Embed Size (px)
Citation preview

Estudio geneacutetico de la progenie del pulpo comuacuten
(Octopus vulgaris) y mejora del alimento vivo para su
cultivo larvario
Lema SUSTAINABLE OCTOPUS
RESUMEN
El pulpo comuacuten (Octopus vulgaris) es una especie de alto valor comercial en Galicia y
de elevado intereacutes cultural y gastronoacutemico En las uacuteltimas deacutecadas la demanda de
pulpo superoacute largamente la capacidad pesquera siendo necesario recurrir a
importaciones En los antildeos 90 se iniciaron una serie de trabajos de cultivo de pulpo en
cautividad cuyo objetivo era analiziar su potencial para la acuicultura Eacutestos
experimentos confirmaron el potencial de esta especie debido a sus interesantes
caracteriacutesticas bioloacutegicas raacutepido crecimiento y ciclo de vida corto alta eficacia de
conversioacuten del alimento elevada fecundidad resistencia a la manipulacioacuten y
adaptacioacuten a la vida en cautividad Aunque los resultados de engorde eran
prometedores y rentables sigue persistiendo un importante cuello de botella en su
cultivo integral el cultivo de las paralarvas planctoacutenicas Por ello el presente trabajo
tuvo como principales objetivos estudiar la hipoacutetesis de paternidad muacuteltiple en las
puestas de pulpo y sus consecuencias a nivel de diversificacioacuten geneacutetica y optimizar
el valor nutricional del alimento vivo para el cultivo de paralarvas evaluaacutendose el
efecto de diferentes dietas sobre el crecimiento y la supervivencia Se ha demostrado
la existencia de muacuteltiple paternidad en O vulgaris Eacutesta puede representar un proceso
que maximiza la recombinacioacuten geneacutetica de la hembra con los varios machos en un
uacutenico acontecimiento reproductivo Por otro lado se ha demostrado que el
enriquecimiento de Artemia con microalgas y aminoaacutecidos esenciales (lisina arginina y
metionina) origina una mejora del peso seco y supervivencia de las paralarvas a los
25 diacuteas de vida El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del
pulpo y de la nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo
integral de esta especie pudiendo igualmente ser uacutetil en el futuro para hacer una
gestioacuten sostenible de las poblaciones naturales
Palabras clave pulpo repoblacioacuten cultivo sostenibilidad geneacutetica diversidad
2
1 Introduccioacuten
11 Biologiacutea y ciclo de vida del pulpo comuacuten (Octopus vulgaris)
El pulpo comuacuten (Octopus vulgaris Cuvier 1797)
tambieacuten denominado pulpo de roca (Fig 1) es
uno de los moluscos cefaloacutepodos mejor
estudiados en todo el mundo destacaacutendose entre
las maacutes de cien especies descritas del geacutenero
Octopus Esta especie ha sido modelo de
investigacioacuten en diversos estudios que incluyen
aacutereas como la neurobiologiacutea la fisiologiacutea animal
bioquiacutemica y comportamiento animal (Young
1971 Wells 1978 Boyle 1983) En los uacuteltimos
15 antildeos se ha enfatizado ademaacutes el estudio de su biologiacutea y ciclo de vida en
cautividad con el objetivo de aplicar estos conocimientos a su cultivo integral (Iglesias
et al 2007) El pulpo es un animal estrictamente marino que se distribuye en aguas
templadas y tropicales de todo el mundo Vive solitario es territorial y realiza
migraciones estacionales acercaacutendose en primavera a zonas maacutes someras para
reproducirse Su ciclo de vida es relativamente corto estimaacutendose entre uno y dos
antildeos en el medio natural aunque seguacuten diferentes autores este periacuteodo puede variar
en funcioacuten de la zona geograacutefica entre 12 y 20 meses en el Mediterraacuteneo (Mangold y
Boletzky 1973) y de 18 a 24 meses en la costa NW de Aacutefrica (Hatanaka 1979) y NE
Atlaacutentico (Smale y Buchan 1981) Los machos suelen presentar mayor longevidad que
las hembras ya que eacutestas al final del periodo de incubacioacuten de los huevos acaban por
perecer La eacutepoca reproductiva de O vulgaris es bastante amplia y variacutea con la zona
geograacutefica En el Mediterraacuteneo Mangold y Boletzky (1973) observaron hembras con
Figura 1 ndash Ejemplar adulto delpulpo comuacuten (Octopus vulgaris)
3
puestas de marzo a octubre mientras que machos maduros se encontraban a lo largo
de todo el antildeo Sin embargo dependiendo de la zona geograacutefica de donde provienen
se han identificado dos picos de puestas al antildeo el primero corresponde a la eacutepoca
marzo-mayo maacutes importante en el AtlaacutenticoMediterraacuteneo y la segunda al comienzo
del otontildeo (septiembre-octubre) maacutes importante en Japoacuten (FAO 2003)
El intereacutes comercial del desarrollo del cultivo del pulpo se debe a que esta especie es
muy demandada en varios paiacuteses de Europa y Asia ademaacutes de un elevado valor
comercial Presenta por otra parte algunas caracteriacutesticas bioloacutegicas muy
interesantes para ser considerado un serio candidato para la acuicultura ciclo de vida
corto (1-2 antildeos) elevadas tasas de crecimiento (entre el 10 y el 115 peso corporal
dia-1 a lo largo de toda su vida) alta tasa de conversioacuten alimentaria (30-60) elevada
fertilidad y muy alta viabilidad de los huevos faacutecil adaptacioacuten a la cautividad
aceptacioacuten de alimentos de bajo valor comercial elevado contenido proteico (70-85
del peso seco) y por uacuteltimo praacutecticamente ausencia de patologiacuteas (Mangold y
Boletzky 1973 Mangold 1983 Boucher-Rodoni et al 1987 Lee 1994 Villanueva
1995 Villanueva et al 1995 Iglesias et al 2000 2007 Vaz-Pires et al 2004) El
ciclo de vida del pulpo comuacuten se representa en la figura 2 El pulpo es una especie
gonocoacuterica es decir tiene los sexos separados y aunque no se aprecian diferencias
considerables de tamantildeo y peso corporal entre machos y hembras presentan un claro
dimorfismo sexual a partir del comienzo de la maduracioacuten sexual El aparato
reproductor de los machos posee un uacutenico testiacuteculo localizado en la parte anterior del
cuerpo donde se producen los espermatozoides que salen por el conducto seminal
deferente que a su vez se conecta con una serie de glaacutendulas En eacutestas los
espermatozoides producidos son empaquetados y rodeados por membranas dando
lugar a los espermatoacuteforos que una vez completamente formados se almacenan en el
saco espermatofoacuterico o bolsa de Needhan de la cual salen a traveacutes del conducto
seminal aferente y del oacutergano terminal (o pene) La estructura maacutes caracteriacutestica del
macho es el tercer brazo derecho que presenta ciertas modificaciones morfoloacutegicas y
4
que por eso recibe el nombre de hectocoacutetilo (Fig 3) Este brazo que tiene la funcioacuten
de transferirdepositar los espermatoacuteforos en los conductos oviductales de la hembra
posee un surco a lo largo de toda su cara ventral por donde van a deslizar los
espermatoacuteforos en el momento de la coacutepula acabando en una zona lisa y aplanada
(liacutegula) a diferencia de los otros brazos que acaban en punta y con ventosas
AdultosSexos separados
Desarrollo embrionarioasymp 25-135 diacuteas
Fase post-eclosioacuten paralarvasEtapa de vida planctoacutenicaasymp 30-60 diacuteas
JuvenilesEtapa bentoacutenica
Juveniles hasta sub-adulto
Figura 2 - Esquema del ciclo de vida del pulpo Octopus vulgaris
Otra de las diferencias morfoloacutegicas entre machos y hembras en la edad adulta y
quizaacutes la maacutes faacutecil de identificar es que los machos poseen unas pocas ventosas de
tamantildeo destacado en los segundos y terceros pares de brazos mientras que en las
hembras las ventosas son de tamantildeo maacutes uniforme a lo largo de todo el brazo
Algunos autores apuntan a que estas grandes ventosas sirven para reconocimiento
sexual en el medio natural antes de que se inicie cualquier tipo de actividad
copulatoria (Hanlon y Messenger 1996)
5
Surc
o es
perm
atof
oacuteric
o
Liacutegu
la
glaacutendula oviductal nordm del brazo
liacutegula
hectocoacutetilo
A B
Macho
Hembra
Figura 3 ndash (A) Esquema de la transferencia de los espermatoacuteforos a traveacutes del hectocoacutetilo
hasta los conductos oviductales (DO) de la hembra (B) Aspecto detallado de la regioacuten distal
del brazo hectocotilizado de Octopus vulgaris mostrando la liacutegula
El sistema reproductor de las hembras consta de un ovario que desemboca en dos
oviductos y de las glaacutendulas oviductales (Fig 3) El ovario se situacutea en la parte
posterior de la cavidad del manto y en eacutel se forman los ovocitos Cuando la hembra es
fecundada los espermatoacuteforos se quedan almacenados en los receptaacuteculos seminales
de las glaacutendulas oviductales Se desconoce o al menos no hay evidencias claras de
que los pulpos tengan un comportamiento de cortejo habiendo sido descritas
situaciones de muacuteltiple transferencia de espermatoacuteforos por varios machos a una sola
hembra (Hanlon y Messenger 1996) Sin embargo hasta la fecha de hoy no hay
evidencias cientiacuteficas de la existencia de paternidad muacuteltiple para esta especie
desconocieacutendose por completo las implicaciones geneacuteticas y poblacionales que dicho
suceso pueda tener Se ha postulado ademaacutes que el uacuteltimo macho en copular con una
determinada hembra seriacutea el uacutenico padre de la progenie de la misma ya que eacuteste se
encargariacutea de eliminar los espermatoacuteforos dejados por otros machos precedentes Las
6
hembras son semelparas (se reproducen una sola vez) desarrollaacutendose el ovario de
forma sincroacutenica La ovulacioacuten se da de una sola vez no habiendo desarrollo y
maduracioacuten de nuevos ovocitos despueacutes de la uacutenica puesta estimada en 100000 a
500000 huevos (Mangold 1987) La fecundacioacuten en los pulpos es interna y se da a
medida que los ovocitos van pasando por el oviducto proximal hasta la zona distal
fertilizaacutendose a su paso por la glaacutendula oviductal donde estaacuten alojados los
espermatoacuteforos La puesta de los huevos puede tardar entre 15 y 30 diacuteas
Posteriormente la hembra pasa a ejercer cuidados maternales como proteccioacuten contra
depredadores y limpieza y oxigenacioacuten de los huevos pudiendo este periodo
prolongarse de 25 a 125 diacuteas dependiendo de la temperatura del agua (Mangold y
Boletzky 1973) Durante este tiempo los huevos de pulpo han de pasar por varias
fases de desarrollo (Fig 4) las cuales han sido clasificadas en XX estadios por Naef
(1928) A lo largo de este periodo las hembras dejan de alimentarse pudiendo perder
hasta el 60 de su peso corporal inicial antes de haber iniciado la puesta (Wodinsky
1978) La eclosioacuten de los huevos libera las paralarvas (Fig 4) teacutermino propuesto por
Young y Harman (1988) que asiacute se denominan porque en realidad no van a sufrir una
verdadera metamorfosis antes de convertirse en juveniles aunque al contrario que en
la vida adulta son planctoacutenicas en sus primeros diacuteas de vida Una vez finalizado el
periodo de incubacioacuten y liberacioacuten de las paralarvas las hembras mueren El tamantildeo
del manto de las paralarvas recieacuten eclosionadas es de alrededor de 2 mm y en cada
brazo poseen 3 ventosas (Boletzky 1987 Villanueva 1995) Se estima que el periodo
de vida planctoacutenico de las paralarvas es de 30 a 60 diacuteas dependiendo de la
temperatura del agua y de la zona geograacutefica (Itami et al 1963 Mangold y Boletzky
1973 Villanueva et al 1995) Se cree que las paralarvas de pulpo al igual que otras
larvas de cefaloacutepodos estudiadas en su medio natural (Vecchione 1987 Passarella y
Hopkins 1991) se alimentan de pequentildeos crustaacuteceos que forman parte del
zooplancton Estas criaturas son depredadoras activas desde el primer diacutea de vida
(Iglesias et al 2006) a pesar de que poseen reservas vitelinas internas que les
7
permiten sobrevivir en ausencia de alimento durante algunos diacuteas (Boletzky 1975) La
absorcioacuten de determinados nutrientes por la piel tambieacuten ha sido sentildealada como un
factor importante en los primeros estadios de vida de las larvas de cefaloacutepodos (Lee
1994) mientras que en otras especies de pulpos como el Octopus dofleini se ha
descrito un comportamiento de alimentacioacuten neustoacutenico es decir las paralarvas se
colocan con los brazos haciacutea la superficie aprovechando la tensioacuten superficial del
agua alimentaacutendose de los detritos orgaacutenicos o presas de esta interfase (Marliave
1981)
A
DC
B
Figura 4 - Fotografiacuteas a la lupa de huevos de pulpo en diferentes estadios de desarrollo y de
una paralarva de pulpo (A) huevos de pulpo (asymp 2 x 1 mm) recieacuten depositados (B) huevos en
una etapa de desarrollo intermedia (estadio X-XII) (C) huevos en un estado de desarrollo
avanzado (estadio XVIII) (D) paralarva de pulpo recieacuten eclosionada (tamantildeo total asymp 3 mm)
8
De los trabajos realizados en laboratorio sobre cultivo de paralarvas de O vulgaris
(Villanueva 1995 Iglesias et al 2004 Carrasco et al 2006) se sabe que a medida
que eacutestas van creciendo la proporcioacuten de los brazos frente al manto va aumentando y
una vez alcanzado un determinado tamantildeo realizan el asentamiento en el fondo
convirtieacutendose asiacute en juveniles bentoacutenicos de morfologiacutea igual que los adultos El
crecimiento de los juveniles hasta la fase adulta puede tardar de 8 a 15 meses
dependiendo mayormente de la temperatura y de la disponibilidad de alimento
(Mangold y Boletzky 1973 Forsythe y Van Heukelem 1987 Semmens et al 2004
Leporati et al 2007) Al igual que muchos otros cefaloacutepodos O vulgaris es una
especie carniacutevora durante todo su ciclo de vida En las fases de juvenil hasta la edad
adulta los pulpos se alimentan de una gran variedad de presas que estaacuten
representadas por la mayor parte de los fila marinos (Mangold 1983) de los cuales
varias especies de crustaacuteceos peces y otros moluscos constituyen las presas maacutes
comunes siendo los cangrejos una de sus favoritas (Nixon 1987) Esta especie
presenta ademaacutes un comportamiento de canibalismo frecuente en el medio natural A
pesar del gran intereacutes que hay en el cultivo industrial de esta especie todaviacutea persiste
un importante cuello de botella por resolver el cultivo de las paralarvas planctoacutenicas
hasta la fase de asentamiento cuando eacutestas se convierten en juveniles bentoacutenicos
12 El principal cuello de botella del cultivo del pulpo (Octopus vulgaris)
Actualmente el principal problema que impide el desarrollo integral del cultivo del pulpo
es la elevada mortalidad que se observa en la fase planctoacutenica A pesar de los intentos
llevados a cabo por diferentes grupos de trabajo de varios paiacuteses en los que se han
utilizado distintas metodologiacuteas de alimentacioacuten y condiciones zooteacutecnicas pocos
fueron los experimentos en los que se ha logrado obtener juveniles bentoacutenicos
(revisado por Iglesias et al 2007) Algunos de los primeros trabajos llevados a cabo a
9
nivel mundial se realizaron en Japoacuten Itami et al (1963) utilizando zoeas de camaroacuten
(Palaemon serrifer) lograron tener pulpos juveniles al cabo de 45 diacuteas con una
supervivencia del 8 Hamazaki et al (1991) publicaba igualmente la obtencioacuten de
juveniles de pulpo bentoacutenicos en condiciones controladas empleando como alimento
vivo juveniles de Artemia (15-2 mm) enriquecidos con Nannochloropsis sp y
antildeadiendo esta misma microalga al tanque de cultivo alcanzando en uno de sus
experimentos en un tanque de 20 m3 un nuacutemero de 23700 juveniles bentoacutenicos
(supervivencia del 29) al cabo de 25 diacuteas de cultivo
Al comienzo de los antildeos 90 se iniciaron tambieacuten en Espantildea un conjunto de
importantes trabajos relacionados con el cultivo del pulpo O vulgaris Los primeros
trabajos publicados sobre el cultivo larvario de cefaloacutepodos en laboratorio han sido
desarrollados por Villanueva (1994 1995) que probando zoeas de distintos
crustaacuteceos decaacutepodos (Pagurus prideaux Liocarcinus depurator y Dardanus arrosor)
logroacute obtener pulpos bentoacutenicos confirmando a las zoeas como un alimento apropiado
para suministrar a larvas de cefaloacutepodos Sin embargo este autor ya poniacutea de
manifiesto la dificultad en obtener zoeas en momentos precisos y los altos costes de
mantenimiento y mano de obra que implicaba esta praacutectica Por otro lado se iniciaban
tambieacuten en Espantildea un conjunto de experimentos de engorde de pulpo a escala de
laboratorio para evaluar el potencial de crecimiento de esta especie afrontaacutendose
entre los antildeos 1997 y 2000 el problema del cultivo de la fase planctoacutenica En los
primeros ensayos realizados Iglesias et al (1999 2000) probaron distintos tipos de
alimento tales como zooplancton del medio natural (copeacutepodos zoeas de crustaacuteceos
y misidaacuteceos) ictioplancton huevos de peces micropellets Artemia y rotiacuteferos y maacutes
tarde zoeas de crustaacuteceos obtenidas en laboratorio a partir de stocks de reproductores
de cangrejo (Carcinus maenas) neacutecora (Necora puber) y camaroacuten (Palaemon
serratus) Sin embargo la mortalidad de las paralarvas fue casi total en los primeros
diacuteas de vida alcanzaacutendose supervivencias maacuteximas del 10 a los 32 diacuteas al emplear
como alimento metanauplios y a continuacioacuten Artemia adulta enriquecida con
10
microalgas pero sin que se lograse alcanzar la fase de pre-asentamiento de las
paralarvas En los intentos llevados a cabo en el IEO de Vigo y maacutes tarde en el Centro
de Experimentacioacuten Pesquera de Asturias se obtuvieron algunos ejemplares de
juveniles bentoacutenicos que han llegado a alcanzar la edad adulta utilizaacutendose como
dieta para suministrar a las paralarvas juveniles de Artemia enriquecidos con
microalgas complementados con zoeas de centolla Maja brachydactyla en momentos
de disponibilidad de eacutestas (Iglesias et al 2002 2004 Carrasco et al 2003 2006) Las
primeras pruebas en las que se ha empleado esta mezcla de presas vivas (Moxica et
al 2002) fueron llevadas a cabo en el IEO de Vigo en un tanque de 9 m3 al que se
aportoacute inicialmente nauplios de Artemia y maacutes tarde juveniles de Artemia (1-4 mm) a
una densidad de presas de 01 Artemia ml-1 complementados con zoeas a partir de la
tercera semana de cultivo (3000 a 5000 zoeas al diacutea) Al tanque se aportaba a diario
una mezcla de microalgas (un 40 de Isochrysis galbana un 40 de Tetraselmis
suecica y un 20 de Chaetoceros sp) con el fin de mantener las presas vivas
constantemente enriquecidas Aunque no se han obtenido pulpos bentoacutenicos en este
experimento los autores refirieron la obtencioacuten de paralarvas en fase de asentamiento
con elevado nuacutemero de ventosas en los brazos (17-18) y un peso seco de 92 plusmn 09
mg a los 52 diacuteas de vida La supervivencia en este experimento fue de 83 al mes de
vida y de 02 al cabo de 52 diacuteas En un experimento posterior en el que se redujo la
escala y se empleoacute un tanque de 1 m3 las paralarvas se alimentaron con Artemia (1-4
mm) enriquecida con Chlorella sp antildeadiendo diariamente al tanque de cultivo una
mezcla de microalgas (Chlorella sp Isochrysis galbana y Chaetoceros sp) Como
complemento de Artemia se suministraron zoeas de centollo (Maja brachydactyla)
cuatro veces a la semana a una concentracioacuten de 001-01 zoeas ml-1 alcanzaacutendose
asiacute una supervivencia del 315 al diacutea 40 y un reducido nuacutemero de juveniles
bentoacutenicos dos semanas maacutes tarde (Iglesias et al 2002 2004) El seguimiento de los
uacutenicos pulpos supervivientes de este experimento hasta la fase adulta y la obtencioacuten
de una puesta de huevos por parte de una hembra originoacute el cierre del ciclo de vida
11
del pulpo en laboratorio por primer vez (Iglesias et al 2004) De forma similar
Carrasco et al (2003 2006) lograron obtener en uno de sus experimentos pulpos
bentoacutenicos a los 60 diacuteas con una supervivencia del 34
Sin embargo la utilizacioacuten de zoeas de crustaacuteceos como alimento vivo supone un
elevado riesgo ya que no se controla la obtencioacuten de las mismas en momentos
precisos ni la cantidad a producir ademaacutes de los elevados costes que conlleva esta
praacutectica por la necesidad de disponer de maacutes recursos materiales fiacutesicos y de
personal (Navarro y Villanueva 2000) Artemia en cambio es un crustaacuteceo faacutecil de
producir los nauplios o metanauplios se obtienen en 24-72 h y su composicioacuten
bioquiacutemica se puede modular mediante teacutecnicas de enriquecimiento al ser un filtrador
continuo obligado y no selectivo La utilizacioacuten de Artemia como presa viva se
consolida por tanto como una de las mejores alternativas para el cultivo de
paralarvas aunque sea necesario seguir trabajando en la modulacioacuten de su valor
nutricional de acuerdo con los posibles requerimientos de las paralarvas
Varios autores han utilizado nauplios de Artemia enriquecidos con distintos productos
comerciales (Navarro y Villanueva 2000 2003 Villanueva et al 2004 Okumura et al
2005) en sus experimentos de cultivo de paralarvas de pulpo siendo el tamantildeo de
estas presas (05-12 mm) adecuado hasta determinado momento del cultivo Sin
embargo se ha demostrado que las paralarvas tienen una preferencia clara por
Artemia de mayor tamantildeo (14 mm) sobre la maacutes pequentildea (08 mm) desde los
primeros diacuteas de vida (Iglesias et al 2006) De entre los productos o suplementos
nutricionales empleados por diversos autores para enriquecer Artemia se encuentran
emulsiones lipiacutedicas de la gama Selco (INVE) u otros aceites de pescado vitaminas
harina de huevos de pescado (BASF) harinas de cereales o productos de la gama Ori
(Skretting) (Navarro y Villanueva 2000 2003 Villanueva et al 2002 Iglesias et al
2004 Okumura et al 2005 Izquierdo et al 2008) pero ninguno ha originado tasas de
supervivencia o pesos secos de las paralarvas semejantes a los obtenidos con la
utilizacioacuten de zoeas de crustaacuteceos Como alternativa a las zoeas o a la Artemia
12
tambieacuten se han probado otros tipos de presas vivas o dietas artificiales para
suministrar a las paralarvas aunque en todo caso este tipo de pruebas se hicieron
complementadas con Artemia enriquecida (revisado por Iglesias et al 2007)
Otro de los principales suplementos empleados muy a menudo por diversos autores
para enriquecercultivar Artemia son las microalgas de las que las maacutes comuacutenmente
utilizadas fueron Nannochloropsis sp Dunaliella viridis Tetraselmis suecica
Isochrysis galbana Chaetoceros sp Chlorella sp (Hamazaki et al 1991 Navarro y
Villanueva 2000 Moxica et al 2002 Iglesias et al 2002 2004 Carrasco et al
2003 2006 Moxica et al 2006)
Hay un cierto consenso entre los grupos de investigadores que se dedican al cultivo
larvario del pulpo sobre que los aspectos nutricionales estaacuten en la base de la elevada
mortalidad observada durante su cultivo (Iglesias et al 2007) Uno de los problemas
que dificulta la interpretacioacuten de los resultados descritos y el avance en la nutricioacuten del
cultivo larvario del pulpo es que muy pocos autores presentan datos de la
composicioacuten bioquiacutemica de las dietas utilizadas y de sus efectos sobre la composicioacuten
de las paralarvas (Moxica et al 2002 Navarro et al 2000 2003 Villanueva et al
2004 Okumura et al 2005) cuando esta informacioacuten es crucial para interpretar los
resultados obtenidos y asiacute poder relacionarlos con la supervivencia y el crecimiento de
las paralarvas
A pesar de los continuos intentos para cerrar el ciclo de vida del pulpo de forma
rentable la mortalidad de las paralarvas durante la fase planctoacutenica sigue siendo casi
total Entre las estrategias propuestas para superar este importante problema estaacuten la
mejora de la composicioacuten de Artemia mediante las teacutecnicas de enriquecimiento
apropiadas y la formulacioacuten de microdietas inertes (Iglesias et al 2007) Importa
pues reducir el tiempo necesario hasta la obtencioacuten de los juveniles de Artemia del
tamantildeo deseado y optimizar su composicioacuten bioquiacutemica con base en los posibles
requerimientos nutricionales de las paralarvas El crecimiento y la composicioacuten
bioquiacutemica de Artemia sp pueden ser fuertemente modificados mediante la utilizacioacuten
13
de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

