upon reinoculation, also generated metastases in the spleen, liver and kidneys. Analysis of the Thyl phenotype indicated that BW-Li cells may also have undergone somatic cell fusion in vivo with host (T?) cells, resulting in the acquisition of metastatic properties. The pattern of cell-cell interactions (adhesion, infiltration) with liver cell monolayers of BW-Li cells and of DCH10-Li (T-cell lymphomas) was identical, and differed from cells of liver metastases of the myeloma-B cell hybridomas which might be based on responses to liver growth signals. Accordingly, the morphology of liver metastases generated by the two categories of hybridomas was different. It appears therefore, that (a) the acquisition of metastatic properties
Address for reprints: Dr. P. Baetselier, Dienst Algemene Biologie, Instituut voor Moleculaire Biologie, Vrije Universiteit Brussel, Paardenstraat 65, 1640 St-Genesius-Rode, Belgium

following somatic cell fusion with normal lymphoreticular cells is of a general significance; (b) somatic cell fusion provides an experimental system for the analysis of molecular properties determining the acquisition of metastatic capability; and (c) it may also represent a mechanism for tumor progression in vivo.
1. Introduction
The question of whether and how properties of tumor cells are altered following somatic hybridiza- tion between neoplastic and normal cells has at- tracted interest ever since the discovery of the tech- nique of somatic cell fusion. Early studies of somatic hybrids derived from fusion of cells of high malignant lines with cells of low malignant lines suggested that the high malignancy of the parental neoplastic cells was the dominant property (1, 2). Subsequent studies demonstrated, however, that the malignant characteristics could behave like a recessive trait in somatic cell hybrids (3, 4). In such hybrids, i tappeared that the neoplastic property was the result of a 'deletion' which was comple- mented by 'healthy' chromosomes of the normal partner.
The relevance of these observations to the in vivo processes of tumor progression remained an open question. Yet, following the initial studies on the outcome of in vitro somatic cell fusion, reports appeared suggesting that tumor cells grown in vivo and in vitro may undergo somatic fusion with the host's normal cells (5-10). In most of these cases, however, the properties acquired by the tumor as a function of somatic hybridization were not studied. Only Goldberger suggested that the generation of metastases by human malignant cells grown in hamsters was associated with somatic cell fusion with hamster cells (11). We suggested that such properties may include the acquisition of metasta- tic competence by nonmetastatic tumor cells, in view of our observations of the phenotypic changes which took place in clones of the T10 sarcoma (12, 13). This tumor, induced by 3-methylcholanthrene in a (C57BL/6J × C3Heb)F1 mouse, manifested two phenotypic patterns when grown in a syn- geneic F1 recipient. Cells of the local tumor ex- pressed at their surfaces predominantly the H-2 b haplotype, whereas cells form lung metastases ex- pressed both the H-2 b and the H-2 k parental
haplotypes. When the local tumor cells were cloned in culture, the majority of the clones ex- pressed the H-2 ~ haplotype and only a minority of clones expressed both parental haplotypes. When tested in vivo, only the latter clones generated lung metastases (14, 15). Clones which phenotypically were H-2 b+ but H-2 k- grew locally, but did not generate metastases. We subsequently tested whether, following serial transplantations in vivo of such clones in syngeneic F1 hybrid mice, altera- tions in the phenotypic properties of such cells would take place. We found that in every single case, a H-2 b+, H-2 k- clone, when transferred se- rially in (H-2 b × H-2k)F1 recipients eventually ex- pressed the two haplotypes (12, 13). Concomitantly with the acquisition of the 'missing' haplotype, the cells acquired metastatic competence. A 'con- ventional' explanation for this alteration in the properties of the cloned tumor cell populations would be a de-repression of a repressed H-2 k haplotype. Yet, the very fact that this change has taken place only following in vivo growth in (H-2b× H-2k)F1 mice and never following pro- longed growth in culture (for more than two years), suggested to us the possibility that the expression of the H-2 k genes may have derived from somatic fusion between the tumor's H-2 k- cells and the host's H-2 k+ somatic cells. But could such somatic hybridization confer metastatic properties on non- metastic tumor cells? To test whether, in principle, such cell fusions may result in metastatic capacity, we chose to study a model system in which somatic hybridization could readily be achieved in culture and in which the normal non-neoplastic partner was a 'circulating' cell, and thus possessed some of the disseminating properties of a metastatic cell. We initially investigated the outcome of hybridiza- tion of nonmetastatic plasmacytoma cells with nor- mal B lymphocytes.

2. Plasmacytoma cells acquire metastatic proper- ties following fusion with spleen B-lymphocytes
In our initial studies (16) we used the BALB/c originated P3-NSI/1-Ag 4-1 (NSI) plasmacytoma cell line, obtained from Dr. C. Milstein, MRC, Cambridge, England. The normal parental cells were B lymphocytes from C57BL/6 female mice which had been immunized either by DNP-KLH or by Micrococcus luteus. Following a booster immu- nization against these antigens, the spleen cells were inoculated to syngeneic recipients which had been exposed to 700 r total body irradiation. The latter received an additional antigenic boost prior to the removal for fusion of their spleen cells 4-5 days later. Fusion of these cells with the NSI plas- macytoma was carried out via polyethylene glycol. Hybrid cells that secreted either anti-DNP anti- bodies (determined by Farr radioimmunoassay), or anti-micrococcus (determined by bacterial ag- glutination), were cloned by limiting dilutions.
Following transplantation to syngeneic F1 hybrid mice, the hybrid cells grew either as ascites or as solid tumors. The hybrid nature of the tumor cells thus produced was evident from the secretion of monoclonal immunoglobulin characteristic of the parental B lymphocyte. This could be determined by electrophoretic analysis of either the sera or the ascites fluid of the tumor-bearing mice. Seven of the 10 hybridornas tested secreted antibodies spe- cific for the antigen against which the donors of the normal B cells had been immunized. Table 1 pres- ents the main properties of the hybridomas used in the following experiments.
All hybridomas produced local tumors following subcutaneous (s.c.) transplantation of 5 x 106 cells into (C57BL/6 x BALB/c)F1 recipients. The tu- mor-bearing animals were either autopsied at death or killed at the terminal stages of tumor growth and the various organs were assayed for visible metastases. The result was that, whereas the NSI plasmacytoma grew locally but did not gener- ate metastases, the hybridomas were found to do so. Thus, Table 2 demonstrates that three hybridomas (HY1-Hy3) generated metastatic nod- ules in the spleen and liver, whereas one (HY8) produced only liver metastases. In fact, the
Table 1. Characteristics of various NSl-derived hybridomas.
Hybridoma 1 Antibody Heavy Antigen specificity secretion chain
class
HYI + IgG1 ? HY2 + IgM ? HY3 2 _ _ HY4 + IgM DNP HY5 + IgG~ DNP HY6 + IgG2 Micrococci HY7 + IgM Micrococci HY8 + IgM Micrococci HY9 + IgM Micrococci HYI0 + IgM Micrococci
i HY1-HY3 and HY6-HY10 were derived from fusion of cells from micrococci-immunized mice and NSI plasmacytoma cells. HY4 and HY5 were derived from fusion of cells from DNP- KLH-immunized mice and NSI plasmacytoma cells. 2 A nonsecretor hybridoma.
hybridomas tested could be classified into two dis- tinct categories: hybridomas which generated met- astases both in the spleen and the liver and hybridomas which generated metastases only in the liver. We subsequently tested (a) whether hybrid cells from spleen or liver metastases of HY2 and HY8 were selected for metastatic competence, and (b) whether HY2 spleen metastases, as distinct from liver metastases, reflect selection for target organ specificities. Tumor cells from spleen or liver nodules of the primary host were trypsinized and inoculated s.c. into secondary F1 mice. The result was that whereas in the primary hosts 25% of the animals manifested metastases (Table 2, exp. 1), in the secondary hosts, grafted with cells from spleen or liver nodules, 100% of the recipients manifested spontaneous metastases (Table 2, exp. 2). Thus, following one transfer generation, selection for a high metastatic capacity was achieved. Cells from spleen nodules of HY2, when regrafted s.c., pro- duced spontaneous metastases both in the spleen and in the liver, and the same was true for liver metastases. Thus, cells from spleen and liver meta- stases do not reflect the pre-existence of two sub- populations with distinct target organ specificities. Yet, it appears that tumor cells from spleen meta- stases were more potent in generating spleen meta-
8
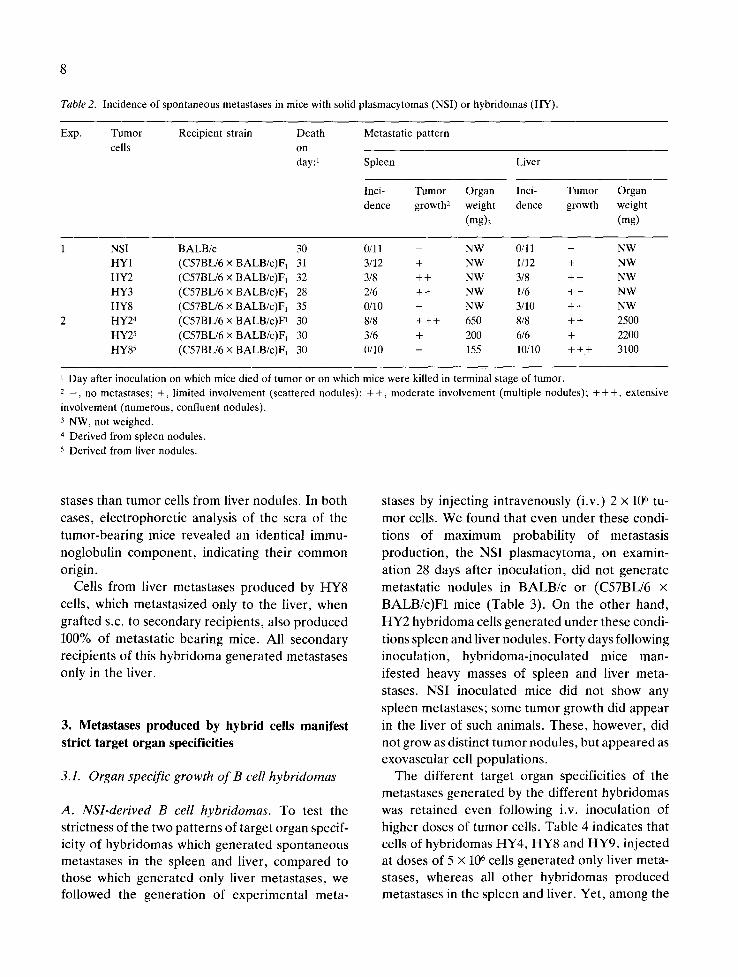
Table 2. Incidence of spontaneous metastases in mice with solid plasmacytomas (NSI) or hybridomas (HY).
