Embed Size (px)
Citation preview

Oscillations in Glycolysis, Cellular Respiration and Communication
BY ARNOLD BOITEUX AND BENNO HESS * Max-Planck-lnstitut fiir Ernahrungsphysiologie, Dortmund
Received 3rd October, 1974
In an attempt to understand oscillatory phenomena in chemical, subcellular and cellular systems, the mechanisms of oscillation as well as the general dynamics of glycolysis, cellular respiration and cyclic-AMP-controlled oscillation in the slime mould Dictyostelirtm discoideum have been studied. The enzymic sources of the oscillations have been identified with glycolysis as well as for the slime mould, and appropriate mathematical models have been developed. The primary reaction mecha- nism of mitochondria1 oscillations is not yet known in detail. However, the studies show unequivo- cally that the conditions for the generation of oscillations can be summarized for all cases in four common statements :
(i) The " primary source " of the oscillations has a nonlinear kinetic characteristic, (ii) The kinetic structure involves simple and multiple types of feed-back interactions. (iii) The system nioves on a limit cycle. (iv) The system operates far from equilibrium and is thermodynamically open.
Recently oscillations in biochemical and biological systems have been observed (for summary see ref. (1)-(4)), recognized as results of thermodynamic conditions, and defined as " dissipative structures " by Glansdorff and Prig~gine.~ Indeed, theory and experiment demonstrate a number of dynamic states in nature, which evolve in open systems operating with a critical degree of non-linearity distant from equilibrium. Three types of behaviour have been observed :
(i) The maintenance of multiple steady states with transitions from one to another.
(ii) The maintenance of rotation on a limit cycle around an unstable singular point. (iii) The maintenance of sustained oscillations coupled to diffusion, resulting in
chemical waves. Recent analyses of biochemical and cellular oscillators provide a remarkable
insight into the molecular source of non-linearity although detailed mechanisms might not be at hand in every case. In some cases, promising progress has been made in biochemical studies of the dynamics observed and of the structures involved as well as in the establishment of quantitative mathematical models. This paper summarizes results obtained in our laboratory in the field of bioenergetic processes and of intercellular communication, in partial collaboration with Dr. A. Goldbeter of the Weizmann Institute of Science, Rehovot, and Dr. G. Gerisch of the Friedrich- Miescher-Laboratorium der Max-Planck-Gesellschaft, Tiibingen.
O S C I L L A T I O N S IN GLYCOLYSIS
At the present time, oscillating glycolysis is the best understood oscillating biochemical system. Since the original observation of NADH-cycles in yeast cells
202
Publ
ishe
d on
01
Janu
ary
1974
. Dow
nloa
ded
by U
nive
rsita
t Pol
itècn
ica
de V
alèn
cia
on 2
6/10
/201
4 08
:07:
19.
View Article Online / Journal Homepage / Table of Contents for this issue
A . BOITEUX AND B . FTESS 203
initialed on the transition from aerobiosis to aiiaerobiosis (for review sec ref. (1 ,)) this phenomenon has received considerable attention. It has been observed in intact cells, in cell ghosts, in cell extracts, and by a variety of analytical methods it can be reduced to its molecular mechanism and satisfactorily simulated by digital computer techniques.
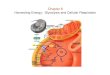
In a suspension of intact cells following the addition of glucose under anaerobic conditions, a number of cyclic metabolic changes can be observed and directly demonstrated by recording the fluorescence of NADH located inside the cells and part of the metabolic process involved in the cyclic phenomena. A typical experiment is shown in the fluorometric record of fig. 1 , which was obtained after a single addition of 100 pmol glucose leading to a series of cyclic changes of NADH-fluorescence,
FIG. 1.-Oscillations in yeast cells grown aerobically for 32 h. The arrow indicates a single addition of glucose. (From ref. (7)).
By means of an injection technique allowing the continuous addition of glucose at a steady rate, undamped oscillations of NADH can be maintained as demonstrated in fig. 2. Whenever suitable glycolytic substrates-either glucose, fructose, sucrose, maltose or trehalose-are injected into a suspension of yeast cells, the cells continue to glycolyze their substrates in an oscillatory manner as long as the injection rate is maintained constant within an upper and lower limit.6* The period of the cycles varies with the experimental conditions between 15 and 75 s.'
FIG. 2.-Qscillatioas in yeast ceIls grown anaerobically for 32 h. Glucose is injected at a constant rate. (From ref. (7)).
Publ
ishe
d on
01
Janu
ary
1974
. Dow
nloa
ded
by U
nive
rsita
t Pol
itècn
ica
de V
alèn
cia
on 2
6/10
/201
4 08
:07:
19.
View Article Online
204 B I O C H E M I C A L OSCILLATIONS I N T I M E AND S P A C E
By a short treatment with toluene the cellular membrane of yeast can be made permeable to low molecular weight substances, whereas the enzymic equipment of the cells is retained within the cellular volume.8 This technique allows a study of the functjon of the low molecular weight substances in the mechanism of glycolytic oscillation. It has been shown that as soon as a lower limit of NAD and adenine nucleotide concentrations is maintained, substrate addition leads to glycolytic oscillations with a period between 30 s and a few minutes, comparable to what is observed in intact cells. To a first approximation the NAD concentration determines the amplitude, while the phosphate/nucleotide system strongly influences period and waveform of the oscillation of NADH.
The complexity of the cellular system can be reduced further without losing essential properties. A cell-free extract obtained by treatment of cells with ultrasonic waves and high speed centrifugation still exhibits self-sustained oscillations when supplied with glucose or fructose at a limiting rate, using the injection technique or a controlled enzymic generation of glucose from disaccharides (see ref. (1) and (7)). The period of the oscillations of NAD fluorescence in the cell-free extract depends primarily on the rate of substrate input and varies between several minutes and approximately 1 h.6 The prolongation of the period observed in cell-free extracts as compared with those observed in the intact suspended cells is mainly due to the dilution of the glycolytic enzymes during preparation of the extract.