RESUMEN
El pulpo comuacuten (Octopus vulgaris) es una especie de alto valor comercial en Galicia y
de elevado intereacutes cultural y gastronoacutemico En las uacuteltimas deacutecadas la demanda de
pulpo superoacute largamente la capacidad pesquera siendo necesario recurrir a
importaciones En los antildeos 90 se iniciaron una serie de trabajos de cultivo de pulpo en
cautividad cuyo objetivo era analiziar su potencial para la acuicultura Eacutestos
experimentos confirmaron el potencial de esta especie debido a sus interesantes
caracteriacutesticas bioloacutegicas raacutepido crecimiento y ciclo de vida corto alta eficacia de
conversioacuten del alimento elevada fecundidad resistencia a la manipulacioacuten y
adaptacioacuten a la vida en cautividad Aunque los resultados de engorde eran
prometedores y rentables sigue persistiendo un importante cuello de botella en su
cultivo integral el cultivo de las paralarvas planctoacutenicas Por ello el presente trabajo
tuvo como principales objetivos estudiar la hipoacutetesis de paternidad muacuteltiple en las
puestas de pulpo y sus consecuencias a nivel de diversificacioacuten geneacutetica y optimizar
el valor nutricional del alimento vivo para el cultivo de paralarvas evaluaacutendose el
efecto de diferentes dietas sobre el crecimiento y la supervivencia Se ha demostrado
la existencia de muacuteltiple paternidad en O vulgaris Eacutesta puede representar un proceso
que maximiza la recombinacioacuten geneacutetica de la hembra con los varios machos en un
uacutenico acontecimiento reproductivo Por otro lado se ha demostrado que el
enriquecimiento de Artemia con microalgas y aminoaacutecidos esenciales (lisina arginina y
metionina) origina una mejora del peso seco y supervivencia de las paralarvas a los
25 diacuteas de vida El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del
pulpo y de la nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo
integral de esta especie pudiendo igualmente ser uacutetil en el futuro para hacer una
gestioacuten sostenible de las poblaciones naturales
Palabras clave pulpo repoblacioacuten cultivo sostenibilidad geneacutetica diversidad
2
1 Introduccioacuten
11 Biologiacutea y ciclo de vida del pulpo comuacuten (Octopus vulgaris)
El pulpo comuacuten (Octopus vulgaris Cuvier 1797)
tambieacuten denominado pulpo de roca (Fig 1) es
uno de los moluscos cefaloacutepodos mejor
estudiados en todo el mundo destacaacutendose entre
las maacutes de cien especies descritas del geacutenero
Octopus Esta especie ha sido modelo de
investigacioacuten en diversos estudios que incluyen
aacutereas como la neurobiologiacutea la fisiologiacutea animal
bioquiacutemica y comportamiento animal (Young
1971 Wells 1978 Boyle 1983) En los uacuteltimos
15 antildeos se ha enfatizado ademaacutes el estudio de su biologiacutea y ciclo de vida en
cautividad con el objetivo de aplicar estos conocimientos a su cultivo integral (Iglesias
et al 2007) El pulpo es un animal estrictamente marino que se distribuye en aguas
templadas y tropicales de todo el mundo Vive solitario es territorial y realiza
migraciones estacionales acercaacutendose en primavera a zonas maacutes someras para
reproducirse Su ciclo de vida es relativamente corto estimaacutendose entre uno y dos
antildeos en el medio natural aunque seguacuten diferentes autores este periacuteodo puede variar
en funcioacuten de la zona geograacutefica entre 12 y 20 meses en el Mediterraacuteneo (Mangold y
Boletzky 1973) y de 18 a 24 meses en la costa NW de Aacutefrica (Hatanaka 1979) y NE
Atlaacutentico (Smale y Buchan 1981) Los machos suelen presentar mayor longevidad que
las hembras ya que eacutestas al final del periodo de incubacioacuten de los huevos acaban por
perecer La eacutepoca reproductiva de O vulgaris es bastante amplia y variacutea con la zona
geograacutefica En el Mediterraacuteneo Mangold y Boletzky (1973) observaron hembras con
Figura 1 ndash Ejemplar adulto delpulpo comuacuten (Octopus vulgaris)
3
puestas de marzo a octubre mientras que machos maduros se encontraban a lo largo
de todo el antildeo Sin embargo dependiendo de la zona geograacutefica de donde provienen
se han identificado dos picos de puestas al antildeo el primero corresponde a la eacutepoca
marzo-mayo maacutes importante en el AtlaacutenticoMediterraacuteneo y la segunda al comienzo
del otontildeo (septiembre-octubre) maacutes importante en Japoacuten (FAO 2003)
El intereacutes comercial del desarrollo del cultivo del pulpo se debe a que esta especie es
muy demandada en varios paiacuteses de Europa y Asia ademaacutes de un elevado valor
comercial Presenta por otra parte algunas caracteriacutesticas bioloacutegicas muy
interesantes para ser considerado un serio candidato para la acuicultura ciclo de vida
corto (1-2 antildeos) elevadas tasas de crecimiento (entre el 10 y el 115 peso corporal
dia-1 a lo largo de toda su vida) alta tasa de conversioacuten alimentaria (30-60) elevada
fertilidad y muy alta viabilidad de los huevos faacutecil adaptacioacuten a la cautividad
aceptacioacuten de alimentos de bajo valor comercial elevado contenido proteico (70-85
del peso seco) y por uacuteltimo praacutecticamente ausencia de patologiacuteas (Mangold y
Boletzky 1973 Mangold 1983 Boucher-Rodoni et al 1987 Lee 1994 Villanueva
1995 Villanueva et al 1995 Iglesias et al 2000 2007 Vaz-Pires et al 2004) El
ciclo de vida del pulpo comuacuten se representa en la figura 2 El pulpo es una especie
gonocoacuterica es decir tiene los sexos separados y aunque no se aprecian diferencias
considerables de tamantildeo y peso corporal entre machos y hembras presentan un claro
dimorfismo sexual a partir del comienzo de la maduracioacuten sexual El aparato
reproductor de los machos posee un uacutenico testiacuteculo localizado en la parte anterior del
cuerpo donde se producen los espermatozoides que salen por el conducto seminal
deferente que a su vez se conecta con una serie de glaacutendulas En eacutestas los
espermatozoides producidos son empaquetados y rodeados por membranas dando
lugar a los espermatoacuteforos que una vez completamente formados se almacenan en el
saco espermatofoacuterico o bolsa de Needhan de la cual salen a traveacutes del conducto
seminal aferente y del oacutergano terminal (o pene) La estructura maacutes caracteriacutestica del
macho es el tercer brazo derecho que presenta ciertas modificaciones morfoloacutegicas y
4
que por eso recibe el nombre de hectocoacutetilo (Fig 3) Este brazo que tiene la funcioacuten
de transferirdepositar los espermatoacuteforos en los conductos oviductales de la hembra
posee un surco a lo largo de toda su cara ventral por donde van a deslizar los
espermatoacuteforos en el momento de la coacutepula acabando en una zona lisa y aplanada
(liacutegula) a diferencia de los otros brazos que acaban en punta y con ventosas
AdultosSexos separados
Desarrollo embrionarioasymp 25-135 diacuteas
Fase post-eclosioacuten paralarvasEtapa de vida planctoacutenicaasymp 30-60 diacuteas
JuvenilesEtapa bentoacutenica
Juveniles hasta sub-adulto
Figura 2 - Esquema del ciclo de vida del pulpo Octopus vulgaris
Otra de las diferencias morfoloacutegicas entre machos y hembras en la edad adulta y
quizaacutes la maacutes faacutecil de identificar es que los machos poseen unas pocas ventosas de
tamantildeo destacado en los segundos y terceros pares de brazos mientras que en las
hembras las ventosas son de tamantildeo maacutes uniforme a lo largo de todo el brazo
Algunos autores apuntan a que estas grandes ventosas sirven para reconocimiento
sexual en el medio natural antes de que se inicie cualquier tipo de actividad
copulatoria (Hanlon y Messenger 1996)
5
Surc
o es
perm
atof
oacuteric
o
Liacutegu
la
glaacutendula oviductal nordm del brazo
liacutegula
hectocoacutetilo
A B
Macho
Hembra
Figura 3 ndash (A) Esquema de la transferencia de los espermatoacuteforos a traveacutes del hectocoacutetilo
hasta los conductos oviductales (DO) de la hembra (B) Aspecto detallado de la regioacuten distal
del brazo hectocotilizado de Octopus vulgaris mostrando la liacutegula
El sistema reproductor de las hembras consta de un ovario que desemboca en dos
oviductos y de las glaacutendulas oviductales (Fig 3) El ovario se situacutea en la parte
posterior de la cavidad del manto y en eacutel se forman los ovocitos Cuando la hembra es
fecundada los espermatoacuteforos se quedan almacenados en los receptaacuteculos seminales
de las glaacutendulas oviductales Se desconoce o al menos no hay evidencias claras de
que los pulpos tengan un comportamiento de cortejo habiendo sido descritas
situaciones de muacuteltiple transferencia de espermatoacuteforos por varios machos a una sola
hembra (Hanlon y Messenger 1996) Sin embargo hasta la fecha de hoy no hay
evidencias cientiacuteficas de la existencia de paternidad muacuteltiple para esta especie
desconocieacutendose por completo las implicaciones geneacuteticas y poblacionales que dicho
suceso pueda tener Se ha postulado ademaacutes que el uacuteltimo macho en copular con una
determinada hembra seriacutea el uacutenico padre de la progenie de la misma ya que eacuteste se
encargariacutea de eliminar los espermatoacuteforos dejados por otros machos precedentes Las
6
hembras son semelparas (se reproducen una sola vez) desarrollaacutendose el ovario de
forma sincroacutenica La ovulacioacuten se da de una sola vez no habiendo desarrollo y
maduracioacuten de nuevos ovocitos despueacutes de la uacutenica puesta estimada en 100000 a
500000 huevos (Mangold 1987) La fecundacioacuten en los pulpos es interna y se da a
medida que los ovocitos van pasando por el oviducto proximal hasta la zona distal
fertilizaacutendose a su paso por la glaacutendula oviductal donde estaacuten alojados los
espermatoacuteforos La puesta de los huevos puede tardar entre 15 y 30 diacuteas
Posteriormente la hembra pasa a ejercer cuidados maternales como proteccioacuten contra
depredadores y limpieza y oxigenacioacuten de los huevos pudiendo este periodo
prolongarse de 25 a 125 diacuteas dependiendo de la temperatura del agua (Mangold y
Boletzky 1973) Durante este tiempo los huevos de pulpo han de pasar por varias
fases de desarrollo (Fig 4) las cuales han sido clasificadas en XX estadios por Naef
(1928) A lo largo de este periodo las hembras dejan de alimentarse pudiendo perder
hasta el 60 de su peso corporal inicial antes de haber iniciado la puesta (Wodinsky
1978) La eclosioacuten de los huevos libera las paralarvas (Fig 4) teacutermino propuesto por
Young y Harman (1988) que asiacute se denominan porque en realidad no van a sufrir una
verdadera metamorfosis antes de convertirse en juveniles aunque al contrario que en
la vida adulta son planctoacutenicas en sus primeros diacuteas de vida Una vez finalizado el
periodo de incubacioacuten y liberacioacuten de las paralarvas las hembras mueren El tamantildeo
del manto de las paralarvas recieacuten eclosionadas es de alrededor de 2 mm y en cada
brazo poseen 3 ventosas (Boletzky 1987 Villanueva 1995) Se estima que el periodo
de vida planctoacutenico de las paralarvas es de 30 a 60 diacuteas dependiendo de la
temperatura del agua y de la zona geograacutefica (Itami et al 1963 Mangold y Boletzky
1973 Villanueva et al 1995) Se cree que las paralarvas de pulpo al igual que otras
larvas de cefaloacutepodos estudiadas en su medio natural (Vecchione 1987 Passarella y
Hopkins 1991) se alimentan de pequentildeos crustaacuteceos que forman parte del
zooplancton Estas criaturas son depredadoras activas desde el primer diacutea de vida
(Iglesias et al 2006) a pesar de que poseen reservas vitelinas internas que les
7
permiten sobrevivir en ausencia de alimento durante algunos diacuteas (Boletzky 1975) La
absorcioacuten de determinados nutrientes por la piel tambieacuten ha sido sentildealada como un
factor importante en los primeros estadios de vida de las larvas de cefaloacutepodos (Lee
1994) mientras que en otras especies de pulpos como el Octopus dofleini se ha
descrito un comportamiento de alimentacioacuten neustoacutenico es decir las paralarvas se
colocan con los brazos haciacutea la superficie aprovechando la tensioacuten superficial del
agua alimentaacutendose de los detritos orgaacutenicos o presas de esta interfase (Marliave
1981)
A
DC
B
Figura 4 - Fotografiacuteas a la lupa de huevos de pulpo en diferentes estadios de desarrollo y de
una paralarva de pulpo (A) huevos de pulpo (asymp 2 x 1 mm) recieacuten depositados (B) huevos en
una etapa de desarrollo intermedia (estadio X-XII) (C) huevos en un estado de desarrollo
avanzado (estadio XVIII) (D) paralarva de pulpo recieacuten eclosionada (tamantildeo total asymp 3 mm)
8
De los trabajos realizados en laboratorio sobre cultivo de paralarvas de O vulgaris
(Villanueva 1995 Iglesias et al 2004 Carrasco et al 2006) se sabe que a medida
que eacutestas van creciendo la proporcioacuten de los brazos frente al manto va aumentando y
una vez alcanzado un determinado tamantildeo realizan el asentamiento en el fondo
convirtieacutendose asiacute en juveniles bentoacutenicos de morfologiacutea igual que los adultos El
crecimiento de los juveniles hasta la fase adulta puede tardar de 8 a 15 meses
dependiendo mayormente de la temperatura y de la disponibilidad de alimento
(Mangold y Boletzky 1973 Forsythe y Van Heukelem 1987 Semmens et al 2004
Leporati et al 2007) Al igual que muchos otros cefaloacutepodos O vulgaris es una
especie carniacutevora durante todo su ciclo de vida En las fases de juvenil hasta la edad
adulta los pulpos se alimentan de una gran variedad de presas que estaacuten
representadas por la mayor parte de los fila marinos (Mangold 1983) de los cuales
varias especies de crustaacuteceos peces y otros moluscos constituyen las presas maacutes
comunes siendo los cangrejos una de sus favoritas (Nixon 1987) Esta especie
presenta ademaacutes un comportamiento de canibalismo frecuente en el medio natural A
pesar del gran intereacutes que hay en el cultivo industrial de esta especie todaviacutea persiste
un importante cuello de botella por resolver el cultivo de las paralarvas planctoacutenicas
hasta la fase de asentamiento cuando eacutestas se convierten en juveniles bentoacutenicos
12 El principal cuello de botella del cultivo del pulpo (Octopus vulgaris)
Actualmente el principal problema que impide el desarrollo integral del cultivo del pulpo
es la elevada mortalidad que se observa en la fase planctoacutenica A pesar de los intentos
llevados a cabo por diferentes grupos de trabajo de varios paiacuteses en los que se han
utilizado distintas metodologiacuteas de alimentacioacuten y condiciones zooteacutecnicas pocos
fueron los experimentos en los que se ha logrado obtener juveniles bentoacutenicos
(revisado por Iglesias et al 2007) Algunos de los primeros trabajos llevados a cabo a
9
nivel mundial se realizaron en Japoacuten Itami et al (1963) utilizando zoeas de camaroacuten
(Palaemon serrifer) lograron tener pulpos juveniles al cabo de 45 diacuteas con una
supervivencia del 8 Hamazaki et al (1991) publicaba igualmente la obtencioacuten de
juveniles de pulpo bentoacutenicos en condiciones controladas empleando como alimento
vivo juveniles de Artemia (15-2 mm) enriquecidos con Nannochloropsis sp y
antildeadiendo esta misma microalga al tanque de cultivo alcanzando en uno de sus
experimentos en un tanque de 20 m3 un nuacutemero de 23700 juveniles bentoacutenicos
(supervivencia del 29) al cabo de 25 diacuteas de cultivo
Al comienzo de los antildeos 90 se iniciaron tambieacuten en Espantildea un conjunto de
importantes trabajos relacionados con el cultivo del pulpo O vulgaris Los primeros
trabajos publicados sobre el cultivo larvario de cefaloacutepodos en laboratorio han sido
desarrollados por Villanueva (1994 1995) que probando zoeas de distintos
crustaacuteceos decaacutepodos (Pagurus prideaux Liocarcinus depurator y Dardanus arrosor)
logroacute obtener pulpos bentoacutenicos confirmando a las zoeas como un alimento apropiado
para suministrar a larvas de cefaloacutepodos Sin embargo este autor ya poniacutea de
manifiesto la dificultad en obtener zoeas en momentos precisos y los altos costes de
mantenimiento y mano de obra que implicaba esta praacutectica Por otro lado se iniciaban
tambieacuten en Espantildea un conjunto de experimentos de engorde de pulpo a escala de
laboratorio para evaluar el potencial de crecimiento de esta especie afrontaacutendose
entre los antildeos 1997 y 2000 el problema del cultivo de la fase planctoacutenica En los
primeros ensayos realizados Iglesias et al (1999 2000) probaron distintos tipos de
alimento tales como zooplancton del medio natural (copeacutepodos zoeas de crustaacuteceos
y misidaacuteceos) ictioplancton huevos de peces micropellets Artemia y rotiacuteferos y maacutes
tarde zoeas de crustaacuteceos obtenidas en laboratorio a partir de stocks de reproductores
de cangrejo (Carcinus maenas) neacutecora (Necora puber) y camaroacuten (Palaemon
serratus) Sin embargo la mortalidad de las paralarvas fue casi total en los primeros
diacuteas de vida alcanzaacutendose supervivencias maacuteximas del 10 a los 32 diacuteas al emplear
como alimento metanauplios y a continuacioacuten Artemia adulta enriquecida con
10
microalgas pero sin que se lograse alcanzar la fase de pre-asentamiento de las
paralarvas En los intentos llevados a cabo en el IEO de Vigo y maacutes tarde en el Centro
de Experimentacioacuten Pesquera de Asturias se obtuvieron algunos ejemplares de
juveniles bentoacutenicos que han llegado a alcanzar la edad adulta utilizaacutendose como
dieta para suministrar a las paralarvas juveniles de Artemia enriquecidos con
microalgas complementados con zoeas de centolla Maja brachydactyla en momentos
de disponibilidad de eacutestas (Iglesias et al 2002 2004 Carrasco et al 2003 2006) Las
primeras pruebas en las que se ha empleado esta mezcla de presas vivas (Moxica et
al 2002) fueron llevadas a cabo en el IEO de Vigo en un tanque de 9 m3 al que se
aportoacute inicialmente nauplios de Artemia y maacutes tarde juveniles de Artemia (1-4 mm) a
una densidad de presas de 01 Artemia ml-1 complementados con zoeas a partir de la
tercera semana de cultivo (3000 a 5000 zoeas al diacutea) Al tanque se aportaba a diario
una mezcla de microalgas (un 40 de Isochrysis galbana un 40 de Tetraselmis
suecica y un 20 de Chaetoceros sp) con el fin de mantener las presas vivas
constantemente enriquecidas Aunque no se han obtenido pulpos bentoacutenicos en este
experimento los autores refirieron la obtencioacuten de paralarvas en fase de asentamiento
con elevado nuacutemero de ventosas en los brazos (17-18) y un peso seco de 92 plusmn 09
mg a los 52 diacuteas de vida La supervivencia en este experimento fue de 83 al mes de
vida y de 02 al cabo de 52 diacuteas En un experimento posterior en el que se redujo la
escala y se empleoacute un tanque de 1 m3 las paralarvas se alimentaron con Artemia (1-4
mm) enriquecida con Chlorella sp antildeadiendo diariamente al tanque de cultivo una
mezcla de microalgas (Chlorella sp Isochrysis galbana y Chaetoceros sp) Como
complemento de Artemia se suministraron zoeas de centollo (Maja brachydactyla)
cuatro veces a la semana a una concentracioacuten de 001-01 zoeas ml-1 alcanzaacutendose
asiacute una supervivencia del 315 al diacutea 40 y un reducido nuacutemero de juveniles
bentoacutenicos dos semanas maacutes tarde (Iglesias et al 2002 2004) El seguimiento de los
uacutenicos pulpos supervivientes de este experimento hasta la fase adulta y la obtencioacuten
de una puesta de huevos por parte de una hembra originoacute el cierre del ciclo de vida
11
del pulpo en laboratorio por primer vez (Iglesias et al 2004) De forma similar
Carrasco et al (2003 2006) lograron obtener en uno de sus experimentos pulpos
bentoacutenicos a los 60 diacuteas con una supervivencia del 34
Sin embargo la utilizacioacuten de zoeas de crustaacuteceos como alimento vivo supone un
elevado riesgo ya que no se controla la obtencioacuten de las mismas en momentos
precisos ni la cantidad a producir ademaacutes de los elevados costes que conlleva esta
praacutectica por la necesidad de disponer de maacutes recursos materiales fiacutesicos y de
personal (Navarro y Villanueva 2000) Artemia en cambio es un crustaacuteceo faacutecil de
producir los nauplios o metanauplios se obtienen en 24-72 h y su composicioacuten
bioquiacutemica se puede modular mediante teacutecnicas de enriquecimiento al ser un filtrador
continuo obligado y no selectivo La utilizacioacuten de Artemia como presa viva se
consolida por tanto como una de las mejores alternativas para el cultivo de
paralarvas aunque sea necesario seguir trabajando en la modulacioacuten de su valor
nutricional de acuerdo con los posibles requerimientos de las paralarvas
Varios autores han utilizado nauplios de Artemia enriquecidos con distintos productos
comerciales (Navarro y Villanueva 2000 2003 Villanueva et al 2004 Okumura et al
2005) en sus experimentos de cultivo de paralarvas de pulpo siendo el tamantildeo de
estas presas (05-12 mm) adecuado hasta determinado momento del cultivo Sin
embargo se ha demostrado que las paralarvas tienen una preferencia clara por
Artemia de mayor tamantildeo (14 mm) sobre la maacutes pequentildea (08 mm) desde los
primeros diacuteas de vida (Iglesias et al 2006) De entre los productos o suplementos
nutricionales empleados por diversos autores para enriquecer Artemia se encuentran
emulsiones lipiacutedicas de la gama Selco (INVE) u otros aceites de pescado vitaminas
harina de huevos de pescado (BASF) harinas de cereales o productos de la gama Ori
(Skretting) (Navarro y Villanueva 2000 2003 Villanueva et al 2002 Iglesias et al
2004 Okumura et al 2005 Izquierdo et al 2008) pero ninguno ha originado tasas de
supervivencia o pesos secos de las paralarvas semejantes a los obtenidos con la
utilizacioacuten de zoeas de crustaacuteceos Como alternativa a las zoeas o a la Artemia
12
tambieacuten se han probado otros tipos de presas vivas o dietas artificiales para
suministrar a las paralarvas aunque en todo caso este tipo de pruebas se hicieron
complementadas con Artemia enriquecida (revisado por Iglesias et al 2007)
Otro de los principales suplementos empleados muy a menudo por diversos autores
para enriquecercultivar Artemia son las microalgas de las que las maacutes comuacutenmente
utilizadas fueron Nannochloropsis sp Dunaliella viridis Tetraselmis suecica
Isochrysis galbana Chaetoceros sp Chlorella sp (Hamazaki et al 1991 Navarro y
Villanueva 2000 Moxica et al 2002 Iglesias et al 2002 2004 Carrasco et al
2003 2006 Moxica et al 2006)
Hay un cierto consenso entre los grupos de investigadores que se dedican al cultivo
larvario del pulpo sobre que los aspectos nutricionales estaacuten en la base de la elevada
mortalidad observada durante su cultivo (Iglesias et al 2007) Uno de los problemas
que dificulta la interpretacioacuten de los resultados descritos y el avance en la nutricioacuten del
cultivo larvario del pulpo es que muy pocos autores presentan datos de la
composicioacuten bioquiacutemica de las dietas utilizadas y de sus efectos sobre la composicioacuten
de las paralarvas (Moxica et al 2002 Navarro et al 2000 2003 Villanueva et al
2004 Okumura et al 2005) cuando esta informacioacuten es crucial para interpretar los
resultados obtenidos y asiacute poder relacionarlos con la supervivencia y el crecimiento de
las paralarvas
A pesar de los continuos intentos para cerrar el ciclo de vida del pulpo de forma
rentable la mortalidad de las paralarvas durante la fase planctoacutenica sigue siendo casi
total Entre las estrategias propuestas para superar este importante problema estaacuten la
mejora de la composicioacuten de Artemia mediante las teacutecnicas de enriquecimiento
apropiadas y la formulacioacuten de microdietas inertes (Iglesias et al 2007) Importa
pues reducir el tiempo necesario hasta la obtencioacuten de los juveniles de Artemia del
tamantildeo deseado y optimizar su composicioacuten bioquiacutemica con base en los posibles
requerimientos nutricionales de las paralarvas El crecimiento y la composicioacuten
bioquiacutemica de Artemia sp pueden ser fuertemente modificados mediante la utilizacioacuten
13
de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

1 Introduccioacuten
11 Biologiacutea y ciclo de vida del pulpo comuacuten (Octopus vulgaris)
El pulpo comuacuten (Octopus vulgaris Cuvier 1797)
tambieacuten denominado pulpo de roca (Fig 1) es
uno de los moluscos cefaloacutepodos mejor
estudiados en todo el mundo destacaacutendose entre
las maacutes de cien especies descritas del geacutenero
Octopus Esta especie ha sido modelo de
investigacioacuten en diversos estudios que incluyen
aacutereas como la neurobiologiacutea la fisiologiacutea animal
bioquiacutemica y comportamiento animal (Young
1971 Wells 1978 Boyle 1983) En los uacuteltimos
15 antildeos se ha enfatizado ademaacutes el estudio de su biologiacutea y ciclo de vida en
cautividad con el objetivo de aplicar estos conocimientos a su cultivo integral (Iglesias
et al 2007) El pulpo es un animal estrictamente marino que se distribuye en aguas
templadas y tropicales de todo el mundo Vive solitario es territorial y realiza
migraciones estacionales acercaacutendose en primavera a zonas maacutes someras para
reproducirse Su ciclo de vida es relativamente corto estimaacutendose entre uno y dos
antildeos en el medio natural aunque seguacuten diferentes autores este periacuteodo puede variar
en funcioacuten de la zona geograacutefica entre 12 y 20 meses en el Mediterraacuteneo (Mangold y
Boletzky 1973) y de 18 a 24 meses en la costa NW de Aacutefrica (Hatanaka 1979) y NE
Atlaacutentico (Smale y Buchan 1981) Los machos suelen presentar mayor longevidad que
las hembras ya que eacutestas al final del periodo de incubacioacuten de los huevos acaban por
perecer La eacutepoca reproductiva de O vulgaris es bastante amplia y variacutea con la zona
geograacutefica En el Mediterraacuteneo Mangold y Boletzky (1973) observaron hembras con
Figura 1 ndash Ejemplar adulto delpulpo comuacuten (Octopus vulgaris)
3
puestas de marzo a octubre mientras que machos maduros se encontraban a lo largo
de todo el antildeo Sin embargo dependiendo de la zona geograacutefica de donde provienen
se han identificado dos picos de puestas al antildeo el primero corresponde a la eacutepoca
marzo-mayo maacutes importante en el AtlaacutenticoMediterraacuteneo y la segunda al comienzo
del otontildeo (septiembre-octubre) maacutes importante en Japoacuten (FAO 2003)
El intereacutes comercial del desarrollo del cultivo del pulpo se debe a que esta especie es
muy demandada en varios paiacuteses de Europa y Asia ademaacutes de un elevado valor
comercial Presenta por otra parte algunas caracteriacutesticas bioloacutegicas muy
interesantes para ser considerado un serio candidato para la acuicultura ciclo de vida
corto (1-2 antildeos) elevadas tasas de crecimiento (entre el 10 y el 115 peso corporal
dia-1 a lo largo de toda su vida) alta tasa de conversioacuten alimentaria (30-60) elevada
fertilidad y muy alta viabilidad de los huevos faacutecil adaptacioacuten a la cautividad
aceptacioacuten de alimentos de bajo valor comercial elevado contenido proteico (70-85
del peso seco) y por uacuteltimo praacutecticamente ausencia de patologiacuteas (Mangold y
Boletzky 1973 Mangold 1983 Boucher-Rodoni et al 1987 Lee 1994 Villanueva
1995 Villanueva et al 1995 Iglesias et al 2000 2007 Vaz-Pires et al 2004) El
ciclo de vida del pulpo comuacuten se representa en la figura 2 El pulpo es una especie
gonocoacuterica es decir tiene los sexos separados y aunque no se aprecian diferencias
considerables de tamantildeo y peso corporal entre machos y hembras presentan un claro
dimorfismo sexual a partir del comienzo de la maduracioacuten sexual El aparato
reproductor de los machos posee un uacutenico testiacuteculo localizado en la parte anterior del
cuerpo donde se producen los espermatozoides que salen por el conducto seminal
deferente que a su vez se conecta con una serie de glaacutendulas En eacutestas los
espermatozoides producidos son empaquetados y rodeados por membranas dando
lugar a los espermatoacuteforos que una vez completamente formados se almacenan en el
saco espermatofoacuterico o bolsa de Needhan de la cual salen a traveacutes del conducto
seminal aferente y del oacutergano terminal (o pene) La estructura maacutes caracteriacutestica del
macho es el tercer brazo derecho que presenta ciertas modificaciones morfoloacutegicas y
4
que por eso recibe el nombre de hectocoacutetilo (Fig 3) Este brazo que tiene la funcioacuten
de transferirdepositar los espermatoacuteforos en los conductos oviductales de la hembra
posee un surco a lo largo de toda su cara ventral por donde van a deslizar los
espermatoacuteforos en el momento de la coacutepula acabando en una zona lisa y aplanada
(liacutegula) a diferencia de los otros brazos que acaban en punta y con ventosas
AdultosSexos separados
Desarrollo embrionarioasymp 25-135 diacuteas
Fase post-eclosioacuten paralarvasEtapa de vida planctoacutenicaasymp 30-60 diacuteas
JuvenilesEtapa bentoacutenica
Juveniles hasta sub-adulto
Figura 2 - Esquema del ciclo de vida del pulpo Octopus vulgaris
Otra de las diferencias morfoloacutegicas entre machos y hembras en la edad adulta y
quizaacutes la maacutes faacutecil de identificar es que los machos poseen unas pocas ventosas de
tamantildeo destacado en los segundos y terceros pares de brazos mientras que en las
hembras las ventosas son de tamantildeo maacutes uniforme a lo largo de todo el brazo
Algunos autores apuntan a que estas grandes ventosas sirven para reconocimiento
sexual en el medio natural antes de que se inicie cualquier tipo de actividad
copulatoria (Hanlon y Messenger 1996)
5
Surc
o es
perm
atof
oacuteric
o
Liacutegu
la
glaacutendula oviductal nordm del brazo
liacutegula
hectocoacutetilo
A B
Macho
Hembra
Figura 3 ndash (A) Esquema de la transferencia de los espermatoacuteforos a traveacutes del hectocoacutetilo
hasta los conductos oviductales (DO) de la hembra (B) Aspecto detallado de la regioacuten distal
del brazo hectocotilizado de Octopus vulgaris mostrando la liacutegula
El sistema reproductor de las hembras consta de un ovario que desemboca en dos
oviductos y de las glaacutendulas oviductales (Fig 3) El ovario se situacutea en la parte
posterior de la cavidad del manto y en eacutel se forman los ovocitos Cuando la hembra es
fecundada los espermatoacuteforos se quedan almacenados en los receptaacuteculos seminales
de las glaacutendulas oviductales Se desconoce o al menos no hay evidencias claras de
que los pulpos tengan un comportamiento de cortejo habiendo sido descritas
situaciones de muacuteltiple transferencia de espermatoacuteforos por varios machos a una sola
hembra (Hanlon y Messenger 1996) Sin embargo hasta la fecha de hoy no hay
evidencias cientiacuteficas de la existencia de paternidad muacuteltiple para esta especie
desconocieacutendose por completo las implicaciones geneacuteticas y poblacionales que dicho
suceso pueda tener Se ha postulado ademaacutes que el uacuteltimo macho en copular con una
determinada hembra seriacutea el uacutenico padre de la progenie de la misma ya que eacuteste se
encargariacutea de eliminar los espermatoacuteforos dejados por otros machos precedentes Las
6
hembras son semelparas (se reproducen una sola vez) desarrollaacutendose el ovario de
forma sincroacutenica La ovulacioacuten se da de una sola vez no habiendo desarrollo y
maduracioacuten de nuevos ovocitos despueacutes de la uacutenica puesta estimada en 100000 a
500000 huevos (Mangold 1987) La fecundacioacuten en los pulpos es interna y se da a
medida que los ovocitos van pasando por el oviducto proximal hasta la zona distal
fertilizaacutendose a su paso por la glaacutendula oviductal donde estaacuten alojados los
espermatoacuteforos La puesta de los huevos puede tardar entre 15 y 30 diacuteas
Posteriormente la hembra pasa a ejercer cuidados maternales como proteccioacuten contra
depredadores y limpieza y oxigenacioacuten de los huevos pudiendo este periodo
prolongarse de 25 a 125 diacuteas dependiendo de la temperatura del agua (Mangold y
Boletzky 1973) Durante este tiempo los huevos de pulpo han de pasar por varias
fases de desarrollo (Fig 4) las cuales han sido clasificadas en XX estadios por Naef
(1928) A lo largo de este periodo las hembras dejan de alimentarse pudiendo perder
hasta el 60 de su peso corporal inicial antes de haber iniciado la puesta (Wodinsky
1978) La eclosioacuten de los huevos libera las paralarvas (Fig 4) teacutermino propuesto por
Young y Harman (1988) que asiacute se denominan porque en realidad no van a sufrir una
verdadera metamorfosis antes de convertirse en juveniles aunque al contrario que en
la vida adulta son planctoacutenicas en sus primeros diacuteas de vida Una vez finalizado el
periodo de incubacioacuten y liberacioacuten de las paralarvas las hembras mueren El tamantildeo
del manto de las paralarvas recieacuten eclosionadas es de alrededor de 2 mm y en cada
brazo poseen 3 ventosas (Boletzky 1987 Villanueva 1995) Se estima que el periodo
de vida planctoacutenico de las paralarvas es de 30 a 60 diacuteas dependiendo de la
temperatura del agua y de la zona geograacutefica (Itami et al 1963 Mangold y Boletzky
1973 Villanueva et al 1995) Se cree que las paralarvas de pulpo al igual que otras
larvas de cefaloacutepodos estudiadas en su medio natural (Vecchione 1987 Passarella y
Hopkins 1991) se alimentan de pequentildeos crustaacuteceos que forman parte del
zooplancton Estas criaturas son depredadoras activas desde el primer diacutea de vida
(Iglesias et al 2006) a pesar de que poseen reservas vitelinas internas que les
7
permiten sobrevivir en ausencia de alimento durante algunos diacuteas (Boletzky 1975) La
absorcioacuten de determinados nutrientes por la piel tambieacuten ha sido sentildealada como un
factor importante en los primeros estadios de vida de las larvas de cefaloacutepodos (Lee
1994) mientras que en otras especies de pulpos como el Octopus dofleini se ha
descrito un comportamiento de alimentacioacuten neustoacutenico es decir las paralarvas se
colocan con los brazos haciacutea la superficie aprovechando la tensioacuten superficial del
agua alimentaacutendose de los detritos orgaacutenicos o presas de esta interfase (Marliave
1981)
A
DC
B
Figura 4 - Fotografiacuteas a la lupa de huevos de pulpo en diferentes estadios de desarrollo y de
una paralarva de pulpo (A) huevos de pulpo (asymp 2 x 1 mm) recieacuten depositados (B) huevos en
una etapa de desarrollo intermedia (estadio X-XII) (C) huevos en un estado de desarrollo
avanzado (estadio XVIII) (D) paralarva de pulpo recieacuten eclosionada (tamantildeo total asymp 3 mm)
8
De los trabajos realizados en laboratorio sobre cultivo de paralarvas de O vulgaris
(Villanueva 1995 Iglesias et al 2004 Carrasco et al 2006) se sabe que a medida
que eacutestas van creciendo la proporcioacuten de los brazos frente al manto va aumentando y
una vez alcanzado un determinado tamantildeo realizan el asentamiento en el fondo
convirtieacutendose asiacute en juveniles bentoacutenicos de morfologiacutea igual que los adultos El
crecimiento de los juveniles hasta la fase adulta puede tardar de 8 a 15 meses
dependiendo mayormente de la temperatura y de la disponibilidad de alimento
(Mangold y Boletzky 1973 Forsythe y Van Heukelem 1987 Semmens et al 2004
Leporati et al 2007) Al igual que muchos otros cefaloacutepodos O vulgaris es una
especie carniacutevora durante todo su ciclo de vida En las fases de juvenil hasta la edad
adulta los pulpos se alimentan de una gran variedad de presas que estaacuten
representadas por la mayor parte de los fila marinos (Mangold 1983) de los cuales
varias especies de crustaacuteceos peces y otros moluscos constituyen las presas maacutes
comunes siendo los cangrejos una de sus favoritas (Nixon 1987) Esta especie
presenta ademaacutes un comportamiento de canibalismo frecuente en el medio natural A
pesar del gran intereacutes que hay en el cultivo industrial de esta especie todaviacutea persiste
un importante cuello de botella por resolver el cultivo de las paralarvas planctoacutenicas
hasta la fase de asentamiento cuando eacutestas se convierten en juveniles bentoacutenicos
12 El principal cuello de botella del cultivo del pulpo (Octopus vulgaris)
Actualmente el principal problema que impide el desarrollo integral del cultivo del pulpo
es la elevada mortalidad que se observa en la fase planctoacutenica A pesar de los intentos
llevados a cabo por diferentes grupos de trabajo de varios paiacuteses en los que se han
utilizado distintas metodologiacuteas de alimentacioacuten y condiciones zooteacutecnicas pocos
fueron los experimentos en los que se ha logrado obtener juveniles bentoacutenicos
(revisado por Iglesias et al 2007) Algunos de los primeros trabajos llevados a cabo a
9
nivel mundial se realizaron en Japoacuten Itami et al (1963) utilizando zoeas de camaroacuten
(Palaemon serrifer) lograron tener pulpos juveniles al cabo de 45 diacuteas con una
supervivencia del 8 Hamazaki et al (1991) publicaba igualmente la obtencioacuten de
juveniles de pulpo bentoacutenicos en condiciones controladas empleando como alimento
vivo juveniles de Artemia (15-2 mm) enriquecidos con Nannochloropsis sp y
antildeadiendo esta misma microalga al tanque de cultivo alcanzando en uno de sus
experimentos en un tanque de 20 m3 un nuacutemero de 23700 juveniles bentoacutenicos
(supervivencia del 29) al cabo de 25 diacuteas de cultivo
Al comienzo de los antildeos 90 se iniciaron tambieacuten en Espantildea un conjunto de
importantes trabajos relacionados con el cultivo del pulpo O vulgaris Los primeros
trabajos publicados sobre el cultivo larvario de cefaloacutepodos en laboratorio han sido
desarrollados por Villanueva (1994 1995) que probando zoeas de distintos
crustaacuteceos decaacutepodos (Pagurus prideaux Liocarcinus depurator y Dardanus arrosor)
logroacute obtener pulpos bentoacutenicos confirmando a las zoeas como un alimento apropiado
para suministrar a larvas de cefaloacutepodos Sin embargo este autor ya poniacutea de
manifiesto la dificultad en obtener zoeas en momentos precisos y los altos costes de
mantenimiento y mano de obra que implicaba esta praacutectica Por otro lado se iniciaban
tambieacuten en Espantildea un conjunto de experimentos de engorde de pulpo a escala de
laboratorio para evaluar el potencial de crecimiento de esta especie afrontaacutendose
entre los antildeos 1997 y 2000 el problema del cultivo de la fase planctoacutenica En los
primeros ensayos realizados Iglesias et al (1999 2000) probaron distintos tipos de
alimento tales como zooplancton del medio natural (copeacutepodos zoeas de crustaacuteceos
y misidaacuteceos) ictioplancton huevos de peces micropellets Artemia y rotiacuteferos y maacutes
tarde zoeas de crustaacuteceos obtenidas en laboratorio a partir de stocks de reproductores
de cangrejo (Carcinus maenas) neacutecora (Necora puber) y camaroacuten (Palaemon
serratus) Sin embargo la mortalidad de las paralarvas fue casi total en los primeros
diacuteas de vida alcanzaacutendose supervivencias maacuteximas del 10 a los 32 diacuteas al emplear
como alimento metanauplios y a continuacioacuten Artemia adulta enriquecida con
10
microalgas pero sin que se lograse alcanzar la fase de pre-asentamiento de las
paralarvas En los intentos llevados a cabo en el IEO de Vigo y maacutes tarde en el Centro
de Experimentacioacuten Pesquera de Asturias se obtuvieron algunos ejemplares de
juveniles bentoacutenicos que han llegado a alcanzar la edad adulta utilizaacutendose como
dieta para suministrar a las paralarvas juveniles de Artemia enriquecidos con
microalgas complementados con zoeas de centolla Maja brachydactyla en momentos
de disponibilidad de eacutestas (Iglesias et al 2002 2004 Carrasco et al 2003 2006) Las
primeras pruebas en las que se ha empleado esta mezcla de presas vivas (Moxica et
al 2002) fueron llevadas a cabo en el IEO de Vigo en un tanque de 9 m3 al que se
aportoacute inicialmente nauplios de Artemia y maacutes tarde juveniles de Artemia (1-4 mm) a
una densidad de presas de 01 Artemia ml-1 complementados con zoeas a partir de la
tercera semana de cultivo (3000 a 5000 zoeas al diacutea) Al tanque se aportaba a diario
una mezcla de microalgas (un 40 de Isochrysis galbana un 40 de Tetraselmis
suecica y un 20 de Chaetoceros sp) con el fin de mantener las presas vivas
constantemente enriquecidas Aunque no se han obtenido pulpos bentoacutenicos en este
experimento los autores refirieron la obtencioacuten de paralarvas en fase de asentamiento
con elevado nuacutemero de ventosas en los brazos (17-18) y un peso seco de 92 plusmn 09
mg a los 52 diacuteas de vida La supervivencia en este experimento fue de 83 al mes de
vida y de 02 al cabo de 52 diacuteas En un experimento posterior en el que se redujo la
escala y se empleoacute un tanque de 1 m3 las paralarvas se alimentaron con Artemia (1-4
mm) enriquecida con Chlorella sp antildeadiendo diariamente al tanque de cultivo una
mezcla de microalgas (Chlorella sp Isochrysis galbana y Chaetoceros sp) Como
complemento de Artemia se suministraron zoeas de centollo (Maja brachydactyla)
cuatro veces a la semana a una concentracioacuten de 001-01 zoeas ml-1 alcanzaacutendose
asiacute una supervivencia del 315 al diacutea 40 y un reducido nuacutemero de juveniles
bentoacutenicos dos semanas maacutes tarde (Iglesias et al 2002 2004) El seguimiento de los
uacutenicos pulpos supervivientes de este experimento hasta la fase adulta y la obtencioacuten
de una puesta de huevos por parte de una hembra originoacute el cierre del ciclo de vida
11
del pulpo en laboratorio por primer vez (Iglesias et al 2004) De forma similar
Carrasco et al (2003 2006) lograron obtener en uno de sus experimentos pulpos
bentoacutenicos a los 60 diacuteas con una supervivencia del 34
Sin embargo la utilizacioacuten de zoeas de crustaacuteceos como alimento vivo supone un
elevado riesgo ya que no se controla la obtencioacuten de las mismas en momentos
precisos ni la cantidad a producir ademaacutes de los elevados costes que conlleva esta
praacutectica por la necesidad de disponer de maacutes recursos materiales fiacutesicos y de
personal (Navarro y Villanueva 2000) Artemia en cambio es un crustaacuteceo faacutecil de
producir los nauplios o metanauplios se obtienen en 24-72 h y su composicioacuten
bioquiacutemica se puede modular mediante teacutecnicas de enriquecimiento al ser un filtrador
continuo obligado y no selectivo La utilizacioacuten de Artemia como presa viva se
consolida por tanto como una de las mejores alternativas para el cultivo de
paralarvas aunque sea necesario seguir trabajando en la modulacioacuten de su valor
nutricional de acuerdo con los posibles requerimientos de las paralarvas
Varios autores han utilizado nauplios de Artemia enriquecidos con distintos productos
comerciales (Navarro y Villanueva 2000 2003 Villanueva et al 2004 Okumura et al
2005) en sus experimentos de cultivo de paralarvas de pulpo siendo el tamantildeo de
estas presas (05-12 mm) adecuado hasta determinado momento del cultivo Sin
embargo se ha demostrado que las paralarvas tienen una preferencia clara por
Artemia de mayor tamantildeo (14 mm) sobre la maacutes pequentildea (08 mm) desde los
primeros diacuteas de vida (Iglesias et al 2006) De entre los productos o suplementos
nutricionales empleados por diversos autores para enriquecer Artemia se encuentran
emulsiones lipiacutedicas de la gama Selco (INVE) u otros aceites de pescado vitaminas
harina de huevos de pescado (BASF) harinas de cereales o productos de la gama Ori
(Skretting) (Navarro y Villanueva 2000 2003 Villanueva et al 2002 Iglesias et al
2004 Okumura et al 2005 Izquierdo et al 2008) pero ninguno ha originado tasas de
supervivencia o pesos secos de las paralarvas semejantes a los obtenidos con la
utilizacioacuten de zoeas de crustaacuteceos Como alternativa a las zoeas o a la Artemia
12
tambieacuten se han probado otros tipos de presas vivas o dietas artificiales para
suministrar a las paralarvas aunque en todo caso este tipo de pruebas se hicieron
complementadas con Artemia enriquecida (revisado por Iglesias et al 2007)
Otro de los principales suplementos empleados muy a menudo por diversos autores
para enriquecercultivar Artemia son las microalgas de las que las maacutes comuacutenmente
utilizadas fueron Nannochloropsis sp Dunaliella viridis Tetraselmis suecica
Isochrysis galbana Chaetoceros sp Chlorella sp (Hamazaki et al 1991 Navarro y
Villanueva 2000 Moxica et al 2002 Iglesias et al 2002 2004 Carrasco et al
2003 2006 Moxica et al 2006)
Hay un cierto consenso entre los grupos de investigadores que se dedican al cultivo
larvario del pulpo sobre que los aspectos nutricionales estaacuten en la base de la elevada
mortalidad observada durante su cultivo (Iglesias et al 2007) Uno de los problemas
que dificulta la interpretacioacuten de los resultados descritos y el avance en la nutricioacuten del
cultivo larvario del pulpo es que muy pocos autores presentan datos de la
composicioacuten bioquiacutemica de las dietas utilizadas y de sus efectos sobre la composicioacuten
de las paralarvas (Moxica et al 2002 Navarro et al 2000 2003 Villanueva et al
2004 Okumura et al 2005) cuando esta informacioacuten es crucial para interpretar los
resultados obtenidos y asiacute poder relacionarlos con la supervivencia y el crecimiento de
las paralarvas
A pesar de los continuos intentos para cerrar el ciclo de vida del pulpo de forma
rentable la mortalidad de las paralarvas durante la fase planctoacutenica sigue siendo casi
total Entre las estrategias propuestas para superar este importante problema estaacuten la
mejora de la composicioacuten de Artemia mediante las teacutecnicas de enriquecimiento
apropiadas y la formulacioacuten de microdietas inertes (Iglesias et al 2007) Importa
pues reducir el tiempo necesario hasta la obtencioacuten de los juveniles de Artemia del
tamantildeo deseado y optimizar su composicioacuten bioquiacutemica con base en los posibles
requerimientos nutricionales de las paralarvas El crecimiento y la composicioacuten
bioquiacutemica de Artemia sp pueden ser fuertemente modificados mediante la utilizacioacuten
13
de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