Exp. Tumor Recipient strain Death Metastatic pattern
cells on day:~ Spleen Liver
Inci- Tumor Organ Inci- Tumor Organ
dence growth: weight dence growth weight
(mg)3 (mg)
1 NSI BALB/c 30 0/11 - NW 0/11
HY1 (C57BL/6 × BALB/c)F 1 31 3/12 + NW 1/12
HY2 (C57BL/6 x BALB/c)F1 32 3/8 + + NW 3/8
HY3 (C57BL/6 × BALB/c)F~ 28 2/6 + + NW 1/6
HY8 (C57BL/6 × BALB/c)F 1 35 0/10 - NW 3/10
2 HY24 (C57BL/6 × BALB/c)F~ 30 8/8 + + + 650 8/8
HY25 (C57BL/6 × BALB/c)F1 30 3/6 + 200 6/6
HY85 (C57BL/6 × BALB/c)F~ 30 0/10 - 155 10/10
- NW
+ NW
+ + NW
+ + NW
+ + NW
+ + 2500
+ 2200
+ + + 3100
1 Day after inoculation on which mice died of tumor or on which mice were killed in terminal stage of tumor. 2 _ , no metastases; +, limited involvement (scattered nodules): + +, moderate involvement (multiple nodules); + + +, extensive
involvement (numerous, confluent nodules). 3 NW, not weighed.
4 Derived from spleen nodules.
5 Derived from liver nodules.
stases than tumor cells from liver nodules. In both cases, electrophoretic analysis of the sera of the tumor-bearing mice revealed an identical immu- noglobulin component, indicating their common origin.
Cells from liver metastases produced by HY8 cells, which metastasized only to the liver, when grafted s.c. to secondary recipients, also produced 100% of metastatic bearing mice. All secondary recipients of this hybridoma generated metastases only in the liver.
3. Metastases produced by hybrid cells manifest strict target organ specificities
3.1. Organ specific growth of B cell hybridomas
A. NSl-derived B cell hybridomas. To test the strictness of the two patterns of target organ specif- icity of hybridomas which generated spontaneous metastases in the spleen and liver, compared to those which generated only liver metastases, we followed the generation of experimental meta-
stases by injecting intravenously (i.v.) 2 × 106 tu- mor cells. We found that even under these condi- tions of maximum probability of metastasis production, the NSI plasmacytoma, on examin- ation 28 days after inoculation, did not generate metastatic nodules in BALB/c or (C57BL/6 × BALB/c)F1 mice (Table 3). On the other hand, HY2 hybridoma cells generated under these condi- tions spleen and liver nodules. Forty days following inoculation, hybridoma-inoculated mice man- ifested heavy masses of spleen and liver meta- stases. NSI inoculated mice did not show any spleen metastases; some tumor growth did appear in the liver of such animals. These, however, did not grow as distinct tumor nodules, but appeared as exovascular cell populations.
The different target organ specificities of the metastases generated by the different hybridomas was retained even following i.v. inoculation of higher doses of tumor cells. Table 4 indicates that cells of hybridomas HY4, HY8 and HY9, injected at doses of 5 × 106 cells generated only liver meta- stases, whereas all other hybridomas produced metastases in the spleen and liver. Yet, among the
9
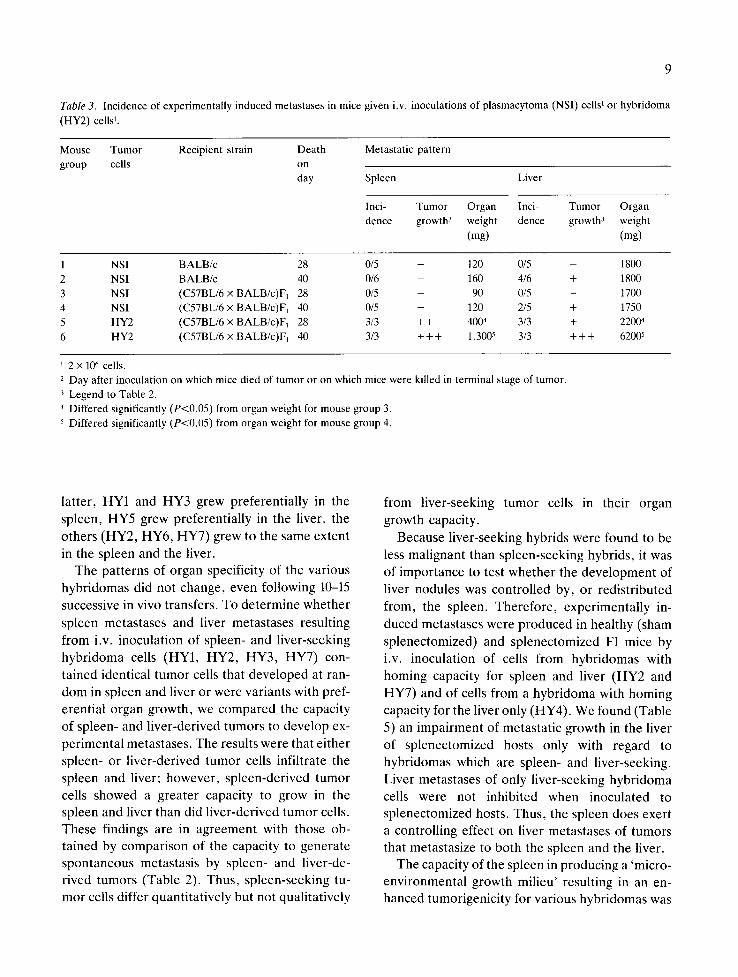
Table 3. Incidence of experimentally induced metastases in mice given i.v. inoculations of plasmacytoma (NSI) cells 1 or hybridoma
(HY2) cells 1.
Mouse Tumor Recipient strain Death Metastatic pattern
group cells on day Spleen Liver
Inci- Tumor Organ Inci- Tumor Organ
dence growth 3 weight dence growth 3 weight
(mg) (mg)
1 NSI BALB/c 28 0/5 - 120 0/5 - 1800
2 NSI BALB/c 40 0/6 - 160 4/6 + 1800
3 NSI (C57BL/6 x BALB/c)F1 28 0/5 - 90 0/5 - 1700
4 NSI (C57BL/6 x BALB/c)F~ 40 0/5 - 120 2/5 + 1750
5 HY2 (C57BL/6 x BALB/c)F~ 28 3/3 + + 4004 3/3 + 22004
6 HY2 (C57BL/6 x BALB/c)F~ 40 3/3 + + + 1,3005 3/3 + + + 6200 s
2 x 106 cells.
2 Day after inoculation on which mice died of tumor or on which mice were killed in terminal stage of tumor.
3 Legend to Table 2.
4 Differed significantly (P<0.05) from organ weight for mouse group 3.
5 Differed significantly (P<0.05) from organ weight for mouse group 4.
latter, HY1 and HY3 grew preferentially in the spleen, HY5 grew preferentially in the liver, the others (HY2, HY6, HY7) grew to the same extent in the spleen and the liver.
The patterns of organ specificity of the various hybridomas did not change, even following 10-15 successive in vivo transfers. To determine whether spleen metastases and liver metastases resulting from i.v. inoculation of spleen- and liver-seeking hybridoma cells (HY1, HY2, HY3, HY7) con- tained identical tumor cells that developed at ran- dom in spleen and liver or were variants with pref- erential organ growth, we compared the capacity of spleen- and liver-derived tumors to develop ex- perimental metastases. The results were that either spleen- or liver-derived tumor cells infiltrate the spleen and liver; however, spleen-derived tumor cells showed a greater capacity to grow in the spleen and liver than did liver-derived tumor cells. These findings are in agreement with those ob- tained by comparison of the capacity to generate spontaneous metastasis by spleen- and liver-de- rived tumors (Table 2). Thus, spleen-seeking tu- mor cells differ quantitatively but not qualitatively
from liver-seeking tumor cells in their organ growth capacity.
Because liver-seeking hybrids were found to be less malignant than spleen-seeking hybrids, it was of importance to test whether the development of liver nodules was controlled by, or redistributed from, the spleen. Therefore, experimentally in- duced metastases were produced in healthy (sham splenectomized) and splenectomized F1 mice by i.v. inoculation of cells from hybridomas with homing capacity for spleen and liver (HY2 and HY7) and of cells from a hybridoma with homing capacity for the liver only (HY4). We found (Table 5) an impairment of metastatic growth in the liver of splenectomized hosts only with regard to hybridomas which are spleen- and liver-seeking. Liver metastases of only liver-seeking hybridoma cells were not inhibited when inoculated to splenectomized hosts. Thus, the spleen does exert a controlling effect on liver metastases of tumors that metastasize to both the spleen and the liver.
The capacity of the spleen in producing a 'micro- environmental growth milieu' resulting in an en- hanced tumorigenicity for various hybridomas was
10
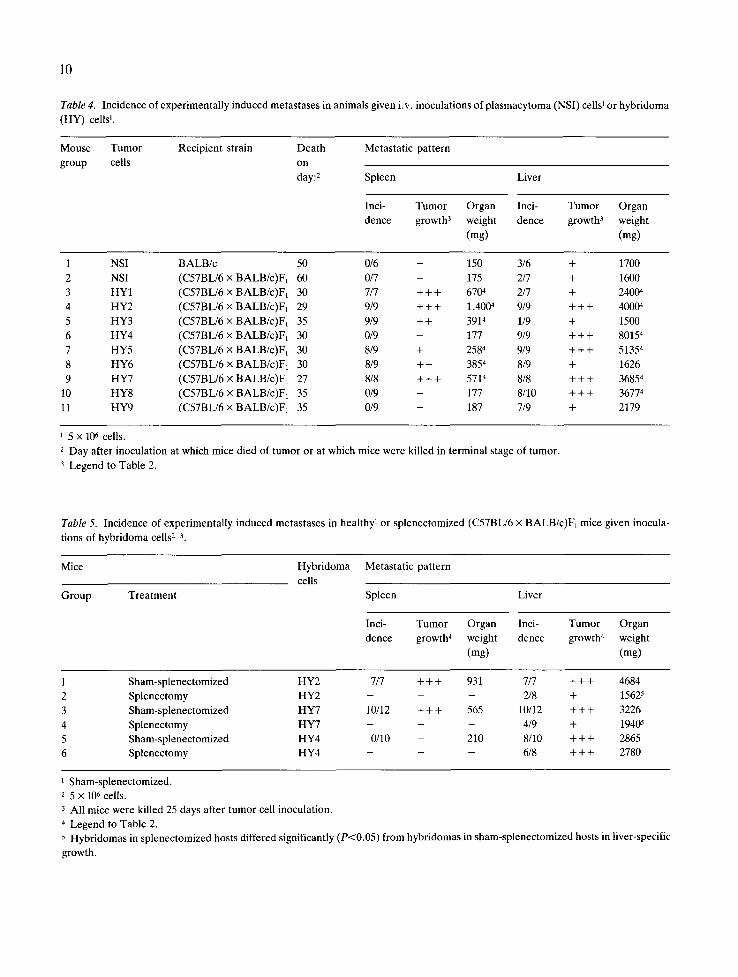
Table 4. Incidence of experimentally induced metastases in animals given i.v. inoculations of plasmacytoma (NSI) cells ~ or hybridoma
(HY) cells 1.
