Embed Size (px)
Citation preview

Euphytica 59 : 1 5 1-156, 1992 .© 1992 Kluwer Academic Publishers . Printed in the Netherlands .
Onion germplasm interacts with sulfur fertility for plant sulfur utilizationand bulb pungency
William M. RandleDepartment of Horticulture, University of Georgia, Athens, GA 30602, USA
Received 1 October 1991 ; accepted 31 January 1992
Key words: Allium cepa, onion, pyruvic acid, sulfur, germplasm, gxe
Summary
Sixty-two onion (Allium cepa L.) entries grown under high (4 .0 mequiv .liter -1 ) and low (0 .1 mequiv.liter -1 )S fertility were evaluated for foliar and bulb S utilization, and pungency . Significant differences weredetected among entries at both S fertility levels for foliar and bulb %S, and enzymatically developedpyruvate (pungency) in bulb tissue. Correlations between foliar %S and enzymatically developed pyruvateand between bulb %S and enzymatically developed pyruvate were poor suggesting that S is differentiallypartitioned into flavor and non-flavor compounds in the bulbs of onion germplasm . Significant S by entryinteractions were found for all variables tested indicating that selection for S utilization and bulb pungency inonions be conducted at S fertility levels indicative of targeted production areas .
Introduction
Consumer acceptance of onions (Allium cepa L.) isinfluenced by flavor quality. Preference for traitscontributing to flavor depends on cultural demandand intended use (Jones & Mann, 1963 ; Rabino-witch, 1988) . While many areas of the world preferpungent onions, demands for mild, sweet onionshas grown in the United States (Alysworth, 1991),Europe and Japan (Anon ., 1991) . Flavor potentialin onions is genetically determined, but is modifiedby the environment . Cultivars exist with a widerange of flavor intensities (Bajaj et al ., 1980; Free-man & Whenham, 1975 ; Lancaster et al ., 1988 ;Schwimmer & Weston, 1961) and empirical obser-vations suggested quantitative control (Jones &Mann, 1963). Year and location influenced flavorintensity in onions (Lancaster et al ., 1988 ; Bedford,1984; Platenius & Knott, 1941) . Temperature (Pla-tenius & Knott, 1941), water supply (Freeman &Mossedeghi, 1973) and sulfur (S) fertility (Freeman
& Mossedeghi, 1970) were specific factors shownto affect flavor intensity in onions . Sulfur fertility isof particular interest because the primary flavorcompounds in onions are sulfur based (Lancaster &Boland, 1990) .
Upon cellular disruption, the unique flavor ofonions arises from the hydrolysis of several flavorprecursors . The flavor precursors, collectivelyknown as S-alk(en)yl cysteine sulfoxides, producethiopropanal-S oxide (the lachrymator), pyruvateand ammonium upon contact with the enzyme allii-nase (Lancaster & Boland, 1990) . The volatile sul-fur compounds are unstable and undergo non-en-zymatic rearrangement to produce up to 80 identi-fied compounds (Boelens et al ., 1971) . Pyruvatehas been shown to be a good estimator of bulbpungency and flavor (Schwimmer & Weston, 1961 ;Schwimmer & Guadagni, 1962) .
Commercial onions are normally grown on soilswith sufficient S to provide the need for plantgrowth and flavor precursor accumulation (Lan-

1 52
caster & Boland, 1990). Sulfur fertility can, how-ever, influence bulb yield and plant dry weight .Freeman & Mossadeghi (1970) found that nutrientsolutions deficient in S or with 0.05 mequiv .liter - 'S reduced plant growth . Dry weigth.plant - ' wasgreatest when S ranged between 0 .1 and 1 .0 me-quiv.liter - ' . A 10% increase in bulb yield was dem-onstrated with the addition of 5 .5 kg.ha- ' S com-pared to 1 .8 kg . ha- ' S (Paterson, 1979) .
Increased demand for sweet, mild onions aroundthe world has been accompanied by increased in-terest in growing and breeding these onions . Pun-gency in onions can be minimized by growing culti-vars with the genetic potential for low pungency,on low S soils, with minimal S fertility . To facilitatethe breeding of sweet, low pungent onions for pro-duction on low S soils, a survey of variation for Suptake and pungency at low S fertility would beuseful. In addition, how S uptake and pungencyvary in response to high and low S fertility woulddemarcate genotype by S interactions in breedingonions of all flavor potentials . Limitations on selec-tion progress from GXE interactions in a givenenvironment has been demonstrated (Allard &Bradshaw, 1964 ; Fernandez, 1991 ; Hill, 1975) . Thisstudy was initiated to investigate how onion germ-plasm of broad genetic background responds tohigh and low S fertility for S uptake and utilizationand for bulb pungency .