puestas de marzo a octubre mientras que machos maduros se encontraban a lo largo
de todo el antildeo Sin embargo dependiendo de la zona geograacutefica de donde provienen
se han identificado dos picos de puestas al antildeo el primero corresponde a la eacutepoca
marzo-mayo maacutes importante en el AtlaacutenticoMediterraacuteneo y la segunda al comienzo
del otontildeo (septiembre-octubre) maacutes importante en Japoacuten (FAO 2003)
El intereacutes comercial del desarrollo del cultivo del pulpo se debe a que esta especie es
muy demandada en varios paiacuteses de Europa y Asia ademaacutes de un elevado valor
comercial Presenta por otra parte algunas caracteriacutesticas bioloacutegicas muy
interesantes para ser considerado un serio candidato para la acuicultura ciclo de vida
corto (1-2 antildeos) elevadas tasas de crecimiento (entre el 10 y el 115 peso corporal
dia-1 a lo largo de toda su vida) alta tasa de conversioacuten alimentaria (30-60) elevada
fertilidad y muy alta viabilidad de los huevos faacutecil adaptacioacuten a la cautividad
aceptacioacuten de alimentos de bajo valor comercial elevado contenido proteico (70-85
del peso seco) y por uacuteltimo praacutecticamente ausencia de patologiacuteas (Mangold y
Boletzky 1973 Mangold 1983 Boucher-Rodoni et al 1987 Lee 1994 Villanueva
1995 Villanueva et al 1995 Iglesias et al 2000 2007 Vaz-Pires et al 2004) El
ciclo de vida del pulpo comuacuten se representa en la figura 2 El pulpo es una especie
gonocoacuterica es decir tiene los sexos separados y aunque no se aprecian diferencias
considerables de tamantildeo y peso corporal entre machos y hembras presentan un claro
dimorfismo sexual a partir del comienzo de la maduracioacuten sexual El aparato
reproductor de los machos posee un uacutenico testiacuteculo localizado en la parte anterior del
cuerpo donde se producen los espermatozoides que salen por el conducto seminal
deferente que a su vez se conecta con una serie de glaacutendulas En eacutestas los
espermatozoides producidos son empaquetados y rodeados por membranas dando
lugar a los espermatoacuteforos que una vez completamente formados se almacenan en el
saco espermatofoacuterico o bolsa de Needhan de la cual salen a traveacutes del conducto
seminal aferente y del oacutergano terminal (o pene) La estructura maacutes caracteriacutestica del
macho es el tercer brazo derecho que presenta ciertas modificaciones morfoloacutegicas y
4
que por eso recibe el nombre de hectocoacutetilo (Fig 3) Este brazo que tiene la funcioacuten
de transferirdepositar los espermatoacuteforos en los conductos oviductales de la hembra
posee un surco a lo largo de toda su cara ventral por donde van a deslizar los
espermatoacuteforos en el momento de la coacutepula acabando en una zona lisa y aplanada
(liacutegula) a diferencia de los otros brazos que acaban en punta y con ventosas
AdultosSexos separados
Desarrollo embrionarioasymp 25-135 diacuteas
Fase post-eclosioacuten paralarvasEtapa de vida planctoacutenicaasymp 30-60 diacuteas
JuvenilesEtapa bentoacutenica
Juveniles hasta sub-adulto
Figura 2 - Esquema del ciclo de vida del pulpo Octopus vulgaris
Otra de las diferencias morfoloacutegicas entre machos y hembras en la edad adulta y
quizaacutes la maacutes faacutecil de identificar es que los machos poseen unas pocas ventosas de
tamantildeo destacado en los segundos y terceros pares de brazos mientras que en las
hembras las ventosas son de tamantildeo maacutes uniforme a lo largo de todo el brazo
Algunos autores apuntan a que estas grandes ventosas sirven para reconocimiento
sexual en el medio natural antes de que se inicie cualquier tipo de actividad
copulatoria (Hanlon y Messenger 1996)
5
Surc
o es
perm
atof
oacuteric
o
Liacutegu
la
glaacutendula oviductal nordm del brazo
liacutegula
hectocoacutetilo
A B
Macho
Hembra
Figura 3 ndash (A) Esquema de la transferencia de los espermatoacuteforos a traveacutes del hectocoacutetilo
hasta los conductos oviductales (DO) de la hembra (B) Aspecto detallado de la regioacuten distal
del brazo hectocotilizado de Octopus vulgaris mostrando la liacutegula
El sistema reproductor de las hembras consta de un ovario que desemboca en dos
oviductos y de las glaacutendulas oviductales (Fig 3) El ovario se situacutea en la parte
posterior de la cavidad del manto y en eacutel se forman los ovocitos Cuando la hembra es
fecundada los espermatoacuteforos se quedan almacenados en los receptaacuteculos seminales
de las glaacutendulas oviductales Se desconoce o al menos no hay evidencias claras de
que los pulpos tengan un comportamiento de cortejo habiendo sido descritas
situaciones de muacuteltiple transferencia de espermatoacuteforos por varios machos a una sola
hembra (Hanlon y Messenger 1996) Sin embargo hasta la fecha de hoy no hay
evidencias cientiacuteficas de la existencia de paternidad muacuteltiple para esta especie
desconocieacutendose por completo las implicaciones geneacuteticas y poblacionales que dicho
suceso pueda tener Se ha postulado ademaacutes que el uacuteltimo macho en copular con una
determinada hembra seriacutea el uacutenico padre de la progenie de la misma ya que eacuteste se
encargariacutea de eliminar los espermatoacuteforos dejados por otros machos precedentes Las
6
hembras son semelparas (se reproducen una sola vez) desarrollaacutendose el ovario de
forma sincroacutenica La ovulacioacuten se da de una sola vez no habiendo desarrollo y
maduracioacuten de nuevos ovocitos despueacutes de la uacutenica puesta estimada en 100000 a
500000 huevos (Mangold 1987) La fecundacioacuten en los pulpos es interna y se da a
medida que los ovocitos van pasando por el oviducto proximal hasta la zona distal
fertilizaacutendose a su paso por la glaacutendula oviductal donde estaacuten alojados los
espermatoacuteforos La puesta de los huevos puede tardar entre 15 y 30 diacuteas
Posteriormente la hembra pasa a ejercer cuidados maternales como proteccioacuten contra
depredadores y limpieza y oxigenacioacuten de los huevos pudiendo este periodo
prolongarse de 25 a 125 diacuteas dependiendo de la temperatura del agua (Mangold y
Boletzky 1973) Durante este tiempo los huevos de pulpo han de pasar por varias
fases de desarrollo (Fig 4) las cuales han sido clasificadas en XX estadios por Naef
(1928) A lo largo de este periodo las hembras dejan de alimentarse pudiendo perder
hasta el 60 de su peso corporal inicial antes de haber iniciado la puesta (Wodinsky
1978) La eclosioacuten de los huevos libera las paralarvas (Fig 4) teacutermino propuesto por
Young y Harman (1988) que asiacute se denominan porque en realidad no van a sufrir una
verdadera metamorfosis antes de convertirse en juveniles aunque al contrario que en
la vida adulta son planctoacutenicas en sus primeros diacuteas de vida Una vez finalizado el
periodo de incubacioacuten y liberacioacuten de las paralarvas las hembras mueren El tamantildeo
del manto de las paralarvas recieacuten eclosionadas es de alrededor de 2 mm y en cada
brazo poseen 3 ventosas (Boletzky 1987 Villanueva 1995) Se estima que el periodo
de vida planctoacutenico de las paralarvas es de 30 a 60 diacuteas dependiendo de la
temperatura del agua y de la zona geograacutefica (Itami et al 1963 Mangold y Boletzky
1973 Villanueva et al 1995) Se cree que las paralarvas de pulpo al igual que otras
larvas de cefaloacutepodos estudiadas en su medio natural (Vecchione 1987 Passarella y
Hopkins 1991) se alimentan de pequentildeos crustaacuteceos que forman parte del
zooplancton Estas criaturas son depredadoras activas desde el primer diacutea de vida
(Iglesias et al 2006) a pesar de que poseen reservas vitelinas internas que les
7
permiten sobrevivir en ausencia de alimento durante algunos diacuteas (Boletzky 1975) La
absorcioacuten de determinados nutrientes por la piel tambieacuten ha sido sentildealada como un
factor importante en los primeros estadios de vida de las larvas de cefaloacutepodos (Lee
1994) mientras que en otras especies de pulpos como el Octopus dofleini se ha
descrito un comportamiento de alimentacioacuten neustoacutenico es decir las paralarvas se
colocan con los brazos haciacutea la superficie aprovechando la tensioacuten superficial del
agua alimentaacutendose de los detritos orgaacutenicos o presas de esta interfase (Marliave
1981)
A
DC
B
Figura 4 - Fotografiacuteas a la lupa de huevos de pulpo en diferentes estadios de desarrollo y de
una paralarva de pulpo (A) huevos de pulpo (asymp 2 x 1 mm) recieacuten depositados (B) huevos en
una etapa de desarrollo intermedia (estadio X-XII) (C) huevos en un estado de desarrollo
avanzado (estadio XVIII) (D) paralarva de pulpo recieacuten eclosionada (tamantildeo total asymp 3 mm)
8
De los trabajos realizados en laboratorio sobre cultivo de paralarvas de O vulgaris
(Villanueva 1995 Iglesias et al 2004 Carrasco et al 2006) se sabe que a medida
que eacutestas van creciendo la proporcioacuten de los brazos frente al manto va aumentando y
una vez alcanzado un determinado tamantildeo realizan el asentamiento en el fondo
convirtieacutendose asiacute en juveniles bentoacutenicos de morfologiacutea igual que los adultos El
crecimiento de los juveniles hasta la fase adulta puede tardar de 8 a 15 meses
dependiendo mayormente de la temperatura y de la disponibilidad de alimento
(Mangold y Boletzky 1973 Forsythe y Van Heukelem 1987 Semmens et al 2004
Leporati et al 2007) Al igual que muchos otros cefaloacutepodos O vulgaris es una
especie carniacutevora durante todo su ciclo de vida En las fases de juvenil hasta la edad
adulta los pulpos se alimentan de una gran variedad de presas que estaacuten
representadas por la mayor parte de los fila marinos (Mangold 1983) de los cuales
varias especies de crustaacuteceos peces y otros moluscos constituyen las presas maacutes
comunes siendo los cangrejos una de sus favoritas (Nixon 1987) Esta especie
presenta ademaacutes un comportamiento de canibalismo frecuente en el medio natural A
pesar del gran intereacutes que hay en el cultivo industrial de esta especie todaviacutea persiste
un importante cuello de botella por resolver el cultivo de las paralarvas planctoacutenicas
hasta la fase de asentamiento cuando eacutestas se convierten en juveniles bentoacutenicos
12 El principal cuello de botella del cultivo del pulpo (Octopus vulgaris)
Actualmente el principal problema que impide el desarrollo integral del cultivo del pulpo
es la elevada mortalidad que se observa en la fase planctoacutenica A pesar de los intentos
llevados a cabo por diferentes grupos de trabajo de varios paiacuteses en los que se han
utilizado distintas metodologiacuteas de alimentacioacuten y condiciones zooteacutecnicas pocos
fueron los experimentos en los que se ha logrado obtener juveniles bentoacutenicos
(revisado por Iglesias et al 2007) Algunos de los primeros trabajos llevados a cabo a
9
nivel mundial se realizaron en Japoacuten Itami et al (1963) utilizando zoeas de camaroacuten
(Palaemon serrifer) lograron tener pulpos juveniles al cabo de 45 diacuteas con una
supervivencia del 8 Hamazaki et al (1991) publicaba igualmente la obtencioacuten de
juveniles de pulpo bentoacutenicos en condiciones controladas empleando como alimento
vivo juveniles de Artemia (15-2 mm) enriquecidos con Nannochloropsis sp y
antildeadiendo esta misma microalga al tanque de cultivo alcanzando en uno de sus
experimentos en un tanque de 20 m3 un nuacutemero de 23700 juveniles bentoacutenicos
(supervivencia del 29) al cabo de 25 diacuteas de cultivo
Al comienzo de los antildeos 90 se iniciaron tambieacuten en Espantildea un conjunto de
importantes trabajos relacionados con el cultivo del pulpo O vulgaris Los primeros
trabajos publicados sobre el cultivo larvario de cefaloacutepodos en laboratorio han sido
desarrollados por Villanueva (1994 1995) que probando zoeas de distintos
crustaacuteceos decaacutepodos (Pagurus prideaux Liocarcinus depurator y Dardanus arrosor)
logroacute obtener pulpos bentoacutenicos confirmando a las zoeas como un alimento apropiado
para suministrar a larvas de cefaloacutepodos Sin embargo este autor ya poniacutea de
manifiesto la dificultad en obtener zoeas en momentos precisos y los altos costes de
mantenimiento y mano de obra que implicaba esta praacutectica Por otro lado se iniciaban
tambieacuten en Espantildea un conjunto de experimentos de engorde de pulpo a escala de
laboratorio para evaluar el potencial de crecimiento de esta especie afrontaacutendose
entre los antildeos 1997 y 2000 el problema del cultivo de la fase planctoacutenica En los
primeros ensayos realizados Iglesias et al (1999 2000) probaron distintos tipos de
alimento tales como zooplancton del medio natural (copeacutepodos zoeas de crustaacuteceos
y misidaacuteceos) ictioplancton huevos de peces micropellets Artemia y rotiacuteferos y maacutes
tarde zoeas de crustaacuteceos obtenidas en laboratorio a partir de stocks de reproductores
de cangrejo (Carcinus maenas) neacutecora (Necora puber) y camaroacuten (Palaemon
serratus) Sin embargo la mortalidad de las paralarvas fue casi total en los primeros
diacuteas de vida alcanzaacutendose supervivencias maacuteximas del 10 a los 32 diacuteas al emplear
como alimento metanauplios y a continuacioacuten Artemia adulta enriquecida con
10
microalgas pero sin que se lograse alcanzar la fase de pre-asentamiento de las
paralarvas En los intentos llevados a cabo en el IEO de Vigo y maacutes tarde en el Centro
de Experimentacioacuten Pesquera de Asturias se obtuvieron algunos ejemplares de
juveniles bentoacutenicos que han llegado a alcanzar la edad adulta utilizaacutendose como
dieta para suministrar a las paralarvas juveniles de Artemia enriquecidos con
microalgas complementados con zoeas de centolla Maja brachydactyla en momentos
de disponibilidad de eacutestas (Iglesias et al 2002 2004 Carrasco et al 2003 2006) Las
primeras pruebas en las que se ha empleado esta mezcla de presas vivas (Moxica et
al 2002) fueron llevadas a cabo en el IEO de Vigo en un tanque de 9 m3 al que se
aportoacute inicialmente nauplios de Artemia y maacutes tarde juveniles de Artemia (1-4 mm) a
una densidad de presas de 01 Artemia ml-1 complementados con zoeas a partir de la
tercera semana de cultivo (3000 a 5000 zoeas al diacutea) Al tanque se aportaba a diario
una mezcla de microalgas (un 40 de Isochrysis galbana un 40 de Tetraselmis
suecica y un 20 de Chaetoceros sp) con el fin de mantener las presas vivas
constantemente enriquecidas Aunque no se han obtenido pulpos bentoacutenicos en este
experimento los autores refirieron la obtencioacuten de paralarvas en fase de asentamiento
con elevado nuacutemero de ventosas en los brazos (17-18) y un peso seco de 92 plusmn 09
mg a los 52 diacuteas de vida La supervivencia en este experimento fue de 83 al mes de
vida y de 02 al cabo de 52 diacuteas En un experimento posterior en el que se redujo la
escala y se empleoacute un tanque de 1 m3 las paralarvas se alimentaron con Artemia (1-4
mm) enriquecida con Chlorella sp antildeadiendo diariamente al tanque de cultivo una
mezcla de microalgas (Chlorella sp Isochrysis galbana y Chaetoceros sp) Como
complemento de Artemia se suministraron zoeas de centollo (Maja brachydactyla)
cuatro veces a la semana a una concentracioacuten de 001-01 zoeas ml-1 alcanzaacutendose
asiacute una supervivencia del 315 al diacutea 40 y un reducido nuacutemero de juveniles
bentoacutenicos dos semanas maacutes tarde (Iglesias et al 2002 2004) El seguimiento de los
uacutenicos pulpos supervivientes de este experimento hasta la fase adulta y la obtencioacuten
de una puesta de huevos por parte de una hembra originoacute el cierre del ciclo de vida
11
del pulpo en laboratorio por primer vez (Iglesias et al 2004) De forma similar
Carrasco et al (2003 2006) lograron obtener en uno de sus experimentos pulpos
bentoacutenicos a los 60 diacuteas con una supervivencia del 34
Sin embargo la utilizacioacuten de zoeas de crustaacuteceos como alimento vivo supone un
elevado riesgo ya que no se controla la obtencioacuten de las mismas en momentos
precisos ni la cantidad a producir ademaacutes de los elevados costes que conlleva esta
praacutectica por la necesidad de disponer de maacutes recursos materiales fiacutesicos y de
personal (Navarro y Villanueva 2000) Artemia en cambio es un crustaacuteceo faacutecil de
producir los nauplios o metanauplios se obtienen en 24-72 h y su composicioacuten
bioquiacutemica se puede modular mediante teacutecnicas de enriquecimiento al ser un filtrador
continuo obligado y no selectivo La utilizacioacuten de Artemia como presa viva se
consolida por tanto como una de las mejores alternativas para el cultivo de
paralarvas aunque sea necesario seguir trabajando en la modulacioacuten de su valor
nutricional de acuerdo con los posibles requerimientos de las paralarvas
Varios autores han utilizado nauplios de Artemia enriquecidos con distintos productos
comerciales (Navarro y Villanueva 2000 2003 Villanueva et al 2004 Okumura et al
2005) en sus experimentos de cultivo de paralarvas de pulpo siendo el tamantildeo de
estas presas (05-12 mm) adecuado hasta determinado momento del cultivo Sin
embargo se ha demostrado que las paralarvas tienen una preferencia clara por
Artemia de mayor tamantildeo (14 mm) sobre la maacutes pequentildea (08 mm) desde los
primeros diacuteas de vida (Iglesias et al 2006) De entre los productos o suplementos
nutricionales empleados por diversos autores para enriquecer Artemia se encuentran
emulsiones lipiacutedicas de la gama Selco (INVE) u otros aceites de pescado vitaminas
harina de huevos de pescado (BASF) harinas de cereales o productos de la gama Ori
(Skretting) (Navarro y Villanueva 2000 2003 Villanueva et al 2002 Iglesias et al
2004 Okumura et al 2005 Izquierdo et al 2008) pero ninguno ha originado tasas de
supervivencia o pesos secos de las paralarvas semejantes a los obtenidos con la
utilizacioacuten de zoeas de crustaacuteceos Como alternativa a las zoeas o a la Artemia
12
tambieacuten se han probado otros tipos de presas vivas o dietas artificiales para
suministrar a las paralarvas aunque en todo caso este tipo de pruebas se hicieron
complementadas con Artemia enriquecida (revisado por Iglesias et al 2007)
Otro de los principales suplementos empleados muy a menudo por diversos autores
para enriquecercultivar Artemia son las microalgas de las que las maacutes comuacutenmente
utilizadas fueron Nannochloropsis sp Dunaliella viridis Tetraselmis suecica
Isochrysis galbana Chaetoceros sp Chlorella sp (Hamazaki et al 1991 Navarro y
Villanueva 2000 Moxica et al 2002 Iglesias et al 2002 2004 Carrasco et al
2003 2006 Moxica et al 2006)
Hay un cierto consenso entre los grupos de investigadores que se dedican al cultivo
larvario del pulpo sobre que los aspectos nutricionales estaacuten en la base de la elevada
mortalidad observada durante su cultivo (Iglesias et al 2007) Uno de los problemas
que dificulta la interpretacioacuten de los resultados descritos y el avance en la nutricioacuten del
cultivo larvario del pulpo es que muy pocos autores presentan datos de la
composicioacuten bioquiacutemica de las dietas utilizadas y de sus efectos sobre la composicioacuten
de las paralarvas (Moxica et al 2002 Navarro et al 2000 2003 Villanueva et al
2004 Okumura et al 2005) cuando esta informacioacuten es crucial para interpretar los
resultados obtenidos y asiacute poder relacionarlos con la supervivencia y el crecimiento de
las paralarvas
A pesar de los continuos intentos para cerrar el ciclo de vida del pulpo de forma
rentable la mortalidad de las paralarvas durante la fase planctoacutenica sigue siendo casi
total Entre las estrategias propuestas para superar este importante problema estaacuten la
mejora de la composicioacuten de Artemia mediante las teacutecnicas de enriquecimiento
apropiadas y la formulacioacuten de microdietas inertes (Iglesias et al 2007) Importa
pues reducir el tiempo necesario hasta la obtencioacuten de los juveniles de Artemia del
tamantildeo deseado y optimizar su composicioacuten bioquiacutemica con base en los posibles
requerimientos nutricionales de las paralarvas El crecimiento y la composicioacuten
bioquiacutemica de Artemia sp pueden ser fuertemente modificados mediante la utilizacioacuten
13
de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