Mouse Tumor Recipient strain Death Metastatic pattern
group cells on
day: 2 Spleen Liver
Inci- Tumor Organ Inci- Tumor Organ
dence growth 3 weight dence growth 3 weight
(mg) (mg)
1 NSI BALB/c 50 0/6 - 150 3/6 + 1700
2 NSI (C57BL/6 x BALB/c)F~ 60 0/7 - 175 2/7 + 1600
3 HY1 (C57BL/6 x BALB/c)F~ 30 7/7 + + + 6704 2/7 + 24004
4 HY2 (C57BL/6 x BALB/c)F~ 29 9/9 + + + 1,4004 9/9 + + + 40004
5 HY3 (C57BL/6 × BALB/c)F~ 35 9/9 + + 3914 1/9 + 1500
6 HY4 (C57BL/6 x BALB/c)F1 30 0/9 - 177 9/9 + + + 80154
7 HY5 (C57BL/6 x BALB/c)F1 30 8/9 + 2584 9/9 + + + 51354
8 HY6 (C57BL/6 x BALB/c)Fx 30 8/9 + + 3854 8/9 + 1626
9 HY7 (C57BL/6 x BALB/c)F1 27 8/8 + + + 5714 8/8 + + + 36854
10 HY8 (C57BL/6 x BALB/c)FI 35 0/9 - 177 8/10 + + + 36774
11 HY9 (C57BL/6 x BALB/c)F1 35 0/9 - 187 7/9 + 2179
i 5 x 106 cells.
2 Day after inoculation at which mice died of tumor or at which mice were killed in terminal stage of tumor.
3 Legend to Table 2.
Table 5. Incidence of experimentally induced metastases in healthy 1 or splenectomized (C57BL/6 x BALB/c)F1 mice given inocula-
tions of hybridoma cells 2, 3.
Mice Hybridoma Metastatic pattern
cells
Group Treatment Spleen Liver
Inci- Tumor Organ Inci- Tumor Organ
dence growth 4 weight dence growth 4 weight
(mg) (mg)
1 Sham-splenectomized HY2 7/7 + + + 931 7/7 + + + 4684
2 Splenectomy HY2 - - - 2/8 + 15625
3 Sham-splenectomized HY7 10/12 + + + 565 10/12 + + + 3226
4 Splenectomy HY7 - - - 4/9 + 19405
5 Sham-splenectomized HY4 0/10 - 210 8/10 + + + 2865
6 Splenectomy HY4 - - - 6/8 + + + 2780
1 Sham-splenectomized. 2 5 x 106 cells.
3 All mice were killed 25 days after tumor cell inoculation.
4 Legend to Table 2. 5 Hybridomas in splenectomized hosts differed significantly (P<0.05) from hybridomas in sham-splenectomized hosts in liver-specific
growth.

reported recently by Witte and Ber (17). Similarly, the growth of a murine leukemia (BCL1) has been reported to be spleen-dependent, since early splenectomy could eradicate completely the pro- liferation of BCL1 cells in the peripheral blood (18). These results suggest that the spleen plays a growth-controlling role in tumor development, or that certain tumor cells may be induced in the spleen to undergo a differentiation event necessary for their spread. In fact, recent studies in our labo- ratory indicate that the microenvironment of the spleen induces maturation processes in B cells and macrophages (19, 20). Such processes may pro- mote the acquisition of properties that control the capacity of leukemias and B cell hybridomas to metastasize to other organs or to proliferate in the periphery.
B. SP2/O-derived B cell hybridomas. We sub- sequently aimed at studying whether a B cell uni- versal fuser other than the NSI, when similarly hybridized to B lymphocytes, would also generate hybridomas of metastatic competence and, if so, whether the target organ specificities would be similar to those observed following crossing of NSI cells and B lymphocytes. We selected the SP2/0 tumor cells, a BALB/c plasmacytoma type cell cur- rently used to generate B cell hybridomas, and fused it with B lymphocytes. However, since the NSI cells and the normal B cell partners in all previous experiments were of two distinct H-2 haplotypes, we now tested whether fusing two par-
11
ental cells of identical H-2 phenotype also resulted in the acquisition of metastatic properties.
BALB/c spleen cells from animals which had been immunized against the micrococcus antigens were fused with the SP2/0 plasmacytoma. Cells of one hybridoma (HYllHll) , thus produced which secreted anti-micrococcus antibodies of the IgM class, were selected for further study. HYl lHl l cells, when inoculated intravenously to syngeneic BALB/c recipients, generated multiple metastatic nodules in the liver and scattered nodules in the spleen (Table 6). The inoculation of cells of the SP2/0 plasmacytoma did not result in tumor growth in any of the recipient organs, when tested 36 days after inoculation. It thus appeared that the l lHl l hybridoma, produced by fusion of SP2/0 plasma- cytoma with a B cell, acquired metastatic proper- ties of similar organ specificities to that of the HYI-10 series, derived from the NSI plasma- cytoma. It should be stressed in this context that the l lHl l hybridoma secreted an antibody of iden- tical antigenic specificity (i.e., micrococcus) and heavy chain isotype as some of the NSI-derived hybridomas (HY7, HY8, HY9).
We subsequently tested whether the spleen and liver nodules represented distinct organ specific variants or whether these populations are random segregants. Hence, hybrid cells from spleen and liver of the primary recipients were re-inoculated to secondary hosts. The results (Table 7) indicate that cells from spleen metastases generated both spleen and liver nodules, as did tumor cells from
Table 6. Incidence of experimentally induced metastases in BALB/c mice given i.v. inoculations of p lasmacytoma (SP2/0) or hybridoma (HYl lH l l ) cells.
Tumor Days 1 Metastatic pat tern
Spleen Liver
Inci- Tumor Organ Inci- Tumor Organ
dence growth 2 weight dence growth 2 weight
(mg) (mg)
SP2/0 36 0/5 - 112 0/5 - 1380 H Y l l H l l 30 2/5 + 130 4/5 + + 1800
1 Day after inoculation on which mice were killed. 2 Legend to Table 2.
12
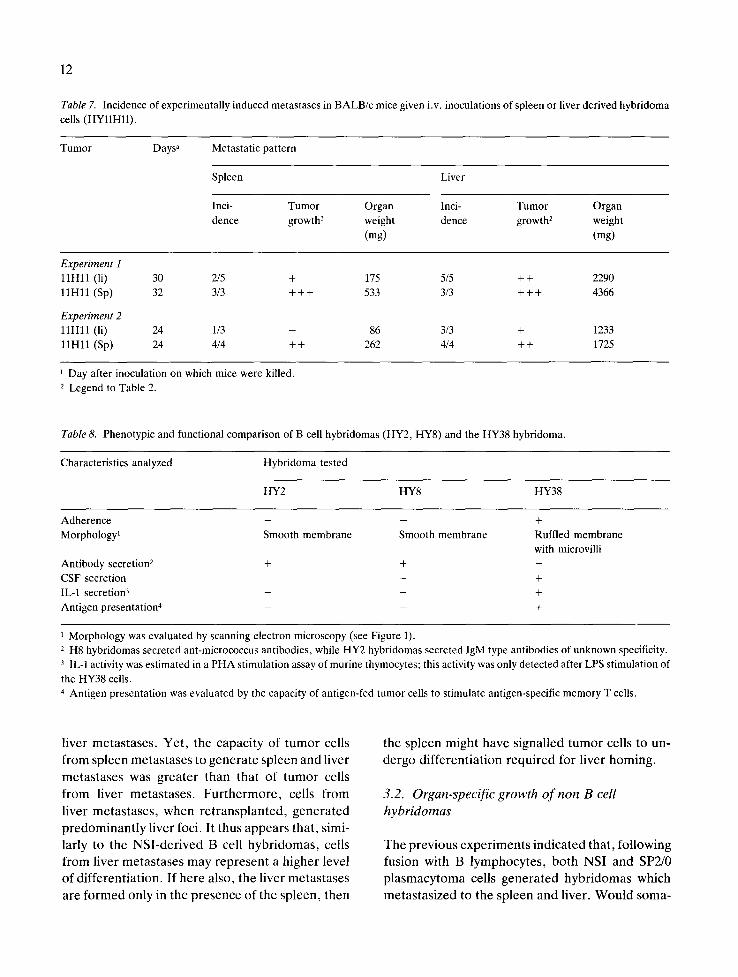
Table 7. Incidence of experimentally induced metastases in BALB/c mice given i.v. inoculations of spleen or liver derived hybridoma cells (HYllHll).
Tumor Days a Metastatic pattern
Spleen Liver
Inci- Tumor Organ Inci- Tumor Organ dence growth 2 weight dence growth 2 weight
(rag) (mg)
Experiment 1 11Hll (li) 30 2/5 + 175 5/5 + + 2290 11H11 (Sp) 32 3/3 + + + 533 3/3 + + + 4366
Experiment 2 11Hll (li) 24 1/3 - 86 3/3 + 1233 11Hll (Sp) 24 4/4 ++ 262 4/4 ++ 1725
Day after inoculation on which mice were killed. 2 Legend to Table 2.
Table 8. Phenotypic and functional comparison of B cell hybridomas (HY2, HY8) and the HY38 hybridoma.