Materials and methods
An evaluation of S uptake and flavor potential wasconducted on 35 U.S. Plant Introductions (PI)from the Northeastern Regional Plant Introduc-tion Station in Geneva, NY, on 25 accessions fromthe National Institute of Agricultural Science inJapan, and on 2 commercial cultivars . The germ-plasm collected represented a broad base of short-,and intermediate-types. Seed of each entry weresown in Fafard #3 artificial media (Fafard Corp .,Anderson, SC) in plastic trays on 15 December .Plants were grown in a greenhouse under naturalphotoperiods and light intensities with night andday temperatures set at 16 and 28 C, respectively .When the first true leaf emerged, seedlings were
fertilized with half strength Hoagland & Arnon(1950) solution . Seedlings were transplanted to15.2 cm diameter pots containing the same mediaon 7 February . The experimental design was a splitplot in three replications with five pots per repli-cation . Sulfur treatments were main plots and en-tries were subplots . Two nutrient regimes wereused to vary the S concentration . A full strengthHoagland's solution with 4 mequiv .liter -' S was thehigh treatment . The low treatment was a modifiedHoagland's solution with 0 .1 mequiv .liter- ' S. A Sconcentration of less than 0 .1 mequiv .liter - ' wasshown to reduce plant growth by up to 50% in thecultivar White Lisbon (Freeman & Mossadeghi,1970) . Magnesium chloride was substituted formagnesium sulfate in the modified solution . Sulfur-ic acid was the S source . The pH of the solutionswas adjusted to 6 .5 with NaOH. Plants were wa-tered twice weekly with the nutrient solutions, sup-plemented with tap water as needed, and the potsleached once a week . Sulfur concentration of thetap water was less than 0 .05 mequiv .liter - ' .
Plants were grown to maturity characterized by asoftening of the leaf sheath area and lodging of thefoliage. Growth in the greenhouse was typical offield-grown plants. Leaf samples were taken fromeach plant of each treatment on 20 March for Sanalysis of the foliage . The most recently, fullyexpanded leaf was selected . Bulbs from replica-tions were considered harvestable when foliagelodged on four of the five plants . Irrigation wasthen withheld and the plants were allowed to dryfor 7 days before the bulbs were harvested and thefoliage and roots severed . The earliest entry wasmature on 1 May and the latest entry matured 21June. The bulbs were cured in paper bags at ambi-ent greenhouse temperatures for 7 days, and storedat room temperature for 30 days before S and pun-gency analyses .
Total %S determinations were made using theprocedures of Jones & Isaac (1972) . Bulb and leaftissues were dried and ground to pass a 20 meshscreen . An iron accelerator and magnesium oxidewere added to a sample and muffled at 500 C for 1hour. Iron and tin accelerators were then addedand combusted on a Leco Sulfur Analyzer (LecoLaboratory Equipment Crop ., St. Joseph, MI) .

Combustion gasses were titrated and sulfur deter-minations made against a standard curve. Enzy-matically developed pyruvate was determinedfrom the modified methods of Schwimmer & West-on (1961) . Bulb tissue was blended with equal vol-umes of water for 3-5 minutes . Equal aliquots oftrichloroacetic acid and onion slurry were com-bined after 20 minutes, incubated for 1 hour, andthen filtered through cheese cloth . One ml of thefiltrate was added to 1 ml of 0.0125% 2,4-dinitro-phenylhydrazine in 2N HCL plus 1 ml water andincubated at 37 C for 10 minutes . Five ml NaOHwas then added before the absorbance was read at420 nm on a spectrophotometer . Pyruvate determi-nations were made against a sodium pyruvate stan-dard curve. Bulb tissue was dried at 70 C for 48hours in a forced air oven for % dry weight deter-minations .
Data were tested by analysis of variance usingthe ANOVA procedures of SAS (Cary, NC) . Sig-nificantly different means were separated by leastsignificant difference at the 5 % level . A correlationmatrix for entries within S fertility and a Spearmanrank correlation for entries among S fertility werecalculated for all variables tested .