que por eso recibe el nombre de hectocoacutetilo (Fig 3) Este brazo que tiene la funcioacuten
de transferirdepositar los espermatoacuteforos en los conductos oviductales de la hembra
posee un surco a lo largo de toda su cara ventral por donde van a deslizar los
espermatoacuteforos en el momento de la coacutepula acabando en una zona lisa y aplanada
(liacutegula) a diferencia de los otros brazos que acaban en punta y con ventosas
AdultosSexos separados
Desarrollo embrionarioasymp 25-135 diacuteas
Fase post-eclosioacuten paralarvasEtapa de vida planctoacutenicaasymp 30-60 diacuteas
JuvenilesEtapa bentoacutenica
Juveniles hasta sub-adulto
Figura 2 - Esquema del ciclo de vida del pulpo Octopus vulgaris
Otra de las diferencias morfoloacutegicas entre machos y hembras en la edad adulta y
quizaacutes la maacutes faacutecil de identificar es que los machos poseen unas pocas ventosas de
tamantildeo destacado en los segundos y terceros pares de brazos mientras que en las
hembras las ventosas son de tamantildeo maacutes uniforme a lo largo de todo el brazo
Algunos autores apuntan a que estas grandes ventosas sirven para reconocimiento
sexual en el medio natural antes de que se inicie cualquier tipo de actividad
copulatoria (Hanlon y Messenger 1996)
5
Surc
o es
perm
atof
oacuteric
o
Liacutegu
la
glaacutendula oviductal nordm del brazo
liacutegula
hectocoacutetilo
A B
Macho
Hembra
Figura 3 ndash (A) Esquema de la transferencia de los espermatoacuteforos a traveacutes del hectocoacutetilo
hasta los conductos oviductales (DO) de la hembra (B) Aspecto detallado de la regioacuten distal
del brazo hectocotilizado de Octopus vulgaris mostrando la liacutegula
El sistema reproductor de las hembras consta de un ovario que desemboca en dos
oviductos y de las glaacutendulas oviductales (Fig 3) El ovario se situacutea en la parte
posterior de la cavidad del manto y en eacutel se forman los ovocitos Cuando la hembra es
fecundada los espermatoacuteforos se quedan almacenados en los receptaacuteculos seminales
de las glaacutendulas oviductales Se desconoce o al menos no hay evidencias claras de
que los pulpos tengan un comportamiento de cortejo habiendo sido descritas
situaciones de muacuteltiple transferencia de espermatoacuteforos por varios machos a una sola
hembra (Hanlon y Messenger 1996) Sin embargo hasta la fecha de hoy no hay
evidencias cientiacuteficas de la existencia de paternidad muacuteltiple para esta especie
desconocieacutendose por completo las implicaciones geneacuteticas y poblacionales que dicho
suceso pueda tener Se ha postulado ademaacutes que el uacuteltimo macho en copular con una
determinada hembra seriacutea el uacutenico padre de la progenie de la misma ya que eacuteste se
encargariacutea de eliminar los espermatoacuteforos dejados por otros machos precedentes Las
6
hembras son semelparas (se reproducen una sola vez) desarrollaacutendose el ovario de
forma sincroacutenica La ovulacioacuten se da de una sola vez no habiendo desarrollo y
maduracioacuten de nuevos ovocitos despueacutes de la uacutenica puesta estimada en 100000 a
500000 huevos (Mangold 1987) La fecundacioacuten en los pulpos es interna y se da a
medida que los ovocitos van pasando por el oviducto proximal hasta la zona distal
fertilizaacutendose a su paso por la glaacutendula oviductal donde estaacuten alojados los
espermatoacuteforos La puesta de los huevos puede tardar entre 15 y 30 diacuteas
Posteriormente la hembra pasa a ejercer cuidados maternales como proteccioacuten contra
depredadores y limpieza y oxigenacioacuten de los huevos pudiendo este periodo
prolongarse de 25 a 125 diacuteas dependiendo de la temperatura del agua (Mangold y
Boletzky 1973) Durante este tiempo los huevos de pulpo han de pasar por varias
fases de desarrollo (Fig 4) las cuales han sido clasificadas en XX estadios por Naef
(1928) A lo largo de este periodo las hembras dejan de alimentarse pudiendo perder
hasta el 60 de su peso corporal inicial antes de haber iniciado la puesta (Wodinsky
1978) La eclosioacuten de los huevos libera las paralarvas (Fig 4) teacutermino propuesto por
Young y Harman (1988) que asiacute se denominan porque en realidad no van a sufrir una
verdadera metamorfosis antes de convertirse en juveniles aunque al contrario que en
la vida adulta son planctoacutenicas en sus primeros diacuteas de vida Una vez finalizado el
periodo de incubacioacuten y liberacioacuten de las paralarvas las hembras mueren El tamantildeo
del manto de las paralarvas recieacuten eclosionadas es de alrededor de 2 mm y en cada
brazo poseen 3 ventosas (Boletzky 1987 Villanueva 1995) Se estima que el periodo
de vida planctoacutenico de las paralarvas es de 30 a 60 diacuteas dependiendo de la
temperatura del agua y de la zona geograacutefica (Itami et al 1963 Mangold y Boletzky
1973 Villanueva et al 1995) Se cree que las paralarvas de pulpo al igual que otras
larvas de cefaloacutepodos estudiadas en su medio natural (Vecchione 1987 Passarella y
Hopkins 1991) se alimentan de pequentildeos crustaacuteceos que forman parte del
zooplancton Estas criaturas son depredadoras activas desde el primer diacutea de vida
(Iglesias et al 2006) a pesar de que poseen reservas vitelinas internas que les
7
permiten sobrevivir en ausencia de alimento durante algunos diacuteas (Boletzky 1975) La
absorcioacuten de determinados nutrientes por la piel tambieacuten ha sido sentildealada como un
factor importante en los primeros estadios de vida de las larvas de cefaloacutepodos (Lee
1994) mientras que en otras especies de pulpos como el Octopus dofleini se ha
descrito un comportamiento de alimentacioacuten neustoacutenico es decir las paralarvas se
colocan con los brazos haciacutea la superficie aprovechando la tensioacuten superficial del
agua alimentaacutendose de los detritos orgaacutenicos o presas de esta interfase (Marliave
1981)
A
DC
B
Figura 4 - Fotografiacuteas a la lupa de huevos de pulpo en diferentes estadios de desarrollo y de
una paralarva de pulpo (A) huevos de pulpo (asymp 2 x 1 mm) recieacuten depositados (B) huevos en
una etapa de desarrollo intermedia (estadio X-XII) (C) huevos en un estado de desarrollo
avanzado (estadio XVIII) (D) paralarva de pulpo recieacuten eclosionada (tamantildeo total asymp 3 mm)
8
De los trabajos realizados en laboratorio sobre cultivo de paralarvas de O vulgaris
(Villanueva 1995 Iglesias et al 2004 Carrasco et al 2006) se sabe que a medida
que eacutestas van creciendo la proporcioacuten de los brazos frente al manto va aumentando y
una vez alcanzado un determinado tamantildeo realizan el asentamiento en el fondo
convirtieacutendose asiacute en juveniles bentoacutenicos de morfologiacutea igual que los adultos El
crecimiento de los juveniles hasta la fase adulta puede tardar de 8 a 15 meses
dependiendo mayormente de la temperatura y de la disponibilidad de alimento
(Mangold y Boletzky 1973 Forsythe y Van Heukelem 1987 Semmens et al 2004
Leporati et al 2007) Al igual que muchos otros cefaloacutepodos O vulgaris es una
especie carniacutevora durante todo su ciclo de vida En las fases de juvenil hasta la edad
adulta los pulpos se alimentan de una gran variedad de presas que estaacuten
representadas por la mayor parte de los fila marinos (Mangold 1983) de los cuales
varias especies de crustaacuteceos peces y otros moluscos constituyen las presas maacutes
comunes siendo los cangrejos una de sus favoritas (Nixon 1987) Esta especie
presenta ademaacutes un comportamiento de canibalismo frecuente en el medio natural A
pesar del gran intereacutes que hay en el cultivo industrial de esta especie todaviacutea persiste
un importante cuello de botella por resolver el cultivo de las paralarvas planctoacutenicas
hasta la fase de asentamiento cuando eacutestas se convierten en juveniles bentoacutenicos
12 El principal cuello de botella del cultivo del pulpo (Octopus vulgaris)
Actualmente el principal problema que impide el desarrollo integral del cultivo del pulpo
es la elevada mortalidad que se observa en la fase planctoacutenica A pesar de los intentos
llevados a cabo por diferentes grupos de trabajo de varios paiacuteses en los que se han
utilizado distintas metodologiacuteas de alimentacioacuten y condiciones zooteacutecnicas pocos
fueron los experimentos en los que se ha logrado obtener juveniles bentoacutenicos
(revisado por Iglesias et al 2007) Algunos de los primeros trabajos llevados a cabo a
9
nivel mundial se realizaron en Japoacuten Itami et al (1963) utilizando zoeas de camaroacuten
(Palaemon serrifer) lograron tener pulpos juveniles al cabo de 45 diacuteas con una
supervivencia del 8 Hamazaki et al (1991) publicaba igualmente la obtencioacuten de
juveniles de pulpo bentoacutenicos en condiciones controladas empleando como alimento
vivo juveniles de Artemia (15-2 mm) enriquecidos con Nannochloropsis sp y
antildeadiendo esta misma microalga al tanque de cultivo alcanzando en uno de sus
experimentos en un tanque de 20 m3 un nuacutemero de 23700 juveniles bentoacutenicos
(supervivencia del 29) al cabo de 25 diacuteas de cultivo
Al comienzo de los antildeos 90 se iniciaron tambieacuten en Espantildea un conjunto de
importantes trabajos relacionados con el cultivo del pulpo O vulgaris Los primeros
trabajos publicados sobre el cultivo larvario de cefaloacutepodos en laboratorio han sido
desarrollados por Villanueva (1994 1995) que probando zoeas de distintos
crustaacuteceos decaacutepodos (Pagurus prideaux Liocarcinus depurator y Dardanus arrosor)
logroacute obtener pulpos bentoacutenicos confirmando a las zoeas como un alimento apropiado
para suministrar a larvas de cefaloacutepodos Sin embargo este autor ya poniacutea de
manifiesto la dificultad en obtener zoeas en momentos precisos y los altos costes de
mantenimiento y mano de obra que implicaba esta praacutectica Por otro lado se iniciaban
tambieacuten en Espantildea un conjunto de experimentos de engorde de pulpo a escala de
laboratorio para evaluar el potencial de crecimiento de esta especie afrontaacutendose
entre los antildeos 1997 y 2000 el problema del cultivo de la fase planctoacutenica En los
primeros ensayos realizados Iglesias et al (1999 2000) probaron distintos tipos de
alimento tales como zooplancton del medio natural (copeacutepodos zoeas de crustaacuteceos
y misidaacuteceos) ictioplancton huevos de peces micropellets Artemia y rotiacuteferos y maacutes
tarde zoeas de crustaacuteceos obtenidas en laboratorio a partir de stocks de reproductores
de cangrejo (Carcinus maenas) neacutecora (Necora puber) y camaroacuten (Palaemon
serratus) Sin embargo la mortalidad de las paralarvas fue casi total en los primeros
diacuteas de vida alcanzaacutendose supervivencias maacuteximas del 10 a los 32 diacuteas al emplear
como alimento metanauplios y a continuacioacuten Artemia adulta enriquecida con
10
microalgas pero sin que se lograse alcanzar la fase de pre-asentamiento de las
paralarvas En los intentos llevados a cabo en el IEO de Vigo y maacutes tarde en el Centro
de Experimentacioacuten Pesquera de Asturias se obtuvieron algunos ejemplares de
juveniles bentoacutenicos que han llegado a alcanzar la edad adulta utilizaacutendose como
dieta para suministrar a las paralarvas juveniles de Artemia enriquecidos con
microalgas complementados con zoeas de centolla Maja brachydactyla en momentos
de disponibilidad de eacutestas (Iglesias et al 2002 2004 Carrasco et al 2003 2006) Las
primeras pruebas en las que se ha empleado esta mezcla de presas vivas (Moxica et
al 2002) fueron llevadas a cabo en el IEO de Vigo en un tanque de 9 m3 al que se
aportoacute inicialmente nauplios de Artemia y maacutes tarde juveniles de Artemia (1-4 mm) a
una densidad de presas de 01 Artemia ml-1 complementados con zoeas a partir de la
tercera semana de cultivo (3000 a 5000 zoeas al diacutea) Al tanque se aportaba a diario
una mezcla de microalgas (un 40 de Isochrysis galbana un 40 de Tetraselmis
suecica y un 20 de Chaetoceros sp) con el fin de mantener las presas vivas
constantemente enriquecidas Aunque no se han obtenido pulpos bentoacutenicos en este
experimento los autores refirieron la obtencioacuten de paralarvas en fase de asentamiento
con elevado nuacutemero de ventosas en los brazos (17-18) y un peso seco de 92 plusmn 09
mg a los 52 diacuteas de vida La supervivencia en este experimento fue de 83 al mes de
vida y de 02 al cabo de 52 diacuteas En un experimento posterior en el que se redujo la
escala y se empleoacute un tanque de 1 m3 las paralarvas se alimentaron con Artemia (1-4
mm) enriquecida con Chlorella sp antildeadiendo diariamente al tanque de cultivo una
mezcla de microalgas (Chlorella sp Isochrysis galbana y Chaetoceros sp) Como
complemento de Artemia se suministraron zoeas de centollo (Maja brachydactyla)
cuatro veces a la semana a una concentracioacuten de 001-01 zoeas ml-1 alcanzaacutendose
asiacute una supervivencia del 315 al diacutea 40 y un reducido nuacutemero de juveniles
bentoacutenicos dos semanas maacutes tarde (Iglesias et al 2002 2004) El seguimiento de los
uacutenicos pulpos supervivientes de este experimento hasta la fase adulta y la obtencioacuten
de una puesta de huevos por parte de una hembra originoacute el cierre del ciclo de vida
11
del pulpo en laboratorio por primer vez (Iglesias et al 2004) De forma similar
Carrasco et al (2003 2006) lograron obtener en uno de sus experimentos pulpos
bentoacutenicos a los 60 diacuteas con una supervivencia del 34
Sin embargo la utilizacioacuten de zoeas de crustaacuteceos como alimento vivo supone un
elevado riesgo ya que no se controla la obtencioacuten de las mismas en momentos
precisos ni la cantidad a producir ademaacutes de los elevados costes que conlleva esta
praacutectica por la necesidad de disponer de maacutes recursos materiales fiacutesicos y de
personal (Navarro y Villanueva 2000) Artemia en cambio es un crustaacuteceo faacutecil de
producir los nauplios o metanauplios se obtienen en 24-72 h y su composicioacuten
bioquiacutemica se puede modular mediante teacutecnicas de enriquecimiento al ser un filtrador
continuo obligado y no selectivo La utilizacioacuten de Artemia como presa viva se
consolida por tanto como una de las mejores alternativas para el cultivo de
paralarvas aunque sea necesario seguir trabajando en la modulacioacuten de su valor
nutricional de acuerdo con los posibles requerimientos de las paralarvas
Varios autores han utilizado nauplios de Artemia enriquecidos con distintos productos
comerciales (Navarro y Villanueva 2000 2003 Villanueva et al 2004 Okumura et al
2005) en sus experimentos de cultivo de paralarvas de pulpo siendo el tamantildeo de
estas presas (05-12 mm) adecuado hasta determinado momento del cultivo Sin
embargo se ha demostrado que las paralarvas tienen una preferencia clara por
Artemia de mayor tamantildeo (14 mm) sobre la maacutes pequentildea (08 mm) desde los
primeros diacuteas de vida (Iglesias et al 2006) De entre los productos o suplementos
nutricionales empleados por diversos autores para enriquecer Artemia se encuentran
emulsiones lipiacutedicas de la gama Selco (INVE) u otros aceites de pescado vitaminas
harina de huevos de pescado (BASF) harinas de cereales o productos de la gama Ori
(Skretting) (Navarro y Villanueva 2000 2003 Villanueva et al 2002 Iglesias et al
2004 Okumura et al 2005 Izquierdo et al 2008) pero ninguno ha originado tasas de
supervivencia o pesos secos de las paralarvas semejantes a los obtenidos con la
utilizacioacuten de zoeas de crustaacuteceos Como alternativa a las zoeas o a la Artemia
12
tambieacuten se han probado otros tipos de presas vivas o dietas artificiales para
suministrar a las paralarvas aunque en todo caso este tipo de pruebas se hicieron
complementadas con Artemia enriquecida (revisado por Iglesias et al 2007)
Otro de los principales suplementos empleados muy a menudo por diversos autores
para enriquecercultivar Artemia son las microalgas de las que las maacutes comuacutenmente
utilizadas fueron Nannochloropsis sp Dunaliella viridis Tetraselmis suecica
Isochrysis galbana Chaetoceros sp Chlorella sp (Hamazaki et al 1991 Navarro y
Villanueva 2000 Moxica et al 2002 Iglesias et al 2002 2004 Carrasco et al
2003 2006 Moxica et al 2006)
Hay un cierto consenso entre los grupos de investigadores que se dedican al cultivo
larvario del pulpo sobre que los aspectos nutricionales estaacuten en la base de la elevada
mortalidad observada durante su cultivo (Iglesias et al 2007) Uno de los problemas
que dificulta la interpretacioacuten de los resultados descritos y el avance en la nutricioacuten del
cultivo larvario del pulpo es que muy pocos autores presentan datos de la
composicioacuten bioquiacutemica de las dietas utilizadas y de sus efectos sobre la composicioacuten
de las paralarvas (Moxica et al 2002 Navarro et al 2000 2003 Villanueva et al
2004 Okumura et al 2005) cuando esta informacioacuten es crucial para interpretar los
resultados obtenidos y asiacute poder relacionarlos con la supervivencia y el crecimiento de
las paralarvas
A pesar de los continuos intentos para cerrar el ciclo de vida del pulpo de forma
rentable la mortalidad de las paralarvas durante la fase planctoacutenica sigue siendo casi
total Entre las estrategias propuestas para superar este importante problema estaacuten la
mejora de la composicioacuten de Artemia mediante las teacutecnicas de enriquecimiento
apropiadas y la formulacioacuten de microdietas inertes (Iglesias et al 2007) Importa
pues reducir el tiempo necesario hasta la obtencioacuten de los juveniles de Artemia del
tamantildeo deseado y optimizar su composicioacuten bioquiacutemica con base en los posibles
requerimientos nutricionales de las paralarvas El crecimiento y la composicioacuten
bioquiacutemica de Artemia sp pueden ser fuertemente modificados mediante la utilizacioacuten
13
de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
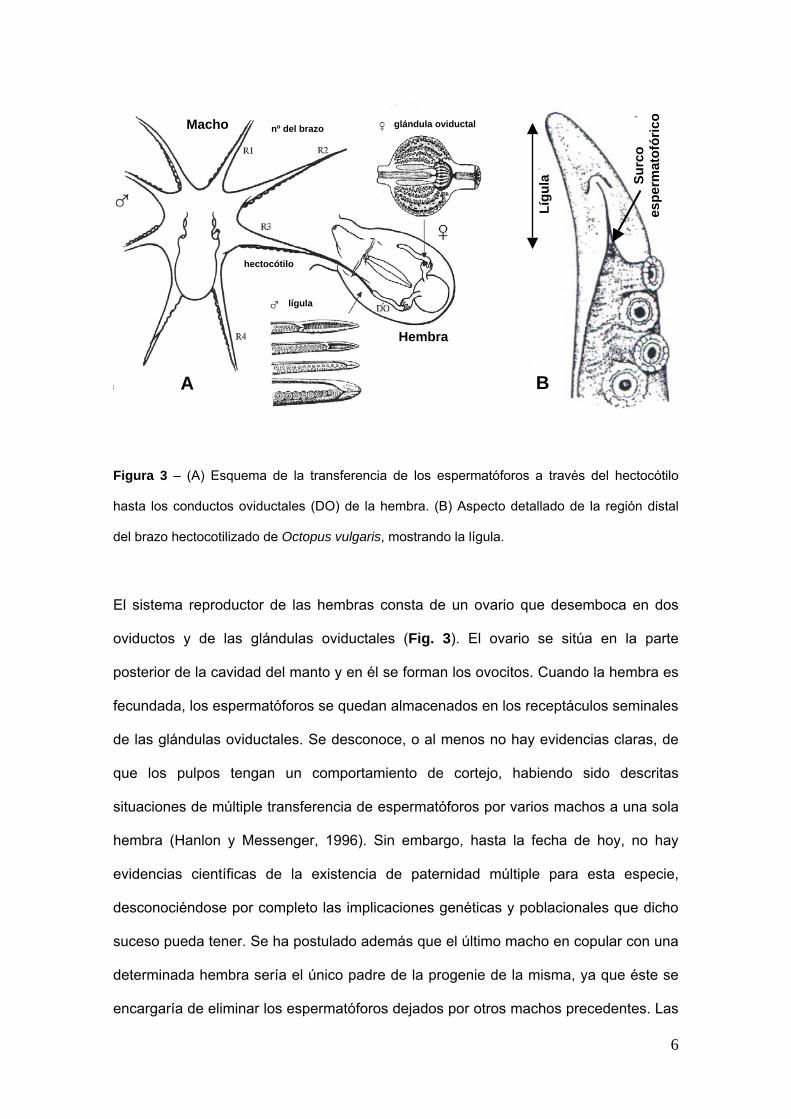
Surc
o es
perm
atof
oacuteric
o
Liacutegu
la
glaacutendula oviductal nordm del brazo
liacutegula
hectocoacutetilo
A B
Macho
Hembra
Figura 3 ndash (A) Esquema de la transferencia de los espermatoacuteforos a traveacutes del hectocoacutetilo
hasta los conductos oviductales (DO) de la hembra (B) Aspecto detallado de la regioacuten distal
del brazo hectocotilizado de Octopus vulgaris mostrando la liacutegula
El sistema reproductor de las hembras consta de un ovario que desemboca en dos
oviductos y de las glaacutendulas oviductales (Fig 3) El ovario se situacutea en la parte
posterior de la cavidad del manto y en eacutel se forman los ovocitos Cuando la hembra es
fecundada los espermatoacuteforos se quedan almacenados en los receptaacuteculos seminales
de las glaacutendulas oviductales Se desconoce o al menos no hay evidencias claras de
que los pulpos tengan un comportamiento de cortejo habiendo sido descritas
situaciones de muacuteltiple transferencia de espermatoacuteforos por varios machos a una sola
hembra (Hanlon y Messenger 1996) Sin embargo hasta la fecha de hoy no hay
evidencias cientiacuteficas de la existencia de paternidad muacuteltiple para esta especie
desconocieacutendose por completo las implicaciones geneacuteticas y poblacionales que dicho
suceso pueda tener Se ha postulado ademaacutes que el uacuteltimo macho en copular con una
determinada hembra seriacutea el uacutenico padre de la progenie de la misma ya que eacuteste se
encargariacutea de eliminar los espermatoacuteforos dejados por otros machos precedentes Las
6
hembras son semelparas (se reproducen una sola vez) desarrollaacutendose el ovario de
forma sincroacutenica La ovulacioacuten se da de una sola vez no habiendo desarrollo y
maduracioacuten de nuevos ovocitos despueacutes de la uacutenica puesta estimada en 100000 a
500000 huevos (Mangold 1987) La fecundacioacuten en los pulpos es interna y se da a
medida que los ovocitos van pasando por el oviducto proximal hasta la zona distal
fertilizaacutendose a su paso por la glaacutendula oviductal donde estaacuten alojados los
espermatoacuteforos La puesta de los huevos puede tardar entre 15 y 30 diacuteas
Posteriormente la hembra pasa a ejercer cuidados maternales como proteccioacuten contra
depredadores y limpieza y oxigenacioacuten de los huevos pudiendo este periodo
prolongarse de 25 a 125 diacuteas dependiendo de la temperatura del agua (Mangold y
Boletzky 1973) Durante este tiempo los huevos de pulpo han de pasar por varias
fases de desarrollo (Fig 4) las cuales han sido clasificadas en XX estadios por Naef
(1928) A lo largo de este periodo las hembras dejan de alimentarse pudiendo perder
hasta el 60 de su peso corporal inicial antes de haber iniciado la puesta (Wodinsky
1978) La eclosioacuten de los huevos libera las paralarvas (Fig 4) teacutermino propuesto por
Young y Harman (1988) que asiacute se denominan porque en realidad no van a sufrir una
verdadera metamorfosis antes de convertirse en juveniles aunque al contrario que en
la vida adulta son planctoacutenicas en sus primeros diacuteas de vida Una vez finalizado el
periodo de incubacioacuten y liberacioacuten de las paralarvas las hembras mueren El tamantildeo
del manto de las paralarvas recieacuten eclosionadas es de alrededor de 2 mm y en cada
brazo poseen 3 ventosas (Boletzky 1987 Villanueva 1995) Se estima que el periodo
de vida planctoacutenico de las paralarvas es de 30 a 60 diacuteas dependiendo de la
temperatura del agua y de la zona geograacutefica (Itami et al 1963 Mangold y Boletzky
1973 Villanueva et al 1995) Se cree que las paralarvas de pulpo al igual que otras
larvas de cefaloacutepodos estudiadas en su medio natural (Vecchione 1987 Passarella y
Hopkins 1991) se alimentan de pequentildeos crustaacuteceos que forman parte del
zooplancton Estas criaturas son depredadoras activas desde el primer diacutea de vida
(Iglesias et al 2006) a pesar de que poseen reservas vitelinas internas que les
7
permiten sobrevivir en ausencia de alimento durante algunos diacuteas (Boletzky 1975) La
absorcioacuten de determinados nutrientes por la piel tambieacuten ha sido sentildealada como un
factor importante en los primeros estadios de vida de las larvas de cefaloacutepodos (Lee
1994) mientras que en otras especies de pulpos como el Octopus dofleini se ha
descrito un comportamiento de alimentacioacuten neustoacutenico es decir las paralarvas se
colocan con los brazos haciacutea la superficie aprovechando la tensioacuten superficial del
agua alimentaacutendose de los detritos orgaacutenicos o presas de esta interfase (Marliave
1981)
A
DC
B
Figura 4 - Fotografiacuteas a la lupa de huevos de pulpo en diferentes estadios de desarrollo y de
una paralarva de pulpo (A) huevos de pulpo (asymp 2 x 1 mm) recieacuten depositados (B) huevos en
una etapa de desarrollo intermedia (estadio X-XII) (C) huevos en un estado de desarrollo
avanzado (estadio XVIII) (D) paralarva de pulpo recieacuten eclosionada (tamantildeo total asymp 3 mm)
8
De los trabajos realizados en laboratorio sobre cultivo de paralarvas de O vulgaris
(Villanueva 1995 Iglesias et al 2004 Carrasco et al 2006) se sabe que a medida
que eacutestas van creciendo la proporcioacuten de los brazos frente al manto va aumentando y
una vez alcanzado un determinado tamantildeo realizan el asentamiento en el fondo
convirtieacutendose asiacute en juveniles bentoacutenicos de morfologiacutea igual que los adultos El
crecimiento de los juveniles hasta la fase adulta puede tardar de 8 a 15 meses
dependiendo mayormente de la temperatura y de la disponibilidad de alimento
(Mangold y Boletzky 1973 Forsythe y Van Heukelem 1987 Semmens et al 2004
Leporati et al 2007) Al igual que muchos otros cefaloacutepodos O vulgaris es una
especie carniacutevora durante todo su ciclo de vida En las fases de juvenil hasta la edad
adulta los pulpos se alimentan de una gran variedad de presas que estaacuten
representadas por la mayor parte de los fila marinos (Mangold 1983) de los cuales
varias especies de crustaacuteceos peces y otros moluscos constituyen las presas maacutes
comunes siendo los cangrejos una de sus favoritas (Nixon 1987) Esta especie
presenta ademaacutes un comportamiento de canibalismo frecuente en el medio natural A
pesar del gran intereacutes que hay en el cultivo industrial de esta especie todaviacutea persiste
un importante cuello de botella por resolver el cultivo de las paralarvas planctoacutenicas
hasta la fase de asentamiento cuando eacutestas se convierten en juveniles bentoacutenicos
12 El principal cuello de botella del cultivo del pulpo (Octopus vulgaris)
Actualmente el principal problema que impide el desarrollo integral del cultivo del pulpo
es la elevada mortalidad que se observa en la fase planctoacutenica A pesar de los intentos
llevados a cabo por diferentes grupos de trabajo de varios paiacuteses en los que se han
utilizado distintas metodologiacuteas de alimentacioacuten y condiciones zooteacutecnicas pocos
fueron los experimentos en los que se ha logrado obtener juveniles bentoacutenicos
(revisado por Iglesias et al 2007) Algunos de los primeros trabajos llevados a cabo a
9
nivel mundial se realizaron en Japoacuten Itami et al (1963) utilizando zoeas de camaroacuten
(Palaemon serrifer) lograron tener pulpos juveniles al cabo de 45 diacuteas con una
supervivencia del 8 Hamazaki et al (1991) publicaba igualmente la obtencioacuten de
juveniles de pulpo bentoacutenicos en condiciones controladas empleando como alimento
vivo juveniles de Artemia (15-2 mm) enriquecidos con Nannochloropsis sp y
antildeadiendo esta misma microalga al tanque de cultivo alcanzando en uno de sus
experimentos en un tanque de 20 m3 un nuacutemero de 23700 juveniles bentoacutenicos
(supervivencia del 29) al cabo de 25 diacuteas de cultivo
Al comienzo de los antildeos 90 se iniciaron tambieacuten en Espantildea un conjunto de
importantes trabajos relacionados con el cultivo del pulpo O vulgaris Los primeros
trabajos publicados sobre el cultivo larvario de cefaloacutepodos en laboratorio han sido
desarrollados por Villanueva (1994 1995) que probando zoeas de distintos
crustaacuteceos decaacutepodos (Pagurus prideaux Liocarcinus depurator y Dardanus arrosor)
logroacute obtener pulpos bentoacutenicos confirmando a las zoeas como un alimento apropiado
para suministrar a larvas de cefaloacutepodos Sin embargo este autor ya poniacutea de
manifiesto la dificultad en obtener zoeas en momentos precisos y los altos costes de
mantenimiento y mano de obra que implicaba esta praacutectica Por otro lado se iniciaban
tambieacuten en Espantildea un conjunto de experimentos de engorde de pulpo a escala de
laboratorio para evaluar el potencial de crecimiento de esta especie afrontaacutendose
entre los antildeos 1997 y 2000 el problema del cultivo de la fase planctoacutenica En los
primeros ensayos realizados Iglesias et al (1999 2000) probaron distintos tipos de
alimento tales como zooplancton del medio natural (copeacutepodos zoeas de crustaacuteceos
y misidaacuteceos) ictioplancton huevos de peces micropellets Artemia y rotiacuteferos y maacutes
tarde zoeas de crustaacuteceos obtenidas en laboratorio a partir de stocks de reproductores
de cangrejo (Carcinus maenas) neacutecora (Necora puber) y camaroacuten (Palaemon
serratus) Sin embargo la mortalidad de las paralarvas fue casi total en los primeros
diacuteas de vida alcanzaacutendose supervivencias maacuteximas del 10 a los 32 diacuteas al emplear
como alimento metanauplios y a continuacioacuten Artemia adulta enriquecida con
10
microalgas pero sin que se lograse alcanzar la fase de pre-asentamiento de las
paralarvas En los intentos llevados a cabo en el IEO de Vigo y maacutes tarde en el Centro
de Experimentacioacuten Pesquera de Asturias se obtuvieron algunos ejemplares de
juveniles bentoacutenicos que han llegado a alcanzar la edad adulta utilizaacutendose como
dieta para suministrar a las paralarvas juveniles de Artemia enriquecidos con
microalgas complementados con zoeas de centolla Maja brachydactyla en momentos
de disponibilidad de eacutestas (Iglesias et al 2002 2004 Carrasco et al 2003 2006) Las
primeras pruebas en las que se ha empleado esta mezcla de presas vivas (Moxica et
al 2002) fueron llevadas a cabo en el IEO de Vigo en un tanque de 9 m3 al que se
aportoacute inicialmente nauplios de Artemia y maacutes tarde juveniles de Artemia (1-4 mm) a
una densidad de presas de 01 Artemia ml-1 complementados con zoeas a partir de la
tercera semana de cultivo (3000 a 5000 zoeas al diacutea) Al tanque se aportaba a diario
una mezcla de microalgas (un 40 de Isochrysis galbana un 40 de Tetraselmis
suecica y un 20 de Chaetoceros sp) con el fin de mantener las presas vivas
constantemente enriquecidas Aunque no se han obtenido pulpos bentoacutenicos en este
experimento los autores refirieron la obtencioacuten de paralarvas en fase de asentamiento
con elevado nuacutemero de ventosas en los brazos (17-18) y un peso seco de 92 plusmn 09
mg a los 52 diacuteas de vida La supervivencia en este experimento fue de 83 al mes de
vida y de 02 al cabo de 52 diacuteas En un experimento posterior en el que se redujo la
escala y se empleoacute un tanque de 1 m3 las paralarvas se alimentaron con Artemia (1-4
mm) enriquecida con Chlorella sp antildeadiendo diariamente al tanque de cultivo una
mezcla de microalgas (Chlorella sp Isochrysis galbana y Chaetoceros sp) Como
complemento de Artemia se suministraron zoeas de centollo (Maja brachydactyla)
cuatro veces a la semana a una concentracioacuten de 001-01 zoeas ml-1 alcanzaacutendose
asiacute una supervivencia del 315 al diacutea 40 y un reducido nuacutemero de juveniles
bentoacutenicos dos semanas maacutes tarde (Iglesias et al 2002 2004) El seguimiento de los
uacutenicos pulpos supervivientes de este experimento hasta la fase adulta y la obtencioacuten
de una puesta de huevos por parte de una hembra originoacute el cierre del ciclo de vida
11
del pulpo en laboratorio por primer vez (Iglesias et al 2004) De forma similar
Carrasco et al (2003 2006) lograron obtener en uno de sus experimentos pulpos
bentoacutenicos a los 60 diacuteas con una supervivencia del 34
Sin embargo la utilizacioacuten de zoeas de crustaacuteceos como alimento vivo supone un
elevado riesgo ya que no se controla la obtencioacuten de las mismas en momentos
precisos ni la cantidad a producir ademaacutes de los elevados costes que conlleva esta
praacutectica por la necesidad de disponer de maacutes recursos materiales fiacutesicos y de
personal (Navarro y Villanueva 2000) Artemia en cambio es un crustaacuteceo faacutecil de
producir los nauplios o metanauplios se obtienen en 24-72 h y su composicioacuten
bioquiacutemica se puede modular mediante teacutecnicas de enriquecimiento al ser un filtrador
continuo obligado y no selectivo La utilizacioacuten de Artemia como presa viva se
consolida por tanto como una de las mejores alternativas para el cultivo de
paralarvas aunque sea necesario seguir trabajando en la modulacioacuten de su valor
nutricional de acuerdo con los posibles requerimientos de las paralarvas
Varios autores han utilizado nauplios de Artemia enriquecidos con distintos productos
comerciales (Navarro y Villanueva 2000 2003 Villanueva et al 2004 Okumura et al
2005) en sus experimentos de cultivo de paralarvas de pulpo siendo el tamantildeo de
estas presas (05-12 mm) adecuado hasta determinado momento del cultivo Sin
embargo se ha demostrado que las paralarvas tienen una preferencia clara por
Artemia de mayor tamantildeo (14 mm) sobre la maacutes pequentildea (08 mm) desde los
primeros diacuteas de vida (Iglesias et al 2006) De entre los productos o suplementos
nutricionales empleados por diversos autores para enriquecer Artemia se encuentran
emulsiones lipiacutedicas de la gama Selco (INVE) u otros aceites de pescado vitaminas
harina de huevos de pescado (BASF) harinas de cereales o productos de la gama Ori
(Skretting) (Navarro y Villanueva 2000 2003 Villanueva et al 2002 Iglesias et al
2004 Okumura et al 2005 Izquierdo et al 2008) pero ninguno ha originado tasas de
supervivencia o pesos secos de las paralarvas semejantes a los obtenidos con la
utilizacioacuten de zoeas de crustaacuteceos Como alternativa a las zoeas o a la Artemia
12
tambieacuten se han probado otros tipos de presas vivas o dietas artificiales para
suministrar a las paralarvas aunque en todo caso este tipo de pruebas se hicieron
complementadas con Artemia enriquecida (revisado por Iglesias et al 2007)
Otro de los principales suplementos empleados muy a menudo por diversos autores
para enriquecercultivar Artemia son las microalgas de las que las maacutes comuacutenmente
utilizadas fueron Nannochloropsis sp Dunaliella viridis Tetraselmis suecica
Isochrysis galbana Chaetoceros sp Chlorella sp (Hamazaki et al 1991 Navarro y
Villanueva 2000 Moxica et al 2002 Iglesias et al 2002 2004 Carrasco et al
2003 2006 Moxica et al 2006)
Hay un cierto consenso entre los grupos de investigadores que se dedican al cultivo
larvario del pulpo sobre que los aspectos nutricionales estaacuten en la base de la elevada
mortalidad observada durante su cultivo (Iglesias et al 2007) Uno de los problemas
que dificulta la interpretacioacuten de los resultados descritos y el avance en la nutricioacuten del
cultivo larvario del pulpo es que muy pocos autores presentan datos de la
composicioacuten bioquiacutemica de las dietas utilizadas y de sus efectos sobre la composicioacuten
de las paralarvas (Moxica et al 2002 Navarro et al 2000 2003 Villanueva et al
2004 Okumura et al 2005) cuando esta informacioacuten es crucial para interpretar los
resultados obtenidos y asiacute poder relacionarlos con la supervivencia y el crecimiento de
las paralarvas
A pesar de los continuos intentos para cerrar el ciclo de vida del pulpo de forma
rentable la mortalidad de las paralarvas durante la fase planctoacutenica sigue siendo casi
total Entre las estrategias propuestas para superar este importante problema estaacuten la
mejora de la composicioacuten de Artemia mediante las teacutecnicas de enriquecimiento
apropiadas y la formulacioacuten de microdietas inertes (Iglesias et al 2007) Importa
pues reducir el tiempo necesario hasta la obtencioacuten de los juveniles de Artemia del
tamantildeo deseado y optimizar su composicioacuten bioquiacutemica con base en los posibles
requerimientos nutricionales de las paralarvas El crecimiento y la composicioacuten
bioquiacutemica de Artemia sp pueden ser fuertemente modificados mediante la utilizacioacuten
13
de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

hembras son semelparas (se reproducen una sola vez) desarrollaacutendose el ovario de
forma sincroacutenica La ovulacioacuten se da de una sola vez no habiendo desarrollo y
maduracioacuten de nuevos ovocitos despueacutes de la uacutenica puesta estimada en 100000 a
500000 huevos (Mangold 1987) La fecundacioacuten en los pulpos es interna y se da a
medida que los ovocitos van pasando por el oviducto proximal hasta la zona distal
fertilizaacutendose a su paso por la glaacutendula oviductal donde estaacuten alojados los
espermatoacuteforos La puesta de los huevos puede tardar entre 15 y 30 diacuteas
Posteriormente la hembra pasa a ejercer cuidados maternales como proteccioacuten contra
depredadores y limpieza y oxigenacioacuten de los huevos pudiendo este periodo
prolongarse de 25 a 125 diacuteas dependiendo de la temperatura del agua (Mangold y
Boletzky 1973) Durante este tiempo los huevos de pulpo han de pasar por varias
fases de desarrollo (Fig 4) las cuales han sido clasificadas en XX estadios por Naef
(1928) A lo largo de este periodo las hembras dejan de alimentarse pudiendo perder
hasta el 60 de su peso corporal inicial antes de haber iniciado la puesta (Wodinsky
1978) La eclosioacuten de los huevos libera las paralarvas (Fig 4) teacutermino propuesto por
Young y Harman (1988) que asiacute se denominan porque en realidad no van a sufrir una
verdadera metamorfosis antes de convertirse en juveniles aunque al contrario que en
la vida adulta son planctoacutenicas en sus primeros diacuteas de vida Una vez finalizado el
periodo de incubacioacuten y liberacioacuten de las paralarvas las hembras mueren El tamantildeo
del manto de las paralarvas recieacuten eclosionadas es de alrededor de 2 mm y en cada
brazo poseen 3 ventosas (Boletzky 1987 Villanueva 1995) Se estima que el periodo
de vida planctoacutenico de las paralarvas es de 30 a 60 diacuteas dependiendo de la
temperatura del agua y de la zona geograacutefica (Itami et al 1963 Mangold y Boletzky
1973 Villanueva et al 1995) Se cree que las paralarvas de pulpo al igual que otras
larvas de cefaloacutepodos estudiadas en su medio natural (Vecchione 1987 Passarella y
Hopkins 1991) se alimentan de pequentildeos crustaacuteceos que forman parte del
zooplancton Estas criaturas son depredadoras activas desde el primer diacutea de vida
(Iglesias et al 2006) a pesar de que poseen reservas vitelinas internas que les
7
permiten sobrevivir en ausencia de alimento durante algunos diacuteas (Boletzky 1975) La
absorcioacuten de determinados nutrientes por la piel tambieacuten ha sido sentildealada como un
factor importante en los primeros estadios de vida de las larvas de cefaloacutepodos (Lee
1994) mientras que en otras especies de pulpos como el Octopus dofleini se ha
descrito un comportamiento de alimentacioacuten neustoacutenico es decir las paralarvas se
colocan con los brazos haciacutea la superficie aprovechando la tensioacuten superficial del
agua alimentaacutendose de los detritos orgaacutenicos o presas de esta interfase (Marliave
1981)
A
DC
B
Figura 4 - Fotografiacuteas a la lupa de huevos de pulpo en diferentes estadios de desarrollo y de
una paralarva de pulpo (A) huevos de pulpo (asymp 2 x 1 mm) recieacuten depositados (B) huevos en
una etapa de desarrollo intermedia (estadio X-XII) (C) huevos en un estado de desarrollo
avanzado (estadio XVIII) (D) paralarva de pulpo recieacuten eclosionada (tamantildeo total asymp 3 mm)
8
De los trabajos realizados en laboratorio sobre cultivo de paralarvas de O vulgaris
(Villanueva 1995 Iglesias et al 2004 Carrasco et al 2006) se sabe que a medida
que eacutestas van creciendo la proporcioacuten de los brazos frente al manto va aumentando y
una vez alcanzado un determinado tamantildeo realizan el asentamiento en el fondo
convirtieacutendose asiacute en juveniles bentoacutenicos de morfologiacutea igual que los adultos El
crecimiento de los juveniles hasta la fase adulta puede tardar de 8 a 15 meses
dependiendo mayormente de la temperatura y de la disponibilidad de alimento
(Mangold y Boletzky 1973 Forsythe y Van Heukelem 1987 Semmens et al 2004
Leporati et al 2007) Al igual que muchos otros cefaloacutepodos O vulgaris es una
especie carniacutevora durante todo su ciclo de vida En las fases de juvenil hasta la edad
adulta los pulpos se alimentan de una gran variedad de presas que estaacuten
representadas por la mayor parte de los fila marinos (Mangold 1983) de los cuales
varias especies de crustaacuteceos peces y otros moluscos constituyen las presas maacutes
comunes siendo los cangrejos una de sus favoritas (Nixon 1987) Esta especie
presenta ademaacutes un comportamiento de canibalismo frecuente en el medio natural A
pesar del gran intereacutes que hay en el cultivo industrial de esta especie todaviacutea persiste
un importante cuello de botella por resolver el cultivo de las paralarvas planctoacutenicas
hasta la fase de asentamiento cuando eacutestas se convierten en juveniles bentoacutenicos
12 El principal cuello de botella del cultivo del pulpo (Octopus vulgaris)
Actualmente el principal problema que impide el desarrollo integral del cultivo del pulpo
es la elevada mortalidad que se observa en la fase planctoacutenica A pesar de los intentos
llevados a cabo por diferentes grupos de trabajo de varios paiacuteses en los que se han
utilizado distintas metodologiacuteas de alimentacioacuten y condiciones zooteacutecnicas pocos
fueron los experimentos en los que se ha logrado obtener juveniles bentoacutenicos
(revisado por Iglesias et al 2007) Algunos de los primeros trabajos llevados a cabo a
9
nivel mundial se realizaron en Japoacuten Itami et al (1963) utilizando zoeas de camaroacuten
(Palaemon serrifer) lograron tener pulpos juveniles al cabo de 45 diacuteas con una
supervivencia del 8 Hamazaki et al (1991) publicaba igualmente la obtencioacuten de
juveniles de pulpo bentoacutenicos en condiciones controladas empleando como alimento
vivo juveniles de Artemia (15-2 mm) enriquecidos con Nannochloropsis sp y
antildeadiendo esta misma microalga al tanque de cultivo alcanzando en uno de sus
experimentos en un tanque de 20 m3 un nuacutemero de 23700 juveniles bentoacutenicos
(supervivencia del 29) al cabo de 25 diacuteas de cultivo
Al comienzo de los antildeos 90 se iniciaron tambieacuten en Espantildea un conjunto de
importantes trabajos relacionados con el cultivo del pulpo O vulgaris Los primeros
trabajos publicados sobre el cultivo larvario de cefaloacutepodos en laboratorio han sido
desarrollados por Villanueva (1994 1995) que probando zoeas de distintos
crustaacuteceos decaacutepodos (Pagurus prideaux Liocarcinus depurator y Dardanus arrosor)
logroacute obtener pulpos bentoacutenicos confirmando a las zoeas como un alimento apropiado
para suministrar a larvas de cefaloacutepodos Sin embargo este autor ya poniacutea de
manifiesto la dificultad en obtener zoeas en momentos precisos y los altos costes de
mantenimiento y mano de obra que implicaba esta praacutectica Por otro lado se iniciaban
tambieacuten en Espantildea un conjunto de experimentos de engorde de pulpo a escala de
laboratorio para evaluar el potencial de crecimiento de esta especie afrontaacutendose
entre los antildeos 1997 y 2000 el problema del cultivo de la fase planctoacutenica En los
primeros ensayos realizados Iglesias et al (1999 2000) probaron distintos tipos de
alimento tales como zooplancton del medio natural (copeacutepodos zoeas de crustaacuteceos
y misidaacuteceos) ictioplancton huevos de peces micropellets Artemia y rotiacuteferos y maacutes
tarde zoeas de crustaacuteceos obtenidas en laboratorio a partir de stocks de reproductores
de cangrejo (Carcinus maenas) neacutecora (Necora puber) y camaroacuten (Palaemon
serratus) Sin embargo la mortalidad de las paralarvas fue casi total en los primeros
diacuteas de vida alcanzaacutendose supervivencias maacuteximas del 10 a los 32 diacuteas al emplear
como alimento metanauplios y a continuacioacuten Artemia adulta enriquecida con
10
microalgas pero sin que se lograse alcanzar la fase de pre-asentamiento de las
paralarvas En los intentos llevados a cabo en el IEO de Vigo y maacutes tarde en el Centro
de Experimentacioacuten Pesquera de Asturias se obtuvieron algunos ejemplares de
juveniles bentoacutenicos que han llegado a alcanzar la edad adulta utilizaacutendose como
dieta para suministrar a las paralarvas juveniles de Artemia enriquecidos con
microalgas complementados con zoeas de centolla Maja brachydactyla en momentos
de disponibilidad de eacutestas (Iglesias et al 2002 2004 Carrasco et al 2003 2006) Las
primeras pruebas en las que se ha empleado esta mezcla de presas vivas (Moxica et
al 2002) fueron llevadas a cabo en el IEO de Vigo en un tanque de 9 m3 al que se
aportoacute inicialmente nauplios de Artemia y maacutes tarde juveniles de Artemia (1-4 mm) a
una densidad de presas de 01 Artemia ml-1 complementados con zoeas a partir de la
tercera semana de cultivo (3000 a 5000 zoeas al diacutea) Al tanque se aportaba a diario
una mezcla de microalgas (un 40 de Isochrysis galbana un 40 de Tetraselmis
suecica y un 20 de Chaetoceros sp) con el fin de mantener las presas vivas
constantemente enriquecidas Aunque no se han obtenido pulpos bentoacutenicos en este
experimento los autores refirieron la obtencioacuten de paralarvas en fase de asentamiento
con elevado nuacutemero de ventosas en los brazos (17-18) y un peso seco de 92 plusmn 09
mg a los 52 diacuteas de vida La supervivencia en este experimento fue de 83 al mes de
vida y de 02 al cabo de 52 diacuteas En un experimento posterior en el que se redujo la
escala y se empleoacute un tanque de 1 m3 las paralarvas se alimentaron con Artemia (1-4
mm) enriquecida con Chlorella sp antildeadiendo diariamente al tanque de cultivo una
mezcla de microalgas (Chlorella sp Isochrysis galbana y Chaetoceros sp) Como
complemento de Artemia se suministraron zoeas de centollo (Maja brachydactyla)
cuatro veces a la semana a una concentracioacuten de 001-01 zoeas ml-1 alcanzaacutendose
asiacute una supervivencia del 315 al diacutea 40 y un reducido nuacutemero de juveniles
bentoacutenicos dos semanas maacutes tarde (Iglesias et al 2002 2004) El seguimiento de los
uacutenicos pulpos supervivientes de este experimento hasta la fase adulta y la obtencioacuten
de una puesta de huevos por parte de una hembra originoacute el cierre del ciclo de vida
11
del pulpo en laboratorio por primer vez (Iglesias et al 2004) De forma similar
Carrasco et al (2003 2006) lograron obtener en uno de sus experimentos pulpos
bentoacutenicos a los 60 diacuteas con una supervivencia del 34
Sin embargo la utilizacioacuten de zoeas de crustaacuteceos como alimento vivo supone un
elevado riesgo ya que no se controla la obtencioacuten de las mismas en momentos
precisos ni la cantidad a producir ademaacutes de los elevados costes que conlleva esta
praacutectica por la necesidad de disponer de maacutes recursos materiales fiacutesicos y de
personal (Navarro y Villanueva 2000) Artemia en cambio es un crustaacuteceo faacutecil de
producir los nauplios o metanauplios se obtienen en 24-72 h y su composicioacuten
bioquiacutemica se puede modular mediante teacutecnicas de enriquecimiento al ser un filtrador
continuo obligado y no selectivo La utilizacioacuten de Artemia como presa viva se
consolida por tanto como una de las mejores alternativas para el cultivo de
paralarvas aunque sea necesario seguir trabajando en la modulacioacuten de su valor
nutricional de acuerdo con los posibles requerimientos de las paralarvas
Varios autores han utilizado nauplios de Artemia enriquecidos con distintos productos
comerciales (Navarro y Villanueva 2000 2003 Villanueva et al 2004 Okumura et al
2005) en sus experimentos de cultivo de paralarvas de pulpo siendo el tamantildeo de
estas presas (05-12 mm) adecuado hasta determinado momento del cultivo Sin
embargo se ha demostrado que las paralarvas tienen una preferencia clara por
Artemia de mayor tamantildeo (14 mm) sobre la maacutes pequentildea (08 mm) desde los
primeros diacuteas de vida (Iglesias et al 2006) De entre los productos o suplementos
nutricionales empleados por diversos autores para enriquecer Artemia se encuentran
emulsiones lipiacutedicas de la gama Selco (INVE) u otros aceites de pescado vitaminas
harina de huevos de pescado (BASF) harinas de cereales o productos de la gama Ori
(Skretting) (Navarro y Villanueva 2000 2003 Villanueva et al 2002 Iglesias et al
2004 Okumura et al 2005 Izquierdo et al 2008) pero ninguno ha originado tasas de
supervivencia o pesos secos de las paralarvas semejantes a los obtenidos con la
utilizacioacuten de zoeas de crustaacuteceos Como alternativa a las zoeas o a la Artemia
12
tambieacuten se han probado otros tipos de presas vivas o dietas artificiales para
suministrar a las paralarvas aunque en todo caso este tipo de pruebas se hicieron
complementadas con Artemia enriquecida (revisado por Iglesias et al 2007)
Otro de los principales suplementos empleados muy a menudo por diversos autores
para enriquecercultivar Artemia son las microalgas de las que las maacutes comuacutenmente
utilizadas fueron Nannochloropsis sp Dunaliella viridis Tetraselmis suecica
Isochrysis galbana Chaetoceros sp Chlorella sp (Hamazaki et al 1991 Navarro y
Villanueva 2000 Moxica et al 2002 Iglesias et al 2002 2004 Carrasco et al
2003 2006 Moxica et al 2006)
Hay un cierto consenso entre los grupos de investigadores que se dedican al cultivo
larvario del pulpo sobre que los aspectos nutricionales estaacuten en la base de la elevada
mortalidad observada durante su cultivo (Iglesias et al 2007) Uno de los problemas
que dificulta la interpretacioacuten de los resultados descritos y el avance en la nutricioacuten del
cultivo larvario del pulpo es que muy pocos autores presentan datos de la
composicioacuten bioquiacutemica de las dietas utilizadas y de sus efectos sobre la composicioacuten
de las paralarvas (Moxica et al 2002 Navarro et al 2000 2003 Villanueva et al
2004 Okumura et al 2005) cuando esta informacioacuten es crucial para interpretar los
resultados obtenidos y asiacute poder relacionarlos con la supervivencia y el crecimiento de
las paralarvas
A pesar de los continuos intentos para cerrar el ciclo de vida del pulpo de forma
rentable la mortalidad de las paralarvas durante la fase planctoacutenica sigue siendo casi
total Entre las estrategias propuestas para superar este importante problema estaacuten la
mejora de la composicioacuten de Artemia mediante las teacutecnicas de enriquecimiento
apropiadas y la formulacioacuten de microdietas inertes (Iglesias et al 2007) Importa
pues reducir el tiempo necesario hasta la obtencioacuten de los juveniles de Artemia del
tamantildeo deseado y optimizar su composicioacuten bioquiacutemica con base en los posibles
requerimientos nutricionales de las paralarvas El crecimiento y la composicioacuten
bioquiacutemica de Artemia sp pueden ser fuertemente modificados mediante la utilizacioacuten
13
de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