Characteristics analyzed Hybridoma tested
HY2 HY8 HY38
Adherence - - + Morphology 1 Smooth membrane Smooth membrane Ruffled membrane
with microvilli Antibody secretion 2 + + - CSF secretion - - + IL-1 secretion 3 - - + Antigen presentation 4 - - +
1 Morphology was evaluated by scanning electron microscopy (see Figure 1). 2 H8 hybridomas secreted ant-micrococcus antibodies, while HY2 hybridomas secreted IgM type antibodies of unknown specificity. 3 IL-1 activity was estimated in a PHA stimulation assay of murine thymocytes; this activity was only detected after LPS stimulation of the HY38 cells. 4 Antigen presentation was evaluated by the capacity of antigen-fed tumor cells to stimulate antigen-specific memory T cells.
l ive r m e t a s t a s e s . Y e t , t h e c a p a c i t y o f t u m o r cel ls
f r o m s p l e e n m e t a s t a s e s to g e n e r a t e s p l e e n and l ive r
m e t a s t a s e s was g r e a t e r t h a n tha t o f t u m o r cel ls
f r o m l ive r m e t a s t a s e s . F u r t h e r m o r e , cel ls f r o m
l ive r m e t a s t a s e s , w h e n r e t r a n s p l a n t e d , g e n e r a t e d
p r e d o m i n a n t l y l ive r foci . I t thus a p p e a r s tha t , s imi-
lar ly to t he N S I - d e r i v e d B cel l h y b r i d o m a s , cel ls
f r o m l ive r m e t a s t a s e s m a y r e p r e s e n t a h i g h e r l eve l
o f d i f f e r e n t i a t i o n . I f h e r e a lso , t h e l ive r m e t a s t a s e s
a r e f o r m e d on ly in t h e p r e s e n c e o f t he sp l een , t h e n
the s p l e e n m i g h t h a v e s igna l l ed t u m o r cel ls to un-
d e r g o d i f f e r e n t i a t i o n r e q u i r e d fo r l i ve r h o m i n g .
3.2. Organ-xpecific growth o f non B cell hybridomas
T h e p r e v i o u s e x p e r i m e n t s i n d i c a t e d tha t , f o l l o w i n g
fu s ion wi th B l y m p h o c y t e s , b o t h N S I and SP2/0
p l a s m a c y t o m a cel ls g e n e r a t e d h y b r i d o m a s wh ich
m e t a s t a s i z e d to t h e s p l e e n and l iver . W o u l d s o m a -
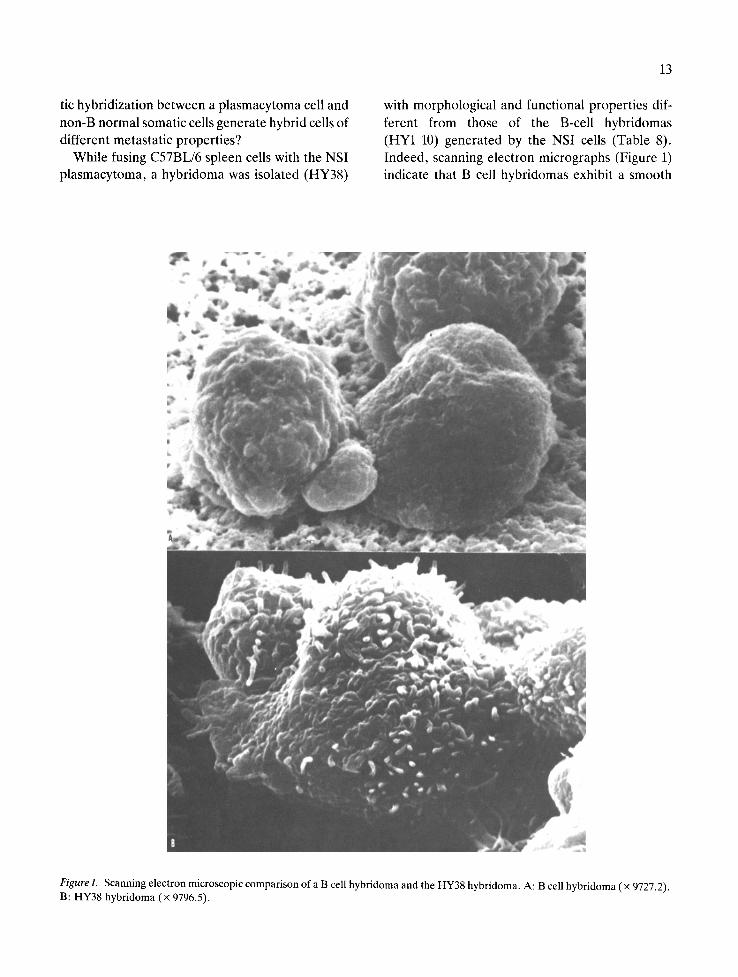
13
tic hybridization between a plasmacytoma cell and non-B normal somatic cells generate hybrid cells of different metastatic properties?
While fusing C57BL/6 spleen cells with the NSI plasmacytoma, a hybridoma was isolated (HY38)
with morphological and functional properties dif- ferent from those of the B-cell hybridomas (HYI-10) generated by the NSI cells (Table 8). Indeed, scanning electron micrographs (Figure 1) indicate that B cell hybridomas exhibit a smooth
Figure 1. Scanning electron microscopic comparison of a B cell hybridoma and the HY38 hybridoma. A: B cell hybridoma (× 9727.2). B: HY38 hybridoma (x 9796.5).

14
cytoplasmic membrane with occasional vesicular extrusions. In contrast, the NSI-derived HY38 hybridoma is an adherent cell with a cytoplasmic membrane which manifests the presence of multi- ple microvilli (finger-like extrusions). Further ana- lyses revealed that this adherent cell does not se- crete antibody and does not express classical B or T cell markers. Functional tests indicated that the HY38 hybridoma exerts accessory cell properties: it presented antigen to antigen-specific memory T cells, it secreted lymphocyte activating factor (LAF or IL-1) and it produced colony-stimulating factor. Hence, it appeared that the HY38 hybridoma may have derived from fusion between a NSI cell and a splenic macrophage or dendritic cell. When tested for its capacity to develop experimental metastases following intravenous inoculation, this hybridoma manifested a pattern of organ-specific growth com- pletely different from that of B cell hybridomas (22). We found (Table 9) that cells of the HY38 did not produce spleen or liver metastases. They did, however, produce heavy masses of lung meta- stases. Thus, fusion between plasmacytoma cells and non-B cells may result in the acquisition of metastatic competence, but the organ specificity of these hybrid cells is different from those of the B cell hybridomas.
3.3. Search for homing molecules controlling the organ specificity of metastatic hybridomas
It is generally believed that cell surface molecules represent 'homing receptors' for both normal and neoplastic cells. Since the metastatic properties of
NSI plasmacytomas were acquired following fu- sion with normal lymphocytes, we tested the effects of agents which were shown to affect lym- phocyte homing on the generation of metastases by the hybridoma cells. Thus, Degos et al. demon- strated that gene products of the MHC of lympho- cytes control their in vivo homing patterns to lym- phoid organs (21). Earlier studies indicated that the removal of sialic acid from lymphocytes impaired their homing properties (23). These two membrane moieties, i.e., sialic acid and H-2 antigens, appear also to play a crucial role in host-tumor interac- tions, as was reported for the Eb-Esb tumor model (24, 25). We therefore tested the possible involve- ment of those membrane molecules in the genera- tion of organ-specific metastases produced by two hybridomas: the HY2 which metastasizes to the spleen and liver, and HY8 which metastasizes to the liver only. It should be recalled that liver meta- stases generated by the HY2 cells is spleen control- led, unlike liver metastases produced by the HY8 cells (Table 4). We treated cells of these two hybridomas with (a) antibodies to the H-2 haplo- type of the parental normal B lymphocytes (anti H-2b), (b) neuraminidase, and (c) Con A. We found (Table 10) that the HY8 cells were not af- fected by any of these treatments. On the other hand, cells of the HY2 completely lost their capac- ity to generate spleen and liver metastases follow- ing anti-H-2 b treatment and significantly reduced this capacity following treatment with neur- aminidase or Con A. It is of interest to note that only HY2 cells whose metastases were spleen con- trolled were affected by agents which affected lym-
Table 9. Incidence of experimental metastases in animals inoculated with B cell (HY2, HYS) and non-B cell (Hy38) hybridomas.
Tumor Metastatic pattern
Spleen Liver Lung
Growth1 Weight (rag) Growth Weight (mg) Growth Weight (rag)
Hy2 + + + 1033 + + + 3392 - 240 HY8 - 242 + + 2621 - 242
HY38 - 128 - 1009 + + + 687
1 Legend to Table 2.
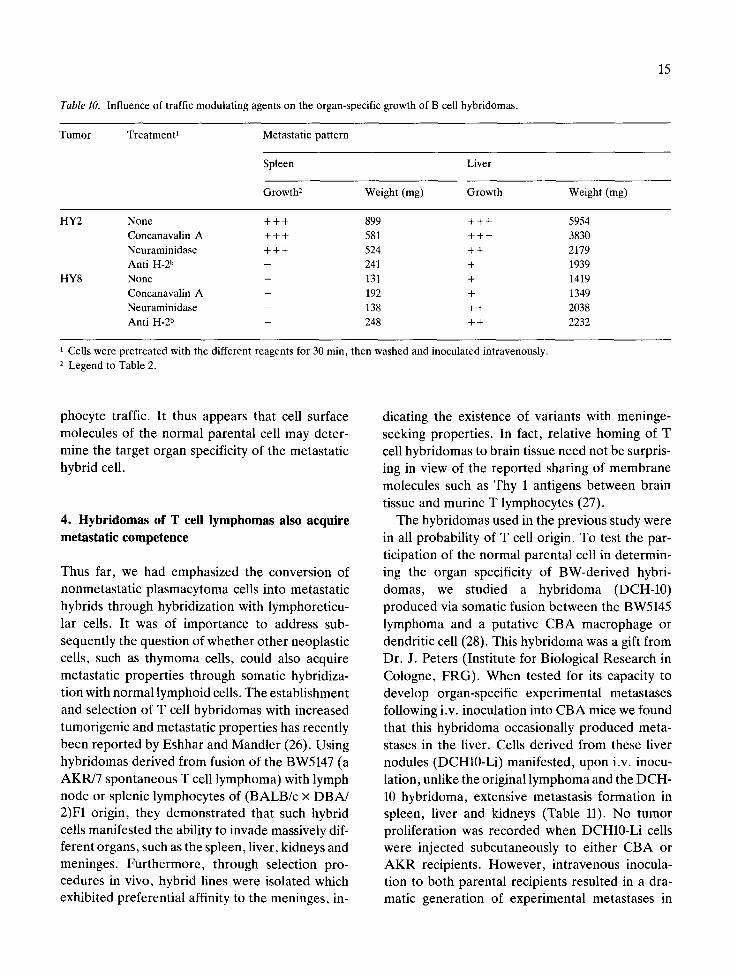
Table 10. Influence of traffic modulating agents on the organ-specific growth of B cell hybridomas.
15
Tumor Treatment 1 Metastatic pattern
Spleen Liver
Growth 2 Weight (mg) Growth Weight (rag)
HY2
HY8
None + + + 899 + + + 5954 Concanavalin A + + + 581 + + + 3830 Neuraminidase + + + 524 + + 2179 Anti H-2 b - 241 + 1939 None - 131 + 1419 Concanavalin A - 192 + 1349 Neuraminidase - 138 + + 2038 Anti H-2 b - 248 + + 2232
1 Cells were pretreated with the different reagents for 30 min, then washed and inoculated intravenously. 2 Legend to Table 2.
phocyte traffic. It thus appears that cell surface molecules of the normal parental cell may deter- mine the target organ specificity of the metastatic hybrid cell.