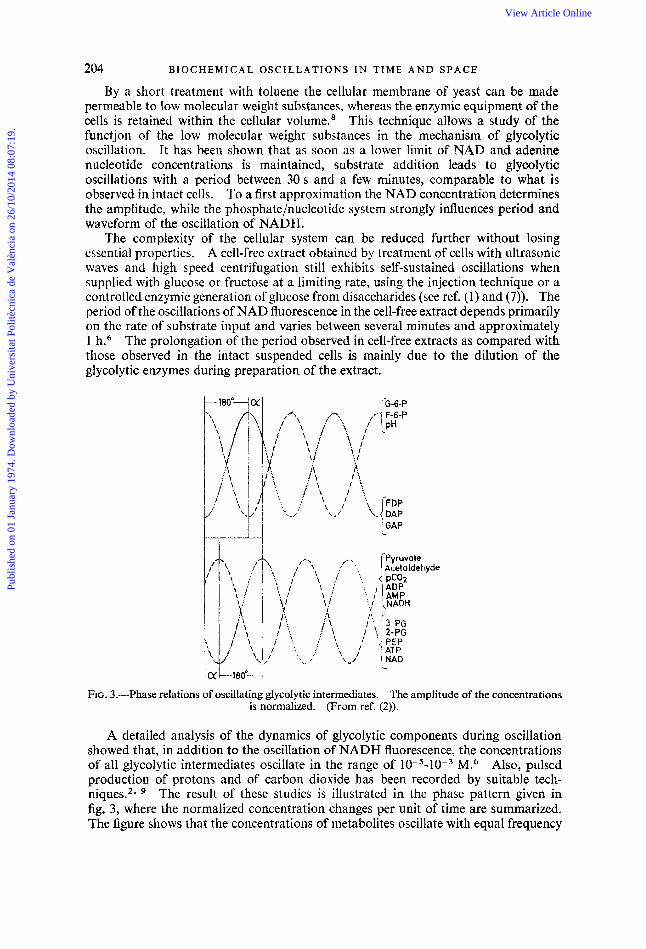
FIG. 3.-Phase relations of oscillating glycolytic intermediates. The amplitude of the concentrations is normalized. (From ref. (2)).
A detailed analysis of the dynamics of glycolytic components during oscillation showed that, in addition to the oscillation of NADH fluorescence, the concentrations of all glycolytic intermediates oscillate in the range of 10-5-10-3 M.6 Also, pulsed production of protons and of carbon dioxide has been recorded by suitable tech- niques.2* The result of these studies is illustrated in the phase pattern given in fig. 3, where the normalized concentration changes per unit of time are summarized. The figure shows that the concentrations of metabolites oscillate with equal frequency
Publ
ishe
d on
01
Janu
ary
1974
. Dow
nloa
ded
by U
nive
rsita
t Pol
itècn
ica
de V
alèn
cia
on 2
6/10
/201
4 08
:07:
19.
View Article Online
A . BOITEUX A N D B. HESS 205
but different phase angles, relative to each other. According to their time dependence they can be classified into two groups in which maxima and minima of the concentra- tions coincide in time. The two groups differ by the phase angle a, which depends on the experimental conditions.6
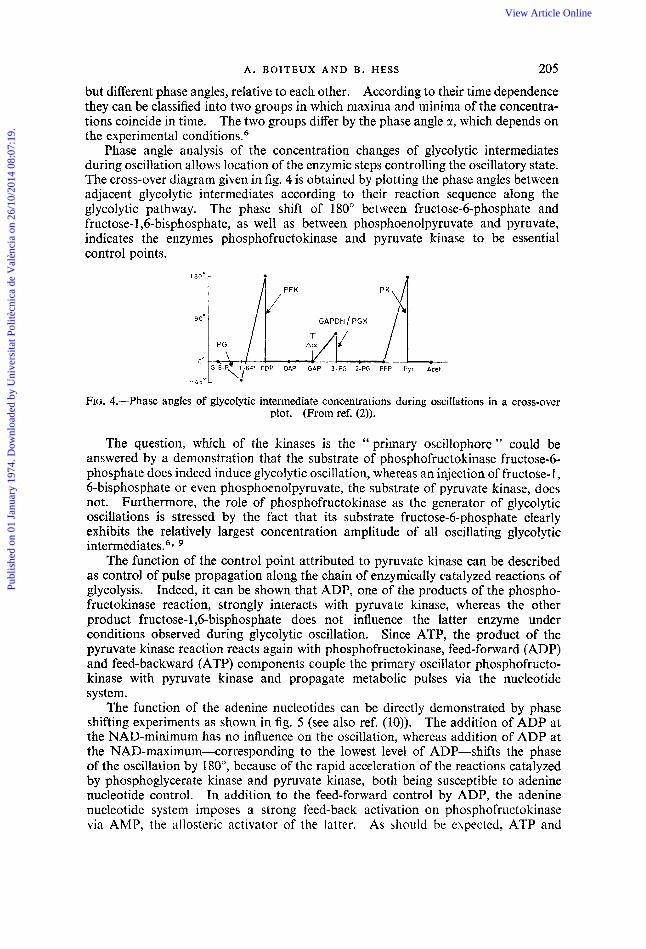
Phase angle analysis of the concentration changes of glycolytic intermediates during oscillation allows location of the enzymic steps controlling the oscillatory state. The cross-over diagram given in fig. 4 is obtained by plotting the phase angles between adjacent glycolytic intermediates according to their reaction sequence along the glycolytic pathway. The phase shift of 180” between fructose-6-phosphate and fructose- 1,6-bisphosphate, as well as between phosphoenolpyruvate and pyruvate, indicates the enzymes phosphofructokinase and pyruvate kiiiase to be essential control points.
FIG. 4.-Phase angles of glycolytic intermediate concentrations during oscillations in a cross-over plot. (From ref. (2)).
The question, which of the kinases is the ‘‘primary oscillophore” could be answered by a demonstration that the substrate of phosphofructokinase fructose-6- phosphate does indeed induce glycolytic oscillation, whereas an injection of fructose-I, 6-bisphosphate or even phosphoenolpyruvate, the substrate of pyruvate kinase, does not. Furthermore, the role of phosphofructokinase as the generator of glycolytic oscillations is stressed by the fact that its substrate fructose-6-phosphate clearly exhibits the relatively largest concentration amplitude of all oscillating glycolytic intermediates.6*
The function of the control point attributed to pyruvate kinase can be described as control of pulse propagation along the chain of enzymically catalyzed reactions of glycolysis. Indeed, it can be shown that ADP, one of the products of the phospho- fructokinase reaction, strongly interacts with pyruvate kinase, whereas the other product fructose-l,6-bisphosphate does not influence the latter enzyme under conditions observed during glycolytic oscillation. Since ATP, the product of the pyruvate kinase reaction reacts again with phosphofructokinase, feed-forward (ADP) and feed-backward (ATP) components couple the primary oscillator phosphofructo- kinase with pyruvate kinase and propagate metabolic pulses via the nucleotide system.