permiten sobrevivir en ausencia de alimento durante algunos diacuteas (Boletzky 1975) La
absorcioacuten de determinados nutrientes por la piel tambieacuten ha sido sentildealada como un
factor importante en los primeros estadios de vida de las larvas de cefaloacutepodos (Lee
1994) mientras que en otras especies de pulpos como el Octopus dofleini se ha
descrito un comportamiento de alimentacioacuten neustoacutenico es decir las paralarvas se
colocan con los brazos haciacutea la superficie aprovechando la tensioacuten superficial del
agua alimentaacutendose de los detritos orgaacutenicos o presas de esta interfase (Marliave
1981)
A
DC
B
Figura 4 - Fotografiacuteas a la lupa de huevos de pulpo en diferentes estadios de desarrollo y de
una paralarva de pulpo (A) huevos de pulpo (asymp 2 x 1 mm) recieacuten depositados (B) huevos en
una etapa de desarrollo intermedia (estadio X-XII) (C) huevos en un estado de desarrollo
avanzado (estadio XVIII) (D) paralarva de pulpo recieacuten eclosionada (tamantildeo total asymp 3 mm)
8
De los trabajos realizados en laboratorio sobre cultivo de paralarvas de O vulgaris
(Villanueva 1995 Iglesias et al 2004 Carrasco et al 2006) se sabe que a medida
que eacutestas van creciendo la proporcioacuten de los brazos frente al manto va aumentando y
una vez alcanzado un determinado tamantildeo realizan el asentamiento en el fondo
convirtieacutendose asiacute en juveniles bentoacutenicos de morfologiacutea igual que los adultos El
crecimiento de los juveniles hasta la fase adulta puede tardar de 8 a 15 meses
dependiendo mayormente de la temperatura y de la disponibilidad de alimento
(Mangold y Boletzky 1973 Forsythe y Van Heukelem 1987 Semmens et al 2004
Leporati et al 2007) Al igual que muchos otros cefaloacutepodos O vulgaris es una
especie carniacutevora durante todo su ciclo de vida En las fases de juvenil hasta la edad
adulta los pulpos se alimentan de una gran variedad de presas que estaacuten
representadas por la mayor parte de los fila marinos (Mangold 1983) de los cuales
varias especies de crustaacuteceos peces y otros moluscos constituyen las presas maacutes
comunes siendo los cangrejos una de sus favoritas (Nixon 1987) Esta especie
presenta ademaacutes un comportamiento de canibalismo frecuente en el medio natural A
pesar del gran intereacutes que hay en el cultivo industrial de esta especie todaviacutea persiste
un importante cuello de botella por resolver el cultivo de las paralarvas planctoacutenicas
hasta la fase de asentamiento cuando eacutestas se convierten en juveniles bentoacutenicos
12 El principal cuello de botella del cultivo del pulpo (Octopus vulgaris)
Actualmente el principal problema que impide el desarrollo integral del cultivo del pulpo
es la elevada mortalidad que se observa en la fase planctoacutenica A pesar de los intentos
llevados a cabo por diferentes grupos de trabajo de varios paiacuteses en los que se han
utilizado distintas metodologiacuteas de alimentacioacuten y condiciones zooteacutecnicas pocos
fueron los experimentos en los que se ha logrado obtener juveniles bentoacutenicos
(revisado por Iglesias et al 2007) Algunos de los primeros trabajos llevados a cabo a
9
nivel mundial se realizaron en Japoacuten Itami et al (1963) utilizando zoeas de camaroacuten
(Palaemon serrifer) lograron tener pulpos juveniles al cabo de 45 diacuteas con una
supervivencia del 8 Hamazaki et al (1991) publicaba igualmente la obtencioacuten de
juveniles de pulpo bentoacutenicos en condiciones controladas empleando como alimento
vivo juveniles de Artemia (15-2 mm) enriquecidos con Nannochloropsis sp y
antildeadiendo esta misma microalga al tanque de cultivo alcanzando en uno de sus
experimentos en un tanque de 20 m3 un nuacutemero de 23700 juveniles bentoacutenicos
(supervivencia del 29) al cabo de 25 diacuteas de cultivo
Al comienzo de los antildeos 90 se iniciaron tambieacuten en Espantildea un conjunto de
importantes trabajos relacionados con el cultivo del pulpo O vulgaris Los primeros
trabajos publicados sobre el cultivo larvario de cefaloacutepodos en laboratorio han sido
desarrollados por Villanueva (1994 1995) que probando zoeas de distintos
crustaacuteceos decaacutepodos (Pagurus prideaux Liocarcinus depurator y Dardanus arrosor)
logroacute obtener pulpos bentoacutenicos confirmando a las zoeas como un alimento apropiado
para suministrar a larvas de cefaloacutepodos Sin embargo este autor ya poniacutea de
manifiesto la dificultad en obtener zoeas en momentos precisos y los altos costes de
mantenimiento y mano de obra que implicaba esta praacutectica Por otro lado se iniciaban
tambieacuten en Espantildea un conjunto de experimentos de engorde de pulpo a escala de
laboratorio para evaluar el potencial de crecimiento de esta especie afrontaacutendose
entre los antildeos 1997 y 2000 el problema del cultivo de la fase planctoacutenica En los
primeros ensayos realizados Iglesias et al (1999 2000) probaron distintos tipos de
alimento tales como zooplancton del medio natural (copeacutepodos zoeas de crustaacuteceos
y misidaacuteceos) ictioplancton huevos de peces micropellets Artemia y rotiacuteferos y maacutes
tarde zoeas de crustaacuteceos obtenidas en laboratorio a partir de stocks de reproductores
de cangrejo (Carcinus maenas) neacutecora (Necora puber) y camaroacuten (Palaemon
serratus) Sin embargo la mortalidad de las paralarvas fue casi total en los primeros
diacuteas de vida alcanzaacutendose supervivencias maacuteximas del 10 a los 32 diacuteas al emplear
como alimento metanauplios y a continuacioacuten Artemia adulta enriquecida con
10
microalgas pero sin que se lograse alcanzar la fase de pre-asentamiento de las
paralarvas En los intentos llevados a cabo en el IEO de Vigo y maacutes tarde en el Centro
de Experimentacioacuten Pesquera de Asturias se obtuvieron algunos ejemplares de
juveniles bentoacutenicos que han llegado a alcanzar la edad adulta utilizaacutendose como
dieta para suministrar a las paralarvas juveniles de Artemia enriquecidos con
microalgas complementados con zoeas de centolla Maja brachydactyla en momentos
de disponibilidad de eacutestas (Iglesias et al 2002 2004 Carrasco et al 2003 2006) Las
primeras pruebas en las que se ha empleado esta mezcla de presas vivas (Moxica et
al 2002) fueron llevadas a cabo en el IEO de Vigo en un tanque de 9 m3 al que se
aportoacute inicialmente nauplios de Artemia y maacutes tarde juveniles de Artemia (1-4 mm) a
una densidad de presas de 01 Artemia ml-1 complementados con zoeas a partir de la
tercera semana de cultivo (3000 a 5000 zoeas al diacutea) Al tanque se aportaba a diario
una mezcla de microalgas (un 40 de Isochrysis galbana un 40 de Tetraselmis
suecica y un 20 de Chaetoceros sp) con el fin de mantener las presas vivas
constantemente enriquecidas Aunque no se han obtenido pulpos bentoacutenicos en este
experimento los autores refirieron la obtencioacuten de paralarvas en fase de asentamiento
con elevado nuacutemero de ventosas en los brazos (17-18) y un peso seco de 92 plusmn 09
mg a los 52 diacuteas de vida La supervivencia en este experimento fue de 83 al mes de
vida y de 02 al cabo de 52 diacuteas En un experimento posterior en el que se redujo la
escala y se empleoacute un tanque de 1 m3 las paralarvas se alimentaron con Artemia (1-4
mm) enriquecida con Chlorella sp antildeadiendo diariamente al tanque de cultivo una
mezcla de microalgas (Chlorella sp Isochrysis galbana y Chaetoceros sp) Como
complemento de Artemia se suministraron zoeas de centollo (Maja brachydactyla)
cuatro veces a la semana a una concentracioacuten de 001-01 zoeas ml-1 alcanzaacutendose
asiacute una supervivencia del 315 al diacutea 40 y un reducido nuacutemero de juveniles
bentoacutenicos dos semanas maacutes tarde (Iglesias et al 2002 2004) El seguimiento de los
uacutenicos pulpos supervivientes de este experimento hasta la fase adulta y la obtencioacuten
de una puesta de huevos por parte de una hembra originoacute el cierre del ciclo de vida
11
del pulpo en laboratorio por primer vez (Iglesias et al 2004) De forma similar
Carrasco et al (2003 2006) lograron obtener en uno de sus experimentos pulpos
bentoacutenicos a los 60 diacuteas con una supervivencia del 34
Sin embargo la utilizacioacuten de zoeas de crustaacuteceos como alimento vivo supone un
elevado riesgo ya que no se controla la obtencioacuten de las mismas en momentos
precisos ni la cantidad a producir ademaacutes de los elevados costes que conlleva esta
praacutectica por la necesidad de disponer de maacutes recursos materiales fiacutesicos y de
personal (Navarro y Villanueva 2000) Artemia en cambio es un crustaacuteceo faacutecil de
producir los nauplios o metanauplios se obtienen en 24-72 h y su composicioacuten
bioquiacutemica se puede modular mediante teacutecnicas de enriquecimiento al ser un filtrador
continuo obligado y no selectivo La utilizacioacuten de Artemia como presa viva se
consolida por tanto como una de las mejores alternativas para el cultivo de
paralarvas aunque sea necesario seguir trabajando en la modulacioacuten de su valor
nutricional de acuerdo con los posibles requerimientos de las paralarvas
Varios autores han utilizado nauplios de Artemia enriquecidos con distintos productos
comerciales (Navarro y Villanueva 2000 2003 Villanueva et al 2004 Okumura et al
2005) en sus experimentos de cultivo de paralarvas de pulpo siendo el tamantildeo de
estas presas (05-12 mm) adecuado hasta determinado momento del cultivo Sin
embargo se ha demostrado que las paralarvas tienen una preferencia clara por
Artemia de mayor tamantildeo (14 mm) sobre la maacutes pequentildea (08 mm) desde los
primeros diacuteas de vida (Iglesias et al 2006) De entre los productos o suplementos
nutricionales empleados por diversos autores para enriquecer Artemia se encuentran
emulsiones lipiacutedicas de la gama Selco (INVE) u otros aceites de pescado vitaminas
harina de huevos de pescado (BASF) harinas de cereales o productos de la gama Ori
(Skretting) (Navarro y Villanueva 2000 2003 Villanueva et al 2002 Iglesias et al
2004 Okumura et al 2005 Izquierdo et al 2008) pero ninguno ha originado tasas de
supervivencia o pesos secos de las paralarvas semejantes a los obtenidos con la
utilizacioacuten de zoeas de crustaacuteceos Como alternativa a las zoeas o a la Artemia
12
tambieacuten se han probado otros tipos de presas vivas o dietas artificiales para
suministrar a las paralarvas aunque en todo caso este tipo de pruebas se hicieron
complementadas con Artemia enriquecida (revisado por Iglesias et al 2007)
Otro de los principales suplementos empleados muy a menudo por diversos autores
para enriquecercultivar Artemia son las microalgas de las que las maacutes comuacutenmente
utilizadas fueron Nannochloropsis sp Dunaliella viridis Tetraselmis suecica
Isochrysis galbana Chaetoceros sp Chlorella sp (Hamazaki et al 1991 Navarro y
Villanueva 2000 Moxica et al 2002 Iglesias et al 2002 2004 Carrasco et al
2003 2006 Moxica et al 2006)
Hay un cierto consenso entre los grupos de investigadores que se dedican al cultivo
larvario del pulpo sobre que los aspectos nutricionales estaacuten en la base de la elevada
mortalidad observada durante su cultivo (Iglesias et al 2007) Uno de los problemas
que dificulta la interpretacioacuten de los resultados descritos y el avance en la nutricioacuten del
cultivo larvario del pulpo es que muy pocos autores presentan datos de la
composicioacuten bioquiacutemica de las dietas utilizadas y de sus efectos sobre la composicioacuten
de las paralarvas (Moxica et al 2002 Navarro et al 2000 2003 Villanueva et al
2004 Okumura et al 2005) cuando esta informacioacuten es crucial para interpretar los
resultados obtenidos y asiacute poder relacionarlos con la supervivencia y el crecimiento de
las paralarvas
A pesar de los continuos intentos para cerrar el ciclo de vida del pulpo de forma
rentable la mortalidad de las paralarvas durante la fase planctoacutenica sigue siendo casi
total Entre las estrategias propuestas para superar este importante problema estaacuten la
mejora de la composicioacuten de Artemia mediante las teacutecnicas de enriquecimiento
apropiadas y la formulacioacuten de microdietas inertes (Iglesias et al 2007) Importa
pues reducir el tiempo necesario hasta la obtencioacuten de los juveniles de Artemia del
tamantildeo deseado y optimizar su composicioacuten bioquiacutemica con base en los posibles
requerimientos nutricionales de las paralarvas El crecimiento y la composicioacuten
bioquiacutemica de Artemia sp pueden ser fuertemente modificados mediante la utilizacioacuten
13
de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

De los trabajos realizados en laboratorio sobre cultivo de paralarvas de O vulgaris
(Villanueva 1995 Iglesias et al 2004 Carrasco et al 2006) se sabe que a medida
que eacutestas van creciendo la proporcioacuten de los brazos frente al manto va aumentando y
una vez alcanzado un determinado tamantildeo realizan el asentamiento en el fondo
convirtieacutendose asiacute en juveniles bentoacutenicos de morfologiacutea igual que los adultos El
crecimiento de los juveniles hasta la fase adulta puede tardar de 8 a 15 meses
dependiendo mayormente de la temperatura y de la disponibilidad de alimento
(Mangold y Boletzky 1973 Forsythe y Van Heukelem 1987 Semmens et al 2004
Leporati et al 2007) Al igual que muchos otros cefaloacutepodos O vulgaris es una
especie carniacutevora durante todo su ciclo de vida En las fases de juvenil hasta la edad
adulta los pulpos se alimentan de una gran variedad de presas que estaacuten
representadas por la mayor parte de los fila marinos (Mangold 1983) de los cuales
varias especies de crustaacuteceos peces y otros moluscos constituyen las presas maacutes
comunes siendo los cangrejos una de sus favoritas (Nixon 1987) Esta especie
presenta ademaacutes un comportamiento de canibalismo frecuente en el medio natural A
pesar del gran intereacutes que hay en el cultivo industrial de esta especie todaviacutea persiste
un importante cuello de botella por resolver el cultivo de las paralarvas planctoacutenicas
hasta la fase de asentamiento cuando eacutestas se convierten en juveniles bentoacutenicos
12 El principal cuello de botella del cultivo del pulpo (Octopus vulgaris)
Actualmente el principal problema que impide el desarrollo integral del cultivo del pulpo
es la elevada mortalidad que se observa en la fase planctoacutenica A pesar de los intentos
llevados a cabo por diferentes grupos de trabajo de varios paiacuteses en los que se han
utilizado distintas metodologiacuteas de alimentacioacuten y condiciones zooteacutecnicas pocos
fueron los experimentos en los que se ha logrado obtener juveniles bentoacutenicos
(revisado por Iglesias et al 2007) Algunos de los primeros trabajos llevados a cabo a
9
nivel mundial se realizaron en Japoacuten Itami et al (1963) utilizando zoeas de camaroacuten
(Palaemon serrifer) lograron tener pulpos juveniles al cabo de 45 diacuteas con una
supervivencia del 8 Hamazaki et al (1991) publicaba igualmente la obtencioacuten de
juveniles de pulpo bentoacutenicos en condiciones controladas empleando como alimento
vivo juveniles de Artemia (15-2 mm) enriquecidos con Nannochloropsis sp y
antildeadiendo esta misma microalga al tanque de cultivo alcanzando en uno de sus
experimentos en un tanque de 20 m3 un nuacutemero de 23700 juveniles bentoacutenicos
(supervivencia del 29) al cabo de 25 diacuteas de cultivo
Al comienzo de los antildeos 90 se iniciaron tambieacuten en Espantildea un conjunto de
importantes trabajos relacionados con el cultivo del pulpo O vulgaris Los primeros
trabajos publicados sobre el cultivo larvario de cefaloacutepodos en laboratorio han sido
desarrollados por Villanueva (1994 1995) que probando zoeas de distintos
crustaacuteceos decaacutepodos (Pagurus prideaux Liocarcinus depurator y Dardanus arrosor)
logroacute obtener pulpos bentoacutenicos confirmando a las zoeas como un alimento apropiado
para suministrar a larvas de cefaloacutepodos Sin embargo este autor ya poniacutea de
manifiesto la dificultad en obtener zoeas en momentos precisos y los altos costes de
mantenimiento y mano de obra que implicaba esta praacutectica Por otro lado se iniciaban
tambieacuten en Espantildea un conjunto de experimentos de engorde de pulpo a escala de
laboratorio para evaluar el potencial de crecimiento de esta especie afrontaacutendose
entre los antildeos 1997 y 2000 el problema del cultivo de la fase planctoacutenica En los
primeros ensayos realizados Iglesias et al (1999 2000) probaron distintos tipos de
alimento tales como zooplancton del medio natural (copeacutepodos zoeas de crustaacuteceos
y misidaacuteceos) ictioplancton huevos de peces micropellets Artemia y rotiacuteferos y maacutes
tarde zoeas de crustaacuteceos obtenidas en laboratorio a partir de stocks de reproductores
de cangrejo (Carcinus maenas) neacutecora (Necora puber) y camaroacuten (Palaemon
serratus) Sin embargo la mortalidad de las paralarvas fue casi total en los primeros
diacuteas de vida alcanzaacutendose supervivencias maacuteximas del 10 a los 32 diacuteas al emplear
como alimento metanauplios y a continuacioacuten Artemia adulta enriquecida con
10
microalgas pero sin que se lograse alcanzar la fase de pre-asentamiento de las
paralarvas En los intentos llevados a cabo en el IEO de Vigo y maacutes tarde en el Centro
de Experimentacioacuten Pesquera de Asturias se obtuvieron algunos ejemplares de
juveniles bentoacutenicos que han llegado a alcanzar la edad adulta utilizaacutendose como
dieta para suministrar a las paralarvas juveniles de Artemia enriquecidos con
microalgas complementados con zoeas de centolla Maja brachydactyla en momentos
de disponibilidad de eacutestas (Iglesias et al 2002 2004 Carrasco et al 2003 2006) Las
primeras pruebas en las que se ha empleado esta mezcla de presas vivas (Moxica et
al 2002) fueron llevadas a cabo en el IEO de Vigo en un tanque de 9 m3 al que se
aportoacute inicialmente nauplios de Artemia y maacutes tarde juveniles de Artemia (1-4 mm) a
una densidad de presas de 01 Artemia ml-1 complementados con zoeas a partir de la
tercera semana de cultivo (3000 a 5000 zoeas al diacutea) Al tanque se aportaba a diario
una mezcla de microalgas (un 40 de Isochrysis galbana un 40 de Tetraselmis
suecica y un 20 de Chaetoceros sp) con el fin de mantener las presas vivas
constantemente enriquecidas Aunque no se han obtenido pulpos bentoacutenicos en este
experimento los autores refirieron la obtencioacuten de paralarvas en fase de asentamiento
con elevado nuacutemero de ventosas en los brazos (17-18) y un peso seco de 92 plusmn 09
mg a los 52 diacuteas de vida La supervivencia en este experimento fue de 83 al mes de
vida y de 02 al cabo de 52 diacuteas En un experimento posterior en el que se redujo la
escala y se empleoacute un tanque de 1 m3 las paralarvas se alimentaron con Artemia (1-4
mm) enriquecida con Chlorella sp antildeadiendo diariamente al tanque de cultivo una
mezcla de microalgas (Chlorella sp Isochrysis galbana y Chaetoceros sp) Como
complemento de Artemia se suministraron zoeas de centollo (Maja brachydactyla)
cuatro veces a la semana a una concentracioacuten de 001-01 zoeas ml-1 alcanzaacutendose
asiacute una supervivencia del 315 al diacutea 40 y un reducido nuacutemero de juveniles
bentoacutenicos dos semanas maacutes tarde (Iglesias et al 2002 2004) El seguimiento de los
uacutenicos pulpos supervivientes de este experimento hasta la fase adulta y la obtencioacuten
de una puesta de huevos por parte de una hembra originoacute el cierre del ciclo de vida
11
del pulpo en laboratorio por primer vez (Iglesias et al 2004) De forma similar
Carrasco et al (2003 2006) lograron obtener en uno de sus experimentos pulpos
bentoacutenicos a los 60 diacuteas con una supervivencia del 34
Sin embargo la utilizacioacuten de zoeas de crustaacuteceos como alimento vivo supone un
elevado riesgo ya que no se controla la obtencioacuten de las mismas en momentos
precisos ni la cantidad a producir ademaacutes de los elevados costes que conlleva esta
praacutectica por la necesidad de disponer de maacutes recursos materiales fiacutesicos y de
personal (Navarro y Villanueva 2000) Artemia en cambio es un crustaacuteceo faacutecil de
producir los nauplios o metanauplios se obtienen en 24-72 h y su composicioacuten
bioquiacutemica se puede modular mediante teacutecnicas de enriquecimiento al ser un filtrador
continuo obligado y no selectivo La utilizacioacuten de Artemia como presa viva se
consolida por tanto como una de las mejores alternativas para el cultivo de
paralarvas aunque sea necesario seguir trabajando en la modulacioacuten de su valor
nutricional de acuerdo con los posibles requerimientos de las paralarvas
Varios autores han utilizado nauplios de Artemia enriquecidos con distintos productos
comerciales (Navarro y Villanueva 2000 2003 Villanueva et al 2004 Okumura et al
2005) en sus experimentos de cultivo de paralarvas de pulpo siendo el tamantildeo de
estas presas (05-12 mm) adecuado hasta determinado momento del cultivo Sin
embargo se ha demostrado que las paralarvas tienen una preferencia clara por
Artemia de mayor tamantildeo (14 mm) sobre la maacutes pequentildea (08 mm) desde los
primeros diacuteas de vida (Iglesias et al 2006) De entre los productos o suplementos
nutricionales empleados por diversos autores para enriquecer Artemia se encuentran
emulsiones lipiacutedicas de la gama Selco (INVE) u otros aceites de pescado vitaminas
harina de huevos de pescado (BASF) harinas de cereales o productos de la gama Ori
(Skretting) (Navarro y Villanueva 2000 2003 Villanueva et al 2002 Iglesias et al
2004 Okumura et al 2005 Izquierdo et al 2008) pero ninguno ha originado tasas de
supervivencia o pesos secos de las paralarvas semejantes a los obtenidos con la
utilizacioacuten de zoeas de crustaacuteceos Como alternativa a las zoeas o a la Artemia
12
tambieacuten se han probado otros tipos de presas vivas o dietas artificiales para
suministrar a las paralarvas aunque en todo caso este tipo de pruebas se hicieron
complementadas con Artemia enriquecida (revisado por Iglesias et al 2007)
Otro de los principales suplementos empleados muy a menudo por diversos autores
para enriquecercultivar Artemia son las microalgas de las que las maacutes comuacutenmente
utilizadas fueron Nannochloropsis sp Dunaliella viridis Tetraselmis suecica
Isochrysis galbana Chaetoceros sp Chlorella sp (Hamazaki et al 1991 Navarro y
Villanueva 2000 Moxica et al 2002 Iglesias et al 2002 2004 Carrasco et al
2003 2006 Moxica et al 2006)
Hay un cierto consenso entre los grupos de investigadores que se dedican al cultivo
larvario del pulpo sobre que los aspectos nutricionales estaacuten en la base de la elevada
mortalidad observada durante su cultivo (Iglesias et al 2007) Uno de los problemas
que dificulta la interpretacioacuten de los resultados descritos y el avance en la nutricioacuten del
cultivo larvario del pulpo es que muy pocos autores presentan datos de la
composicioacuten bioquiacutemica de las dietas utilizadas y de sus efectos sobre la composicioacuten
de las paralarvas (Moxica et al 2002 Navarro et al 2000 2003 Villanueva et al
2004 Okumura et al 2005) cuando esta informacioacuten es crucial para interpretar los
resultados obtenidos y asiacute poder relacionarlos con la supervivencia y el crecimiento de
las paralarvas
A pesar de los continuos intentos para cerrar el ciclo de vida del pulpo de forma
rentable la mortalidad de las paralarvas durante la fase planctoacutenica sigue siendo casi
total Entre las estrategias propuestas para superar este importante problema estaacuten la
mejora de la composicioacuten de Artemia mediante las teacutecnicas de enriquecimiento
apropiadas y la formulacioacuten de microdietas inertes (Iglesias et al 2007) Importa
pues reducir el tiempo necesario hasta la obtencioacuten de los juveniles de Artemia del
tamantildeo deseado y optimizar su composicioacuten bioquiacutemica con base en los posibles
requerimientos nutricionales de las paralarvas El crecimiento y la composicioacuten
bioquiacutemica de Artemia sp pueden ser fuertemente modificados mediante la utilizacioacuten
13
de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

nivel mundial se realizaron en Japoacuten Itami et al (1963) utilizando zoeas de camaroacuten
(Palaemon serrifer) lograron tener pulpos juveniles al cabo de 45 diacuteas con una
supervivencia del 8 Hamazaki et al (1991) publicaba igualmente la obtencioacuten de
juveniles de pulpo bentoacutenicos en condiciones controladas empleando como alimento
vivo juveniles de Artemia (15-2 mm) enriquecidos con Nannochloropsis sp y
antildeadiendo esta misma microalga al tanque de cultivo alcanzando en uno de sus
experimentos en un tanque de 20 m3 un nuacutemero de 23700 juveniles bentoacutenicos
(supervivencia del 29) al cabo de 25 diacuteas de cultivo
Al comienzo de los antildeos 90 se iniciaron tambieacuten en Espantildea un conjunto de
importantes trabajos relacionados con el cultivo del pulpo O vulgaris Los primeros
trabajos publicados sobre el cultivo larvario de cefaloacutepodos en laboratorio han sido
desarrollados por Villanueva (1994 1995) que probando zoeas de distintos
crustaacuteceos decaacutepodos (Pagurus prideaux Liocarcinus depurator y Dardanus arrosor)
logroacute obtener pulpos bentoacutenicos confirmando a las zoeas como un alimento apropiado
para suministrar a larvas de cefaloacutepodos Sin embargo este autor ya poniacutea de
manifiesto la dificultad en obtener zoeas en momentos precisos y los altos costes de
mantenimiento y mano de obra que implicaba esta praacutectica Por otro lado se iniciaban
tambieacuten en Espantildea un conjunto de experimentos de engorde de pulpo a escala de
laboratorio para evaluar el potencial de crecimiento de esta especie afrontaacutendose
entre los antildeos 1997 y 2000 el problema del cultivo de la fase planctoacutenica En los
primeros ensayos realizados Iglesias et al (1999 2000) probaron distintos tipos de
alimento tales como zooplancton del medio natural (copeacutepodos zoeas de crustaacuteceos
y misidaacuteceos) ictioplancton huevos de peces micropellets Artemia y rotiacuteferos y maacutes
tarde zoeas de crustaacuteceos obtenidas en laboratorio a partir de stocks de reproductores
de cangrejo (Carcinus maenas) neacutecora (Necora puber) y camaroacuten (Palaemon
serratus) Sin embargo la mortalidad de las paralarvas fue casi total en los primeros
diacuteas de vida alcanzaacutendose supervivencias maacuteximas del 10 a los 32 diacuteas al emplear
como alimento metanauplios y a continuacioacuten Artemia adulta enriquecida con
10
microalgas pero sin que se lograse alcanzar la fase de pre-asentamiento de las
paralarvas En los intentos llevados a cabo en el IEO de Vigo y maacutes tarde en el Centro
de Experimentacioacuten Pesquera de Asturias se obtuvieron algunos ejemplares de
juveniles bentoacutenicos que han llegado a alcanzar la edad adulta utilizaacutendose como
dieta para suministrar a las paralarvas juveniles de Artemia enriquecidos con
microalgas complementados con zoeas de centolla Maja brachydactyla en momentos
de disponibilidad de eacutestas (Iglesias et al 2002 2004 Carrasco et al 2003 2006) Las
primeras pruebas en las que se ha empleado esta mezcla de presas vivas (Moxica et
al 2002) fueron llevadas a cabo en el IEO de Vigo en un tanque de 9 m3 al que se
aportoacute inicialmente nauplios de Artemia y maacutes tarde juveniles de Artemia (1-4 mm) a
una densidad de presas de 01 Artemia ml-1 complementados con zoeas a partir de la
tercera semana de cultivo (3000 a 5000 zoeas al diacutea) Al tanque se aportaba a diario
una mezcla de microalgas (un 40 de Isochrysis galbana un 40 de Tetraselmis
suecica y un 20 de Chaetoceros sp) con el fin de mantener las presas vivas
constantemente enriquecidas Aunque no se han obtenido pulpos bentoacutenicos en este
experimento los autores refirieron la obtencioacuten de paralarvas en fase de asentamiento
con elevado nuacutemero de ventosas en los brazos (17-18) y un peso seco de 92 plusmn 09
mg a los 52 diacuteas de vida La supervivencia en este experimento fue de 83 al mes de
vida y de 02 al cabo de 52 diacuteas En un experimento posterior en el que se redujo la
escala y se empleoacute un tanque de 1 m3 las paralarvas se alimentaron con Artemia (1-4
mm) enriquecida con Chlorella sp antildeadiendo diariamente al tanque de cultivo una
mezcla de microalgas (Chlorella sp Isochrysis galbana y Chaetoceros sp) Como
complemento de Artemia se suministraron zoeas de centollo (Maja brachydactyla)
cuatro veces a la semana a una concentracioacuten de 001-01 zoeas ml-1 alcanzaacutendose
asiacute una supervivencia del 315 al diacutea 40 y un reducido nuacutemero de juveniles
bentoacutenicos dos semanas maacutes tarde (Iglesias et al 2002 2004) El seguimiento de los
uacutenicos pulpos supervivientes de este experimento hasta la fase adulta y la obtencioacuten
de una puesta de huevos por parte de una hembra originoacute el cierre del ciclo de vida
11
del pulpo en laboratorio por primer vez (Iglesias et al 2004) De forma similar
Carrasco et al (2003 2006) lograron obtener en uno de sus experimentos pulpos
bentoacutenicos a los 60 diacuteas con una supervivencia del 34
Sin embargo la utilizacioacuten de zoeas de crustaacuteceos como alimento vivo supone un
elevado riesgo ya que no se controla la obtencioacuten de las mismas en momentos
precisos ni la cantidad a producir ademaacutes de los elevados costes que conlleva esta
praacutectica por la necesidad de disponer de maacutes recursos materiales fiacutesicos y de
personal (Navarro y Villanueva 2000) Artemia en cambio es un crustaacuteceo faacutecil de
producir los nauplios o metanauplios se obtienen en 24-72 h y su composicioacuten
bioquiacutemica se puede modular mediante teacutecnicas de enriquecimiento al ser un filtrador
continuo obligado y no selectivo La utilizacioacuten de Artemia como presa viva se
consolida por tanto como una de las mejores alternativas para el cultivo de
paralarvas aunque sea necesario seguir trabajando en la modulacioacuten de su valor
nutricional de acuerdo con los posibles requerimientos de las paralarvas
Varios autores han utilizado nauplios de Artemia enriquecidos con distintos productos
comerciales (Navarro y Villanueva 2000 2003 Villanueva et al 2004 Okumura et al
2005) en sus experimentos de cultivo de paralarvas de pulpo siendo el tamantildeo de
estas presas (05-12 mm) adecuado hasta determinado momento del cultivo Sin
embargo se ha demostrado que las paralarvas tienen una preferencia clara por
Artemia de mayor tamantildeo (14 mm) sobre la maacutes pequentildea (08 mm) desde los
primeros diacuteas de vida (Iglesias et al 2006) De entre los productos o suplementos
nutricionales empleados por diversos autores para enriquecer Artemia se encuentran
emulsiones lipiacutedicas de la gama Selco (INVE) u otros aceites de pescado vitaminas
harina de huevos de pescado (BASF) harinas de cereales o productos de la gama Ori
(Skretting) (Navarro y Villanueva 2000 2003 Villanueva et al 2002 Iglesias et al
2004 Okumura et al 2005 Izquierdo et al 2008) pero ninguno ha originado tasas de
supervivencia o pesos secos de las paralarvas semejantes a los obtenidos con la
utilizacioacuten de zoeas de crustaacuteceos Como alternativa a las zoeas o a la Artemia
12
tambieacuten se han probado otros tipos de presas vivas o dietas artificiales para
suministrar a las paralarvas aunque en todo caso este tipo de pruebas se hicieron
complementadas con Artemia enriquecida (revisado por Iglesias et al 2007)
Otro de los principales suplementos empleados muy a menudo por diversos autores
para enriquecercultivar Artemia son las microalgas de las que las maacutes comuacutenmente
utilizadas fueron Nannochloropsis sp Dunaliella viridis Tetraselmis suecica
Isochrysis galbana Chaetoceros sp Chlorella sp (Hamazaki et al 1991 Navarro y
Villanueva 2000 Moxica et al 2002 Iglesias et al 2002 2004 Carrasco et al
2003 2006 Moxica et al 2006)
Hay un cierto consenso entre los grupos de investigadores que se dedican al cultivo
larvario del pulpo sobre que los aspectos nutricionales estaacuten en la base de la elevada
mortalidad observada durante su cultivo (Iglesias et al 2007) Uno de los problemas
que dificulta la interpretacioacuten de los resultados descritos y el avance en la nutricioacuten del
cultivo larvario del pulpo es que muy pocos autores presentan datos de la
composicioacuten bioquiacutemica de las dietas utilizadas y de sus efectos sobre la composicioacuten
de las paralarvas (Moxica et al 2002 Navarro et al 2000 2003 Villanueva et al
2004 Okumura et al 2005) cuando esta informacioacuten es crucial para interpretar los
resultados obtenidos y asiacute poder relacionarlos con la supervivencia y el crecimiento de
las paralarvas
A pesar de los continuos intentos para cerrar el ciclo de vida del pulpo de forma
rentable la mortalidad de las paralarvas durante la fase planctoacutenica sigue siendo casi
total Entre las estrategias propuestas para superar este importante problema estaacuten la
mejora de la composicioacuten de Artemia mediante las teacutecnicas de enriquecimiento
apropiadas y la formulacioacuten de microdietas inertes (Iglesias et al 2007) Importa
pues reducir el tiempo necesario hasta la obtencioacuten de los juveniles de Artemia del
tamantildeo deseado y optimizar su composicioacuten bioquiacutemica con base en los posibles
requerimientos nutricionales de las paralarvas El crecimiento y la composicioacuten
bioquiacutemica de Artemia sp pueden ser fuertemente modificados mediante la utilizacioacuten
13
de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

microalgas pero sin que se lograse alcanzar la fase de pre-asentamiento de las
paralarvas En los intentos llevados a cabo en el IEO de Vigo y maacutes tarde en el Centro
de Experimentacioacuten Pesquera de Asturias se obtuvieron algunos ejemplares de
juveniles bentoacutenicos que han llegado a alcanzar la edad adulta utilizaacutendose como
dieta para suministrar a las paralarvas juveniles de Artemia enriquecidos con
microalgas complementados con zoeas de centolla Maja brachydactyla en momentos
de disponibilidad de eacutestas (Iglesias et al 2002 2004 Carrasco et al 2003 2006) Las
primeras pruebas en las que se ha empleado esta mezcla de presas vivas (Moxica et
al 2002) fueron llevadas a cabo en el IEO de Vigo en un tanque de 9 m3 al que se
aportoacute inicialmente nauplios de Artemia y maacutes tarde juveniles de Artemia (1-4 mm) a
una densidad de presas de 01 Artemia ml-1 complementados con zoeas a partir de la
tercera semana de cultivo (3000 a 5000 zoeas al diacutea) Al tanque se aportaba a diario
una mezcla de microalgas (un 40 de Isochrysis galbana un 40 de Tetraselmis
suecica y un 20 de Chaetoceros sp) con el fin de mantener las presas vivas
constantemente enriquecidas Aunque no se han obtenido pulpos bentoacutenicos en este
experimento los autores refirieron la obtencioacuten de paralarvas en fase de asentamiento
con elevado nuacutemero de ventosas en los brazos (17-18) y un peso seco de 92 plusmn 09
mg a los 52 diacuteas de vida La supervivencia en este experimento fue de 83 al mes de
vida y de 02 al cabo de 52 diacuteas En un experimento posterior en el que se redujo la
escala y se empleoacute un tanque de 1 m3 las paralarvas se alimentaron con Artemia (1-4
mm) enriquecida con Chlorella sp antildeadiendo diariamente al tanque de cultivo una
mezcla de microalgas (Chlorella sp Isochrysis galbana y Chaetoceros sp) Como
complemento de Artemia se suministraron zoeas de centollo (Maja brachydactyla)
cuatro veces a la semana a una concentracioacuten de 001-01 zoeas ml-1 alcanzaacutendose
asiacute una supervivencia del 315 al diacutea 40 y un reducido nuacutemero de juveniles
bentoacutenicos dos semanas maacutes tarde (Iglesias et al 2002 2004) El seguimiento de los
uacutenicos pulpos supervivientes de este experimento hasta la fase adulta y la obtencioacuten
de una puesta de huevos por parte de una hembra originoacute el cierre del ciclo de vida
11
del pulpo en laboratorio por primer vez (Iglesias et al 2004) De forma similar
Carrasco et al (2003 2006) lograron obtener en uno de sus experimentos pulpos
bentoacutenicos a los 60 diacuteas con una supervivencia del 34
Sin embargo la utilizacioacuten de zoeas de crustaacuteceos como alimento vivo supone un
elevado riesgo ya que no se controla la obtencioacuten de las mismas en momentos
precisos ni la cantidad a producir ademaacutes de los elevados costes que conlleva esta
praacutectica por la necesidad de disponer de maacutes recursos materiales fiacutesicos y de
personal (Navarro y Villanueva 2000) Artemia en cambio es un crustaacuteceo faacutecil de
producir los nauplios o metanauplios se obtienen en 24-72 h y su composicioacuten
bioquiacutemica se puede modular mediante teacutecnicas de enriquecimiento al ser un filtrador
continuo obligado y no selectivo La utilizacioacuten de Artemia como presa viva se
consolida por tanto como una de las mejores alternativas para el cultivo de
paralarvas aunque sea necesario seguir trabajando en la modulacioacuten de su valor
nutricional de acuerdo con los posibles requerimientos de las paralarvas
Varios autores han utilizado nauplios de Artemia enriquecidos con distintos productos
comerciales (Navarro y Villanueva 2000 2003 Villanueva et al 2004 Okumura et al
2005) en sus experimentos de cultivo de paralarvas de pulpo siendo el tamantildeo de
estas presas (05-12 mm) adecuado hasta determinado momento del cultivo Sin
embargo se ha demostrado que las paralarvas tienen una preferencia clara por
Artemia de mayor tamantildeo (14 mm) sobre la maacutes pequentildea (08 mm) desde los
primeros diacuteas de vida (Iglesias et al 2006) De entre los productos o suplementos
nutricionales empleados por diversos autores para enriquecer Artemia se encuentran
emulsiones lipiacutedicas de la gama Selco (INVE) u otros aceites de pescado vitaminas
harina de huevos de pescado (BASF) harinas de cereales o productos de la gama Ori
(Skretting) (Navarro y Villanueva 2000 2003 Villanueva et al 2002 Iglesias et al
2004 Okumura et al 2005 Izquierdo et al 2008) pero ninguno ha originado tasas de
supervivencia o pesos secos de las paralarvas semejantes a los obtenidos con la
utilizacioacuten de zoeas de crustaacuteceos Como alternativa a las zoeas o a la Artemia
12
tambieacuten se han probado otros tipos de presas vivas o dietas artificiales para
suministrar a las paralarvas aunque en todo caso este tipo de pruebas se hicieron
complementadas con Artemia enriquecida (revisado por Iglesias et al 2007)
Otro de los principales suplementos empleados muy a menudo por diversos autores
para enriquecercultivar Artemia son las microalgas de las que las maacutes comuacutenmente
utilizadas fueron Nannochloropsis sp Dunaliella viridis Tetraselmis suecica
Isochrysis galbana Chaetoceros sp Chlorella sp (Hamazaki et al 1991 Navarro y
Villanueva 2000 Moxica et al 2002 Iglesias et al 2002 2004 Carrasco et al
2003 2006 Moxica et al 2006)
Hay un cierto consenso entre los grupos de investigadores que se dedican al cultivo
larvario del pulpo sobre que los aspectos nutricionales estaacuten en la base de la elevada
mortalidad observada durante su cultivo (Iglesias et al 2007) Uno de los problemas
que dificulta la interpretacioacuten de los resultados descritos y el avance en la nutricioacuten del
cultivo larvario del pulpo es que muy pocos autores presentan datos de la
composicioacuten bioquiacutemica de las dietas utilizadas y de sus efectos sobre la composicioacuten
de las paralarvas (Moxica et al 2002 Navarro et al 2000 2003 Villanueva et al
2004 Okumura et al 2005) cuando esta informacioacuten es crucial para interpretar los
resultados obtenidos y asiacute poder relacionarlos con la supervivencia y el crecimiento de
las paralarvas
A pesar de los continuos intentos para cerrar el ciclo de vida del pulpo de forma
rentable la mortalidad de las paralarvas durante la fase planctoacutenica sigue siendo casi
total Entre las estrategias propuestas para superar este importante problema estaacuten la
mejora de la composicioacuten de Artemia mediante las teacutecnicas de enriquecimiento
apropiadas y la formulacioacuten de microdietas inertes (Iglesias et al 2007) Importa
pues reducir el tiempo necesario hasta la obtencioacuten de los juveniles de Artemia del
tamantildeo deseado y optimizar su composicioacuten bioquiacutemica con base en los posibles
requerimientos nutricionales de las paralarvas El crecimiento y la composicioacuten
bioquiacutemica de Artemia sp pueden ser fuertemente modificados mediante la utilizacioacuten
13
de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