4. Hybridomas of T cell lymphomas also acquire metastatic competence
Thus far, we had emphasized the conversion of nonmetastatic plasmacytoma cells into metastatic hybrids through hybridization with lymphoreticu- lar cells. It was of importance to address sub- sequently the question of whether other neoplastic cells, such as thymoma cells, could also acquire metastatic properties through somatic hybridiza- tion with normal lymphoid cells. The establishment and selection of T cell hybridomas with increased tumorigenic and metastatic properties has recently been reported by Eshhar and Mandler (26). Using hybridomas derived from fusion of the BW5147 (a AKR/7 spontaneous T cell lymphoma) with lymph node or splenic lymphocytes of (BALB/c x DBA/ 2)F1 origin, they demonstrated that such hybrid cells manifested the ability to invade massively dif- ferent organs, such as the spleen, liver, kidneys and meninges. Furthermore, through selection pro- cedures in vivo, hybrid lines were isolated which exhibited preferential affinity to the meninges, in-
dicating the existence of variants with meninge- seeking properties. In fact, relative homing of T cell hybridomas to brain tissue need not be surpris- ing in view of the reported sharing of membrane molecules such as Thy 1 antigens between brain tissue and murine T lymphocytes (27).
The hybridomas used in the previous study were in all probability of T cell origin. To test the par- ticipation of the normal parental cell in determin- ing the organ specificity of BW-derived hybri- domas, we studied a hybridoma (DCH-10) produced via somatic fusion between the BW5145 lymphoma and a putative CBA macrophage or dendritic cell (28). This hybridoma was a gift from Dr. J. Peters (Institute for Biological Research in Cologne, FRG). When tested for its capacity to develop organ-specific experimental metastases following i.v. inoculation into CBA mice we found that this hybridoma occasionally produced meta- stases in the liver. Cells derived from these liver nodules (DCH10-Li) manifested, upon i.v. inocu- lation, unlike the original lymphoma and the DCH- 10 hybridoma, extensive metastasis formation in spleen, liver and kidneys (Table 11). No tumor proliferation was recorded when DCH10-Li cells were injected subcutaneously to either CBA or A K R recipients. However , intravenous inocula- tion to both parental recipients resulted in a dra- matic generation of experimental metastases in
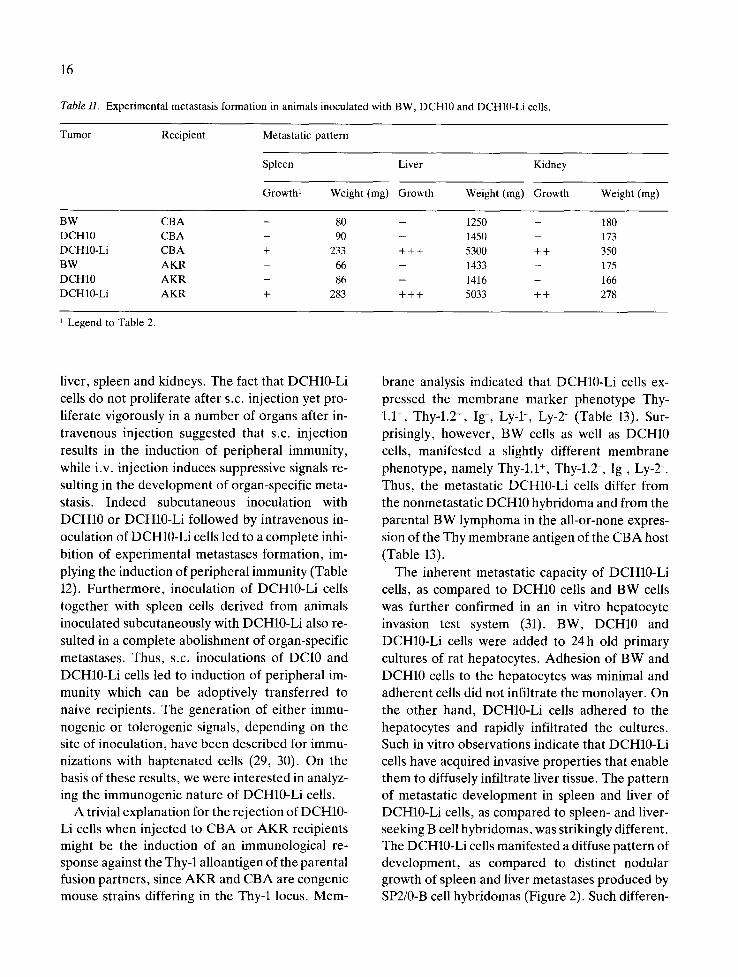
16
Table H. Experimental metastasis formation in animals inoculated with BW, DCH10 and DCH10-Li cells.
Tumor Recipient Metastatic pattern
Spleen Liver Kidney
Growth 1 Weight (mg) Growth Weight (mg) Growth Weight (mg)
BW CBA - 80 - 1250 - 180 DCH10 CBA - 90 - 1450 - 173 DCH10-Li CBA + 233 + + + 5300 + + 350 BW AKR - 66 - 1433 - 175 DCH10 AKR - 86 - 1416 - 166 DCH10-Li AKR + 283 + + + 5033 + + 278
Legend to Table 2.
liver, spleen and kidneys. The fact that DCH10-Li cells do not proliferate after s.c. injection yet pro- liferate vigorously in a number of organs after in- travenous injection suggested that s.c. injection results in the induction of peripheral immunity, while i.v. injection induces suppressive signals re- sulting in the development of organ-specific meta- stasis. Indeed subcutaneous inoculation with DCH10 or DCH10-Li followed by intravenous in- oculation of DCH10-Li cells led to a complete inhi- bition of experimental metastases formation, im- plying the induction of peripheral immunity (Table 12). Furthermore, inoculation of DCH10-Li cells together with spleen cells derived from animals inoculated subcutaneously with DCH10-Li also re- sulted in a complete abolishment of organ-specific metastases. Thus, s.c. inoculations of DC10 and DCH10-Li cells led to induction of peripheral im- munity which can be adoptively transferred to naive recipients. The generation of either immu- nogenic or tolerogenic signals, depending on the site of inoculation, have been described for immu- nizations with haptenated cells (29, 30). On the basis of these results, we were interested in analyz- ing the immunogenic nature of DCH10-Li cells.
A trivial explanation for the rejection of DCH10- Li cells when injected to CBA or AKR recipients might be the induction of an immunological re- sponse against the Thy-1 alloantigen of the parental fusion partners, since A K R and CBA are congenic mouse strains differing in the Thy-1 locus. Mem-
brane analysis indicated that DCH10-Li cells ex- pressed the membrane marker phenotype Thy- 1.1 ÷, Thy-l.2 ÷, I g , Ly-l-, Ly-2- (Table 13). Sur- prisingly, however, BW cells as well as DCH10 cells, manifested a slightly different membrane phenotype, namely Thy-l.1 ÷, Thy-l.2-, Ig-, Ly-2-. Thus, the metastatic DCH10-Li cells differ from the nonmetastatic DCH10 hybridoma and from the parental BW lymphoma in the all-or-none expres- sion of the Thy membrane antigen of the CBA host (Table 13).
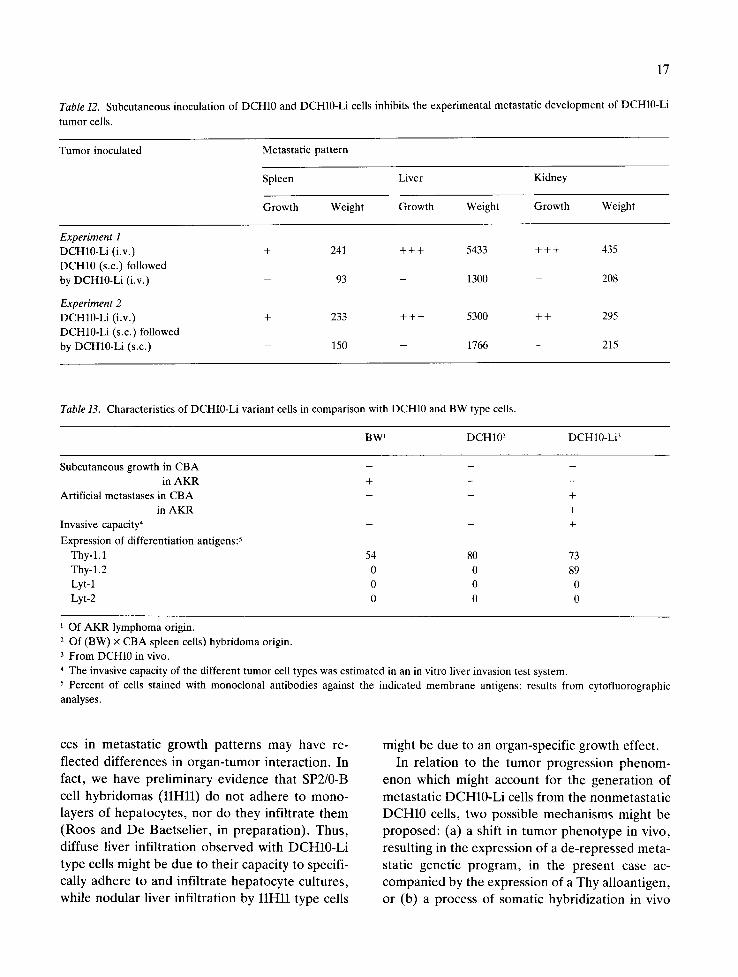
The inherent metastatic capacity of DCH10-Li cells, as compared to DCH10 cells and BW cells was further confirmed in an in vitro hepatocyte invasion test system (31). BW, DCH10 and DCH10-Li cells were added to 24h old primary cultures of rat hepatocytes. Adhesion of BW and DCH10 cells to the hepatocytes was minimal and adherent cells did not infiltrate the monolayer. On the other hand, DCH10-Li cells adhered to the hepatocytes and rapidly infiltrated the cultures. Such in vitro observations indicate that DCH10-Li cells have acquired invasive properties that enable them to diffusely infiltrate liver tissue. The pattern of metastatic development in spleen and liver of DCH10-Li cells, as compared to spleen- and liver- seeking B cell hybridomas, was strikingly different. The DCH10-Li cells manifested a diffuse pattern of development, as compared to distinct nodular growth of spleen and liver metastases produced by SP2/0-B cell hybridomas (Figure 2). Such differen-
17
Table 12. Subcutaneous inoculation of DCH10 and DCH10-Li cells inhibits the experimental metastatic development of DCH10-Li
tumor cells.
