Embed Size (px)
Citation preview

Neuropsychologia, Vol. 25, No. 18, pp. 299-304, 1987. Printed in Great Britain.
0028-3932/x7 f3.00+0.00 CN 1987 Pergamon Journals Ltd.
NOTE
P300 FROM IDENTICAL TWINS*
JOHN POLICH~ and TIM BURNS
Division of Prechnical Neuroscience and Endocrinology, Scripps Clinic and Research Foundation, La Jolla, CA 92031, U.S.A.
(Received 14 April 1986; Accepted 26 August 1986)
Abstract---The P300 component was obtained from ten pairs of identical twins and matched control subjects with an auditory discrimination task. The event-related brain potentials (ERPs) from the identical twins were strikingly similar in amplitude and latency compared to control subject pairs. The data suggest that individual variations in ERP waveform morphology are determined by the structure of the neurophysiological mechanisms responsible for P300 generation.
THE P300 or P3 component of the event-related brain potential (ERP) is a large (l&20 pV), positive-going waveform elicited whenever subjects discriminate stimuli. Possibly originating in the hippocampus and amygdaloid structures, [IO, 16,303, its production has been linked to the psychological processes underlying memory updating when the neural representation of the stimulus environment is modified to accommodate incoming sensory information [S, 61. Because the P3 component appears to be an electrophysiological measure of fundamental cognitive activity, its possible use as an evaluative index of population differences in mental capability has been widely explored. For example, P3 latency has been found to decrease with increases in cognitive development stemming from maturation in children [4, 111. Adult aging [2,9, 19,251 and neurological impairment [13, 18,27, 3 I] have been correlated with increases in P3 latency. In addition, P3 amplitude variations have been associated with individual differences in memory function 112,241, differential inheritability of alcoholism [l, 7,213, and the residual effects of alcohol consumption [15, 231. Thus, the P3 ERP component is beginning to provide an electrophysiological index of the cognitive processes that covary with physiological changes.
An auditory stimulus paradigm in which the subject is required to discriminate a target tone from a nontarget or standard tone often is employed for these investigations. This procedure reliably produces an easily quantifiable P3 component, although stimulus parameters can affect its amplitude and latency [I 7,261. It is clear from many reports [2,24,29], however, that even within homogeneous populations the P3 component from this simple discrimination task varies greatly across individuals: P3 amplitude can range from just a few to more than 25 pV, while peak latency ranges from 250 to as much as 375 msec [20]. Although many studies have attempted to relate differences in P3 amplitude and latency to neuropsychological variables, neurophysiological differences such as cranium size, skull thickness, and/or the physical location and structure of the neural apparatus which produces the P3 could also affect component measures. However, these factors have remained largely unexplored.
The present study was conducted to provide a preliminary basis for assessing the general role of neurophysiological factors as contributors to individual differences in P3 morphology. Previous evoked potential studies have found marked waveform similarity for monozygotic twins using repetitive stimulus presentations 13, 141. Comparison of mono- and dyzygotic twin auditory evoked potentials have suggested a strong genetic basis for this similarity [28]. A single ERP study employing a few pairs of subjects noted greater similarity for P3 latency of the twins relative to controls, but no amplitude data were reported nor were the intra-pair correlations for the twins or controls given [32]. If variation in P3 morphology is determined in part by variation in biological structures, then identical twins should manifest highly similar waveforms relative to control pairs since they share their neurophysiological makeup. The degree of their similarity may then provide some insight into the normative P3 variation due to non-cognitive factors.
*The research reported here was supported by the J. M. McDonald Foundation and NIAAA grant AA6420 and is publication number 4082BCR from the Research Institute of Scripps Clinic.
tcorrespondence to be addressed to John Polich, Preclinical Neuroscience (BCRl), Scripps Clinic and Research Foundation, 10666 N. Torrey Pines Road, La Jolla, CA 92037, U.S.A.
299

300 NOTE
METHODS Subjects
Ten pairs ofidentical twins were obtained by verbal advertisement. Their monozygocity was carefully ascertained with background questions and visual inspection by independent observers, since blood tests could not be performed. Twins who expressed doubt or who did not appear identical were excluded from the sample (three pairs of 13 contacted). This evaluation of physical similarity and background information produced perfect agreement between the different observers. Control subjects were matched to the twins on sex, age, educational level, race, and alcohol consumption. Subjects ranged in age from 18-30 years. All subjects reported being free from neurological or psychiatric disorders and received course credit or were paid for their participation.