del pulpo en laboratorio por primer vez (Iglesias et al 2004) De forma similar
Carrasco et al (2003 2006) lograron obtener en uno de sus experimentos pulpos
bentoacutenicos a los 60 diacuteas con una supervivencia del 34
Sin embargo la utilizacioacuten de zoeas de crustaacuteceos como alimento vivo supone un
elevado riesgo ya que no se controla la obtencioacuten de las mismas en momentos
precisos ni la cantidad a producir ademaacutes de los elevados costes que conlleva esta
praacutectica por la necesidad de disponer de maacutes recursos materiales fiacutesicos y de
personal (Navarro y Villanueva 2000) Artemia en cambio es un crustaacuteceo faacutecil de
producir los nauplios o metanauplios se obtienen en 24-72 h y su composicioacuten
bioquiacutemica se puede modular mediante teacutecnicas de enriquecimiento al ser un filtrador
continuo obligado y no selectivo La utilizacioacuten de Artemia como presa viva se
consolida por tanto como una de las mejores alternativas para el cultivo de
paralarvas aunque sea necesario seguir trabajando en la modulacioacuten de su valor
nutricional de acuerdo con los posibles requerimientos de las paralarvas
Varios autores han utilizado nauplios de Artemia enriquecidos con distintos productos
comerciales (Navarro y Villanueva 2000 2003 Villanueva et al 2004 Okumura et al
2005) en sus experimentos de cultivo de paralarvas de pulpo siendo el tamantildeo de
estas presas (05-12 mm) adecuado hasta determinado momento del cultivo Sin
embargo se ha demostrado que las paralarvas tienen una preferencia clara por
Artemia de mayor tamantildeo (14 mm) sobre la maacutes pequentildea (08 mm) desde los
primeros diacuteas de vida (Iglesias et al 2006) De entre los productos o suplementos
nutricionales empleados por diversos autores para enriquecer Artemia se encuentran
emulsiones lipiacutedicas de la gama Selco (INVE) u otros aceites de pescado vitaminas
harina de huevos de pescado (BASF) harinas de cereales o productos de la gama Ori
(Skretting) (Navarro y Villanueva 2000 2003 Villanueva et al 2002 Iglesias et al
2004 Okumura et al 2005 Izquierdo et al 2008) pero ninguno ha originado tasas de
supervivencia o pesos secos de las paralarvas semejantes a los obtenidos con la
utilizacioacuten de zoeas de crustaacuteceos Como alternativa a las zoeas o a la Artemia
12
tambieacuten se han probado otros tipos de presas vivas o dietas artificiales para
suministrar a las paralarvas aunque en todo caso este tipo de pruebas se hicieron
complementadas con Artemia enriquecida (revisado por Iglesias et al 2007)
Otro de los principales suplementos empleados muy a menudo por diversos autores
para enriquecercultivar Artemia son las microalgas de las que las maacutes comuacutenmente
utilizadas fueron Nannochloropsis sp Dunaliella viridis Tetraselmis suecica
Isochrysis galbana Chaetoceros sp Chlorella sp (Hamazaki et al 1991 Navarro y
Villanueva 2000 Moxica et al 2002 Iglesias et al 2002 2004 Carrasco et al
2003 2006 Moxica et al 2006)
Hay un cierto consenso entre los grupos de investigadores que se dedican al cultivo
larvario del pulpo sobre que los aspectos nutricionales estaacuten en la base de la elevada
mortalidad observada durante su cultivo (Iglesias et al 2007) Uno de los problemas
que dificulta la interpretacioacuten de los resultados descritos y el avance en la nutricioacuten del
cultivo larvario del pulpo es que muy pocos autores presentan datos de la
composicioacuten bioquiacutemica de las dietas utilizadas y de sus efectos sobre la composicioacuten
de las paralarvas (Moxica et al 2002 Navarro et al 2000 2003 Villanueva et al
2004 Okumura et al 2005) cuando esta informacioacuten es crucial para interpretar los
resultados obtenidos y asiacute poder relacionarlos con la supervivencia y el crecimiento de
las paralarvas
A pesar de los continuos intentos para cerrar el ciclo de vida del pulpo de forma
rentable la mortalidad de las paralarvas durante la fase planctoacutenica sigue siendo casi
total Entre las estrategias propuestas para superar este importante problema estaacuten la
mejora de la composicioacuten de Artemia mediante las teacutecnicas de enriquecimiento
apropiadas y la formulacioacuten de microdietas inertes (Iglesias et al 2007) Importa
pues reducir el tiempo necesario hasta la obtencioacuten de los juveniles de Artemia del
tamantildeo deseado y optimizar su composicioacuten bioquiacutemica con base en los posibles
requerimientos nutricionales de las paralarvas El crecimiento y la composicioacuten
bioquiacutemica de Artemia sp pueden ser fuertemente modificados mediante la utilizacioacuten
13
de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

tambieacuten se han probado otros tipos de presas vivas o dietas artificiales para
suministrar a las paralarvas aunque en todo caso este tipo de pruebas se hicieron
complementadas con Artemia enriquecida (revisado por Iglesias et al 2007)
Otro de los principales suplementos empleados muy a menudo por diversos autores
para enriquecercultivar Artemia son las microalgas de las que las maacutes comuacutenmente
utilizadas fueron Nannochloropsis sp Dunaliella viridis Tetraselmis suecica
Isochrysis galbana Chaetoceros sp Chlorella sp (Hamazaki et al 1991 Navarro y
Villanueva 2000 Moxica et al 2002 Iglesias et al 2002 2004 Carrasco et al
2003 2006 Moxica et al 2006)
Hay un cierto consenso entre los grupos de investigadores que se dedican al cultivo
larvario del pulpo sobre que los aspectos nutricionales estaacuten en la base de la elevada
mortalidad observada durante su cultivo (Iglesias et al 2007) Uno de los problemas
que dificulta la interpretacioacuten de los resultados descritos y el avance en la nutricioacuten del
cultivo larvario del pulpo es que muy pocos autores presentan datos de la
composicioacuten bioquiacutemica de las dietas utilizadas y de sus efectos sobre la composicioacuten
de las paralarvas (Moxica et al 2002 Navarro et al 2000 2003 Villanueva et al
2004 Okumura et al 2005) cuando esta informacioacuten es crucial para interpretar los
resultados obtenidos y asiacute poder relacionarlos con la supervivencia y el crecimiento de
las paralarvas
A pesar de los continuos intentos para cerrar el ciclo de vida del pulpo de forma
rentable la mortalidad de las paralarvas durante la fase planctoacutenica sigue siendo casi
total Entre las estrategias propuestas para superar este importante problema estaacuten la
mejora de la composicioacuten de Artemia mediante las teacutecnicas de enriquecimiento
apropiadas y la formulacioacuten de microdietas inertes (Iglesias et al 2007) Importa
pues reducir el tiempo necesario hasta la obtencioacuten de los juveniles de Artemia del
tamantildeo deseado y optimizar su composicioacuten bioquiacutemica con base en los posibles
requerimientos nutricionales de las paralarvas El crecimiento y la composicioacuten
bioquiacutemica de Artemia sp pueden ser fuertemente modificados mediante la utilizacioacuten
13
de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

de microalgas de composicioacuten bioquiacutemica mejorada producidas en cultivos
semicontinuos (Faacutebregas et al 1996b Faacutebregas et al 2001)
En el caso de las paralarvas de pulpo se ha verificado que un tamantildeo de Artemia de
15 a 40 mm es el maacutes adecuado para suministrar a lo largo de su cultivo (Iglesias et
al 2004 2006 2007 Carrasco et al 2006) Los tamantildeos maacutes pequentildeos se utilizariacutean
en los primeros diacuteas de vida incrementando el tamantildeo de las presas a medida que las
paralarvas se hacen maacutes grandes Aunque otros autores hayan observado que
tamantildeos inferiores a 15 mm son aptos para suministrar a las paralarvas en sus
primeros diacuteas de vida (Navarro y Villanueva 2000 2003 Villanueva et al 2002
2004 Okumura et al 2005) ese tamantildeo se vuelve poco atractivo a partir de los 20
diacuteas de cultivo La optimizacioacuten del crecimiento de nauplios de Artemia hasta los
tamantildeos maacutes adecuados y la mejora del perfil nutricional de los juveniles de Artemia
son importantes metas a alcanzar en el cultivo larvario del pulpo con el fin de facilitar
la gestioacuten y disponibilidad del alimento vivo durante los experimentos y mejorar las
tasas de crecimiento y supervivencia de las paralarvas
En el presente trabajo se hizo en primer lugar un estudio geneacutetico de la progenie de
hembras de pulpo intentando confirmar la hipoacutetesis de la muacuteltiple paternidad en esta
especie dados los dos patrones de comportamiento reproductivo que son conflictivos
en este aspecto acoplamiento con muacuteltiples individuos y papel del hectocoacutetilo para
eliminar los anteriores espermatoacuteforos Esta informacioacuten sobre los aspectos
reproductivos es de gran importancia para una gestioacuten racional de los stocks
seleccioacuten de estrategias de repoblacioacuten y debe ser considerado en los estudios de
geneacutetica de poblaciones y de conservacioacuten asiacute como en el disentildeo y gestioacuten de la
acuicultura de O vulgaris Permitiriacutea ademaacutes desvelar algunas incoacutegnitas sobre
aspectos de la biologiacutea reproductiva del pulpo estrategias de transferencia de pools
geneacuteticos dinaacutemica de poblaciones etceacutetera
14
En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

En segundo lugar se enfocoacute la optimizacioacuten de la composicioacuten nutricional de Artemia
como principal presa viva de las paralarvas teniendo en cuenta los posibles
requerimientos nutricionales de eacutestas evaluando los efectos de diferentes dietas sobre
el crecimiento y la supervivencia de paralarvas cultivadas en tanques a pequentildea
escala Para la mejora de la composicioacuten nutricional de Artemia se usaron microalgas
cultivadas en condiciones optimizadas y controladas empleando ademaacutes suplementos
purificados como aminoaacutecidos esenciales libres
2 Material y meacutetodos
21 Estudio geneacutetico de la progenie
Un total de 124 individuos de O vulgaris salvajes adultos fueron capturados y
mantenidos en jaulas suspendidas de una batea en la riacutea de Vigo (Galicia Espantildea)
Las muestras se tomaron de puestas obtenidas en estas condiciones de cultivo
conteniendo cada una cuatro racimos con 10 a 15 huevos asiacute como del tejido
muscular de las hembras correspondientes Un total de 41 muestras de tejido
muscular fueron seleccionadas para el genotipado de la poblacioacuten Los huevos de
cada puesta fueron fijados en etanol del 90 en tubos separados Once embriones
fueron diseccionados y situados por separado en 15 ml de etanol
El ADN total fue aislado de cada embrioacuten asiacute como del tejido muscular de las hembras
y de otros especiacutemenes usando el sistema NucPrepreg en un ABI PRISMTM 6100
Nucleic Acid PrepStation (Applied Biosystems) Los embriones las hembras y las
muestras de la poblacioacuten fueron genotipados en los loci de los microsateacutelites Oct3
(repeticioacuten dinucleotiacutedica) y Ov12 (repeticioacuten tetranucleotiacutedica) usando los cebadores
microOct3FmicroOct3R y microOct12FmicroOct12R (Greatorex et al 2000) marcados con los
15
fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

fluocrocromos 6-FAM y TET respectivamente Las condiciones de la PCR realizada
en un termociclador GeneAmp 9700 (Applied Biosystem) incluyoacute un paso inicial de
desnaturalizacioacuten a 96ordmC durante 5 minutos seguido por 35 ciclos de
desnaturalizacioacuten a 96ordmC durante 25 segundos annealing a 54ordmC durante 25
segundos y extensioacuten a 72ordmC durante 25 segundos Los productos de PCR
resultantes junto con un marcador de tamantildeo GeneScan-350 ROX (Applied
Biosystem) fueron separados y detectados en un secuenciador automaacutetico de ADN
ABI Prism 377 (Applied Biosystems) La deteccioacuten de alelos y la estimacioacuten de
tamantildeos se llevoacute a cabo con el software GenScan y Genotyper (Applied Biosystems)
Las frecuencias aleacutelicas y la heterozigosis en las madres sus descendientes
muestreados y la muestra de la poblacioacuten fueron estimados con CERVUS v30
(Kalinowski et al 2007) El test de equilibrio de Hardy-Weinberg (HWE) fue realizado
solamente en la muestra de la poblacioacuten para asumir la independencia del muestreo
Los alelos maternos y paternos en ambos loci fueron deducidos usando el criterio de
exclusioacuten (Adam y Ardren 2008) comparando el genotipo materno con el genotipo de
cada descendiente
La paternidad muacuteltiple fue evidenciada cuando el nuacutemero de alelos paternos en un
uacutenico racimo simple fue superior a dos Como alelos paternos se incluyen aquellos
que no son alelos maternos alelos homocigoacuteticos o aquellos alelos heterozigoacuteticos
que son ideacutenticos a los de la madre Las probabilidades previstas de exclusioacuten el
nuacutemero miacutenimo de padres posibles y sus genotipos deducidos maacutes probables fueron
obtenidos usando GERUD v20 (Jones 2005) La probabilidad de los genotipos
paternos fue obtenida con patrones de segregacioacuten y frecuencias genotiacutepicas en la
poblacioacuten Las relaciones entre pares de huevos del mismo racimo fueron estimadas
por medio del parentesco entre pares (r) (Queller y Goodnight 1989) con Kingroup
(Konovalov et al 2004) seguacuten lo dispuesto en el programa KINSHIP (Goodnight y
Queller 1999) Asiacute la muacuteltiple paternidad produce medio-hermanos maternos con un
16
valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

valor previsto de r=025 mientras que el valor para los hermanos completos es de
alrededor de r=05
22 Sistemas de cultivo de las microalgas
Las microalgas se cultivaron en reacutegimen semicontinuo con agua de mar (35 ppt)
autoclavada (121ordmC durante 15 min) enriquecida con medio de cultivo ALGAL-1 (40
mM NaNO3) y con una tasa de renovacioacuten diaria de un 30 del volumen total
(Faacutebregas et al 2001) Los cultivos se realizaron con un ciclo de luzoscuridad de
12h12h con flujo de aire constante y el pH se mantuvo entre 75-80 mediante la
inyeccioacuten automaacutetica de CO2 Los sistemas de cultivo fueron botellas de 1L para los
ensayos de enriquecimiento de Artemia con diferentes microalgas (Fig 5) o
botellones de cristal de 6 L para el cultivo de las cepas seleccionadas para la
produccioacuten y enriquecimiento de Artemia como alimento vivo de las paralarvas de
pulpo
Figura 5 - Cultivo de diferentes cepas de microalgas marinas en botellas de 1 L para los
ensayos de enriquecimiento de juveniles de Artemia De izquierda a derecha Tetraselmis
suecica Isochrysis galbana Isochrysis T-ISO y Rhodomonas lens
17
Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

Los caacutelculos de la densidad celular se hicieron mediante el recuento de aliacutecuotas al
microscopio de contraste de fase (Nikon Labophot) en caacutemaras Improved Neubaeur
(Brand)
23 Ensayos de enriquecimiento de juveniles de Artemia
Los nauplios de Artemia sp (AF INVE Beacutelgica) inicialmente con un tamantildeo de 05
mm se alimentaron con Tetraselmis suecica durante 3 y 4 diacuteas hasta alcanzar
tamantildeos aproximados de 14 mm y 25 mm respectivamente El cultivo se hizo en
vasos de 1 L de volumen total con 700 ml de agua a un temperatura constante de
-1265plusmn05 ordmC y densidad inicial de 5 nauplios ml Para el enriquecimiento final de 24 h
con las diferentes microalgas se formaron grupos por triplicado a los que se
suministroacute igual cantidad (en peso seco) de microalgas por Artemia Los grupos
comparados fueron ATET Artemia enriquecida con Tetraselmis suecica AISO
enriquecida con Isochrysis galbana AT-ISO enriquecida con Isochrysis aff galbana
clon T-ISO y ARHO enriquecida con Rhodomonas lens Se muestrearon grupos de
juveniles de Artemia para los anaacutelisis de composicioacuten bioquiacutemica congelaacutendose eacutestas
a -20ordmC
24 Ensayo de cultivo de paralarvas de pulpo
Para el cultivo de paralarvas de pulpo se utilizaron nueve tanques tronco-coacutenicos de
color blanco con 50 L de volumen de agua Se utilizoacute un circuito de agua semi-cerrado
cuyo volumen total era de 600 L y se establecioacute un caudal de 11 L h-1 El porcentaje de
purga diario ha sido de un 10 La temperatura se mantuvo constante a 195plusmn05 ordmC
mediante la colocacioacuten de termorreguladores en el tanque reservorio Se establecioacute un
fotoperiacuteodo de 18h luz6h oscuridad y las densidad inicial de paralarvas fue de 10
18
paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

paralarvas L-1 Para producir el alimento vivo (juveniles de Artemia) se cultivaron
microalgas marinas en botellones de 6 L que serviacutean tanto para el crecimiento de
Artemia como para su enriquecimiento final Se ha empleado la microalga
Rhodomonas lens para el crecimiento de los nauplios de Artemia hasta la fase de
juvenil (hasta 15 mm) ya que eacutesta ha dado muy buenos resultados de crecimiento de
Artemia en comparacioacuten con otras cepas (resultados no descritos en este trabajo) La
produccioacuten de Artemia se ha hecho en pequentildeos tanques de 12 L de volumen total En
el ensayo de cultivo de paralarvas se han probado tres dietas a base de juveniles de
Artemia (16-28 mm) suministrados en tres tomas a lo largo del diacutea (12h00 16h00 y
20h00) La densidad de presas en los tanques era de 005 ind ml-1 diacutea-1 Los grupos de
paralarvas comparados fueron grupo control (ANANO) - Artemia enriquecida con la
microalga Nannochloropsis gaditana grupo AR+I - Artemia enriquecida con una
mezcla de las microalgas Rhodomonas lens e Isochrysis galbana (7030 en porcentaje
de peso seco) y AR+I+AA - la misma mezcla de microalgas del grupo anterior
suplementada con AA libres disueltos en el agua en las siguientes concentraciones
34 mM lisina 25 mM arginina y 15 mM metionina Antes de ser suministradas a las
paralarvas las artemias se enriquecieron durante 24 h con las microalgas
correspondientes continuaacutendose el proceso de enriquecimiento a lo largo del diacutea de
su utilizacioacuten hasta la distribucioacuten de las tomas del alimento Al grupo de artemias que
recibioacute los AA libres disueltos en el agua se antildeadiacutean los AA por la mantildeana
recogieacutendose las artemias del tanque de enriquecimiento en las horas de alimentacioacuten
por lo que el tiempo total de enriquecimiento variaba de 3 a 9 h
Se determinoacute el peso seco de las paralarvas a los diacuteas 0 15 y 25 mediante el
muestreo de 10 individuos por tanque La supervivencia hasta el diacutea 15 se calculoacute a
traveacutes del recuento de las larvas muertas mientras que a partir del diacutea 20 se optoacute por
contar los individuos vivos presentes en los tanques
19
25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

25 Anaacutelisis de composicioacuten bioquiacutemica
El contenido proteico de los juveniles de Artemia se determinoacute por el meacutetodo de Lowry
(Lowry et al 1951) sometieacutendose las muestras a una hidroacutelisis alcalina durante 1h a
95-100ordmC mientras que en los ensayos de paralarvas la determinacioacuten de la proteiacutena
total se derivoacute del contenido en nitroacutegeno de las muestras multiplicado por el factor
625 (Nx625) Para el anaacutelisis elemental de C-N-H se utilizoacute un auto-analizador Carlo
Erba EA 1108 equipado con un auto-muestreo AS200 Los liacutepidos totales de las
muestras de Artemia se extrajeron con una solucioacuten 12 (vv) de cloroformo-metanol
(Bligh y Dyer 1959) y se calcularon por diferencia de peso despueacutes de secar los
extractos lipiacutedicos en una estufa a 37ordmC Los carbohidratos se cuantificaron por el
meacutetodo fenol-sulfuacuterico (Dubois et al 1956) Para la determinacioacuten de los aacutecidos
grasos los extractos de liacutepidos totales se sometieron a metanoliacutesis (5 aacutecido
clorhiacutedrico en metanol) durante 25 h a 85 ordmC (Sato y Murata 1988) seguido de
recuperacioacuten de los eacutesteres de aacutecidos grasos con hexano Eacutestos se identificaron y
cuantificaron por cromatografiacutea de gases-espectrometriacutea de masas (GC-MS) usando
un equipo Perkin-Elmer 800-8000 Series y una columna OmegawaxTM 250 fused silica
(Supelco) de 30 m y 025 mm de diaacutemetro Para la cuantificacioacuten de los aminoaacutecidos
(AA) totales de los juveniles de Artemia se prepararon muestras para determinar la
fraccioacuten de AA libres (AAL) y de AA proteicos (AAP) Para ello se sometieron
muestras liofilizadas (10-20 mg) a aacutecido tricloroaceacutetico (6) despueacutes tratadas con HCl
6 M a 105 ordmC durante 24 h La solucioacuten obtenida se evaporoacute en un rotavapor siendo
posteriormente resuspendida en una solucioacuten de HCl 20 mM Tanto los AAL como los
AAP fueron analizados por cromatografiacutea de fase-reversa (Gilson HPLC) conectada a
un equipo de muestreo automaacutetico ASTED y con deteccioacuten por fluorescencia
(reactivos OPA y FMOC) El aacutecido L-α-amino-n-butiacuterico (a-ABA) se usoacute como estaacutendar
interno Los picos resultantes se analizaron a traveacutes del software Gilson Unipoint 715
20
El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

El AAP triptoacutefano no se pudo cuantificar ya que se destruye por accioacuten de la hidrolisis
aacutecida Todos los anaacutelisis se realizaron por triplicado
26 Anaacutelisis estadiacutesticos
Los anaacutelisis estadiacutesticos se hicieron con el software SPSS 150 (SPSS Inc) Los
datos de tamantildeo de las artemias y de porcentajes de composicioacuten bioquiacutemica se
compararon por anaacutelisis de varianza (ANOVA) seguido de test post-hoc de Tukey-
Kramer con un nivel de significancia del 005 Antes de efectuar estas comparaciones
los valores de porcentajes fueron transformados al arco-seno de su raiacutez cuadrada con
el fin de obtener una distribucioacuten normal de los valores (Zar 1999)
21
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
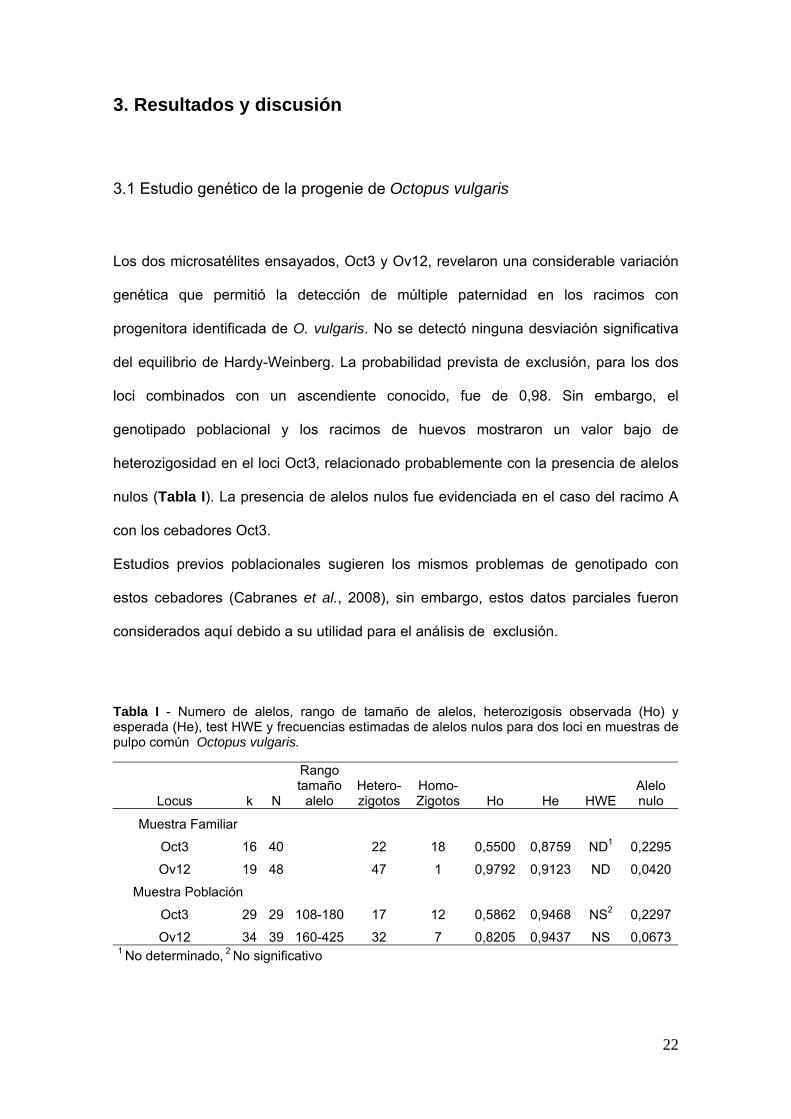
3 Resultados y discusioacuten
31 Estudio geneacutetico de la progenie de Octopus vulgaris
Los dos microsateacutelites ensayados Oct3 y Ov12 revelaron una considerable variacioacuten
geneacutetica que permitioacute la deteccioacuten de muacuteltiple paternidad en los racimos con
progenitora identificada de O vulgaris No se detectoacute ninguna desviacioacuten significativa
del equilibrio de Hardy-Weinberg La probabilidad prevista de exclusioacuten para los dos
loci combinados con un ascendiente conocido fue de 098 Sin embargo el
genotipado poblacional y los racimos de huevos mostraron un valor bajo de
heterozigosidad en el loci Oct3 relacionado probablemente con la presencia de alelos
nulos (Tabla I) La presencia de alelos nulos fue evidenciada en el caso del racimo A
con los cebadores Oct3
Estudios previos poblacionales sugieren los mismos problemas de genotipado con
estos cebadores (Cabranes et al 2008) sin embargo estos datos parciales fueron
considerados aquiacute debido a su utilidad para el anaacutelisis de exclusioacuten
Tabla I - Numero de alelos rango de tamantildeo de alelos heterozigosis observada (Ho) y esperada (He) test HWE y frecuencias estimadas de alelos nulos para dos loci en muestras de pulpo comuacuten Octopus vulgaris
Locus k N
Rango tamantildeo
alelo Hetero- Homo- zigotos Zigotos Ho He HWE
Alelo nulo
Muestra Familiar
Oct3 16 40 22 18 05500 08759 ND1 02295
Ov12 19 48 47 1 09792 09123 ND 00420
Muestra Poblacioacuten
Oct3 29 29 108-180 17 12 05862 09468 NS2 02297
Ov12 34 39 160-425 32 7 08205 09437 NS 006731 No determinado 2 No significativo
22
Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

Cuatro hembras (A-D) y sus muestras de huevos respectivos (N=11) fueron
genotipados para los dos loci Dentro del racimo A la hembra mostroacute un solo alelo
(165) pero el estado de homozigosis puede ser desechado debido a la presencia de
un alelo simple diferente en uno de los embriones (no5) Por lo tanto Oct3 muestra
cinco diferentes alelos un alelo nulo materno y quizaacutes uno o maacutes alelos nulos
paternos En este racimo A el genotipado con Ov12 dio lugar a dos alelos maternos y
a 7 alelos paternos En el racimo B Oct3 muestra cinco alelos tres de los cuales son
diferentes a los maternos Cinco diferentes alelos fueron detectados para el marcador
Ov12 El racimo C mostroacute tres alelos adicionales para Oct3 y cuatro para el Ov12
ademaacutes de los dos alelos detectados en la hembra Finalmente el racimo D mostroacute
cinco y seis diferentes alelos para los marcadores Oct3 y Ov12 respectivamente
Los valores estimados de parentesco entre pares (r) por comparaciones dentro de
cada racimo fueron dispersos con un valor medio le 04 (Fig 6) sugiriendo la
presencia de medio-hermanos en cada progenie Los valores medios maacutes bajos de r
fueron obtenidos para el racimo 1 (r=023) 3 y 4 (r=030) Para los descendientes el
nuacutemero miacutenimo de padres es de 3 a 4 y su genotipo fue deducido del valor estimado
de verosimilitud (Tabla II) La progenie engendrada por cada macho va de 2 a 7 Por lo
tanto maacutes de dos alelos paternos fueron encontrados en todas las masas de huevos
analizadas para ambos loci microsateacutelites apoyando asiacute la hipoacutetesis de la muacuteltiple
paternidad en el pulpo del Atlaacutentico O vulgaris Este comportamiento reproductivo
parece ser una regla general porque maacutes de dos contribuciones paternas fueron
detectadas en todos los descendientes analizados
23
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
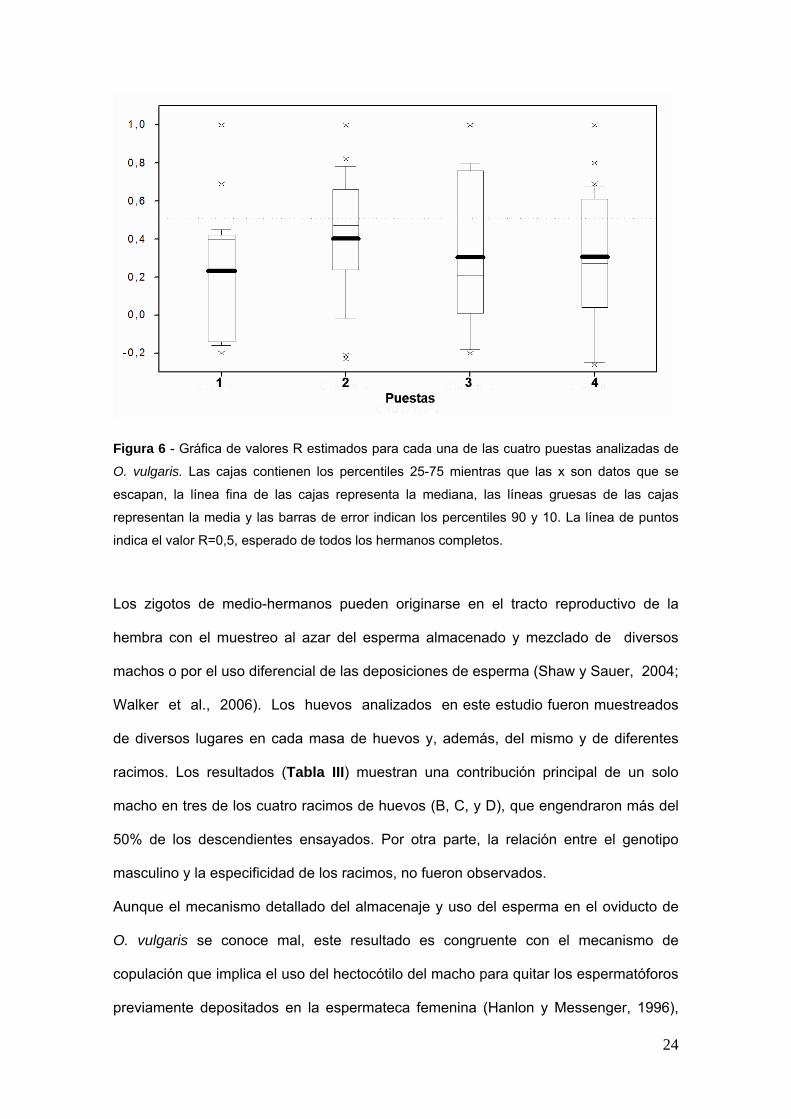
Figura 6 - Graacutefica de valores R estimados para cada una de las cuatro puestas analizadas de
O vulgaris Las cajas contienen los percentiles 25-75 mientras que las x son datos que se
escapan la liacutenea fina de las cajas representa la mediana las liacuteneas gruesas de las cajas
representan la media y las barras de error indican los percentiles 90 y 10 La liacutenea de puntos
indica el valor R=05 esperado de todos los hermanos completos
Los zigotos de medio-hermanos pueden originarse en el tracto reproductivo de la
hembra con el muestreo al azar del esperma almacenado y mezclado de diversos
machos o por el uso diferencial de las deposiciones de esperma (Shaw y Sauer 2004
Walker et al 2006) Los huevos analizados en este estudio fueron muestreados
de diversos lugares en cada masa de huevos y ademaacutes del mismo y de diferentes
racimos Los resultados (Tabla III) muestran una contribucioacuten principal de un solo
macho en tres de los cuatro racimos de huevos (B C y D) que engendraron maacutes del
50 de los descendientes ensayados Por otra parte la relacioacuten entre el genotipo
masculino y la especificidad de los racimos no fueron observados
Aunque el mecanismo detallado del almacenaje y uso del esperma en el oviducto de
O vulgaris se conoce mal este resultado es congruente con el mecanismo de
copulacioacuten que implica el uso del hectocoacutetilo del macho para quitar los espermatoacuteforos
previamente depositados en la espermateca femenina (Hanlon y Messenger 1996)
24
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
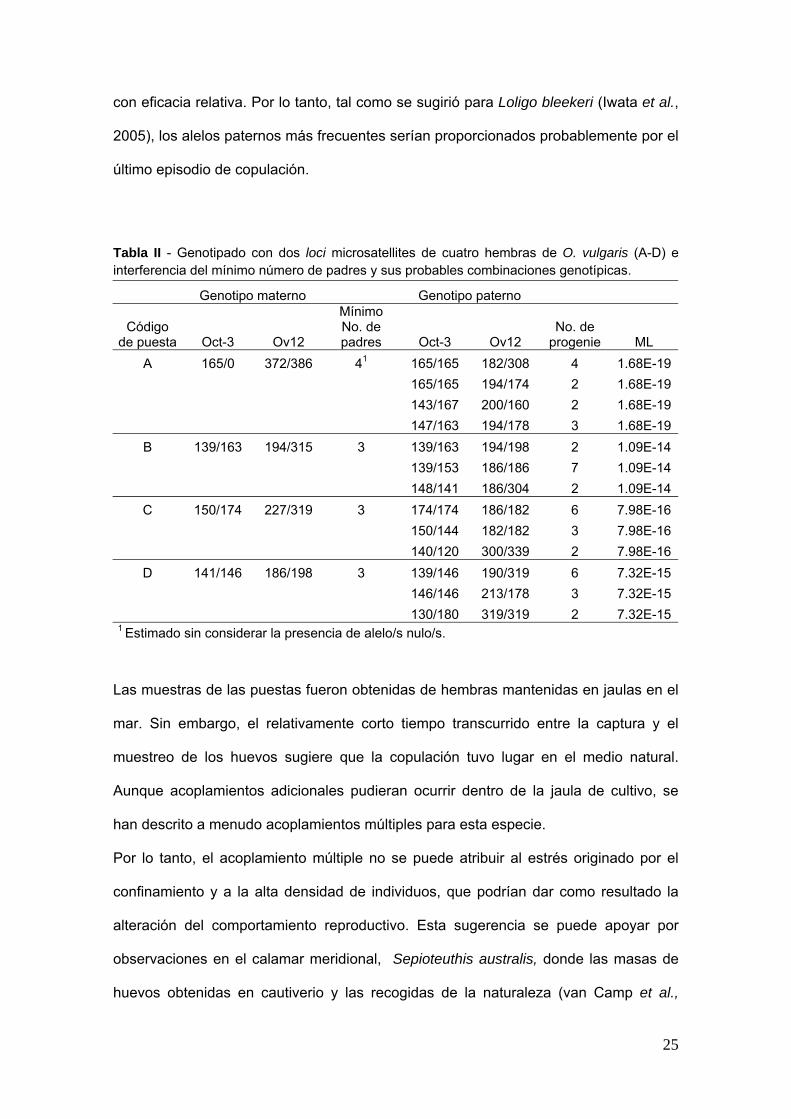
con eficacia relativa Por lo tanto tal como se sugirioacute para Loligo bleekeri (Iwata et al
2005) los alelos paternos maacutes frecuentes seriacutean proporcionados probablemente por el
uacuteltimo episodio de copulacioacuten
Tabla II - Genotipado con dos loci microsatellites de cuatro hembras de O vulgaris (A-D) e interferencia del miacutenimo nuacutemero de padres y sus probables combinaciones genotiacutepicas
Genotipo materno Genotipo paterno Miacutenimo No de padres
Coacutedigo de puesta Oct-3 Ov12 Oct-3 Ov12
No de progenie ML
1A 1650 372386 4 165165 182308 4 168E-19 165165 194174 2 168E-19 143167 200160 2 168E-19 147163 194178 3 168E-19
B 139163 194315 3 139163 194198 2 109E-14 139153 186186 7 109E-14 148141 186304 2 109E-14
C 150174 227319 3 174174 186182 6 798E-16 150144 182182 3 798E-16 140120 300339 2 798E-16
D 141146 186198 3 139146 190319 6 732E-15 146146 213178 3 732E-15 130180 319319 2 732E-15
1 Estimado sin considerar la presencia de alelos nulos
Las muestras de las puestas fueron obtenidas de hembras mantenidas en jaulas en el
mar Sin embargo el relativamente corto tiempo transcurrido entre la captura y el
muestreo de los huevos sugiere que la copulacioacuten tuvo lugar en el medio natural
Aunque acoplamientos adicionales pudieran ocurrir dentro de la jaula de cultivo se
han descrito a menudo acoplamientos muacuteltiples para esta especie
Por lo tanto el acoplamiento muacuteltiple no se puede atribuir al estreacutes originado por el
confinamiento y a la alta densidad de individuos que podriacutean dar como resultado la
alteracioacuten del comportamiento reproductivo Esta sugerencia se puede apoyar por
observaciones en el calamar meridional Sepioteuthis australis donde las masas de
huevos obtenidas en cautiverio y las recogidas de la naturaleza (van Camp et al
25
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
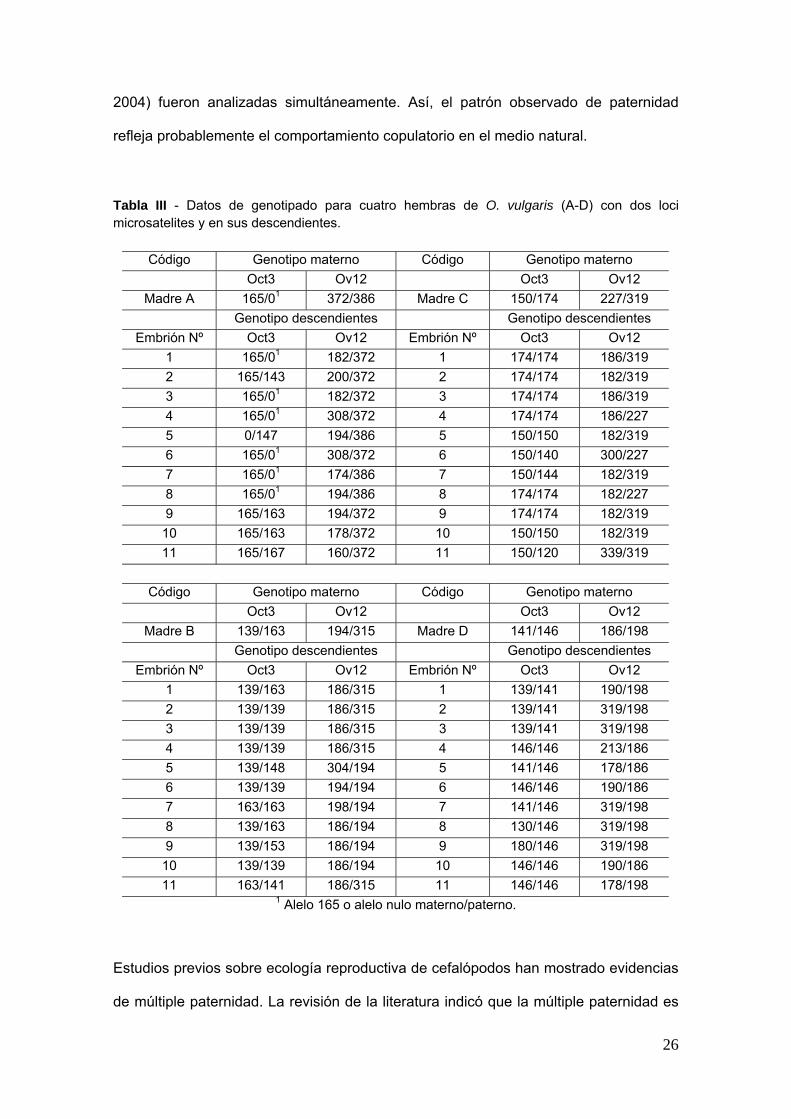
2004) fueron analizadas simultaacuteneamente Asiacute el patroacuten observado de paternidad
refleja probablemente el comportamiento copulatorio en el medio natural
Tabla III - Datos de genotipado para cuatro hembras de O vulgaris (A-D) con dos loci microsatelites y en sus descendientes
Coacutedigo Genotipo materno Coacutedigo Genotipo materno Oct3 Ov12 Oct3 Ov12
Madre A 16501 372386 Madre C 150174 227319 Genotipo descendientes Genotipo descendientes
Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12 1 16501 182372 1 174174 186319 2 165143 200372 2 174174 182319 3 16501 182372 3 174174 186319 4 16501 308372 4 174174 186227 5 0147 194386 5 150150 182319 6 16501 308372 6 150140 300227 7 16501 174386 7 150144 182319 8 16501 194386 8 174174 182227 9 165163 194372 9 174174 182319
10 165163 178372 10 150150 182319 11 165167 160372 11 150120 339319
Coacutedigo Genotipo materno Coacutedigo Genotipo materno
Oct3 Ov12 Oct3 Ov12 Madre B 139163 194315 Madre D 141146 186198
Genotipo descendientes Genotipo descendientes Embrioacuten Nordm Oct3 Ov12 Embrioacuten Nordm Oct3 Ov12
1 139163 186315 1 139141 190198 2 139139 186315 2 139141 319198 3 139139 186315 3 139141 319198 4 139139 186315 4 146146 213186 5 139148 304194 5 141146 178186 6 139139 194194 6 146146 190186 7 163163 198194 7 141146 319198 8 139163 186194 8 130146 319198 9 139153 186194 9 180146 319198
10 139139 186194 10 146146 190186 11 163141 186315 11 146146 178198
1 Alelo 165 o alelo nulo maternopaterno
Estudios previos sobre ecologiacutea reproductiva de cefaloacutepodos han mostrado evidencias
de muacuteltiple paternidad La revisioacuten de la literatura indicoacute que la muacuteltiple paternidad es
26
consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