Tumor inoculated Metastatic pattern
Spleen Liver Kidney
Growth Weight Growth Weight Growth Weight
Experiment 1 DCH10-Li (i.v.) DCH10 (s.c.) followed by DCH10-Li (i.v.)
Experiment 2 DCH10-Li (i.v.) DCH10-Li (s.c.) followed by DCH10-Li (s.c.)
+ 241 +++ 5433 +++ 435
- 93 - 1300 - 208
+ 233 +++ 5300 ++ 295
- 150 - 1766 - 215
Table 13. Characteristics of DCH10-Li variant cells in comparison with DCH10 and BW type cells.
BW 1 DCH102 DCH10-Li 3
Subcutaneous growth in CBA in AKR +
Artificial metastases in CBA in AKR
Invasive capacity 4
Expression of differentiation antigens: 5 Thy-l.1 54 Thy-l.2 0 Lyt-1 0 Lyt-2 0
80 73 0 89 0 0 0 0
1 Of AKR lymphoma origin. 2 Of (BW) × CBA spleen cells) hybridoma origin. 3 From DCH10 in vivo. 4 The invasive capacity of the different tumor cell types was estimated in an in vitro liver invasion test system. 5 Percent of cells stained with monoclonal antibodies against the indicated membrane antigens; results from cytofluorographic analyses.
ces in metasta t ic growth pa t te rns may have re-
flected differences in o rgan - tumor in teract ion. In
fact, we have pre l iminary evidence that SP2/0-B
cell hybr idomas ( l l H l l ) do not adhere to mono-
layers of hepatocytes , no r do they infi l trate them
(Roos and De Baetsel ier , in prepara t ion) . Thus ,
diffuse liver inf i l t ra t ion observed with DCH10-Li
type cells might be due to their capacity to specifi-
cally adhere to and infi l t rate hepatocyte cultures,
while nodu la r liver inf i l t ra t ion by l l H l l type cells
might be due to an organ-specific growth effect.
In re la t ion to the tumor progress ion phe nom-
e n o n which might account for the genera t ion of
metastat ic DCH10-Li cells f rom the nonmetas ta t i c
DCH10 cells, two possible mechanisms might be
proposed: (a) a shift in t umor pheno type in vivo,
resul t ing in the expression of a de-repressed meta-
static genet ic program, in the present case ac-
compan ied by the expression of a Thy a l loant igen,
or (b) a process of somatic hybr id iza t ion in vivo
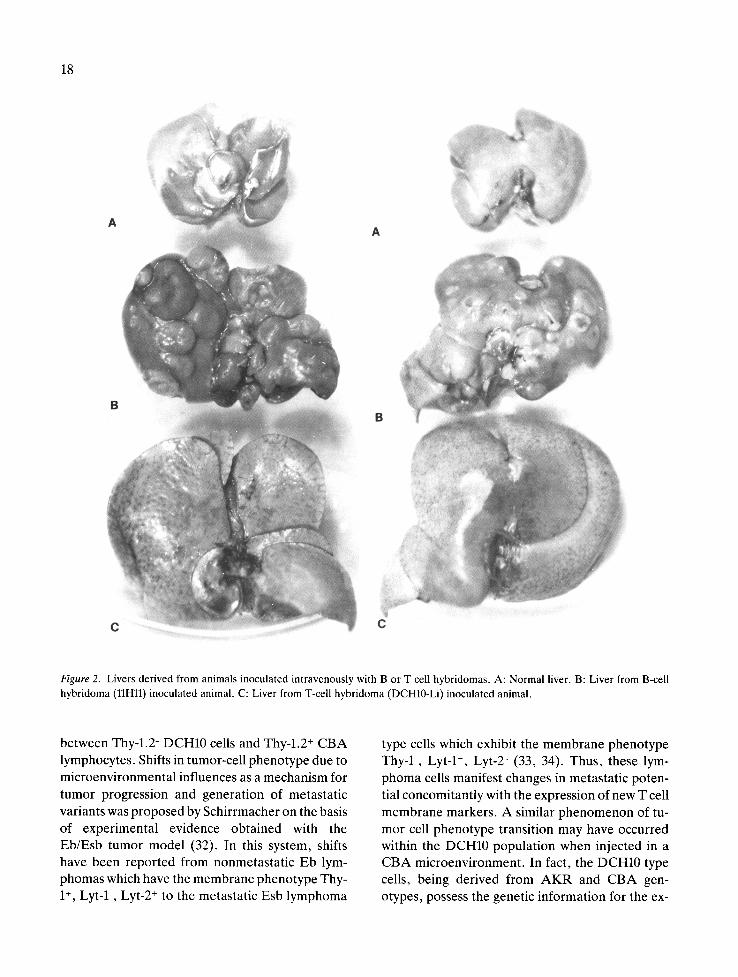
18
Figure 2. Livers derived from animals inoculated intravenously with B or T cell hybridomas. A: Normal liver. B: Liver from B-cell hybridoma (llHll) inoculated animal. C: Liver from T-cell hybridoma (DCH10-Li) inoculated animal.
between Thy-l.2- DCH10 cells and Thy-l.2 + CBA lymphocytes. Shifts in tumor-cell phenotype due to microenvironmental influences as a mechanism for tumor progression and generation of metastatic variants was proposed by Schirrmacher on the basis of experimental evidence obtained with the Eb/Esb tumor model (32). In this system, shifts have been reported from nonmetastatic Eb lym- phomas which have the membrane phenotype Thy- 1 +, Lyt-l-, Lyt-2 + to the metastatic Esb lymphoma
type cells which exhibit the membrane phenotype Thy-l-, Lyt-1 ÷, Lyt-2- (33, 34). Thus, these lym- phoma cells manifest changes in metastatic poten- tial concomitantly with the expression of new T cell membrane markers. A similar phenomenon of tu- mor cell phenotype transition may have occurred within the DCH10 population when injected in a CBA microenvironment. In fact, the DCH10 type cells, being derived from A K R and CBA gen- otypes, possess the genetic information for the ex-

pression of Thy-l.2 membrane antigen and its ex- pression might be induced in vivo. However, the alternative explanation for the generation of meta- static DCH10-Li, namely in situ somatic hybridiza- tion cannot be excluded. In fact, based on docu- mented observations and recent experimental evidence, we tend to favor this last possibility, as will be discussed in the next section.
5. In vivo hybridization: A model for tumor progression?
Fusion in vivo between tumor cells and host cells has been shown to occur in a variety of animal models. Unfortunately, none of the tumor cell-host cell hybrids thus formed were examined in any detail for changes in their relative tumorigenic or metastatic capabilities. There is, however, a recent study which implies that the in vivo formation of tumor-host cell hybrids could lead to the emer- gence of more malignant tumor cell variants as defined by metastatic ability. We refer to the re- sults of Kerbei et al. (35, 36) who, using a non- metastatic variant (MDW-D4) of the highly meta- static MDAY-D2 DBA/2 (H-2 d) mouse tumor, demonstrated that such variants became metastatic in vivo after a cellular change took place, such as extinction of recessive lectin or drug sensitivity and acquisition of a higher number of chromosomes. Supportive evidence for a spontaneous cell fusion in vivo between the MDW4 tumor cells and host cells was provided by the observation that growth of MDW4 tumor cells in either (H-2k X H-2d)F1 mice or (H-2k)--~H-2 d bone marrow radiation chimeras led to the appearance of MDW4 derived cells which express H-2 k antigens (37). Thus in this experimental tumor model, tumor progression might have arisen as a consequence of tumor-host cell fusion in vivo followed by chromosome segre- gation.
In view of the results obtained with the DCH10 tumor system, in which transplantation of Thy-l.1 + DCH10 cells to a CBA recipient led to the ap- pearance of the Thy-l.1 +, Thy-l.2 + DCH10-Li met- astatic variant, we were interested in testing whether transplantation of the Thy-l.1 + BW lym-
19
phoma to CBA recipients might lead to analogous phenomena. In fact after i.v. injection of BW cells to CBA mice, we found that one animal developed a tumor mass in the liver. Cells derived from this liver nodule termed BW-Li, produced upon i.v. injection to CBA mice metastases in the spleen, liver, kidneys and ovaries in 100% of the recipients. Furthermore, BW-Li cells, in contrast to BW cells, were found to invade in vitro monolayers of hepatocytes confirming their inherent invasive po- tential (De Baetselier, Roos et al., in preparation). Membrane analyses of BW-Li cells revealed that such cells were Thy-l.1 +, Thy-l.2 ÷, Ly-1 +, Ly-2-, as compared to BW cells which exhibited the mem- brane phenotype Thy-l.1 +, Thy-l.2-, Ly-l- , Ly-2-. Thus, BW-Li cells express the Thy-l.1 marker of the BW lymphoma and the Thy-l.2 marker of the CBA recipient. These results provide supportive evidence for a spontaneous fusion in vivo between the BW cells and Thy-l.2 + CBA host cells. The acquisition of T cell membrane markers of the CBA host (i.e., Thy-l.2, Ly-1) may have led to the expression of metastatic and invasive capabilities. This phenomenon then reflects our observations obtained with in vitro somatic hybridizations.
Regarding the process of in vivo hybridization as a possible mechanism for generation of tumor het- erogeneity, tumor progression and metastasis, a number of conditions should be fulfilled in order to increase the probability of such a process to occur in vivo: (a) The tumor cell or tumor cell variant should be a good fuser. In fact, the experimental data summarized in this review deal with plas- macytoma and lymphoma cells which have been selected on the basis of their capacity to fuse effi- ciently with normal somatic cells. Hence, in vivo cellular fusion processes may happen only with certain fusable clones of tumor cells. (b) The pre- sence of fusion-inducing substances, such as viruses might be a prerequisite for in vivo fusion; (c) the nature of the infiltrating host cell might be of cardi- nal importance in determining the emergence of new metastatic cells within a local tumor cell popu- lation. In this context one should be aware of the fact that non-malignant cells within neoplasms ex- hibit heterogeneity which is apparently related to the heterogeneity of the tumor cell population.