Recording conditions
ERPs were elicited with binaural 1000 and 2000 Hz tones delivered at 60 dB SPL and having a 9.9 msec rise/fall with 50 msec plateau times. The tones were presented in a random series once every second, with the low tone designated as the standard and the high tone designated as the target stimulus. Electroencephalographic (EEG) activity was recorded at the Cz electrode site of the l&20 system using a gold-plated electrode affixed with electrode paste and tape, referred to linked earlobes with a forehead ground. Additional electrodes were placed at the outer canthus and below the left eye to monitor electro-ocular (EOG) activity. Impedance for all recording sites was 10 kR or less. The filter bandpass was 0.5 to 30 Hz (3 db down, 12 db octave/slope). The EEG was digitized at 1.5 msec per point for 768 msec with a 75 msec prestimulus baseline. Waveforms were averaged on-line by a Nicolet Pathfinder II which also controlled the stimulus presentation and artifact rejection. Trials on which the EEG or EOG exceeded +45 PV were automatically rejected.
Procedure
All subjects were recorded while sitting down with their eyes closed and required to move the index finger of their right hand whenever they detected a target tone. Two blocks of 100 trials with 20% target stimuli were obtained to ensure waveform replicability and facilitate accurate component identification. All subjects performed the task with virtually no errors and very few artifactual responses. Care was taken that the twin and control subjects received exactly the same treatment during the recording session.
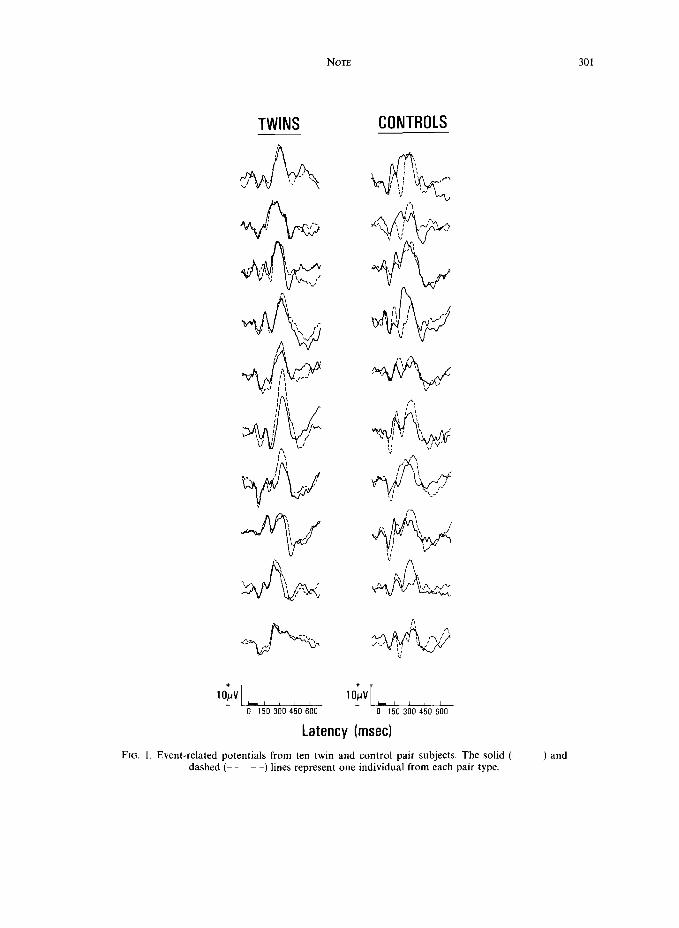
RESULTS The target tone ERP data from the ten twin and control pairs are presented in Fig. 1. Solid and dashed lines
represent the waveforms obtained from two different individuals. Each individual waveform is comprised of the 40 target trials obtained from the sum of the two blocks. The amplitudes and latencies for the Nl, P2, N2 and P3 components were measured at the most negative or positive peak within the latency ranges of 80--120, 140-240, 15G240 and 230-370 msec, respectively. Amplitudes were assessed relative to the prestimulus baseline, while latencies were taken as the time point defining the component peak.