consecuencia de competencia masculina y del acoplamiento muacuteltiple en calamares
tales como Loligo forbesi (Shaw y Boyle 1997) Loligo pealeii (Buresch et al 2001)
Loligo bleekeri (Iwata et al 2005) en las jibias gigantes australianas Sepia apama
(Naud et al 2004) y en el calamar meridional Sepioteuthis australis (van Camp et al
2004) Sin embargo todas estas especies pertenecen al superorden Decabrachia y
por lo tanto nuestros resultados para O vulgaris son la primera evidencia de muacuteltiple
paternidad en el superorden Octobrachia lo que tambieacuten apoya la generalizacioacuten de
este comportamiento de acoplamiento conservado evolutivamente en cefaloacutepodos
coleoides
Por lo tanto la muacuteltiple paternidad en O vulgaris es un aspecto que debe ser tomado
en cuenta en geneacutetica poblacional y de conservacioacuten puesto que afecta al tamantildeo
eficaz de la poblacioacuten (Ne) (Karl 2008) y tambieacuten en el disentildeo y gestioacuten del cultivo
(para tener un cociente adecuado de machoshembras y seleccioacuten de reproductores)
32 Evaluacioacuten de la composicioacuten bioquiacutemica de juveniles de Artemia
enriquecidos con diferentes microalgas marinas
La utilizacioacuten de diferentes especies de microalgas para enriquecer los juveniles de
Artemia (tamantildeo inicial de 25 mm) durante 24 h no originoacute diferencias significativas
en el tamantildeo final de los individuos (Tabla IV) En la figura 7 se puede apreciar el
color del tracto digestivo de las artemias en funcioacuten de la microalga filtrada
27
AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

AISO
ARHOAT-ISO
ATET
Figura 7 - Juveniles de Artemia (27-35 mm) enriquecidos con las diferentes microalgas
marinas vistos a la lupa Se puede apreciar de forma notable el cambio de color del contenido
digestivo en funcioacuten del alimento ingerido ATET Artemia enriquecida con Tetraselmis suecica
AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida
con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens
Se han encontrado diferencias significativas en la composicioacuten nutricional de los
juveniles de Artemia (Tabla IV) El contenido proteico fue muy similar entre los grupos
(64-68 del peso seco) mientras que el contenido en liacutepidos varioacute entre el 10
(ARHO) y el 18 (AT-ISO) En cuanto al contenido en carbohidratos los individuos
enriquecidos con R lens presentaron claramente los valores maacutes bajos de todos La
mejor relacioacuten proteiacutena energiacutea se observoacute en el grupo enriquecido con Rhodomonas
(ARHO) El porcentaje maacutes elevado del aacutecido graso 205n-3 (EPA) se encontroacute en los
individuos del grupo AISO (146) seguido del grupo ARHO (Tabla V) El valor maacutes
elevado de 226n-3 (DHA) se observoacute en el grupo AT-ISO (29 del total de AG)
28
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
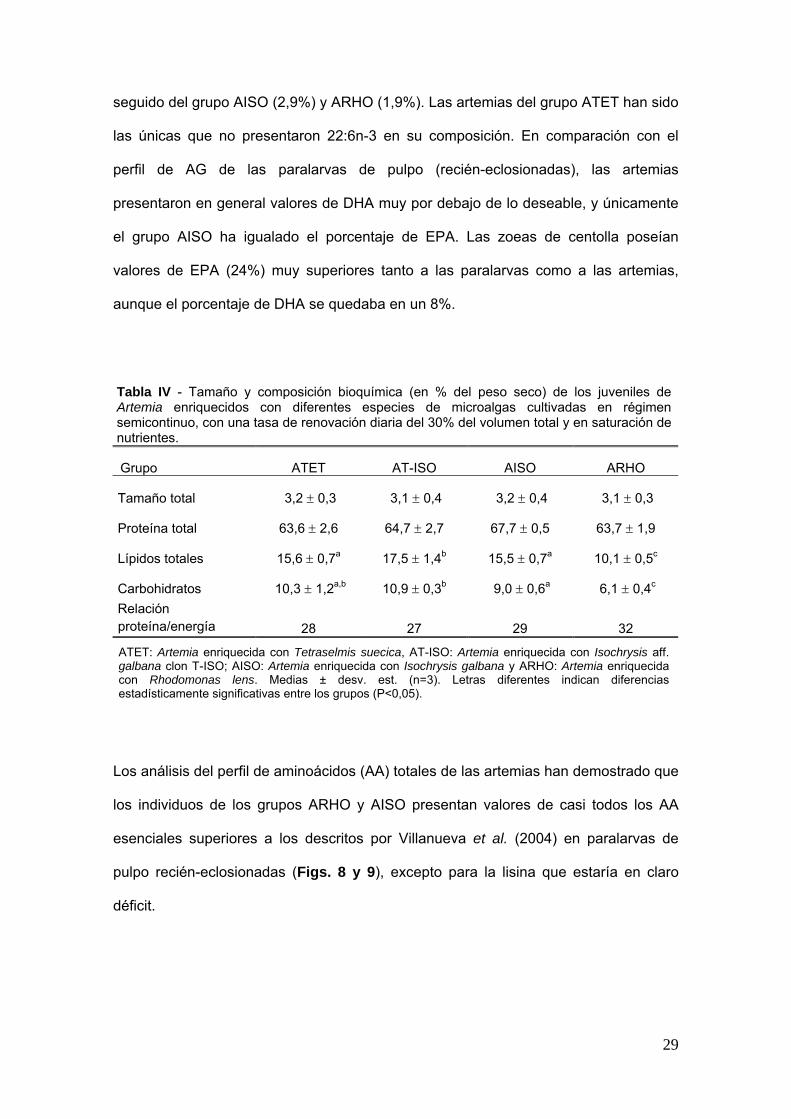
seguido del grupo AISO (29) y ARHO (19) Las artemias del grupo ATET han sido
las uacutenicas que no presentaron 226n-3 en su composicioacuten En comparacioacuten con el
perfil de AG de las paralarvas de pulpo (recieacuten-eclosionadas) las artemias
presentaron en general valores de DHA muy por debajo de lo deseable y uacutenicamente
el grupo AISO ha igualado el porcentaje de EPA Las zoeas de centolla poseiacutean
valores de EPA (24) muy superiores tanto a las paralarvas como a las artemias
aunque el porcentaje de DHA se quedaba en un 8
Tabla IV - Tamantildeo y composicioacuten bioquiacutemica (en del peso seco) de los juveniles de Artemia enriquecidos con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 del volumen total y en saturacioacuten de nutrientes
Grupo ATET AT-ISO AISO ARHO
Tamantildeo total 32 plusmn 03 31 plusmn 04 32 plusmn 04 31 plusmn 03
Proteiacutena total 636 plusmn 26 647 plusmn 27 677 plusmn 05 637 plusmn 19
Liacutepidos totales 156 plusmn 07a 175 plusmn 14b 155 plusmn 07a 101 plusmn 05c
Carbohidratos 103 plusmn 12ab 109 plusmn 03b a c 90 plusmn 06 61 plusmn 04Relacioacuten proteiacutenaenergiacutea 28 27 29 32
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO Artemia enriquecida con Rhodomonas lens Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
Los anaacutelisis del perfil de aminoaacutecidos (AA) totales de las artemias han demostrado que
los individuos de los grupos ARHO y AISO presentan valores de casi todos los AA
esenciales superiores a los descritos por Villanueva et al (2004) en paralarvas de
pulpo recieacuten-eclosionadas (Figs 8 y 9) excepto para la lisina que estariacutea en claro
deacuteficit
29
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
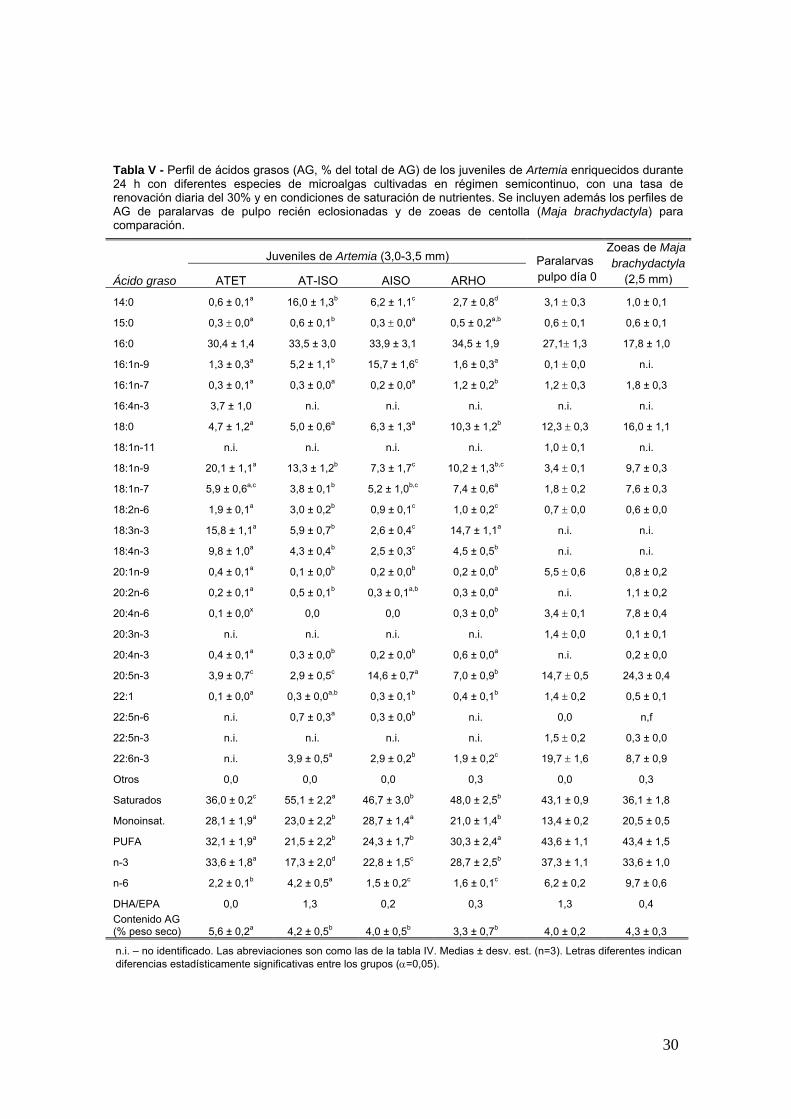
30
Tabla V - Perfil de aacutecidos grasos (AG del total de AG) de los juveniles de Artemia enriquecidos durante 24 h con diferentes especies de microalgas cultivadas en reacutegimen semicontinuo con una tasa de renovacioacuten diaria del 30 y en condiciones de saturacioacuten de nutrientes Se incluyen ademaacutes los perfiles de AG de paralarvas de pulpo recieacuten eclosionadas y de zoeas de centolla (Maja brachydactyla) para comparacioacuten
Juveniles de Artemia (30-35 mm)
Aacutecido graso ATET AT-ISO AISO ARHO
Paralarvas pulpo diacutea 0
Zoeas de Maja brachydactyla
(25 mm)
140 06 plusmn 01a 160 plusmn 13b 62 plusmn 11c 27 plusmn 08d 31 plusmn 03 10 plusmn 01
150 03 plusmn 00a 06 plusmn 01b 03 plusmn 00a 05 plusmn 02ab 06 plusmn 01 06 plusmn 01
160 304 plusmn 14 335 plusmn 30 339 plusmn 31 345 plusmn 19 271plusmn 13 178 plusmn 10
161n-9 13 plusmn 03a 52 plusmn 11b 157 plusmn 16c 16 plusmn 03a 01 plusmn 00 ni
161n-7 03 plusmn 01a 03 plusmn 00a 02 plusmn 00a 12 plusmn 02b 12 plusmn 03 18 plusmn 03
164n-3 37 plusmn 10 ni ni ni ni ni
180 47 plusmn 12a 50 plusmn 06a 63 plusmn 13a 103 plusmn 12b 123 plusmn 03 160 plusmn 11
181n-11 ni ni ni ni 10 plusmn 01 ni
181n-9 201 plusmn 11a 133 plusmn 12b 73 plusmn 17c 102 plusmn 13bc 34 plusmn 01 97 plusmn 03
181n-7 59 plusmn 06ac 38 plusmn 01b 52 plusmn 10bc 74 plusmn 06a 18 plusmn 02 76 plusmn 03
182n-6 19 plusmn 01a 30 plusmn 02b 09 plusmn 01c 10 plusmn 02c 07 plusmn 00 06 plusmn 00
183n-3 158 plusmn 11a 59 plusmn 07b 26 plusmn 04c 147 plusmn 11a ni ni
184n-3 98 plusmn 10a 43 plusmn 04b 25 plusmn 03c 45 plusmn 05b ni ni
201n-9 04 plusmn 01a 01 plusmn 00b 02 plusmn 00b 02 plusmn 00b 55 plusmn 06 08 plusmn 02
202n-6 02 plusmn 01a 05 plusmn 01b 03 plusmn 01ab 03 plusmn 00a ni 11 plusmn 02
204n-6 01 plusmn 00x 00 00 03 plusmn 00b 34 plusmn 01 78 plusmn 04
203n-3 ni ni ni ni 14 plusmn 00 01 plusmn 01
204n-3 04 plusmn 01a 03 plusmn 00b 02 plusmn 00b 06 plusmn 00a ni 02 plusmn 00
205n-3 39 plusmn 07c 29 plusmn 05c 146 plusmn 07a 70 plusmn 09b 147 plusmn 05 243 plusmn 04
221 01 plusmn 00a 03 plusmn 00ab 03 plusmn 01b 04 plusmn 01b 14 plusmn 02 05 plusmn 01
225n-6 ni 07 plusmn 03a 03 plusmn 00b ni 00 nf
225n-3 ni ni ni ni 15 plusmn 02 03 plusmn 00
226n-3 ni 39 plusmn 05a 29 plusmn 02b 19 plusmn 02c 197 plusmn 16 87 plusmn 09
Otros 00 00 00 03 00 03
Saturados 360 plusmn 02c 551 plusmn 22a 467 plusmn 30b 480 plusmn 25b 431 plusmn 09 361 plusmn 18
Monoinsat 281 plusmn 19a 230 plusmn 22b 287 plusmn 14a 210 plusmn 14b 134 plusmn 02 205 plusmn 05
PUFA 321 plusmn 19a 215 plusmn 22b 243 plusmn 17b 303 plusmn 24a 436 plusmn 11 434 plusmn 15
n-3 336 plusmn 18a 173 plusmn 20d 228 plusmn 15c 287 plusmn 25b 373 plusmn 11 336 plusmn 10
n-6 22 plusmn 01b 42 plusmn 05a 15 plusmn 02c 16 plusmn 01c 62 plusmn 02 97 plusmn 06
DHAEPA 00 13 02 03 13 04 Contenido AG ( peso seco) 56 plusmn 02a 42 plusmn 05b 40 plusmn 05b 33 plusmn 07b 40 plusmn 02 43 plusmn 03
ni ndash no identificado Las abreviaciones son como las de la tabla IV Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (α=005)
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
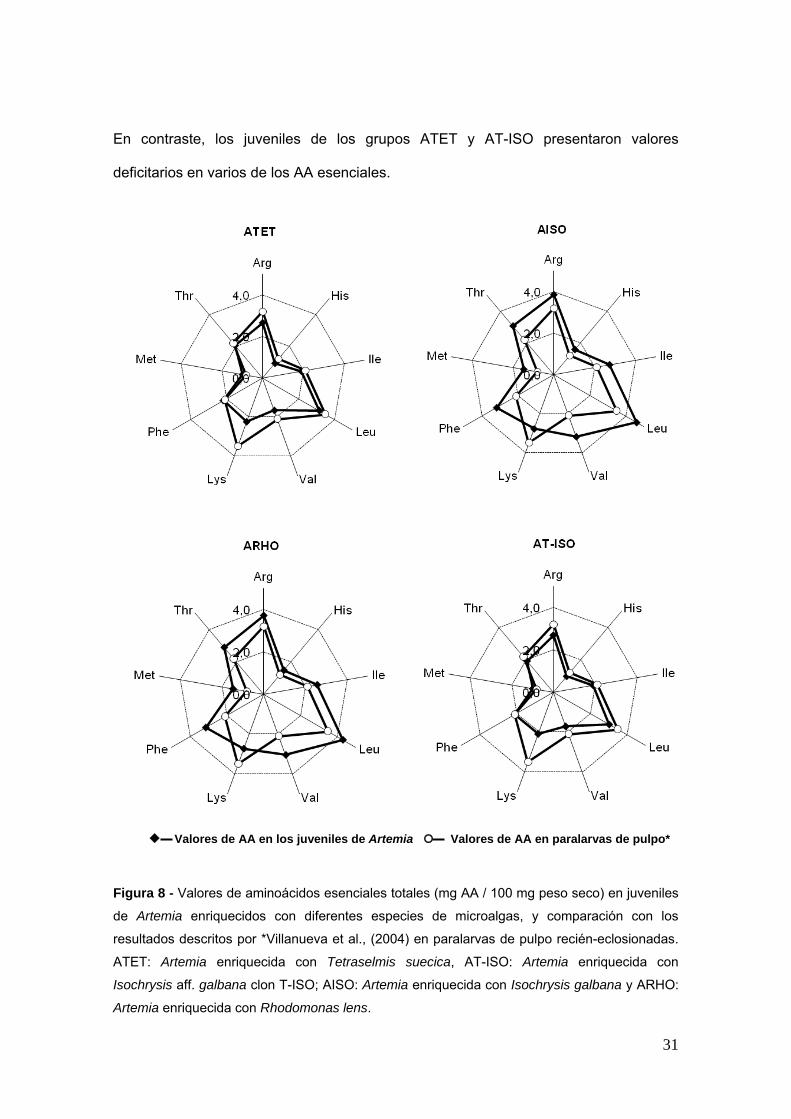
En contraste los juveniles de los grupos ATET y AT-ISO presentaron valores
deficitarios en varios de los AA esenciales
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 8 - Valores de aminoaacutecidos esenciales totales (mg AA 100 mg peso seco) en juveniles
de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con los
resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-eclosionadas
ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia enriquecida con
Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis galbana y ARHO
Artemia enriquecida con Rhodomonas lens
31
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
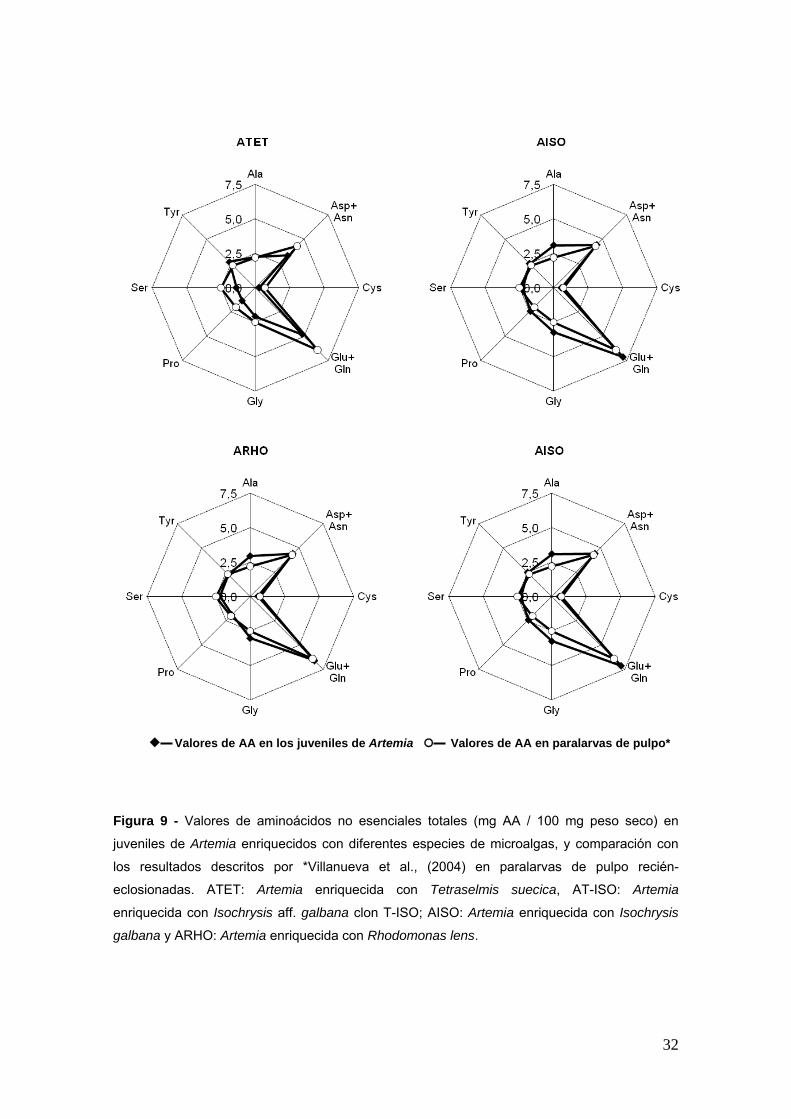
Valores de AA en los juveniles de Artemia Valores de AA en paralarvas de pulpo
Figura 9 - Valores de aminoaacutecidos no esenciales totales (mg AA 100 mg peso seco) en
juveniles de Artemia enriquecidos con diferentes especies de microalgas y comparacioacuten con
los resultados descritos por Villanueva et al (2004) en paralarvas de pulpo recieacuten-
eclosionadas ATET Artemia enriquecida con Tetraselmis suecica AT-ISO Artemia
enriquecida con Isochrysis aff galbana clon T-ISO AISO Artemia enriquecida con Isochrysis
galbana y ARHO Artemia enriquecida con Rhodomonas lens
32
Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

Los cefaloacutepodos tienen una gran dependencia en el consumo de dietas ricas en
proteiacutena debido a sus elevadas tasas de crecimiento y a la utilizacioacuten de aminoaacutecidos
como fuente de energiacutea (Houlihan et al 1990 Lee 1994) Es por eso aconsejable
seleccionar presas ricas en proteiacutena o con una relacioacuten proteiacutenaenergiacutea elevada En
este trabajo el contenido proteico de las artemias enriquecidas con diferentes
microalgas varioacute entre el 60 y el 65 similar al contenido proteico de estadios
iniciales de vida del pulpo (Villanueva et al 2004) La utilizacioacuten de cultivos
semicontinuos de microalgas en condiciones optimizadas incrementa su contenido en
proteiacutenas que se refleja a su vez en la composicioacuten de Artemia sp (Faacutebregas et al
2001) La mejor relacioacuten proteiacutenaenergiacutea se observoacute en las artemias del grupo ARHO
siendo este factor importante para los cefaloacutepodos Se ha demostrado que en sepias
juveniles la utilizacioacuten de dietas con la mayor relacioacuten proteiacutenaenergiacutea originaba las
mejores tasas de crecimiento de los individuos (Lee 1994) Admitiendo que un alto
contenido proteico o relacioacuten proteiacutenaenergiacutea en las presas vivas es fundamental
para el buen crecimiento de las paralarvas importa averiguar si el perfil de
aminoaacutecidos de las presas se ajusta a los posibles requerimientos de aquellas
Villanueva et al (2004) han analizado el perfil de aminoaacutecidos totales de paralarvas de
pulpo recieacuten eclosionadas y de juveniles salvajes suponiendo este estudio un buen
punto de partida para comparar los perfiles de aminoaacutecidos tanto cualitativos como
cuantitativos de presas vivas y de paralavas En este trabajo los primeros anaacutelisis de
aminoaacutecidos totales de las artemias enriquecidas con diferentes microalgas han
demostrado la existencia de diferencias apreciables entre los grupos de artemias y un
posible deacuteficit del aminoaacutecido esencial lisina en los juveniles de Artemia enriquecidos
con R lens o I galbana (los dos mejores grupos de Artemia) En caso de que la lisina
fuese el primer aminoaacutecido limitante el crecimiento de las paralarvas estariacutea
condicionado por la cantidad y disponibilidad de eacuteste en las presas por lo que se
tendriacutea que enriquecer las artemias con este aminoaacutecido esencial u otros que podriacutean
pasar a ser el limitante hasta conseguir el balance adecuado El uso de microalgas
33
para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

para enriquecer presas vivas en acuicultura es una teacutecnica habitual y que vuelve a
tener particular intereacutes ya que los perfiles de aminoaacutecidos (tanto en cantidad como en
calidad) suelen ser mejores que el uso de otros enriquecedores comerciales (Aragatildeo
et al 2004)
Los porcentajes de los PUFAs 205n-3 y 226n-3 en las artemias fueron en general
muy inferiores a los observados en las paralarvas o en las zoeas de centolla como
presa de comparacioacuten ya que eacutestas han sido indicadas como adecuadas para el
cultivo de paralarvas (Iglesias et al 2004 Carrasco et al 2006) Las paralarvas de
pulpo presentan porcentajes muy elevados de los aacutecidos grasos 205n-3 (asymp14) y de
226n-3 (asymp21) y requerimientos en fosfoliacutepidos y colesterol igualmente elevados
(Navarro y Villanueva 2000) por lo que se requiere incrementar los valores de estos
AG en las artemias con emulsiones lipiacutedicas especiacuteficas y muy ricas en estos PUFAs
Sin embargo el uso de microalgas de composicioacuten controlada como dieta para
enriquecer las artemias (enriquecimiento con R lens o con I galbana) ha originado
porcentajes de 205n-3 y 226n-3 iguales o superiores a los valores descritos por
Navarro y Villanueva (2000) en Artemia sp (1-3 mm) enriquecida con emulsiones
lipiacutedicas comerciales (Selcos INVE)
33 Efectos de la dieta sobre el crecimiento y la supervivencia de paralarvas de
pulpo
La composicioacuten bioquiacutemica de los juveniles de Artemia utilizados para alimentar las
paralarvas se presenta en la tabla VII No se encontraron diferencias significativas en
el contenido de proteiacutenas liacutepidos y carbohidratos de las diferentes dietas Sin
embargo la fraccioacuten de AAL fue considerablemente superior en los grupos AR+I y
AR+I+AA que en el grupo control (Plt005)
34
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
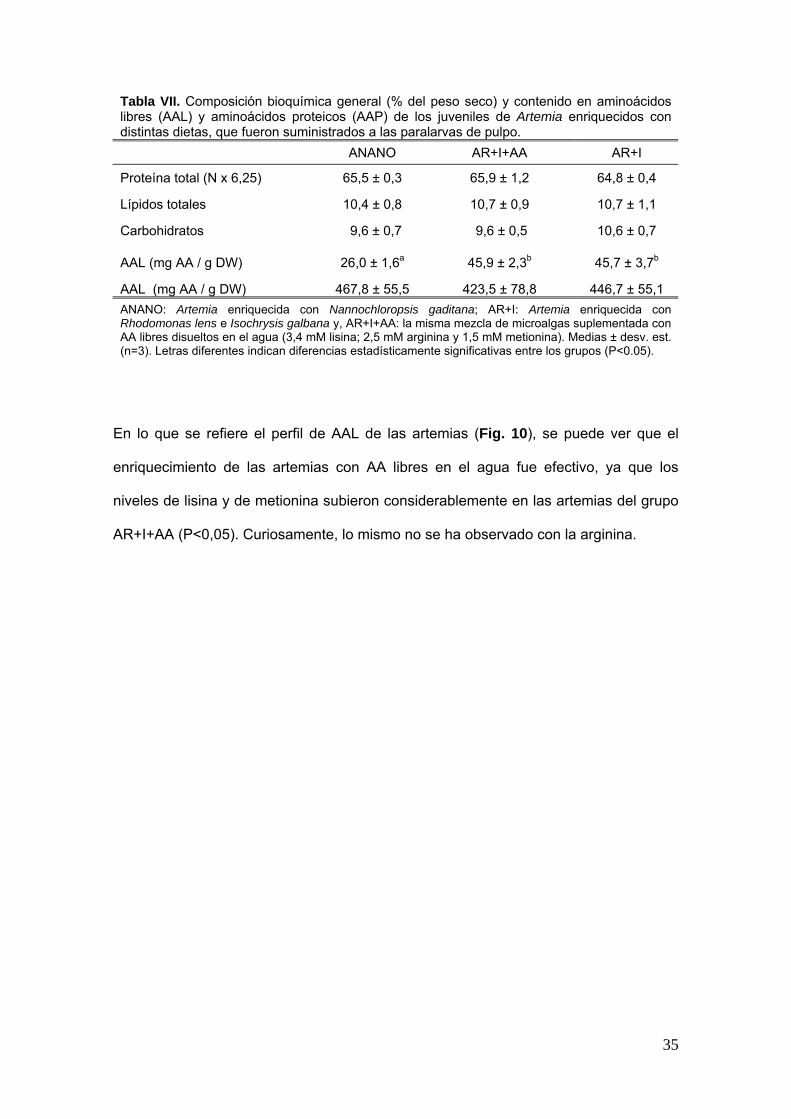
Tabla VII Composicioacuten bioquiacutemica general ( del peso seco) y contenido en aminoaacutecidos libres (AAL) y aminoaacutecidos proteicos (AAP) de los juveniles de Artemia enriquecidos con distintas dietas que fueron suministrados a las paralarvas de pulpo
ANANO AR+I+AA AR+I
Proteiacutena total (N x 625) 655 plusmn 03 659 plusmn 12 648 plusmn 04
Liacutepidos totales 104 plusmn 08 107 plusmn 09 107 plusmn 11
Carbohidratos 96 plusmn 07 96 plusmn 05 106 plusmn 07
AAL (mg AA g DW) 260 plusmn 16a 459 plusmn 23b 457 plusmn 37b
AAL (mg AA g DW) 4678 plusmn 555 4235 plusmn 788 4467 plusmn 551 ANANO Artemia enriquecida con Nannochloropsis gaditana AR+I Artemia enriquecida con Rhodomonas lens e Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En lo que se refiere el perfil de AAL de las artemias (Fig 10) se puede ver que el
enriquecimiento de las artemias con AA libres en el agua fue efectivo ya que los
niveles de lisina y de metionina subieron considerablemente en las artemias del grupo
AR+I+AA (Plt005) Curiosamente lo mismo no se ha observado con la arginina
35
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
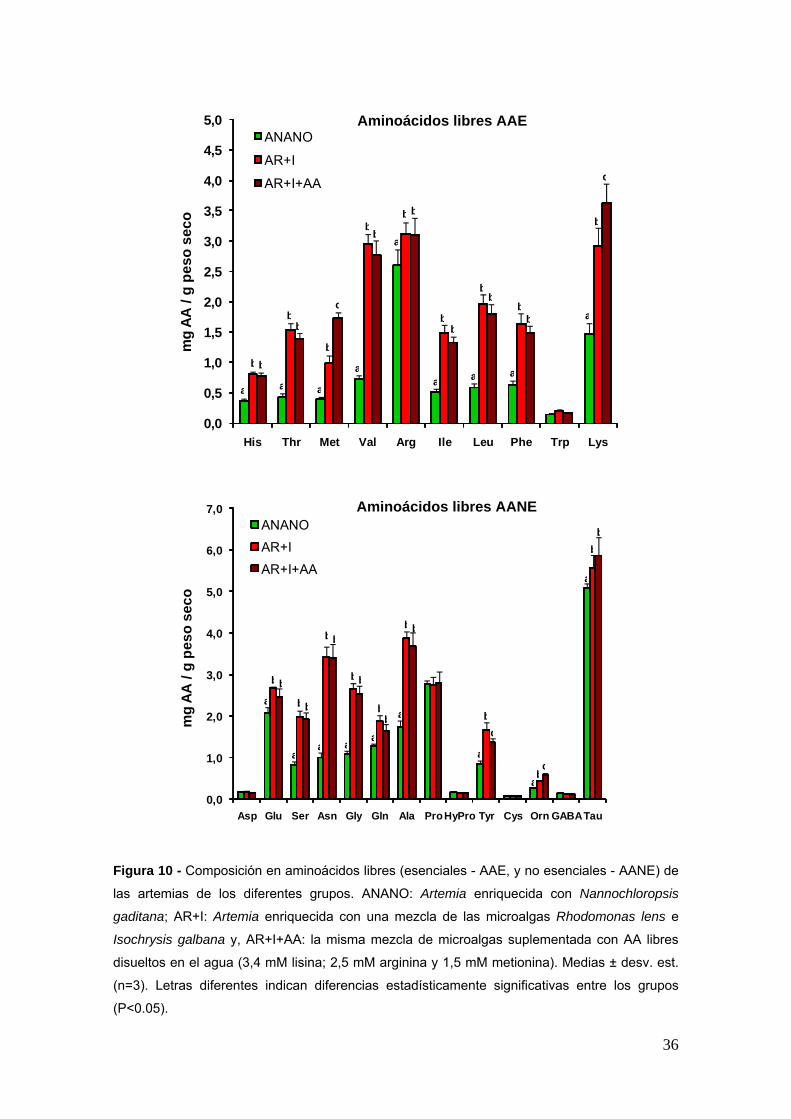
00
05
10
15
20
25
30
35
40
45
50
His Thr Met Val Arg Ile Leu Phe Trp Lys
mg
AA
g pe
so s
eco
Amino acidos libres AAEANANO
AR+I
AR+I+AA
a
b b
bb
a
c
b
a
bb
a
b b
a
bb
a
bb
a
bb
a
c
b
a
Aminoaacutecidos libres AAE
00
10
20
30
40
50
60
70
Asp Glu Ser Asn Gly Gln Ala ProHyPro Tyr Cys Orn GABATau
mg
AA
g pe
so s
eco
Amino acidos libres AANEANANOAR+IAR+I+AA
abb
a
bb
bb
bb
b
a a a
bb
cb
cb
a
a
a
bb
a
b
Aminoaacutecidos libres AANE
Figura 10 - Composicioacuten en aminoaacutecidos libres (esenciales - AAE y no esenciales - AANE) de
las artemias de los diferentes grupos ANANO Artemia enriquecida con Nannochloropsis
gaditana AR+I Artemia enriquecida con una mezcla de las microalgas Rhodomonas lens e
Isochrysis galbana y AR+I+AA la misma mezcla de microalgas suplementada con AA libres
disueltos en el agua (34 mM lisina 25 mM arginina y 15 mM metionina) Medias plusmn desv est
(n=3) Letras diferentes indican diferencias estadiacutesticamente significativas entre los grupos
(Plt005)
36
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
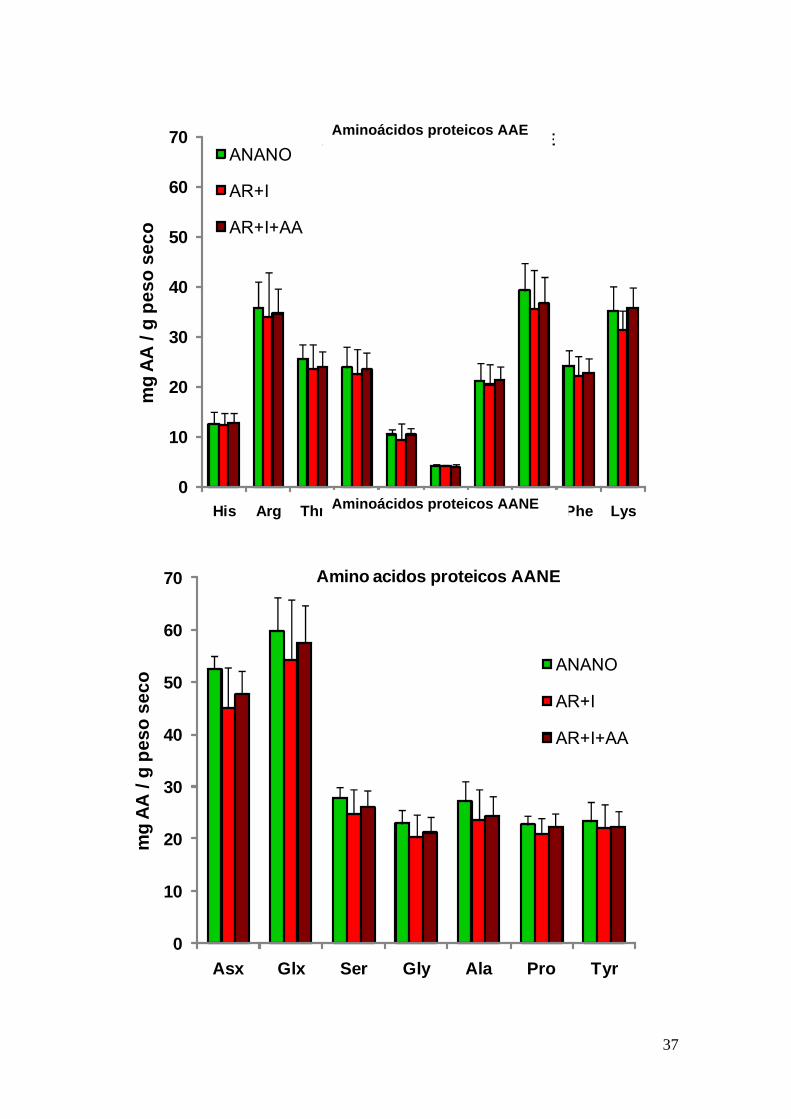
0
10
20
30
40
50
60
70
His Arg Thr Val Met Cys Ile Leu Phe Lys
mg
AA
g pe
so s
eco
Amino acidos proteicos AAEAminoaacutecidos proteicos AAE
ANANO
AR+I
AR+I+AA
Aminoaacutecidos proteicos AANE
0
10
20
30
40
50
60
70
Asx Glx Ser Gly Ala Pro Tyr
mg
AA
g pe
so s
eco
Amino acidos proteicos AANE
ANANO
AR+I
AR+I+AA
37
Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