Results and discussion
Analysis of variance established highly significantdifferences (p = 0.01) among S treatments . Thehigh S treatment resulted in higher concentrationsof foliar and bulb sulfur, enzymatically producedpyruvate and dry weight percentage . Higher per-centages of sulfur were present in the sampled fo-liar tissue compared to the bulb tissue . Averagefoliar %S was 0 .91 and 0 .59 at the high and low Streatments, respectively . Bulb % S averaged 0 .60 atthe high S treatment and 0 .33 at the low S treat-ment . This range of values is similar to that previ-ously reported for bulb % S (Christensen et al .,1968). Sulfur compounds in onion are thought to berecycled during growth and development with leafblades supplying developing bulb scale tissue (Lan-caster et al ., 1986) . In addition, S compounds differwith age of the blade and bulb (Platenius, 1935 ;Lancaster et al ., 1984) and location of the scale
153
within the bulb (Freeman, 1975) . Pyruvate concen-tration averaged 6.8 gmol .g-1 f.w . at the high Streatment and 3 .7 gmol .g-1 f.w. at the low S treat-ment . Only slight differences were found for dryweight at the high (10 .9%) and low (10.7%) Streatments. Freeman & Mossadeghi (1970) foundthat % dry weight of the onion cultivar WhitheLisbon, decreased from 8 .0 to 7 .3 when grown in0.1 to 3 mequiv.liter -1 S nutrient solutions, al-though it was not know if the difference was statis-tically significant .
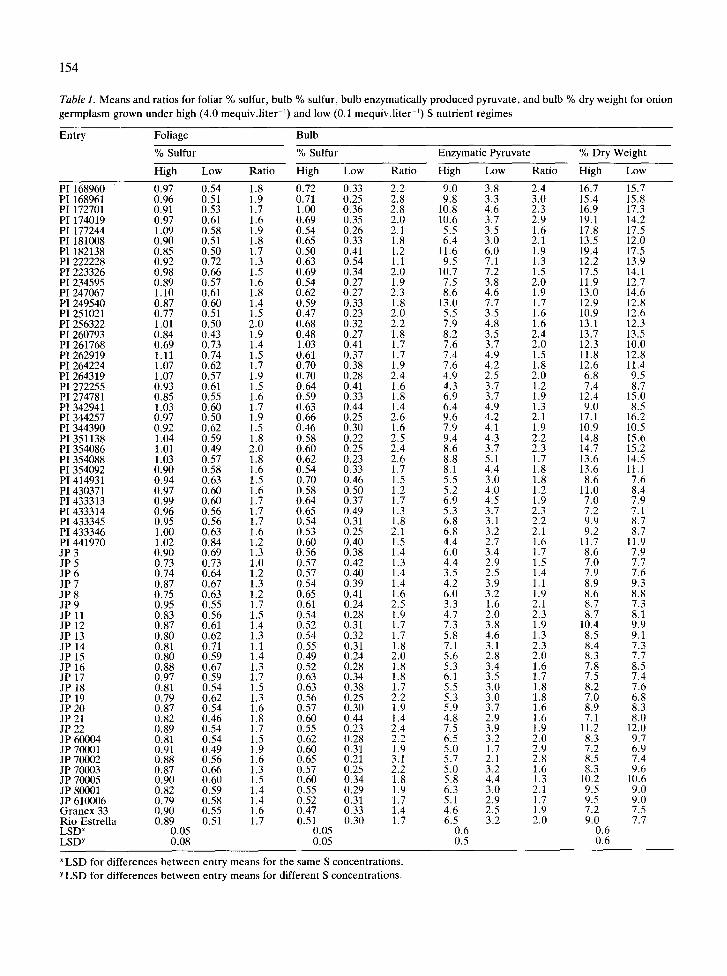
Highly significant differences (p = 0 .01) werefound among entries for all variables tested . Foliar%S ranged from 1.11 (PI 262919) to 0 .69 (PI261768) at the high S treatment and from 0 .84 (PI441970) to 0 .43 (PI 260793) at the low S treatment(Table 1) . Bulb %S ranged from 1 .03 (PI 261768) to0.46 (PI 344390) at the high S treatment and from0.54 (PI 222228) to 0.21 (JP 70002) at the low Streatment. Phenotypic variability for percent Swhen plants are grown under different S fertilitylevels has not been previously described . The cor-relation coefficients between foliar and bulb %Swere poor, but highly significant at both S levels(Table 2) .