The function of the adenine nucleotides can be directly demonstrated by phase shifting experiments as shown in fig. 5 (see also ref. (10)). The addition of ADP at the NAD-minimum has no influence on the oscillation, whereas addition of ADP at the NAD-maximum-corresponding to the lowest level of ADP-shifts the phase of the oscillation by 180”, because of the rapid acceleration of the reactions catalyzed by phosphoglycerate kinase and pyruvate kinase, both being susceptible to adenine nucleotide control. In addition to the feed-forward control by ADP, the adenine nucleotide system imposes a strong feed-back activation on phosphofructokinase via AMP, the allosteric activator of the latter. As should be expected, ATP and
Publ
ishe
d on
01
Janu
ary
1974
. Dow
nloa
ded
by U
nive
rsita
t Pol
itècn
ica
de V
alèn
cia
on 2
6/10
/201
4 08
:07:
19.
View Article Online
206 BIOCHEMICAL OSCILLATlONS I N T l M E AND SPACE
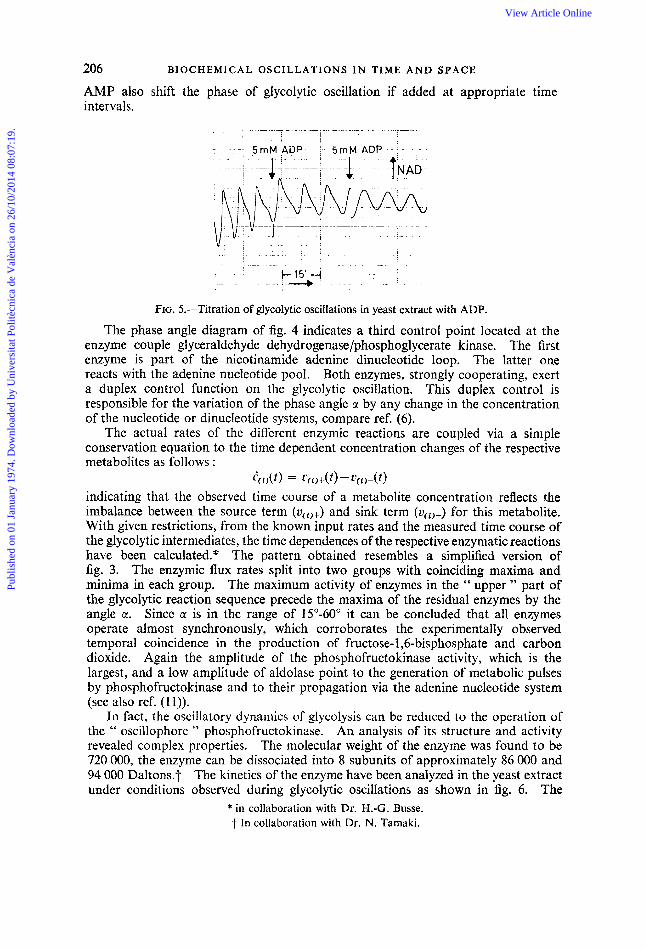
AMP also shift the phase of glycolytic oscillation if added at appropriate intervals.
time
.- . . . . .. -. ._ . . .. .
. .. : . I b 1 5 ' -4
: . - - i-- . . . . .
FIG. 5.-Titration of glycolytic oscillations in yeast extract with ADP.
The phase angle diagram of fig. 4 indicates a third control point located at the enzyme couple glyceraldehyde dehydrogenase/phosphoglycerate kinase. The first enzyme is part of the nicotinamide adenine dinucleotide loop. The latter one reacts with the adenine nucleotide pool. Both enzymes, strongly cooperating, exert a duplex control function on the glycolytic oscillation. This duplex control is responsible for the variation of the phase angle a by any change in the concentration of the nucleotide or dinucleotide systems, compare ref. (6).
The actual rates of the different enzymic reactions are coupled via a simple conservation equation to the time dependent concentration changes of the respective metabolites as follows :
indicating that the observed time course of a metabolite concentration reflects the imbalance between the source term (vci,+) and sink term (uci)-) for this metabolite. With given restrictions, from the known input rates and the measured time course of the glycolytic intermediates, the time dependences of the respective enzymatic reactions have been calculated.* The pattern obtained resembles a simplified version of fig. 3. The enzymic flux rates split into two groups with coinciding maxima and minima in each group. The maximum activity of enzymes in the '' upper " part of the glycolytic reaction sequence precede the maxima of the residual enzymes by the angle a. Since cc is in the range of 15"-60" it can be concluded that all enzymes operate almost synchronously, which corroborates the experimentally observed temporal coincidence in the product ion of fructose- 1,6-bisphosphate and carbon dioxide. Again the amplitude of the phosphofructokinase activity, which is the largest, and a low amplitude of aldolase point to the generation of metabolic pulses by phosphofructokinase and to their propagation via the adenine nucleotide system (see also ref. (1 1)).
In fact, the oscillatory dynamics of glycolysis can be reduced to the operation of the " oscillophore " phosphofructokinase. An analysis of its structure and activity revealed complex properties. The molecular weight of the enzyme was found to be 720 000, the enzyme can be dissociated into 8 subunits of approximately 86 000 and 94 000 Daltons.7 The kinetics of the enzyme have been analyzed in the yeast extract under conditions observed during glycolytic oscillations as shown in fig. 6. The
eci,(t) = ~ 7 ~ i ) + ( t ) - ~ c i , - ( t >
* in collaboration with Dr. H.-G. Busse. -f In collaboration with Dr. N. Tamaki.
Publ
ishe
d on
01
Janu
ary
1974
. Dow
nloa
ded
by U
nive
rsita
t Pol
itècn
ica
de V
alèn
cia
on 2
6/10
/201
4 08
:07:
19.