Figura 11 - Composicioacuten en aminoaacutecidos proteicos (esenciales - AAE y no esenciales - AANE)
en las artemias de los diferentes grupos Las abreviaciones son como las de la figura anterior
Medias plusmn desv est (n=3) Letras diferentes indican diferencias estadiacutesticamente significativas
entre los grupos (Plt005)
38
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
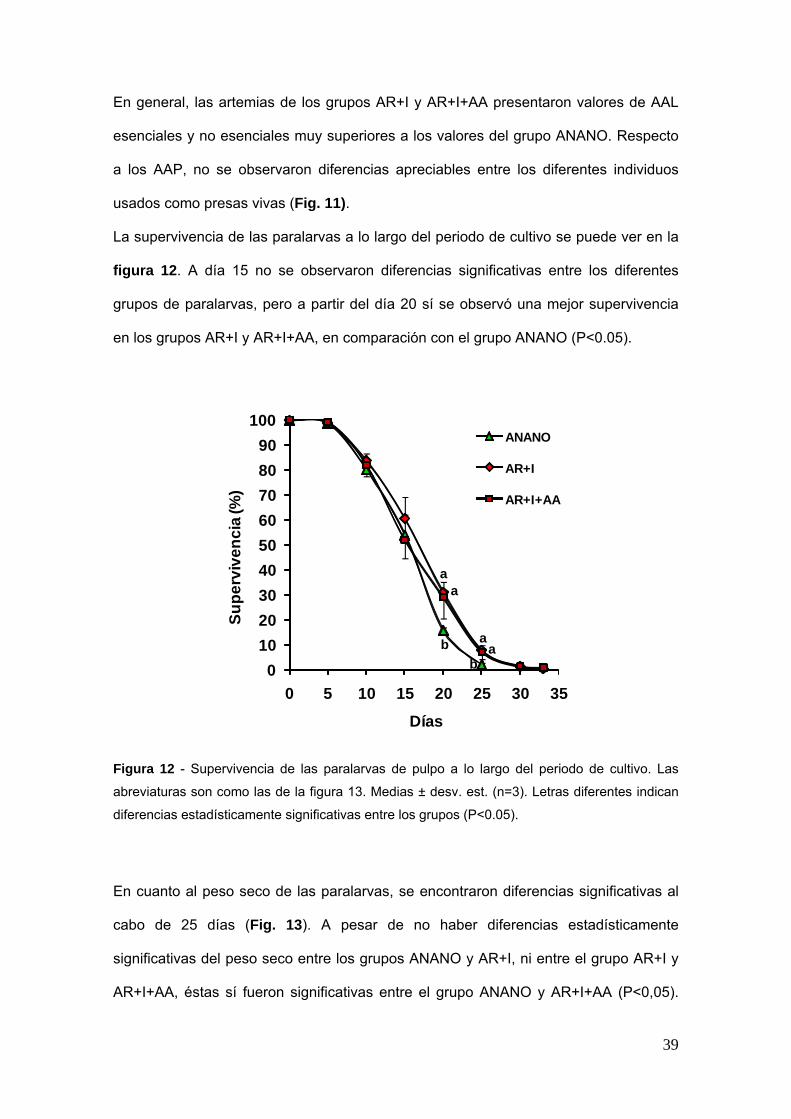
En general las artemias de los grupos AR+I y AR+I+AA presentaron valores de AAL
esenciales y no esenciales muy superiores a los valores del grupo ANANO Respecto
a los AAP no se observaron diferencias apreciables entre los diferentes individuos
usados como presas vivas (Fig 11)
La supervivencia de las paralarvas a lo largo del periodo de cultivo se puede ver en la
figura 12 A diacutea 15 no se observaron diferencias significativas entre los diferentes
grupos de paralarvas pero a partir del diacutea 20 siacute se observoacute una mejor supervivencia
en los grupos AR+I y AR+I+AA en comparacioacuten con el grupo ANANO (Plt005)
0102030405060708090
100
0 5 10 15 20 25 30 35
Supe
rviv
enci
a (
)
Diacuteas
ANANO
AR+I
AR+I+AA
aa
b aa
b
Figura 12 - Supervivencia de las paralarvas de pulpo a lo largo del periodo de cultivo Las
abreviaturas son como las de la figura 13 Medias plusmn desv est (n=3) Letras diferentes indican
diferencias estadiacutesticamente significativas entre los grupos (Plt005)
En cuanto al peso seco de las paralarvas se encontraron diferencias significativas al
cabo de 25 diacuteas (Fig 13) A pesar de no haber diferencias estadiacutesticamente
significativas del peso seco entre los grupos ANANO y AR+I ni entre el grupo AR+I y
AR+I+AA eacutestas siacute fueron significativas entre el grupo ANANO y AR+I+AA (Plt005)
39
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
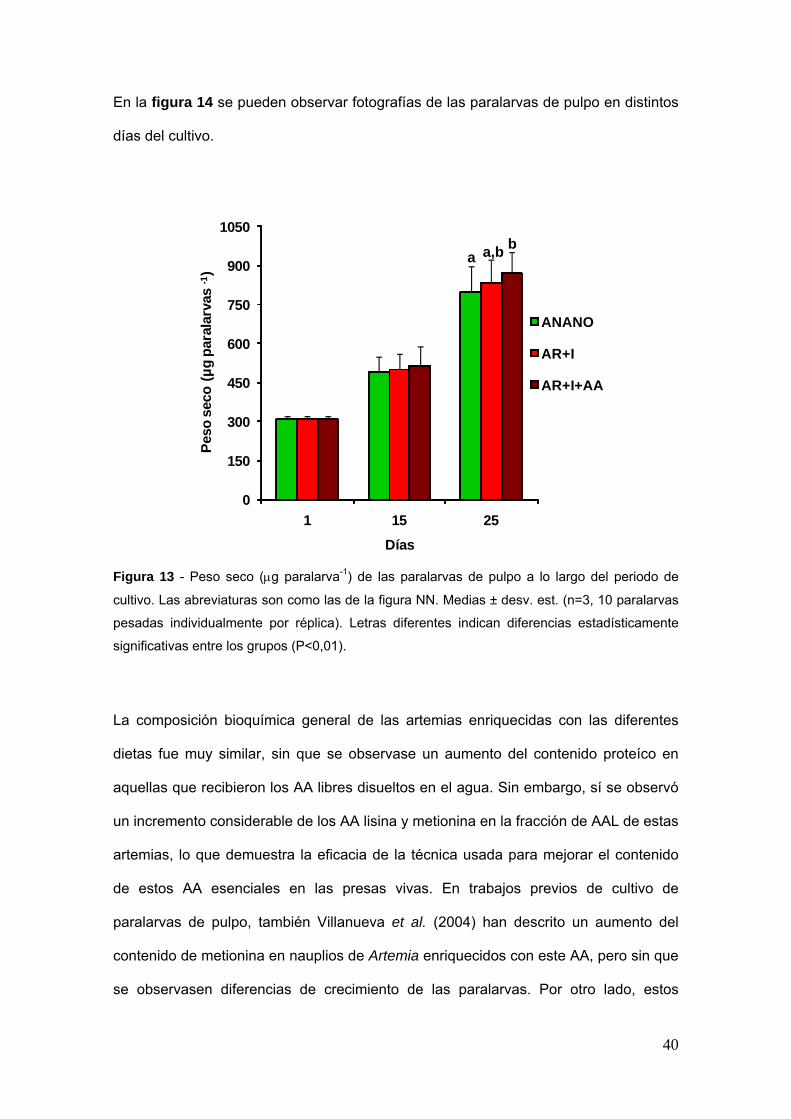
En la figura 14 se pueden observar fotografiacuteas de las paralarvas de pulpo en distintos
diacuteas del cultivo
0
150
300
450
600
750
900
1050
1 15 25
Peso
sec
o (micro
g pa
rala
rvas
-1)
Diacuteas
ANANO
AR+I
AR+I+AA
abab
Figura 13 - Peso seco (μg paralarva-1) de las paralarvas de pulpo a lo largo del periodo de
cultivo Las abreviaturas son como las de la figura NN Medias plusmn desv est (n=3 10 paralarvas
pesadas individualmente por reacuteplica) Letras diferentes indican diferencias estadiacutesticamente
significativas entre los grupos (Plt001)
La composicioacuten bioquiacutemica general de las artemias enriquecidas con las diferentes
dietas fue muy similar sin que se observase un aumento del contenido proteiacuteco en
aquellas que recibieron los AA libres disueltos en el agua Sin embargo siacute se observoacute
un incremento considerable de los AA lisina y metionina en la fraccioacuten de AAL de estas
artemias lo que demuestra la eficacia de la teacutecnica usada para mejorar el contenido
de estos AA esenciales en las presas vivas En trabajos previos de cultivo de
paralarvas de pulpo tambieacuten Villanueva et al (2004) han descrito un aumento del
contenido de metionina en nauplios de Artemia enriquecidos con este AA pero sin que
se observasen diferencias de crecimiento de las paralarvas Por otro lado estos
40
autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

autores habiacutean probado disolver AA directamente en los tanques de cultivo de las
paralarvas pero tampoco notaron mejoras en el crecimiento de las mismas Sin
embargo la mejor supervivencia observada para este grupo podriacutea estar relacionada
con la capacidad de las paralarvas en absorber AA a traveacutes de la piel (Lee 1994) que
les dariacutea alguna posible ventaja en la obtencioacuten de energiacutea
El peso seco de las paralarvas observado en este trabajo a los 25 diacuteas de vida (798-
870 microg paralarva-1) fue superior a los valores descritos por Villanueva et al (2004) en
paralarvas alimentadas con nauplios de Artemia enriquecidos con Super-Selco (INVE)
o complementados con metionina disuelta o que recibieron los AA disueltos en el
agua (650-718 microg paralarva-1) lo que indica que las dietas usadas en este estudio
para enriquecer las artemias (microalgas y una mezcla de AA esenciales libres) fueron
maacutes adecuadas y ventajosas Todo indica que vehicular AA esenciales a traveacutes de
Artemia es una teacutecnica maacutes eficaz que el uso de AA disueltos en el agua por lo que se
deberiacutea de profundizar esta liacutenea de investigacioacuten
En este trabajo aunque no se observasen diferencias estadiacutesticamente significativas
en el peso seco de las paralarvas en las dos primeras semanas de cultivo a pesar de
haber ya una cierta tendencia para el efecto beneacutefico de antildeadir los AA esenciales siacute
se han encontrado diferencias importantes al cabo de 25 diacuteas El grupo control
alimentando con juveniles de Artemia enriquecidos con Nannochloropsis previamente
probado por otros autores y descrito como un protocolo mejorado para el cultivo de
paralarvas (Hamazaki et al 1991 Moxica et al 2006) ha generado peores resultados
que la utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) aunque no se registrasen diferencias estadiacutesticamente significativas En
contraste el uso combinado de la mezcla R lens e I galbana complementada con los
AA esenciales lisina metionina y arginina originoacute resultados claramente ventajosos
para el cultivo de las paralarvas
41
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48
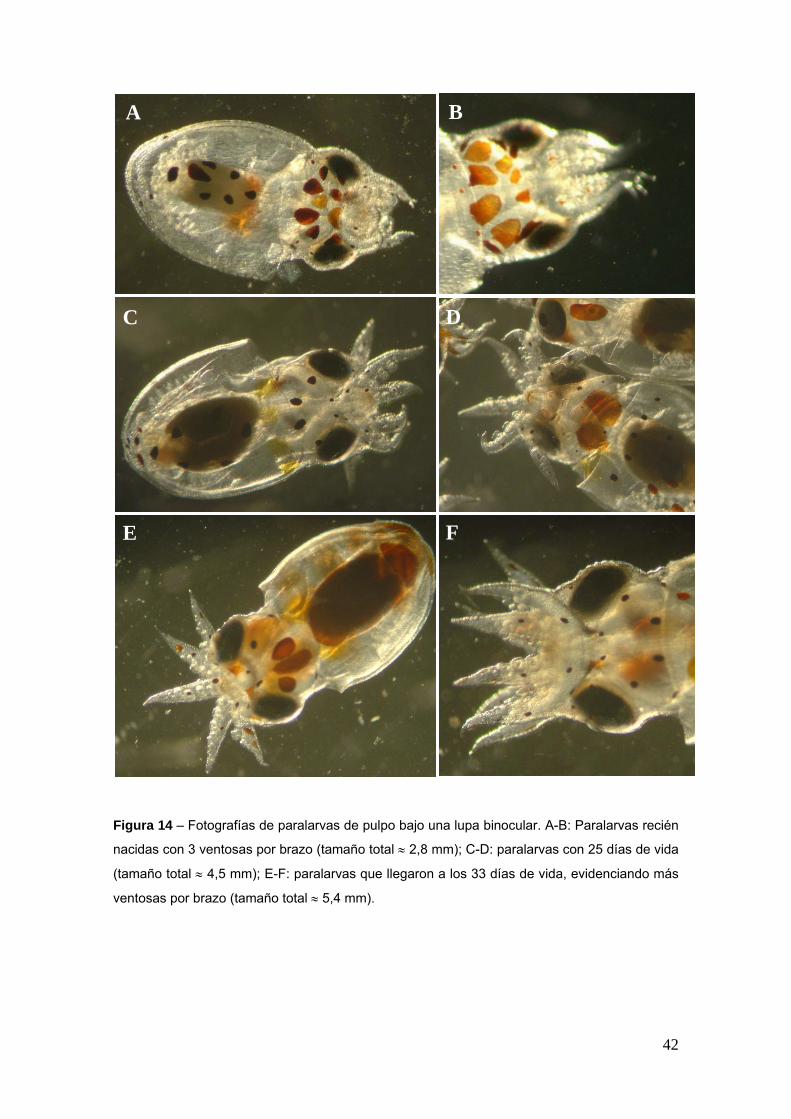
A B
C D
E F
Figura 14 ndash Fotografiacuteas de paralarvas de pulpo bajo una lupa binocular A-B Paralarvas recieacuten
nacidas con 3 ventosas por brazo (tamantildeo total asymp 28 mm) C-D paralarvas con 25 diacuteas de vida
(tamantildeo total asymp 45 mm) E-F paralarvas que llegaron a los 33 diacuteas de vida evidenciando maacutes
ventosas por brazo (tamantildeo total asymp 54 mm)
42
Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

Alcanzar un balance adecuado de AA en las dietas iniciales de peces u de otras
especies diana es un reto importante en acuicultura (Aragatildeo et al 2004) debido a las
elevadas exigencias nutricionales de las larvas
Estos organismos en sus etapas iniciales de vida requieren presas muy bien
balanceadas en proteiacutena y AA esenciales en liacutepidos estructurales (especialmente en
PUFAs) y en pigmentos y minerales ya que sus aparatos digestivos estaacuten a menudo
auacuten poco desarrollados y a penas son capaces de sintetizar ciertos compuestos o de
recurrir a las viacuteas metaboacutelicas necesarias para ese fin Estos factores aliados a las
altas tasas de crecimiento que suelen tener obligan a la ingestioacuten de presas muy
completas nutricionalmente Por ello la modulacioacuten de la composicioacuten nutricional de
Artemia sigue siendo clave para mejorar el crecimiento y la supervivencia de las
paralarvas hasta que se pueda cerrar el ciclo de vida del pulpo en cautiverio de forma
rentable y controlada La formulacioacuten de microdietas inertes es otra estrategia clave
para complementar el uso de Artemia como presa viva con el fin de aportar a las
paralarvas todos los nutrientes esenciales requeridos para su buen desarrollo
43
Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

Conclusiones
Ha sido posible demostrar la existencia de muacuteltiple paternidad en O vulgaris La
paternidad muacuteltiple puede por lo tanto representar un proceso que maximiza la
recombinacioacuten geneacutetica con los muacuteltiples machos en un uacutenico acontecimiento
reproductivo El cuidado exhaustivo del huevo garantiza ademaacutes un nivel adecuado de
supervivencia de una descendencia geneacuteticamente diversa
En cuanto a la mejora del valor nutricional de las presas vivas el enriquecimiento de
juveniles de Artemia con distintas especies de microalgas ha generado diferencias
considerables en su composicioacuten bioquiacutemica
En el ensayo de cultivo de paralarvas con diferentes dietas se observoacute que los
juveniles de Artemia enriquecidos con Nannochloropsis gaditana previamente
utilizados por otros autores y descritas como protocolo mejorado para el cultivo de
paralarvas han generado peores resultados de crecimiento y supervivencia que la
utilizacioacuten de juveniles enriquecidos con Rhodomonas lens e Isochrysis galbana
(AR+I) La complementacioacuten de las estas artemias con aminoaacutecidos esenciales (lisina
arginina y metionina) disueltos en el agua supuso ademaacutes una mejora del peso seco
de las paralarvas por lo que seriacutea uacutetil profundizar esta nueva liacutenea de investigacioacuten
El conocimiento de aspectos baacutesicos de la biologiacutea reproductiva del pulpo y de la
nutricioacuten de las paralarvas es fundamental para avanzar en el cultivo integral de esta
especie pudiendo igualmente ser uacutetil en el futuro para regular las pesqueriacuteas y hacer
una gestioacuten sostenible de las poblaciones naturales
44
Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

Referencias citadas
Adam GJ Ardren WR (2008) Methods of parentage analysis in natural populations Mol Ecol 12 2511-2523
Aragatildeo C Conceiccedilatildeo LEC Dinis MT Fyhn HJ 2004 Amino acid pools of rotifers and Artemia under different conditions nutritional implications for fish larvae Aquaculture 234 429-445
Bligh EG Dyer WJ 1959 A rapid method of total lipid extraction and purification Can J Biochem Physiol 37 911-917
Boletzky S v 1975 A contribution to the study of the yolk absorption in the Cephalopoda Springer-Verlag 1975 Z Morph Tiere 80 229-246
Boletzky Sv 1987 Embryonic phase In Boyle PR (Ed) Cephalopod Life Cycles Comparative Reviews Vol II Academic Press London pp 23ndash 25
Boletzky Sv Hanlon RT 1983 A review of the laboratory maintenance rearing and culture of cephalopod molluscs Memoirs of the National Museum 44 147-187
Boyle PR (Ed) 1983 Cephalopod Life Cycles Comparative Reviews Vol II Academic Press Inc London 441 pp
Buresch KM Hanlon RT Maxwell MR Ring S (2001) Microsatellite DNA markers indicate a high frequency of multiple paternity within individual field-collected egg capsules of the squid Loligo pealeii Mar Ecol Prog Ser 210 161-165
Cabranes C Fernandez-Rueda P Martinez JL (2008) Genetic structure of Octopus vulgaris around the Iberian Peninsula and Canary Islands as indicated by microsatellite DNA variation ICES J Mar Sci 65 12-16
Carrasco JF Rodriacuteguez C Rodriacuteguez M 2003 Cultivo intensivo de paralarvas de pulpo (Octopus vulgaris Cuvier 1797) utilizando como base de la alimentacioacuten zoeas vivas de crustaacuteceos IX Congreso Nacional de Acuicultura Caacutediz Espantildea 12ndash16 Maio
Carrasco JF Arronte JC Rodriacuteguez C 2006 Paralarval rearing of the common octopus Octopus vulgaris (Cuvier) Aquac Res 37 1601ndash1605
Faacutebregas J Herrero C Cabezas B Abalde J 1986 Biomass production and biochemical composition in mass culture of the marine microalga Isochrysis galbana Parke at varying nutrient concentrations Aquaculture 53 101-113
Faacutebregas J Otero A Morales E Cordero B Patintildeo M 1996 Tetraselmis suecica cultured in different nutrient concentrations varies in nutritional value to Artemia Aquaculture 143 197-204
Faacutebregas J Otero A Dominguez A Patintildeo M 2001 Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species Mar Biotechnol 3 256-263
Food and Agriculture Organization of the United Nations (FAO) 2003 Species identification sheets httpwwwfaoorgfigisservletFiRefServletds=speciesampfid=3571
Forsythe JW Van Heukelem WF 1987 Growth In Cephalopod Life Cycles (Ed Boyle PR) Vol II Comparative Reviews Academic Press Inc (London) Ltd 135-156
Ginzburg M 1987 Dunaliella a green alga adapted to salt Adv Bot Res 14 93-183
Goodnight KF Queller DC (1999) Computer software for performing likelihood tests of pedigree relationship using genetic markers Mol Ecol 8 1231-1234
Greatorex EC Jones CS Murphy J Key LN Emery AM Boyle PR (2000) Microsatellite markers for investigating population structure in Octopus vulgaris (Mollusca Cephalopoda) Mol Ecol 9 641-642
Hamazaki H Fukunaga K Yoshida Y Maruyama K 1991 Effects of marine Nannochloropsis sp on survival and growth on rearing pelagic paralarvae of
45
Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

Octopus vulgaris and results of mass culture in the tank of 20 metric tons Saibai-Giken 19 75-84 (in Japanese)
Hanlon RT Messenger 1996 Cephalopod Behaviour University Press Cambridge 232 pp
Hatanaka H 1979 Studies on the fisheries biology of common octopus off northwest coast of Africa Bull Far Seas Fish Res Lab 17 13ndash124
Houlihan DF McMillan DN Agnisola C Trara Genoino I Foti L 1990 Protein synthesis and growth in Octopus vulgaris Mar Biol 106 251-259
Iglesias J Saacutenchez FJ Otero JJ 1997 Primeras experiencias sobre el cultivo integral del pulpo (Octopus vulgaris) en el Instituto Espantildeol de Oceanografiacutea En Costa J Abellan E Garciacutea B Ortega A y Zamara S (Eds) Actas del VI Congreso Nacional de Acuicultura Cartagena ISBN 84-491-0323-1 pp 221-226
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2000 Culture of octopus (Octopus vulgaris Cuvier) Present knowledge problems and perspectives Cah Options Meacutediterr 47 313ndash321
Iglesias J Otero JJ Moxica C Saacutenchez FJ 2002 Paralarvae culture of octopus (Octopus vulgaris Cuvier) using Artemia and zoeas and first data on juvenile growth up to eight months of age European Aquaculture Society Special Publication No 32 268-269
Iglesias J Saacutenchez FJ Otero JJ Moxica C 2003 El cultivo del pulpo (Octopus vulgaris Cuvier) Monografiacutea Ministerio de Agricultura Pesca y Alimentacioacuten Instituto Espantildeol de Oceanografiacutea 23 pp
Iglesias J Otero JJ Moxica C Fuentes L Saacutenchez FJ 2004 The completed life cycle of the octopus (Octopus vulgaris Cuvier) under culture conditions paralarvae rearing using Artemia and zoeae and first data on juvenile growth up to eight months of age Aquacult Int 12 481-487
Iglesias J Fuentes L Saacutenchez J Otero JJ Moxica C Lago MJ 2006 First feeding of Octopus vulgaris Cuvier 1797 paralarvae using Artemia Effect of prey size prey density and feeding frequency Aquaculture 261 817-822
Iglesias J Saacutenchez J Bersanob J Carrasco J Dhont J Fuentes L Linares F Muntildeoz J Okumura S Roo J van der Meeren T Vidal E Villanueva R 2007a Rearing of Octopus vulgaris paralarvae Present status bottlenecks and trends Aquaculture 266 1-15
Izquierdo M Chattou MA Roo J Socorro J Estefanell J Fernaacutendez-Palacios H 2008 Use of different commercial Artemia sp enrichments in Octopus vulgaris paralarval rearing at initial planktonic phase effects on growth survival and fatty acid composition of the paralarvae XIII International Symposium on Fish Nutrition and Feeding Florianopolis 1-5 Junio 2008 Brazil
Itami K Izawa Y Maeda S Nakay K 1963 Notes on the laboratory culture of the octopus culture Bull Jpn Soc Sci Fish 29 514-520 (en japoneacutes resumen en ingleacutes)
Iwata Y Munehara H Sakurai Y (2005) Dependence of paternity rates on alternative reproductive behaviours in the squid Loligo bleekeri Mar Ecol Prog Ser 298 219-228
Jones AG (2005) Gerud20 A computer program for the reconstruction of parental genotypes form half-sib progeny arrays with known or unknown parents Mol Ecol Notes 5 708-711
Kalinowski ST Taper ML Marshall TC (2007) Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment Mol Ecol 16 1099-1106
Karl SA (2008) The effect of multiple paternity on the genetically effective size of a population Mol Ecol 17 3973-3977
Kochert G 1978 Carbohydrate determination by the phenol-sulfuric acid method In Hellebust JA Craigie JS (Eds) Handbook of Phycology Methods
46
Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

Physiological and Biochemical methods Cambridge University Press London pp 95-97
Konovalov DA Manning C Henshaw MT (2004) KINGROUP a program for pedigree relationship reconstruction and kin group assignments using genetic markers Mol Ecol Notes 4 779-782
Lee PG 1994 Nutrition of cephalopods fuelling the system Mar Freshw Behav Physiol 25 35ndash51
Leporati SC Pecl GT Semmens JM 2007 Cephalopod hatchling growth the effects of initial size and seasonal temperatures Mar Biol 151 1375ndash1383
Lowry OH Resobrough HJ Farr AL Randall RJ 1951 Protein measurement with the Folin-Phenol reagent J Biol Chem 193 265-275
Mangold K 1983 Food feeding and growth in cephalopods Memoirs of the National Museum of Victoria 44 81-93
Mangold K Boletzky Sv 1973 New data on reproductive biology and growth of Octopus vulgaris Mar Biol 19 7ndash 12
Marliave J B 1981 Neustonic feeding in early larvae of Octopus dofleini (Wuumllker) The Veliger 23 350ndash351
Moxica C Linares F Otero JJ Iglesias J Saacutenchez FJ 2002 Cultivo intensivo de paralarvas de pulpo Octopus vulgaris Cuvier 1797 en tanques de 9 m3 Bol Inst Esp Oceanogr 18 31-36 (in Spanish English abstract)
Moxica C Fuentes L Hernaacutendez J Iglesias J Lago MJ Otero JJ and Saacutenchez FJ 2006 Efecto de Nannochloropsis sp en la supervivencia y crecimiento de paralarvas de pulpo Octopus vulgaris IX Foro dos Recursos Marintildeos e da Acuicultura das Riacuteas Galegas 10 y 11 de Octubre de 2006 O Grove Espantildea pp 255ndash269
Naef A 1928 Die Cephalopoden (Embriologie) Fauna e Flora del Golfo di Napoli Roma Bardi Monografiacutea (I2) 35 pp 1-364
National Research Council 1993 Dietary Requirements In Nutrient Requirements of Fish National Academy Press Washington DC pp 3-33
Naud MJ Hanlon RT Hall KC Shaw PW Havenhand JN (2004) Behavioural and genetic assessment of reproductive success in a spawning aggregation of the Australian giant cuttlefish Sepia apama Anim Behav 67 1043-1050
Navarro J Villanueva R 2000 Lipid and fatty acid composition of early stages of cephalopods an approach to their lipid requirements Aquaculture 183 161-177
Navarro J Villanueva R 2003 The fatty acid composition of Octopus vulgaris paralarvae reared with live and inert food deviation from their natural fatty acid profile Aquaculture 219 613-631
Nixon M 1987 Cephalopods diets In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Comparative Reviews London Academic Press pp 201ndash219
Okumura S Kurihara A Iwamoto A Takeuchi T 2005 Improved survival and growth in Octopus vulgaris paralarvae by feeding large type Artemia and Pacific sandeel Ammodytes personatus Improved survival and growth of common octopus paralarvae Aquaculture 244 147-157
Passarella KP Hopkins T L 1991 Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico Bull Mar Sci 49 683-659
Queller DC Goodnight KF (1989) Estimating relatedness using genetic markers Evolution 43 258-275
Sato N Murata N 1988 Membrane lipids In Packer L Glazer AN (Eds) Methods in Enzymology Vol 167 Academic Press New York pp 251-259
Semmens JM Pecl GT Villanueva R Jouffre D Sobrino I Wood JB Rigby PR 2004 Understanding octopus growth patterns variability and physiology Mar Fresh Res 55 367-377
Smale MJ Buchan PR 1981 Biology of Octopus vulgaris off the east coast of South Africa Mar Biol 65 1ndash12
47
Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48

Shaw PW Boyle PR (1997) Multiple paternity within the brood of single females of Loligo forbesi (Cephalopoda Loliginidae) demonstrated with microsatellite DNA markers Mar Ecol Prog Ser 160 279-282
Shaw PW Sauer WHH (2004) Multiple paternity and complex fertilization dynamics in the squid Loligo vulgaris reynaudii Mar Ecol Prog Ser 270 173-179
van Camp LM Donnellan SC Dyer AR Fairweather PG (2004) Multiple paternity in field- and captive-laid egg strands of Sepioteuthis australis (Cephalopoda Loliginidae) Mar Freshw Res 55 819-823
Vaz-Pires P Seixas P Barbosa A 2004 Aquaculture potential of the common octopus (Octopus vulgaris Cuvier 1797) a review Aquaculture 238 221-238
Vecchione M 1987 Juvenile ecology In Boyle PR (Ed) Cephalopod Life Cycles Vol 2 Academic Press London pp 61-84
Villanueva R 1994 Decapod crab zoeae as food for rearing cephalopod paralarvae Aquaculture 128 143ndash152
Villanueva R 1995 Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement Can J Fish Aquat Sci 52 2639-2650
Villanueva R Bustamante P 2006 Composition in essential and non-essential elements of early stages of cephalopods and dietary effects on the elemental profiles of Octopus vulgaris paralarvae Aquaculture 261 225-240
Villanueva R Nozais C Boletzky SV 1995 The planktonic life of octopuses Nature 377 107
Villanueva R Koueta N Riba J Boucaud-Camou E 2002 Growth and proteolytic activity of Octopus vulgaris paralarvae with different food rations during first feeding using Artemia nauplii and compound diets Aquaculture 205 269ndash286
Villanueva R Riba J Ruiacutez-Capillas C Gonzaacutelez AV Baeta M 2004 Amino acid composition of early stages of cephalopods and effect of amino acid dietary treatments on Octopus vulgaris paralarvae Aquaculture 242 455-478
Villanueva R Escudero JM Deulofeu R Bozzano A Casoliva C 2009 Vitamin A and E content in early stages of cephalopods and their dietary effects in Octopus vulgaris paralarvae Aquaculture 286 277-282
Walker D Power AJ Sweeney-Reeves MJ Avise JC (2006) Multiple paternity and female sperm usage along egg-case strings of the knobbed whelk Busycon carica (Mollusca Melongenidae) Mar Biol 151 53-61
Wells MJ 1978 Octopus Physiology and Behaviour of an Advanced Invertebrate Chapman and Hall London 417 pp
Wodinsky J 1978 Feeding behavior of broody female Octopus vulgaris Anim Behav 26 808-813
Young JZ 1971 The anatomy of the nervous system of Octopus vulgaris Claredon Press Oxford 690 pp
Young RE Harman RF 1988 lsquolsquoLarvarsquorsquo lsquolsquoparalarvarsquorsquo and lsquolsquosubadultrsquorsquo in cephalopod terminology Malacologia 29 (1) 201ndash 207
Zar J H 1999 Biostatistical analysis 4th Ed Prentice Hall Inc New Jersey 660 pp
48





