Enzymatically produced pyruvate ranged from13 .0 gmol .g-1 f.w. (PI 249540) to 3 .3 gmol .g-1 f.w .(JP 9) at the high S treatment and from 7.7 gmol .g-1 f.w. (PI 249540) to 1 .6gmol.g-1 f.w. (JP 9) atthe low S treatment . Pyruvate concentrations fromthe high S treatment are representative of pheno-typic differences previously reported for field-grown fresh market onions, although values below5.3gmol.g-1 f.w. were rare (Bajaj et al., 1980 ;Freeman & Whenham, 1975 ; Lancaster et al .,1988; Schwimmer & Guadagni, 1962 ; Schwimmer& Weston, 1961). Phenotypic variability for en-zymatic pyruvate at low S fertility has not beenpreviously described. However, a single cultivargrown in sand culture at 0 .1 mequiv .liter -1 S had aenzymatic pyruvate concentration of 0 .92 gmol .g-1f.w. (Freeman & Mossadeghi, 1970) . Correlationsbetween bulb %S and enzymatic pyruvate at highand low S fertility were poor, but highly significant(Table 2) suggesting that onion germplasm parti-tions S into flavor and non-flavor compounds dif-ferently . As an example, PIs 264224 and 264319
154
Table 1 . Means and ratios for foliar % sulfur, bulb % sulfur, bulb enzymatically produced pyruvate, and bulb % dry weight for oniongermplasm grown under high (4 .0 mequiv .liter -1) and low (0.1 mequiv .liter -1 ) S nutrient regimes
"LSD for differences between entry means for the same S concentrations .Y LSD for differences between entry means for different S concentrations .
Entry Foliage Bulb
% Sulfur Enzymatic Pyruvate % Dry Weight% Sulfur
High Low Ratio High Low Ratio High LowHigh Low Ratio
PI 168960 0.97 0.54 1 .8 0 .72 0 .33 2 .2 9 .0 3 .8 2 .4 16 .7 15 .7PI 168961 0.96 0 .51 1 .9 0 .71 0 .25 2 .8 9 .8 3 .3 3 .0 15 .4 15 .8PI 172701 0 .91 0 .53 1 .7 1 .00 0 .36 2 .8 10 .8 4 .6 2 .3 16 .9 17 .3PI 174019 0.97 0 .61 1 .6 0 .69 0 .35 2 .0 10 .6 3 .7 2 .9 19 .1 14 .2PI 177244 1.09 0.58 1 .9 0 .54 0 .26 2 .1 5 .5 3 .5 1 .6 17 .8 17 .5PI 181008 0.90 0 .51 1 .8 0 .65 0 .33 1 .8 6 .4 3 .0 2 .1 13 .5 12 .0PI 182138 0.85 0.50 1 .7 0 .50 0 .41 1 .2 11 .6 6 .0 1 .9 19 .4 17 .5PI 222228 0.92 0.72 1 .3 0 .63 0 .54 1.1 9 .5 7 .1 1 .3 12 .2 13 .9PI 223326 0.98 0.66 1 .5 0 .69 0 .34 2 .0 10 .7 7 .2 1 .5 17 .5 14 .1PI 234595 0.89 0.57 1 .6 0 .54 0 .27 1 .9 7 .5 3 .8 2 .0 11 .9 12 .7PI 247067 1.10 0 .61 1 .8 0 .62 0 .27 2 .3 8 .6 4 .6 1 .9 13 .0 14 .6PI 249540 0.87 0.60 1 .4 0 .59 0 .33 1 .8 13 .0 7 .7 1 .7 12 .9 12 .8PI 251021 0.77 0 .51 1 .5 0 .47 0 .23 2 .0 5 .5 3 .5 1 .6 10 .9 12 .6PI 256322 1 .01 0 .50 2 .0 0 .68 0 .32 2 .2 7 .9 4 .8 1 .6 13 .1 12 .3PI 260793 0.84 0 .43 1 .9 0 .48 0 .27 1 .8 8 .2 3 .5 2 .4 13 .7 13 .5PI 261768 0.69 0.73 1 .4 1 .03 0 .41 1 .7 7 .6 3 .7 2 .0 12 .3 10 .0PI 262919 1.11 0.74 1 .5 0 .61 0 .37 1 .7 7 .4 4 .9 1 .5 11 .8 12 .8PI 264224 1.07 0.62 1 .7 0 .70 0 .38 1 .9 7 .6 4 .2 1 .8 12 .6 11 .4PI 264319 1.07 0.57 1 .9 0 .70 0 .28 2 .4 4 .9 2 .5 2 .0 6 .8 9 .5PI 272255 0.93 0 .61 1 .5 0 .64 0 .41 1 .6 4 .3 3 .7 1 .2 7 .4 8 .7PI 274781 0.85 0 .55 1 .6 0 .59 0 .33 1 .8 6 .9 3 .7 1 .9 12 .4 15 .0PI 342941 1.03 0.60 1 .7 0 .63 0 .44 1 .4 6 .4 4 .9 1 .3 9 .0 8 .5PI 344257 0.97 0.50 1 .9 0 .66 0 .25 2 .6 9 .6 4 .2 2 .1 17 .1 16 .2PI 344390 0.92 0.62 1 .5 0 .46 0 .30 1 .6 7 .9 4 .1 1 .9 10 .9 10 .5PI 351138 1.04 0.59 1 .8 0 .58 0 .22 2 .5 9 .4 4 .3 2 .2 14 .8 15 .6PI 354086 1 .01 0 .49 2 .0 0 .60 0 .25 2 .4 8 .6 3 .7 2 .3 14 .7 15 .2PI 354088 1.03 0.57 1 .8 0 .62 0 .23 2 .6 8 .8 5 .1 1 .7 13 .6 14 .5PI 354092 0.90 0.58 1 .6 0 .54 0 .33 1 .7 8 .1 4 .4 1 .8 13 .6 11 .1PI 414931 0.94 0.63 1 .5 0 .70 0.46 1 .5 5 .5 3 .0 1 .8 8 .6 7 .6PI 430371 0.97 0.60 1 .6 0 .58 0 .50 1 .2 5 .2 4 .0 1 .2 11 .0 8 .4PI 433313 0.99 0.60 1 .7 0 .64 0 .37 1 .7 6 .9 4 .5 1 .9 7 .0 7 .9PI 433314 0.96 0.56 1 .7 0 .65 0 .49 1 .3 5 .3 3 .7 2 .3 7 .2 7 .1PI 433345 0.95 0.56 1 .7 0 .54 0 .31 1 .8 6 .8 3 .1 2 .2 9 .9 8 .7PI 433346 1 .00 0.63 1 .6 0 .53 0 .25 2 .1 6 .8 3 .2 2 .1 9 .2 8 .7PI 441970 1.02 0.84 1 .2 0 .60 0.40 1 .5 4 .4 2 .7 1 .6 11 .7 11 .9JP 3 0.90 0.69 1 .3 0 .56 0 .38 1 .4 6 .0 3 .4 1 .7 8 .6 7 .9JP 5 0.73 0 .73 1 .0 0 .57 0 .42 1 .3 4 .4 2 .9 1 .5 7 .0 7 .7JP 6 0.74 0.64 1 .2 0 .57 0 .40 1 .4 3 .5 2 .5 1 .4 7 .9 7 .6JP 7 0.87 0.67 1 .3 0 .54 0 .39 1 .4 4 .2 3 .9 1 .1 8 .9 9 .3JP 8 0.75 0.63 1 .2 0 .65 0 .41 1 .6 6 .0 3 .2 1 .9 8 .6 8 .8JP 9 0 .95 0 .55 1 .7 0 .61 0 .24 2 .5 3 .3 1 .6 2 .1 8 .7 7 .3JP 11 0 .83 0 .56 1 .5 0 .54 0 .28 1 .9 4 .7 2 .0 2 .3 8 .7 8 .1JP 12 0.87 0 .61 1 .4 0 .52 0 .31 1 .7 7 .3 3 .8 1 .9 10 .4 9 .9JP 13 0.80 0.62 1 .3 0 .54 0 .32 1 .7 5 .8 4 .6 1 .3 8 .5 9 .1JP 14 0 .81 0.71 1 .1 0 .55 0 .31 1 .8 7 .1 3 .1 2 .3 8 .4 7 .3JP 15 0 .80 0.59 1 .4 0 .49 0 .24 2 .0 5 .6 2 .8 2 .0 8 .3 7 .7JP 16 0 .88 0.67 1 .3 0 .52 0 .28 1 .8 5 .3 3 .4 1 .6 7 .8 8 .5JP 17 0 .97 0 .59 1 .7 0 .63 0 .34 1 .8 6 .1 3 .5 1 .7 7 .5 7 .4JP 18 0 .81 0.54 1 .5 0 .63 0 .38 1 .7 5 .5 3 .0 1 .8 8 .2 7 .6JP 19 0 .79 0.62 1 .3 0 .56 0 .25 2 .2 5 .3 3 .0 1 .8 7 .0 6 .8JP 20 0 .87 0.54 1 .6 0 .57 0 .30 1 .9 5 .9 3 .7 1 .6 8 .9 8 .3JP 21 0.82 0.46 1 .8 