View Article Online
A . B O l T E U X A N D B . HESS
10”’-
lo-’-
2 IO-?
fl to-L-
K
2 2
C .- ..I
1 o-s-
1 d6-
207
Limit Conditions [mM]
ATP ADP AMP A . 1 1.5 1.1
8: 3 0,5 0,l
1 2 3 4 5 6 i [F-6-P]/103 M-’
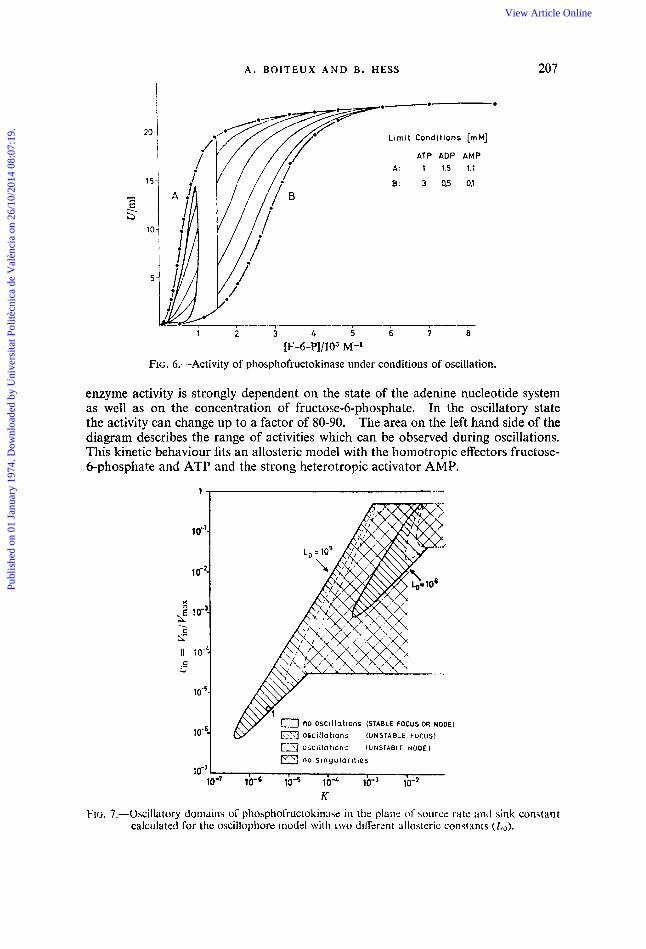
FIG. 6.-Activity of phosphofructokinase under conditions of oscillation.
enzyme activity is strongly dependent on the state of the adenine nucleotide system as well as on the concentration of fructose-6-phosphate. In the oscillatory state the activity can change up to a factor of 80-90. The area on the left hand side of the diagram describes the range of activities which can be observed during oscillations. This kinetic behaviour fits an allosteric model with the homotropic effectors fructose- 6-phosphate and ATP and the strong heterotropic activator AMP.
.....
’I
0 no oscillations (STABLE FOCUS OR NODE)
m oscillations (UNSTABLE FOCUS)
&+Y ‘ 0 no oscillations (STABLE FOCUS OR NODE)
m oscillations (UNSTABLE FOCUS)
osciilutions (UNSTABLE NODE) a no s ingular i t ies
K FIG. 7.-Oscillatory domains of phosphofructokinase in the plan; of sourcz rate and sink con$tm t
calculated for the osciilophore model with two dilferent allosteric constants (Lo) .
Publ
ishe
d on
01
Janu
ary
1974
. Dow
nloa
ded
by U
nive
rsita
t Pol
itècn
ica
de V
alèn
cia
on 2
6/10
/201
4 08
:07:
19.
View Article Online
208 BIOCHEMICAL OSCILLATIONS I N T I M E A N D SPACE
On the basis of the allosteric properties, a mathematical model has been developed, which is open and exhibits oscillatory self-excitation.12 The model displays a variety of dynamic domains, which depend on the source rate, the sink constant and the allosteric constant Lo (see fig. 7).* It is obvious that oscillations can only occur at conditions distant from equilibrium. The model describes in detail both period and amplitude of glycolytic oscillations as well as their change observed experimentally on variation of the injection rate of substrate.
TABLE 1 .-INTERACTION OF GLYCOLYTIC OSCILLATOR ( Tl ) WITH MODULATED RATE OF SUBSTRATE INJECTION ( T2). CALCULATED FROM EXPERIMENTS WITH YEAST EXTRACT
relation of periods interaction
entrainment on periodic (T2) pulsatory bursts of some cycles with the autonomous period TI
1 . 7 ~ Ti > 7'2 > 1 . 2 ~ Ti no synchronisation ; period and amplitude variable
2-2 N T,/n (n = 1.2, . . .) entrainment on exactly IZ x T2
Recently, entrainment of the glycolytic oscillophore to a periodically varied injection rate has been demonstrated experimentally. This behaviour is predicted from the common properties of non-linear oscillators. Whether, and what type of, entrainment is observed depends on the relation of the periods of the oscillophore TI and of the input rate T2 as shown in table 1. Given a stochastic variation of this rate the oscillophore settles at its autonomous frequency. Indeed, a com- parison between the biochemical experiments and the properties of the model reveal a remarkable congruence.
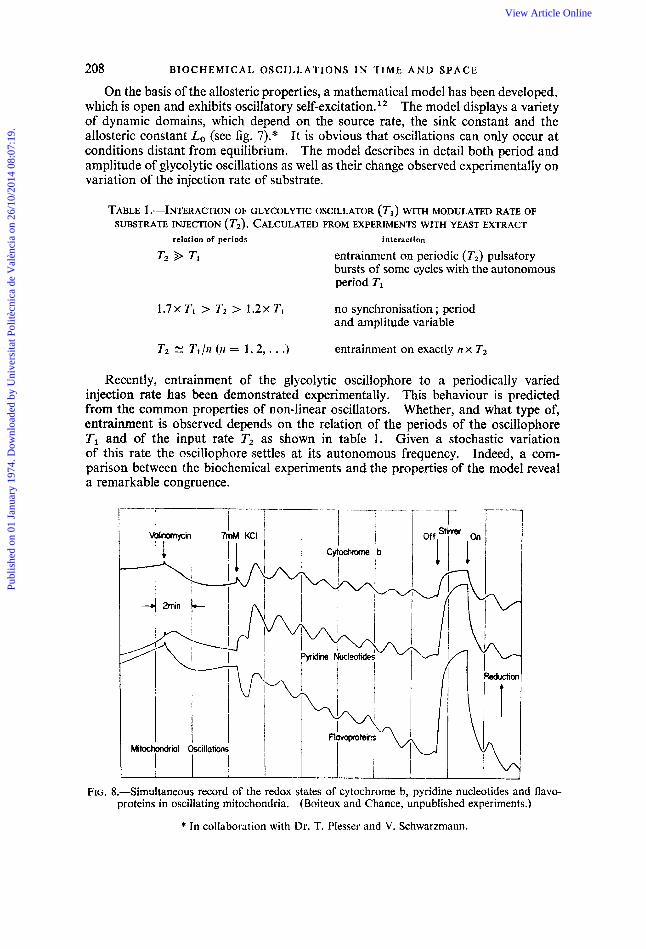
FIG. 8.-Simultaneous record of the redox states of cytochrome b, pyridine nucleotides and flavo- proteins in oscillating mitochondria. (Boiteux and Chance, unpublished experiments.)