20
Using subclones of a BALB/c C3H mouse mamm- ary carcinoma, Heppner et al. demonstrated that the type and percentage of lymphocytes infiltrating tumors varied among different lines (38). Similarly using metastatic and nonmetastatic clones of the T10 sarcoma we found (Eisenbach et al., submit- ted) that local tumors produced by low metastatic clones were heavily infiltrated by macrophages, whereas local tumors produced by high metastatic clones were not. Thus, depending on the nature of a specific tumor cell population, different types of lymphoreticular cells might be selectively at- tracted. Furthermore, not only the nature of the infiltrating host cell might be of importance in con- ferring metastatic properties via cell fusion, but also the state of.activation or differentiation of the putative fusion partner. Indeed, activated T cells are endowed with more invasive properties than normal T cells, such as the potential to invade hepatocyte monolayers (39) or vascular endo- thelial cell monolayers and to degrade sulfated proteoglycans (40). This might be of relevance with regard to the BW-Li type cells, which were found to express differentiation markers of mature T cells (i.e., Lyt-1), unlike the parental BW cells, and to infiltrate hepatocyte monolayers in a way compar- able to activated T cells. Considering these param- eters, i.e., tumor-mediated selective infiltration, phenotype of infiltrating cells and differentiation status of the infiltrating cells, one could assume that an enhancement of the local presence of cer- tain types of normal somatic cells within the local tumor might increase the incidence of tumor-host cell fusion in vivo. Indicative evidence for this is provided by the following experiments. In an at- tempt to analyse the effect of the local presence of macrophages within a polyclonal metastasizing tu- mor, the 3LL Lewis lung carcinoma, we serially injected s.c. 3LL cells admixed with macrophages. After s.c. transfer of 3LL tumors with normal per- itoneal macrophages for six transplant generations, the tumor cells (3LL/R6) were found to be more immunogenic in the presence of macrophages and in many instances tumor rejection was observed (41, 42). When continuing this macrophage-medi- ated selection procedure for nine additional trans- plant generations we found that the tumor cells
(3LL/R15) were strongly tumorigenic when inocul- ated with or without macrophages. In fact, the 3LL/R15 tumor cells appeared to be a new tumor variant with completely new characteristics as com- pared to the 3LL and 3LL/R6 tumor cells. 3LL/R15 type cells are small, nonadherent cells which ex- press both H-2K and H-2D gene products of the H-2 b haplotype, while 3LL and 3LL/R6 type cells are large, adherent cells which express mainly the H-2D b alloantigens. Furthermore, 3LL/R15 type cells manifest a completely new metastatic pattern. Whereas 3LL and 3LL/R6 cells metastasize only to one single organ, the lungs, the 3LL/R15 cells gen- erated spontaneous metastases in the lymph nodes and artificial metastases in the kidney, liver and spleen (Table 14). What is the origin of such a tumor variant which differs completely in tumori- genic and metastatic properties? On the basis of morphological criteria and of expression of cell surface markers we suggest that the 3LL/R15 cells were generated via an in vivo fusion process be- tween 3LL cells and infiltrating host cells. How- ever, since these experiments were carried out in a completely syngeneic system and since the 3LL cells were not tagged with genetic markers, we cannot exclude other mechanisms which might ac- count for the induction of 3LL/R15 type cells. In- deed one should consider other possibilities such as the phenomenon of 'carcinogenic tumors' (43, 44), in which transplantation of tumor cells results in tumor formation by host cells or 'host cell mediated mutagenesis', in which macrophage production of oxygen radicals might result in tumor cell muta- genesis (45, 46). Be the mechanism underlying the generation of 3LL/R15 type cells as it may, we want to stress that the induction of such a variant has occurred through tumor growth in an environment enriched with syngeneic host cells. In such an en- vironment the probability of host cell-tumor inter- action, including host cell-tumor fusion, is ex- pected to increase.
6. Conclusions and implications
We have reviewed our studies indicating that soma- tic cell hybridization of nonmetastatic plasma-
cytomas or lymphoma cells with normal lympho- cytes or macrophages results in hybrid cells which manifest metastatic properties. The metastases produced by the different types of hybridomas manifested different target organ specificities (Table 15). With regard to a given type of neoplas- tic partner, the phenotype of the somatic normal partner seems to determine the target organ specif- icity of its metastases. This is deduced from our demonstration that cells of two different plas- macytomas, the NSI and the SP2/0, when hybri- dized with B-lymphocytes yielded hybrids which metastasize to the spleen and liver, whereas a hybrid in which the normal partner seemed to have been a macrophage, metastasized to the lungs. The role of the normal partner in controlling the target organ specificity is further supported by our dem- onstration that antibodies directed against the H-2
21
phenotype of the normal partner in the HY2 hybridoma prevented metastasis formation in a homing system which was expected to be control- led by gene products of the normal parental lym- phocyte MHC.
The application of the technique of somatic hybridization to generate metastatic cells from nonmetastatic neoplasms was recently adopted by two other groups of investigators. Kerbel et al. (37) reported that cells of the DBA/2 originated non- metastatic MDW4 tumor acquired metastatic properties following in vitro fusion with either spleen or bone marrow-derived cells. Similarly, using the Eb/Esb system, Larizza et al. (to be pub- lished) demonstrated that fusion of Eb cells with syngeneic macrophages resulted in the generation of metastatic cell lines.
In all our experiments, and in those of others,
Table 14. Organ-specific growth of 3LL, 3LL(R) and 3LL(R)15 cells after intravenous inoculation.
Tumor Metastatic pattern
Spleen Liver Lung Kidney
Growth I Weight Growth Weight Growth Weight Growth Weight (mg) (mg) (rag) (mg)
3LL - 129 - 1912 + + + 920 - 146 3LL(R) 15 + 291 + 1927 - 267 + + + 625 3LL(R) - 217 - 1474 + + + 1022 - 188
i Legend to Table 2.
Table 15. Origin and metastatic pattern of different somatic hybrids.
Tumor cells Origin Metastatic pattern
HYI-10 (Balb/c NSI plasmacytoma × Liver C57BL/6 B lymphocyte) Spleen and liver
11Hll (Balb/c SP210 plastocytoma x Balb/c B lymphocyte) Spleen and liver
HY38 (Balb/c NSI plasmacytoma × C57BL/6 accessory cell (?)) Lungs
DCH10-Li (BW derived hybridoma × CBA T cell (?) in vivo) Spleen, liver, kidneys
BW-Li (BW lymphoma x CBA T cell (?) Spleen, liver, kidneys, in vivo) ovaries

22
somatic hybrids which acquired metastatic compe- tence appear to have derived from the lympho- reticular system. Two obvious questions emerge: (a) Can cells of nonmetastatic tumors of non- lymphoid origin also acquire metastatic properties through somatic hybridization with normal lym- phoid cells? (b) Can somatic cells of nonlymphoid phenotype confer, via somatic hybridization, meta- static properties on nonmetastatic tumor cells? We tend to suggest that the 'circulating' capacity of the normal parental partners such as lymphocytes might represent molecular properties which deter- mine the acquisition of metastatic competence via somatic cell fusion. Lymphocytes share with dis- seminating tumor cells a number of properties rele- vant to the metastatic process: the capacity to migr- ate via the blood circulation, to survive while migrating, to penetrate blood vessels, to invade different tissues and to home to defined regions of the lymphoid system. Hence, fusion of lympho- cytes with neoplastic cells may confer such proper- ties on the hybrid tumor cells, resulting in the ac- quisition of metastatic competence.
Since some of these functions seem to be deter- mined by membrane properties, the generation of metastatic hybridomas may provide us with a use- ful experimental system for the analysis of the func- tion of defined membrane molecules in controlling the metastatic pattern of different tumors. In fact, phenomena such as a similar target organ specif- icity for metastases of different tumors, may be based on different membrane properties. Thus, we found that two hybridomas of two distinct cellular origins, both of which metastasize preferentially to the liver, manifest two distinct patterns of tumor cell-target organ interactions. The DCH10-Li cells - of a T cell hybridoma- adhere to hepatocytes and rapidly infiltrate hepatocyte monolayers. On the other hand, the SP2/0-B cell hybridomas, which also generate liver metastases, do not adhere to hepatocytes, nor do they infiltrate them. In the first case of the DCH10-Li, liver metastases might de- rive from specific cell-cell recognition between the tumor and the hepatocytes, whereas in the second system of the B-cell hybridomas, liver metastases may be controlled by liver-specific growth signals. Such different interactions may account for the
different patterns of liver metastases: the nodular growth of the liver metastases of the B-cell hybri- domas, as distinct from the diffuse growth of the DCH10-Li cells.
Thus far, most experimental studies on cancer metastasis have compared particular tumor cells and variants thereof. Such variants have been iso- lated, for instance by cloning in vitro (47), by se- lecting for organ specificity (48) and by resistance to plant lectins (49, 50) or immune effectors (51-53). In many cases, however, one is confronted with (a) the phenotypic instability of the metastatic variants (54) and (b) the question of the exact origin of the metastatic variant, namely whether the putative variant is a real descendant of the primary tumor or a newly induced tumor. The use of somatic hybridization as a tool to engineer sets of malignant cells having the same origin but different metastatic characteristics may provide a new useful model to analyze cell surface properties of import- ance for metastasis.
Is somatic cell hybridization just an experimental tool for the study of cellular and molecular deter- minants which control tumor metastasis, or does it also represent a process through which nonmeta- static tumor cells may acquire in vivo metastatic properties? Kerbel et al., using drug resistant cells of a nonmetastatic tumor variant, reported re- cently that such cells became metastatic in vivo and the properties acquired by the tumor cells could be attributed to tumor-host cell fusion which might have taken place in vivo (37). In our studies of the BW-Li cells, we indicated that the tumor cells, while acquiring metastatic competence following in vivo growth, acquired phenotypic properties of cell surface molecules which could also be ex- plained by spontaneous fusion between BW cells and Thy-l.2 + CBA host cells. It is clear that at this stage it is impossible to evaluate to what extent such in vivo fusions constitute, in terms of inci- dence, a significant process of conversion of non- metastatic to metastatic tumor cells. Yet it is also clear that the observations reported in the present review make the question of whether and to what extent tumor progression in vivo involves somatic cell fusion worth investigating.

Acknowledgements
This investigation was supported by Grant No. 28139, awarded by the National Cancer Institute, Department of Health and Human Services of the United States and by the Hermann and Lilly Schil- ling Foundation, Essen, FRG. Dienst Algemeen Biologie VUB was supported by a grant of the ASKL Kankerfonds.
References
1. Barski G, Sorieul S, Cornefert F: 'Hybrid'-type cells in combined cultures of two different mammalian cell strains. J Natl Cancer Inst 26: 1269-1291, 1961.
2. Barski G, Cornefert F: Characteristics of 'hybrid'-type cell lines obtained from mixed cultures in vitro. J Natl Cancer Inst 28: 801-821, 1962.
3. Harris H, Mimmer O J, Klein G, Worst P, Tachibana, T: Suppression of malignancy by cell fusion. Nature 223: 363-368, 1969.
4. Klein, G. Bregula U, Wiener F, Harris H: The analyses of malignancy by cell fusion. I. Hybrids between tumor cells and L cell derivatives. J Cell Sci 8: 659-672, 1971.
5. Wiener F, Feny6 EM, Klein G: Fusion of tumor cells with host cells. Nature New Biol 238: 155-159, 1972.
6. Wiener F, Feny6 EM, Klein G: Tumor-host cell hybrids in radiochimeras. Proc Natl Scad Sci USA 71: 147-152, 1974.
7. Feny6 EM, Wiener F, Klein G, Harris H: Selection of tumor-host cell hybrids from polyoma virus- and methyl- cholanthrene-induced sarcomas. J Natl Cancer Inst 51: 1865-1875, 1973.