0 .60 0 .44 1 .4 4 .8 2 .9 1 .6 7 .1 8 .0JP 22 0.89 0 .54 1 .7 0 .55 0 .23 2 .4 7 .5 3 .9 1 .9 11 .2 12 .0JP 60004 0 .81 0.54 1 .5 0 .62 0 .28 2 .2 6 .5 3 .2 2 .0 8 .3 9 .7JP 70001 0 .91 0.49 1 .9 0 .60 0 .31 1 .9 5 .0 1 .7 2 .9 7 .2 6 .9JP 70002 0.88 0 .56 1 .6 0 .65 0 .21 3 .1 5 .7 2 .1 2 .8 8 .5 7 .4JP 70003 0.87 0 .66 1 .3 0 .57 0 .25 2 .2 5 .0 3 .2 1 .6 8 .3 9 .6JP 70005 0.90 0.60 1 .5 0 .60 0 .34 1 .8 5 .8 4 .4 1 .3 10 .2 10 .6JP 80001 0 .82 0.59 1 .4 0 .55 0 .29 1 .9 6 .3 3 .0 2 .1 9 .5 9 .0JP 610006 0.79 0.58 1 .4 0 .52 0 .31 1 .7 5 .1 2 .9 1 .7 9 .5 9 .0Granex 33 0.90 0 .55 1 .6 0 .47 0 .33 1 .4 4 .6 2 .5 1 .9 7 .2 7 .5Rio Estrella 0.89 0.51 1 .7 0 .51 0 .30 1 .7 6 .5 3 .2 2 .0 9.0 7 .7LSDx 0.05 0 .05 0 .6 0 .6LSDY 0.08 0 .05 0 .5 0 .6

had identical foliar and bulb %S values at high Sfertility, but differed significantly for enzymaticpyruvate concentration (Table 1) . Whether the dif-ferential accumulation of S into non-flavor com-pounds is in flavor precursor intermediates, such asthe y-glutamyl peptides, other types of sulfur com-pounds (Lancaster et al ., 1989) is yet to be deter-mined. Correlations between foliar %S and en-zymatic pyruvate were also poor and non-signif-icant at the low S level, but significant at the high Slevel. Percent dry weight ranged from 19 .4 (PI182138) to 6.8 (PI 264319) at the high S level andfrom 17.5 (PI's 177244 and 182138) to 6.8 (JP 19) atthe low S level . These concentrations are repre-sentative of phenotypic differences previously re-ported for dry matter accumulation in field-grownonions (Darbyshire & Henry, 1979 ; Darbyshire &Steer, 1988) . Dry matter correlated poorly withfoliar and bulb % S at high and low sulfur fertilityand pyruvate at low S fertility, yet was highly signif-icant (Table 2) . The correlation, however, betweendry matter and pyruvate at high S fertility wasmoderate to good and highly significant .
Analysis of variance also established highly sig-nificant differences (p = 0 .01) for the interactionbetween S and entries for all variables tested . Ra-tios for foliar %S between high and low S fertility
Table 2 . Correlation coefficients (r) between foliar and bulb %sulfur, bulb enzymatically produced pyruvate and bulb % dryweight across 62 onion entries grown under high (4 .0 mequiv .li-ter - ') and low (0 .1 mequiv .liter -1 ) S nutrition
"S•* * Not significant or significant at the 1% level, respectively .I Foliage sampled a the onset of bulbing .Y Mature, cured bulbs were sampled .