* In collaboration with Dr. T. Plesser and V. Schwarzmann.
Publ
ishe
d on
01
Janu
ary
1974
. Dow
nloa
ded
by U
nive
rsita
t Pol
itècn
ica
de V
alèn
cia
on 2
6/10
/201
4 08
:07:
19.
View Article Online
A . BOITEUX AND €3. HESS 209 In partial summary, one now notes that analysis and simulation of the oscillating
glycolysis reveal a number of basic principles, which obviously are necessary condi- tions for this phenomenon :
(i) The " primary source " of the oscillations has a non-linear kinetic character- istic.
(ii) The kinetic structure involves some type of feed-back interaction. (iii) The system moves on a limit cycle. (iv) The system operates far from equilibrium and is thermodynamically open.
OSCILLATION IN MITOCHONDRIAL RESPIRATION
The control system of cellular respiration possesses an inherent potential to break into self-sustained oscillations. In a number of laboratories, this property has been observed during studies of intact cells as well as of isolated mitochondria.1 In a typical experiment, fig. 8 demonstrates the dynamics of some components of the respiratory chain in suspensions of pigeon heart mitochondria, supplied with substrate and oxygen. Since an integral part of the oscillatory system is the rate of ion transport across the mitochondrial membrane, which must match the rate of electron flow through the respiratory chain, the oscillation can be influenced by interaction with the mechanism of ion transport. Therefore, in this experiment valinomycin, an ionophoric antibiotic, is used to activate the potassium transport. It is shown, that the oscillations start right away, as soon as potassium ions are added. The use of an ionophore, however, is not obligatory. Oscillations can be induced as well by other methods critically matching the rate of ion transport and electron flow. As shown in the figure, the components of the respiratory chain, cytochrome b, pyridine nucleotides and flavoproteins are oxidized and reduced simultaneously with a period of about 70 s, all three responding with identical kinetics to the change of oxygen potential, which is maintained by vigorous stirring. The mitochondrial system is complex enough to show a variety of dynamic states. In a phase plane plot of the redox
0 1 40 0 10 20 30
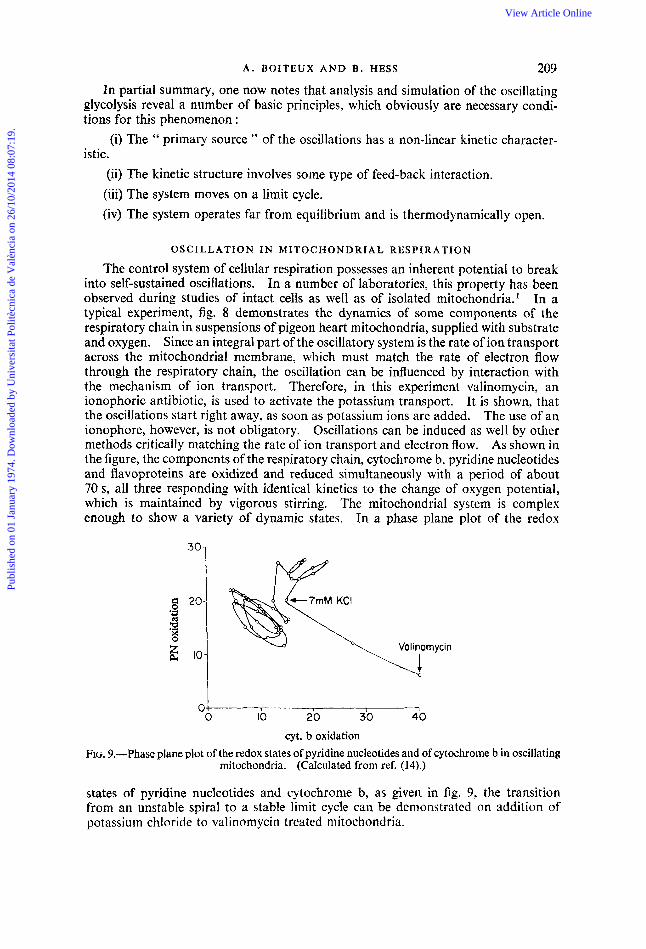
cyt. b oxidation FIG. 9.-Phase plane plot of the redox states of pyridine nucleotides and of cytochrome b in oscillating
mitochondria. (Calculated from ref. (141.)
states of pyridine nucleotides and cytochrome b, as given in fig. 9, the transition from an unstable spiral to a stable limit cycle can be demonstrated on addition of potassium chloride to valinoinycin treated mitochondria.
Publ
ishe
d on
01
Janu
ary
1974
. Dow
nloa
ded
by U
nive
rsita
t Pol
itècn
ica
de V
alèn
cia
on 2
6/10
/201
4 08
:07:
19.