8. Ber R, Wiener F, Feny6 EM: Proof of in vivo fusion of murine tumor cells with host cells by universal fusers. J Natl Cancer Inst 60: 931-933, 1978.
9. Lala PK, Santer V, Rahil KS: Spontaneous fusion between Ehrlich ascites tumor cells and host cells in vivo: Kinetics of hybridization, and concurrent changes in the histocom- patibility profile of the tumor after propagation in different host strains. Eur J Cancer 16: 487-510, 1979.
10. Marshall MJ, Shone DG, Windle JM, Worsfold M: Spon- taneous fusion of malignant and host mouse cells in culture detected by phosphoglucose isomerase isoenzymes (GPI). Br J Cancer 46: 811-816, 1982.
11. Goldenberg DM, Pavia RA, Tsao MC: In vivo hybridiza- tion of human tumor and normal hamster cells. Nature 250: 649-651, 1974.
12. Katzav S, De Baetselier P, Tartakovsky B, Segal S, Feld- man M: Immunogenetic properties of tumor metastases. In: Vitetta ES (ed) B and T cell Tumors. Academic Press, New York, 1982, pp 233-245.
13. Katzav S, De Baetselier P, Tartakovsky B, Gorelik E, Segal S, Feldman M: Immunogenetic determinants control-
23
ling the metastatic properties of tumor cells. In: Fabris N (ed) Immunoregulation. Plenum Press, New York, 1983, pp 453-463.
14. De Baetselier P, Katzav S, Gorelik E, Feldman M, Segal S: Differential expression of H-2 gene products in tumor cells is associated with their metastatic properties. Nature 288: 179-181, 1980.
15. Katzav S, De Baetselier P, Gorelik E, Feldman M, Segal S: Immunologic control of metastasis formation by a methyl- cholanthrene-induced tumor (T10) in mice: Differential ex- pression of H-2 gene products. Transpl Proc XIII: 742-746, 1981.
16. De Baetselier P, Gorelik E, Eshhar Z, Ron Y, Katzav S, Feldman M, Segal S: Metastatic properties conferred on nonmetastatic tumors by hybridization of spleen B lympho- cytes with plasmacytoma cells. J Natl Cancer Inst 67: 1079-1087, 1981.
17. Witte PL, Ber R: Improved efficiency of hybridoma ascites production by intrasplenic inoculation in mice. J Natl Can- cer Inst 70: 575-577, 1983.
18. Slavin S, Morecki S, Weiss L: The role of the spleen in tumor growth: Kinetics of the murine B cell leukemia (BCL1). J Immunol 124: 586--589, I980.
19. Ron Y, De Baetselier P, Segal S: Involvement of the spleen in murine B cell differentiation. Eur J Immunol 11: 94-99, 1981.
20. Ron Y, De Baetselier P, Feldman M, Segal S: Involvement of the spleen in the control of the immunogenic and pha- gocytic function of thioglycollate-induced macrophages. Eur J Immunol 11: 608-611, 1981.
21. De Baetselier P, Gorelik E, Eshhar Z, Ron Y, Katzav S, Feldman M, Segal S: Organ-specific homing of B cell hybridomas. In: Nieuwenhuis P, van den Broek AA, Hanna MG (eds) In vivo immunology. Plenum, New York, 1982, pp 179-185.
22. Degos L, Pla M, Colombani JM: H-2 restriction for lym- phocyte homing into lymph nodes. Eur J Immunol 9: 808-814, 1979.
23. Woodruff J J, Gesner BM: The effect of neuraminidase on the fate of transfused lymphocytes. J Exp Med 129: 551-567, 1969.
24. Schirrmacher V, Cheingsong-Popv R, Arnheiter H: Hepatocyte tumor cell interaction in vitro. I. Conditions for rosette formation and inhibition by anti H-2 antibodies. J Exp Med 151: 984-988, 1980.
25. Fogel M, Altevogt P, Schirrmacher V: Metastatic potential severely altered by changes in tumor cell adhesiveness and cell-surface sialylation. J Exp Med 157: 371-376, 1983.
26. Eshhar Z, Mandler R: Changes in tumorigenic and meta- static pattern of thymoma cells induced by their hybridiza- tion to normal lymphocytes. In: Vitetta ES (ed) B and T cell tumors. Academic Press, New York, 1982, pp 313--318.
27. Williams AF, Barclay AN, Letarte-Muirhead M, Morris RJ: Rat Thy-1 antigens from thymus and brain: Their tissue distribution, purification and chemical composition. Cold Spring Harbor Symp Quant Biol 41: 51-6l, 1976.

24
28. Peters JH: Hybridomas of mouse dendritic cells (DC) ex- pressing phenotypic markers of DC including growth-stim- ulating action on T-lymphocytes. In: Pusch K, Kirchner H (eds) Mechanisms of lymphocyte activation. Elsevier/ North Holland, 1981, pp 537-540.
29. Britz JS, Askenase PW, Ptak W, Steinman RM, Gershon RK: Specialized antigen-presenting cells. Splenic dendritic cells and peritoneal exudate cells induced by mycobacteria activate effector T cells that are resistant to suppresion. J Exp Med 155: 1344-1356, 1982.
30. Claman HN, Miller SD, Conlov PJ, Moorhedd JW: Con- trol of experimental contact sensitivity. Adv in lmmuno130: 121-158, 1980.
31. Roos E, van de Pavert I, Middelhoop OP: Infiltration of tumor cells into cultures of isolated hepatocytes. J Cell Sci 47: 385-397, 1981.
32. Schirrmacher V: Shifts in tumor cell phenotypes induced by signals from the microenvironment. Relevance for the im- munobiology of cancer metastasis. Immunobiology 157: 89-98, 1980.
33. Schirrmacher V, Fogel M, Russmann E, Bosslet K, AI- tevogt P, Beck L: Antigenic variation in cancer metastasis: Immune escape versus immune control. Cancer Metastasis Rev 1: 241-274, 1982.
34. Altevogt P, Kurnick JT, Kimura AK, Bosslet K, Schirr- macker V: Different expression of Lyt differentiation anti- gens and cell surface glycoproteins by a murine T lym- phoma line and its high metastatic variant. Eur J Immunol 12: 300-307, 1982.
35. Dennis J, Donaghue TP, Florian M, Kerbel RS: Apparent reversion of stable in vitro genetic markers detected in tumor cells from spontaneous metastases. Nature 292: 242-245, 1981.
36. Kerbel RS, Dennis JW, Lagarde AE, Frost P: Tumor pro- gression in metastasis: An experimental approach using lectin resistant tumor variants. Cancer Metastasis Rev 1: 93-140, 1982.
37. Kerbel RS, Lagarde AE, Dennis JW, Donaghue TP: Spon- taneous host cell × tumor cell hybridization in vivo: Contri- bution to the emergence of metastatic cell variants. Mol Cell Biol 3: 523-538, 1983.
38. Rios A, Laux D, Heppner GH: Patterns of lymphocyte infiltration in tumor sublines of a single mammary ade- nocarcinoma (Abstract). Proc AACR 28: 1021, 1982.
39. Roos E, Van de Pavert IV: Antigen-activated T lympho- cytes infiltrate hepatocyte cultures in a manner comparable to liver-colonizing lymphosarcoma cells. Clin Exp Metast 1: 173-180, 1983.
40. Vlodavsky I, Ariav Y, Fuks Z, Altevogt P, Schirrmacher V: Lymphoma cell-mediated degradation of sulfated proteo-
glycans in the subendothelial basa lamina: Relation to tu- mor cell metastasis. Cancer Res (in press).
41. De Baetselier P, Kapon A, Feldman M, Segal S: Activated macrophages modulate the tumorigenicity of 3LL tumor cells. In: Levy E (ed) Advances in pathology, Vol 1. Per- gamon Press, Oxford & New York, 1982, pp 305-308.
42. De Baetselier P, Kapon A, Katzav S, Feldman M, Segal S: Selection of an immunogenic 3LL tumor subline following serial growth in vivo in the local presence of peritoneal macrophages. In: Norman S J, Sorkin E (eds) Macrophages and natural killer cells. Plenum Publ Corp, New York, 1982, pp 281-288.
43. Kerbel RS, Florian M, Man, MS, Dennis J, McKenzie IFC: Carcinogenicity of tumor cell populations: Origin of a puta- tive H-2 isoantigenic loss variant tumor. J Natl Cancer Inst 64: 1221-1230, 1981.
44. Frost P, Kerbel RS, Tartamella-Blondo R: Generation of highly metastatic tumors in DBA/2 mice. Invasion Meta- stasis 1: 22-33, 1981.
45. Weitzman SA, Stossel TP: Mutation caused by human pha- gocytes. Science 212: 546-547, 1981.
46. Weitzman SA, Stossel TP: Effects of oxygen radical scav- engers and antioxidants on phagocyte-induced muta- genesis. J Immunol 128: 2770-2772, 1982.
47. Kripke ML, Gruys E, Fidler IJ: Metastatic heterogeneity of cells from an ultraviolet light-induced murine fibrosarcoma of recent origin. Cancer Res 38L: 2962-2967, 1978.
48. Nicolson GL, Winkelhake JL: Organ specificity of blood- - borne metastasis determined by cell adhesion. Nature 299:
230, 1975. 49. Burger MM: The cell surface and metastasis. In: Biology of
the cancer cell. Kugler, Amsterdam, 1980, pp 193-208. 50. Kerbel RS, Dennis JW, Lagarde AE, Frost P: Tumor pro-
gression in metastasis. An experimental approach using lectin-resistant tumor variants. Cancer Metastasis Rev 2: 99-140, 1982.
51. Fidler IJ, Gersten DM, Bodmen MB: Characterization in vivo and in vitro of tumor cells selected for resistance to syngeneic lymphocyte-mediated cytotoxicity. Cancer Res. 36: 3160-3169, 1976.
52. Frost P, Kerbel RS: Immunoselection in vitro of a non- metastatic variant from a highly metastatic tumor. Int J Cancer 27: 381-385, 1981.
53. Bosslet K, Schirrmacher V: Escape of metastasizing clonal tumor cell variants from tumor-specific cytolytic T lympho- cytes. I Exp Med 154: 557-562, 1981.
54. Poste G, Doll J, Fidler IJ: Interactions between clonal subpopulations affect the stability of the metastatic phe- notype in polyclonal populations of B16 melanoma cells. Proc Natl Acad Sci USA 78: 6226-6230, 1981.





