1 55
were above 1 .0, except for JP 5 which was appar-ently unaffected by S fertility (Table 1) . Forty-five
entries has ratios above 1 .5 suggesting that mostonion germplasm is meaningfully influenced by Snutrition for foliar %S . Three entries were rela-tively unaffected by S fertility (PI 441970, JP 6, JP8), having ratios of 1 .2 or below . On the otherhand, several entries were greatly affected by Sfertility for foliar %S (PI 168960, PI 177244, PI256322, PI 260793, PI 264319, PI 344257, PI354086, JP 70001), having ratios of 1 .9 or above .The Spearman rank correlation between high andlow S fertility for foliar %S was very poor (r, =0.11) and nonsignificant suggesting that the onionentries differentially responded to the level of Sfertility .
Ratios between high and low S fertility for bulb%S were all above 1 .0 (Table 1) . The distributionand magnitude of ratios for bulb %S, however,were much greater than the ratios for foliar %S .Twenty-two entries had bulb %S ratios above 2 .0,whereas only 11 entries had ratios below 1 .5. Sever-al entries were greatly affected by S fertility (PI168961, PI 172701, JP 70002), having ratios above2.8 . Entries which were relatively unaffected by Sfertility were PI 182138, PI 222228, and PI 430371,each having ratios below 1 .3 . The Spearman rankcorrelation for bulb %S between entries grown athigh and low S fertility was poor (r s = 0 .33), albeithighly significant (p = 0 .01) .
The distribution and magnitude of the ratios forenzymatically developed pyruvate at high and lowS fertility were similar to those for bulb % S (Table1). However, the Spearman rank correlation be-tween high and low S fertility for enzymaticallydeveloped pyruvate was moderate (r s = 0.49) andhighly significant (p = 0 .01), suggesting that en-zymatically produced pyruvate was more stableamong S fertility levels than either foliar or bulb%S. Entries which were greatly affected by S fertil-ity were PI 168961, PI 174019, JP 70001 and JP70002, each having ratios above 2 .8. Several entrieswere relatively unaffected by S fertility for enzy-matically produced pyruvate (PI 272255, PI430371, JP 7), having ratios below 1 .3 .
Phenotypic variation for foliar and bulb % S, andpungency was found in onion germplasm grown at
Variable 1 2 3 4
1 Foliar% Sx - 0.33**High S
0.27** 0 .36**2 Bulb% SY - 0.30** 0 .29**3 Pyruvate 0.76**4% DM
1 Foliar% S - 0.31**Low S
0.13" 1 -0.19**2 Bulb% S - 0.29** -0 .18*3 Pyruvate 0.49**4% DM

156
high and low S fertility . A significant interactionbetween onion germplasm and S fertility has beendemonstrated for foliar and bulb %S and enzymat-ically developed pyruvate (pungency) . Therefore,when selecting onions for S utilization or pungen-cy, it is important that it be done at S fertility levelsindicative of tartgeted production areas .
References
Allard, R.W. & A.D. Bradshaw, 1964. Implications of geno-type-environmental interactions in plant breeding . Crop Sci .4 : 503-508 .
Anonymous, 1991 . Japanese to sample Walla sweet during sea-son. Onion World 7(5) : 16-17 .
Aylsworth, J.D ., 1991. Sweet onion showdown. Am. Veg .Grower 39(9) : 32-34 .
Bajaj, K.L ., G. Kaur, J . Singh & S .P .S . Gill, 1980 . Chemicalevaluation of some important varieties of onion (Allium cepaL .) . Qual . Plant . Plant Foods Hum . Nutr . 30 : 117-122 .
Bedford, L.V., 1984 . Dry matter and pungency tests on Britishgrown onions . J . Natl . Inst . Agric . Bot . 16 : 58-61 .
Boelens, M ., P .J . de Valois, H .J . Wobben & A . van der Gen,1971 . Volatile flavour compounds from onion . J . Agric . FoodChem. 19 : 984-991 .
Christensen, R.E ., R.M. Beckman & J .J . Birdsall, 1968 . Somemineral elements of commercial spices and herbs as deter-mined by direct reading emission spectroscopy . J . Assoc . Off .Anal. Chem . 51 :1003-1008 .
Darbyshire, B . & R .J . Henry, 1979 . The association of fructanswith high percentage dry weight in onion cultivars suitable fordehydrating . J . Sci . Food Agr . 30 : 1035-1038 .
Darbyshire, B . & B .T. Steer, 1988 . Carbohydrate biochemistry .p . 1-16 . In : H . D . Rabinowitch & J .L. Brewster (Eds) . Vol . 3 :Onions and allied crops. CRC Press, Boca Raton, Fla .