View Article Online
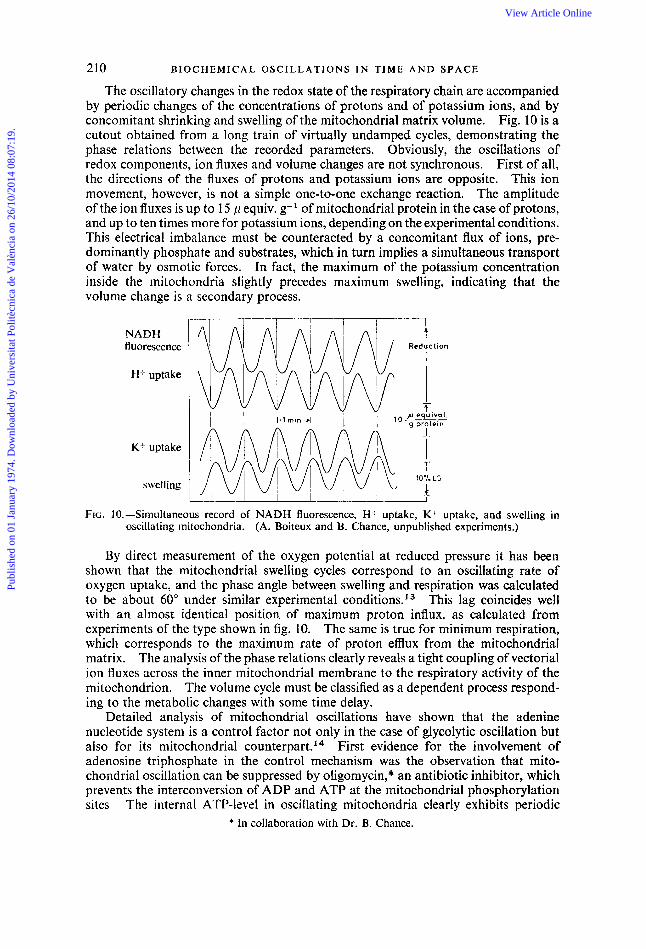
210 B I O C H E M I C A L O S C I L L A T I O N S IN T I M E A N D S P A C E
The oscillatory changes in the redox state of the respiratory chain are accompanied by periodic changes of the concentrations of protons and of potassium ions, and by concomitant shrinking and swelling of the mitochondria1 matrix volume. Fig. 10 is a cutout obtained from a long train of virtually undamped cycles, demonstrating the phase relations between the recorded parameters. Obviously, the osciIlations of redox components, ion fluxes and volume changes are not synchronous. First of all, the directions of the fluxes of protons and potassium ions are opposite. This ion movement, however, is not a simple one-to-one exchange reaction. The amplitude of the ion fluxes is up to 15 p equiv. g-' of mitochondrial protein in the case of protons, and up to ten times more for potassium ions, depending on the experimental conditions. This electrical imbalance must be counteracted by a concomitant flux of ions, pre- dominantly phosphate and substrates, which in turn implies a simultaneous transport of water by osmotic forces. In fact, the maximum of the potassium concentration inside the mitochondria slightly precedes maximum swelling, indicating that the volume change is a secondary process.
NADH fluorescence
K+ uptake
swell i ng
-1 t I
Reduction
g protein I
T 10% LS
h -J
FIG. IO. -Siniultaneous record of NADH fluorescence, H uptake, K+ uptake, and swelling in oscillating mitochondria, (A. Boiteux and B. Chance, unpublished experiments.)
By direct measurement of the oxygen potential at reduced pressure it has been shown that the mitochondrial swelling cycles correspond to an oscillating rate of oxygen uptake, and the phase angle between swelling and respiration was calculated to be about 60" under similar experimental condition^.'^ This lag coincides well with an almost identical position of maximum proton influx, as calculated from experiments of the type shown in fig. 10. The same is true for minimum respiration, which corresponds to the maximum rate of proton efflux from the mitochondrial matrix. The analysis of the phase relations clearly reveals a tight coupling of vectorial ion fluxes across the inner mitochondrial membrane to the respiratory activity of the mitochondrion. The volume cycle must be classified as a dependent process respond- ing to the metabolic changes with some time delay.
Detailed analysis of mitochondrial oscillations have shown that the adenine nucleotide system is a control factor not only in the case of glycolytic oscillation but also for its mitochondrial co~nterpar t . '~ First evidence for the involvement of adenosine triphosphate in the control mechanism was the observation that mito- chondrial oscillation can be suppressed by oligomycin,* an antibiotic inhibitor, which prevents the interconversion of ADP and ATP at the mitochondrial phosphorylation sites The internal ATP-level in oscillating mitochondria clearly exhibits periodic
* In collaboration with Dr. B. Chance.
Publ
ishe
d on
01
Janu
ary
1974
. Dow
nloa
ded
by U
nive
rsita
t Pol
itècn
ica
de V
alèn
cia
on 2
6/10
/201
4 08
:07:
19.
View Article Online
A . BOITEUX AND B . HESS 21 1
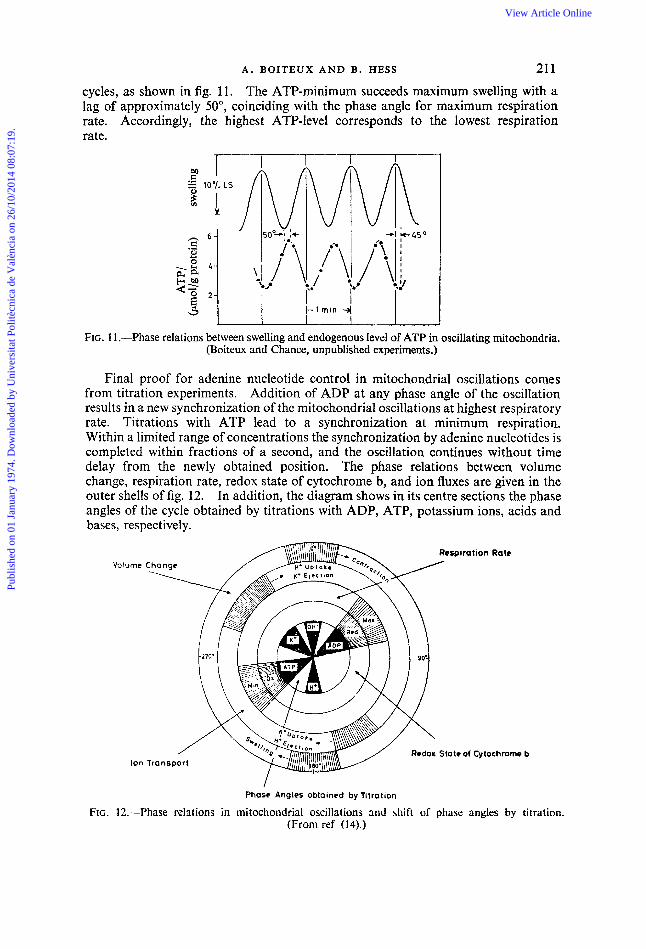
cycles, as shown in fig. 1 1 . The ATP-minimum succeeds maximum swelling with a lag of approximately 50°, coinciding with the phase angle for maximum respiration rate. Accordingly, the highest ATP-level corresponds to the lowest respiration rate.
FIG. 11 .--Phase relations between swelling and endogenous level of ATP in oscillating mitochondria. (Boiteux and Chance, unpublished experiments.)