Fernandez, G.C.J ., 1991 . Analysis of genotype x environmentinteraction by stability estimates . HortSci . 26 : 947-950 .
Freeman, G.G ., 1975 . Distribution of flavour components inonion (Allium cepa L.), Leek (Allium porrum) and garlic(Allium sativum) . J . Sci . Food Agric . 26 : 471-481 .
Freeman, G .G. & N . Mossadeghi, 1970 . Effect of sulphatenutrition on flavour components of onion (Allium cepa) . J .Sci . Food Agr . 21 : 610-615 .
Freeman, G . G . & N . Mossadeghi, 1973 . Studies on the relation-ship between water regime and flavour strength in water cress(Rorippa nasturium-aquaticum [L] Hayek,) cabbage (Brassi-
ca oleracea var . capitata) and onion (Allium cepa) . J . Hort .Sci . 48 : 365-378 .
Freeman, G.G. & R .J . Whenham, 1975. A rapid spectrophoto-metric method of determination of thiopropanal S-oxide (la-chrymator) in onion (Allium cepa L .) and its significance inflavor studies . J . Sci . Food Agric . 26 : 1529-1543 .
Hill, J ., 1975 . Genotype-environment interaction - a challengeto plant breeding . J . Agric . Sci . 85 : 477-499.
Hoagland, D .R. & D . 1 . Arnon, 1950 . The water culture methodfor growing plants without s oil . Calif. Agr . Expt . Sta . Circ .34 7 .
Jones, J .B . & R .A. Isaac, 1972 . Determination of sulfur in plantmaterial using a Leco Sulfur Analyzer . J . Agric. Food Chem .20:1292-1294 .
Jones, H . A . & L .K . Mann, 1963 . Onions and their allies . Bota-ny, cultivation, and utilization. Interscience Publ ., NY.
Lancaster, J .E. & M .J . Boland, 1990 . Flavor biochemistry . p .33-72 . In : H.D. Rabinowitch & J.L. Brewster (Eds) . Vol . 3 :Onions and allied crops. CRC Press, Boca Raton, Fla .
Lancaster, J .E ., B .J . McCallion & M .L. Shaw, 1984 . The levelsof S-alk(en)yl-L-cysteine sulphoxides during the growth ofthe onion (Allium cepa L .) . J . Sci . Food Agric . 35 : 415-420 .
Lancaster, J .E ., B .J . McCallion & M .L. Shaw, 1986. The dy-namics of the flavour precursors, the S-alk(en)yl-L-cysteinesulphoxides, during leaf blade and scale development in theonion (Allium cepa) . Physiol . Plant 66: 293-297 .
Lancaster, J .E ., P .F . Reay, J .D . Mann, W .D. Bennett & J .R .Sedcole, 1988 . Quality in New Zealand-grown onion bulbs - asurvey of chemical and physical characteristics. New ZealandJ Expt . Agric . 16 : 279-285 .
Lancaster, J .E ., P.H .S . Reynolds, M .L. Shaw, E.M. Dom-misse & J. Munro, 1989 . Intracellular localization of thebiosynthetic pathway to flavour precursors in onion . Phy-tochem . 28: 461-464 .
Paterson, D .R ., 1979 . Sulfur fertilization effects on onion yieldand pungency . Tex . Agr . Expt . Sta. Prog . Rep . 355 1 .
Platenius, H ., 1935 . A method for estimating the volatile sulfurcontent and pungency of onions . J . Agric . Res . 51 : 847-853 .
Platenius, H . & J .E . Knott, 1941 . Factors affecting onion pun-gency . J . Agric . Res . 62 : 371-379 .
Rabinowitch, H.D., 1988 . Genetics and breeding: state of theart or too slow but not too late . p . 57-69 . In : Eucarpia 4thAllium Symposium . Wellsbourne, Warwick, UK .
Schwimmer, S . & D.G. Guadagni, 1962 . Relation betweenolfactory threshold concentration and pyruvic acid content ofonion juice . J . Food Sci . 27 : 94-97 .
Schwimmer, S . & W.J . Weston, 1961 . Enzymatic developmentof pyruvic acid in onion as a measure of pungency . J . Agric .Food Chem . 9 : 301-304 .





