Final proof for adenine nucleotide control in mitochondrial oscillations comes from titration experiments. Addition of ADP at any phase angle of the oscillation results in a new synchronization of the mitochondrial oscillations at highest respiratory rate. Titrations with ATP lead to a synchronization at minimum respiration. Within a limited range of concentrations the synchronization by adenine nucleotides is completed within fractions of a second, and the oscillation continues without time delay from the newly obtained position. The phase relations between volume change, respiration rate, redox state of cytochrome b, and ion fluxes are given in the outer shells of fig. 12. In addition, the diagram shows in its centre sections the phase angles of the cycle obtained by titrations with ADP, ATP, potassium ions, acids and bases, respectively.
Volume
Ion
Phase Angles obtained by Titration
FIG. 12.-Phase relations in mitochondrial oscillations and shift of phase angles by titration. (From ref (14).)
Publ
ishe
d on
01
Janu
ary
1974
. Dow
nloa
ded
by U
nive
rsita
t Pol
itècn
ica
de V
alèn
cia
on 2
6/10
/201
4 08
:07:
19.
View Article Online

212 BIOCHEMICAL OSCILLATIONS I N T I M E A N D SPACE
From these experiments it is obvious that the ADP/ATP-ratio in the mitochondria1 matrix is a control factor of the oscillations, though not the only one. Experiments with valinomycin clearly demonstrate that the activity of the potassium carrier determines both amplitude and period of the o~cillation.'~ Thus, the rate of ion transport across the mitochondrial membrane is another control factor in addition to the endogenous adenosine phosphate system. In fact, membrane-bound carrier systems are the receptor sites for controlling and synchronizing signals in suspending medium and cytosol, respectively. By titrations with acids and bases (cf. fig. 12) the external synchronizer of the mitochondrial population was shown to be a proton gradient l4 across the membrane. This gradient developing along with the proton flux serves as a very effective positive feed-back signal to the respiratory system, promoting strong self-amplification. Fig. 12 shows that an addition of protons shifts the cycle to the onset of swelling and to further proton efflux from the matrix space. Addition of hydroxyl ions immediately shifts the mitochondrial system to the position, where it starts shrinking and taking up protons from the medium.
The study demonstrates that the mitochondrial energy production is tightly controlled by a multiplicity of signals from the cytosolic space, the rate of ion fluxes across the inner membrane being a critical controlling parameter. The detection of the very effective feed-back action of a proton gradient raises the question of the rela- tion between the membrane charge and the structural and functional changes at critical control sites. Though the mechanism of generation of mitochondrial oscillations is not yet known in detail, it is obvious, that the statements from points (i)-(iv) of the previous section also hold for this case. (1) The involvement of vectorial transport processes for controlling reaction steps leads to non-linear kinetics. (2) The self-exciting proton gradient is the basis of the feed-back structure of the membrane system. (3) Fig. 9 shows that the system moves on a limit cycle. (4) Approximation towards the equilibrium state by substrate exhaustion or oxygen limitation leads to rapid damping of the oscillation, which can be maintained only if a minimal flux is provided. The system is open for the source, namely oxygen, and for a number of sink reactions, leading to the production of water, carbon dioxide, and heat.
OSCILLATIONS IN CELLULAR COMMUNICATION
The source and nature of the spatial organization of living systems is one of the fundamental problems of the biological sciences. More than 20 years ago the question of stability of biological systems with respect to diffusion was analyzed by Turing and later by Glansdorff and Prigogine.' Nowadays it is known that rotation of chemical and biochemical systems around an unstable singular point coupled to diffusion does occur, resulting in chemical waves. The generation of chemical waves has been observed in the case of the classical Belousov-Zhabotinsky reaction (see ref. ( I) and (4)) and it has been shown that this system is able to transmit information.
Recently, it has been found that the propagation of biochemical waves plays a significant role in the process of morphogenesis of the slime mould Dictyustelium discoideum.'' Within its life cycle the slime mould passes through a growth phase in which the organism exists in the state of single ameboid cells in a given life territory. After the end of the growth phase the single cells aggregate in response to chemotactic stimuli and finally form a multi-cellular body which differentiates to become the fruiting body, which is the source of the next generation.
This is an example of self-organization of spatial patterns by chemical communica- tion starting with a layer of randomly distributed identical cells. The aggregation territories are controlled by the centres which release chemical pulses of cyclic AMP
Publ
ishe
d on
01
Janu
ary
1974
. Dow
nloa
ded
by U
nive
rsita
t Pol
itècn
ica
de V
alèn
cia
on 2
6/10
/201
4 08
:07:
19.
View Article Online
A . BOITEUX AND B. HESS 213
with a frequency of 0.2-0.3 min-I. The pulses are propagated from cell to cell as excitation waves which spread in a uniform layer with a constant speed of 40-50 pm m i d . The waves are either concentric or spiral shaped. Formally the system can be treated as a set of diffusion coupled oscillators in which the DictyusteZium cells operate as the oscillating element by receiving, amplifying and ejecting periodically concentration gradients of cyclic AMP.
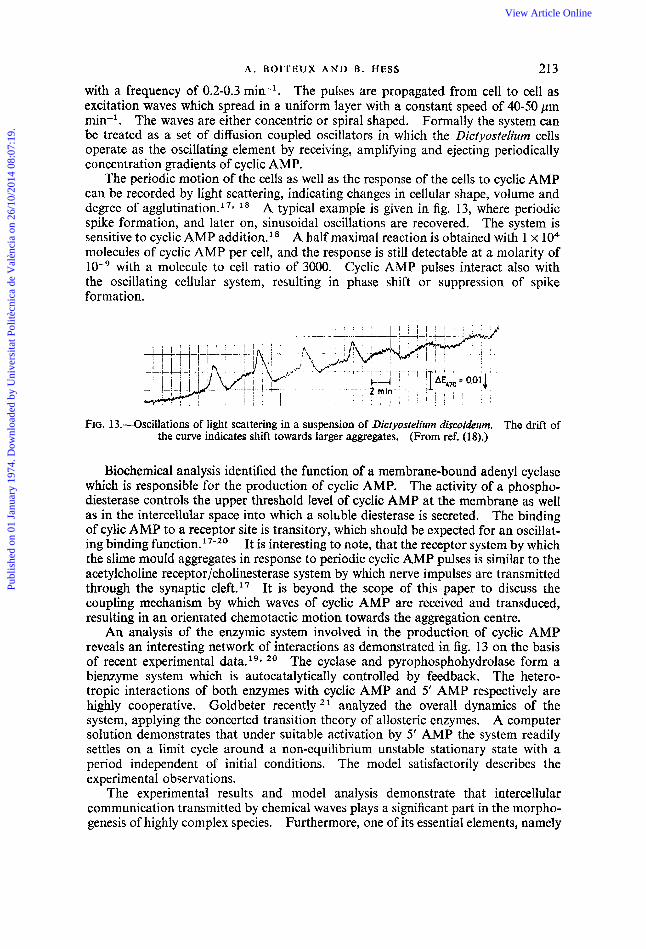
The periodic motion of the cells as well as the response of the cells to cyclic AMP can be recorded by light scattering, indicating changes in cellular shape, volume and degree of aggl~tination. '~~ l 8 A typical example is given in fig. 13, where periodic spike formation, and later on, sinusoidal oscillations are recovered. The system is sensitive to cyclic AMP addition.l* A half maximal reaction is obtained with 1 x lo4 molecules of cyclic AMP per cell, and the response is still detectable at a molarity of
with a molecule to cell ratio of 3000. Cyclic AMP pulses interact also with the oscillating cellular system, resulting in phase shift or suppression of spike formation.
FIG. 13.-Oscillations of light scattering in a suspension of Dictyostelium discoideum. The drift of the curve indicates shift towards larger aggregates, (From ref. (18).)
Biochemical analysis identified the function of a membrane-bound adenyl cyclase which is responsible for the production of cyclic AMP. The activity of a phospho- diesterase controls the upper threshold level of cyclic AMP at the membrane as well as in the intercellular space into which a solhble diesterase is secreted. The binding of cylic AMP to a receptor site is transitory, which should be expected for an oscillat- ing binding function.' 7-20 It is interesting to note, that the receptor system by which the slime mould aggregates in response to periodic cyclic AMP pulses is similar to the acetylcholine receptor/cholinesterase system by which nerve impulses are transmitted through the synaptic ~ 1 e f t . l ~ It is beyond the scope of this paper to discuss the coupling mechanism by which waves of cyclic AMP are received and transduced, resulting in an orientated chemotactic motion towards the aggregation centre.
An analysis of the enzymic system involved in the production of cyclic AMP reveals an interesting network of interactions as demonstrated in fig. 13 on the basis of recent experimental data. 19* 2o The cyclase and pyrophosphohydrolase form a bienzyme system which is autocatalytically controlled by feedback. The hetero- tropic interactions of both enzymes with cyclic AMP and 5' AMP respectively are highly cooperative. Goldbeter recently analyzed the overall dynamics of the system, applying the concerted transition theory of allosteric enzymes. A computer solution demonstrates that under suitable activation by 5' AMP the system readily settles on a limit cycle around a non-equilibrium unstable stationary state with a period independent of initial conditions. The model satisfactorily describes the experimental observations.
The experimental results and model analysis demonstrate that intercellular communication transmitted by chemical waves plays a significant part in the morpho- genesis of highly complex species. Furthermore, one of its essential elements, namely
Publ
ishe
d on
01
Janu
ary
1974
. Dow
nloa
ded
by U
nive
rsita
t Pol
itècn
ica
de V
alèn
cia
on 2
6/10
/201
4 08
:07:
19.
View Article Online

214 BIOCHEMICAL OSCILLATIONS IN T I M E A N D SPACE
the pulsewise production of the transmitting chemicals, could be reduced and identified as the periodic function of the cyclic AMP producing enzyme in the cellular wall. Indeed, nonlinear chemical and biocheinical oscillators are well suitable to serve as devices to measure time, to memorize chemicals and to synchronize chemical activities.
J( A T P
Adeny lcyc lase Py ropho s-phohydro la se
Cyc l ic AMP
\ G-AMP +
? Phosphodiesterase 1
FIG. 14.-Enzyme network for the control of the cyclic AMP-level. (From ref. (17)).
B. Hess and A. Boiteux, Ann. Rev. Biochem., 1971, 40,237. B. Chance, E. K. we, A. K. Ghosh and B. Hess, Biological and Biochemical Oscillators (Academic Press, New York and London, 1973). B. Hess, A. Boiteux, H.-G. Busse and G. Gerisch, Membranes, Dissbatiue Structures in Evolution (Wiley-Interscience New York, 1974) in press. G. Nicolis and J. Portnow, Chem. Rev., 1973,73, 365. P. Glansdorff and I. Prigogine, Thermodynamic Theory of Structure, Stability and Fluctuations (Wiley-Interscience, N.Y., 1971). B. Hess, A. Boiteux and 3. Kriiger, A h . Enzyme Regul., 1969, 7, 149. ’ B. Bess and A. Boiteux, Hoppe-Seyler’s 2. Physiol. Chew., 1968,349, 1567. R. E. Reeves and A. Sols, Biochem. Biophys. Res. Comm. 1973’50,459. B. Hess and A. Boiteux, Biochim. Biophys. Acta Library, 1968, 11, 148.
lo B. Chance, R. Schoner and S. Elsaesser, Proc. Natl. Acad. Sci. USA, 1964, 52, 335. l 1 B. Hess, Nova Acta Leopoldina, 1968, 33, 195. l 2 A. Goldbeter and R. Lefever, Biophys., 1972, 12, 1302. l3 A. Boiteux and H. Degn, Hoppe-Seyler’s 2. Physiol. Chem., 1972, 353, 696. l4 A. Boiteux, Ergebn. exp. Medizin 9 (Volk und Gesundheit, Berlin, 1972), p. 347. l5 A. M. Turing, Phil. Trans. B, 1952,237,37. l6 H.-G. Busse and B. Hess, Nature, 1973, 244,203. l7 G. Gerisch, D. Malchow and B. Hess, Cell Communicatioiz and cyclic AMP Regulation during
Aggregation of the Slime Mould Dictyostelium Discoideum (Biochemistry of Sensory Functions) (Springer Verlag, Heidelberg, Berlin, New York) in press.
l8 F. Gerisch and B. Hess, Proc. Natl. Acad. Sci. USA, 1974,71, 2118. l9 E. F. Rossomando and M. Sussman, Proc. Natl. h a d . Sci. USA, 1973, 70, 1254. 2o D. Malchow and G. Gerisch, Proc. Natl. Acad. Sci. USA, 1974, 71, 2423. 2 1 A. Goldbeter, Nature, 1974.
Publ
ishe
d on
01
Janu
ary
1974
. Dow
nloa
ded
by U
nive
rsita
t Pol
itècn
ica
de V
alèn
cia
on 2
6/10
/201
4 08
:07:
19.
View Article Online