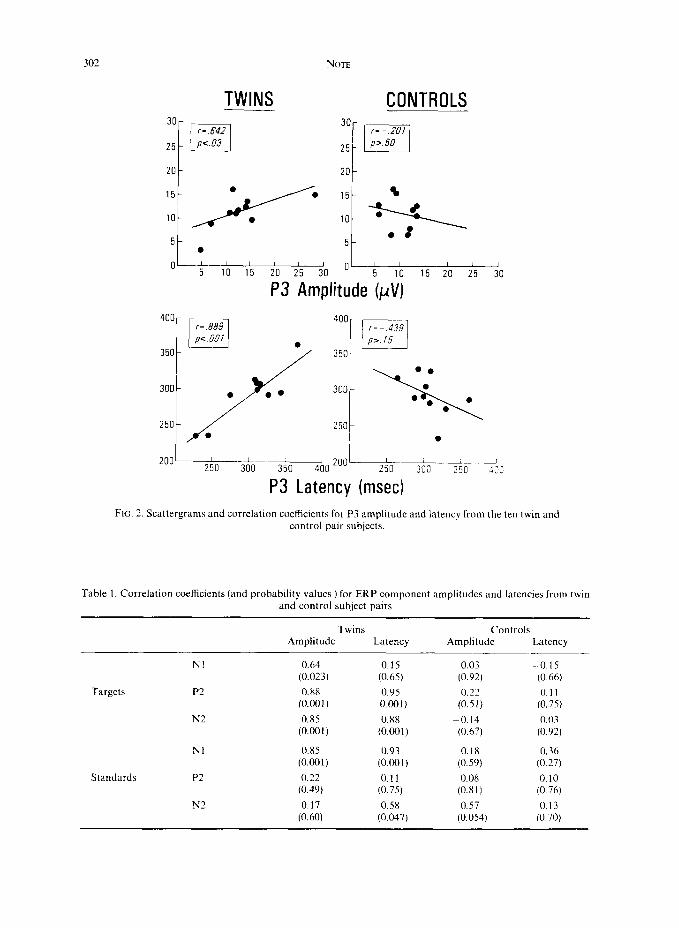
Pearson r correlation coefficients were computed on the subject pairs from each group separately for the amplitude and latency from each component. The results are presented in Fig. 2 for the P3 and Table 1 for the other components, with the significance level of each noted. These results indicated that the amplitude and latency of target components (except for Nl latency) obtained from the twin pairs were significantly correlated, whereas none of the control pair components were reliably related. For the ERP components obtained from the standard tones, the Nl and N2 from the twin pairs yielded significant correlations, while none of the control pair data demonstrated such an effect.
DISCUSSION The similarity of the P3 components obtained from identical twins is striking and consistent with previous twin
study reports for evoked responses [3, 14, 281 and endogenous components [32]. The variability of the P3 in the control subjects illustrates the variation of the ERP typically observed from this simple auditory discrimination task 12, 24,291. The morphological similarity in the twin waveforms compared to the control pair ERPs suggests that individual differences in the P3 component are determined by the underlying neurophysiological structures associated with its generation.
Although it can be argued that the twin pairs are mentally alike and that this variable could account for the observed ERP consistency, the simple two-tone discrimination paradigm employed in the present study requires relatively low-level cognitive processes, vis-a-vis evaluation, recognition, or memory retrieval tasks. Because a wide variety of P3 values can be obtained from a homogeneous sample (i.e., university students) with a narrow range of cognitive ability [20], similarity in the electrophysiological responses between the twin pairs compared to the matched controls most likely stems from their identical neurophysiological structure rather than cognitive backgrounds or strategies. In addition, the significant correlations for the Nl, P2 and N2 amplitudes and latencies
NOTE 301
TWINS CONTROLS
l&VI!, , / , , lo;V~_, , , , - 0 150 300 450 600 0 150 300 450 600
Latency (msec)
FIG. 1. Event-related potentials from ten twin and control pair subjects. The solid ( -) and dashed (- - ~ -) lines represent one individual from each pair type.
302 NOTE
TWINS CONTROLS
I r=.642 PC.03
30 r- -.201
25 ‘II 1
PD.50
20 25 30
P3 Amplitude (pV)
r=.889 II 400 r
pc.OOI l
350
0 / I
0. 300
r= - 439 _,I p,. 15
2ooL---i / ’ 250 300 350 400
200~‘~ 250 300 35000
P3 Latency (msec)
FIG. 2. Scattergrams and correlation coefficients for P3 amplitude and latency from the ten twin and control pair subjects.
Table I. Correlation coefficients (and probability values ) for ERP component amplitudes and latencies from twin and control subject pairs
Targets
Nl
P2
N2
Twins Controls Amplitude Latency Amplitude Latency
0.64 0.15 -0.03 -. 0. I5 (0.023) (0.65) (0.92) (0.66)
0.88 0.95 PO.22 0. I 1 (0.001) 0.001) (0.51) (0.75)
0.85 0.88 -0.14 0.03 (0.001) (0.001) (0.67) (0.92)
Standards
NI 0.85 0.93 0.18 0.36 (0.001) (O.cOl) (0.59) (0.27)
P2 0.22 0.11 0.08 ~ 0. IO (0.49) (0.75) (0.81) (0.76)
N2 0.17 0.58 0.57 0.13 (0.60) (0.047) (0.054) (0.70)

NOTE 303
from the twin pairs (8 of 12) compared to the control subjects (0 of 12) also implies an underlying structural similarity, since these components are more sensitive to exogenous rather than cognitive variables 161. If these conclusions are accurate, then the role of physiological variables in the production of the P3 ERP component needs to be ascertained more precisely in order to develop further electrophysiological methods for the evaluation of human cognitive function.
5. 6.
I.
8.
9
10.
11. 12.
13.
14.
1.5.
16.
17.
18.
19.
20. 21. 22.
23.
REFERENCES
BEGLEITER, H., PORJESZ, B., BIHARI, B. and KISSIN, B. Event-related brain potentials in boys at risk for alcoholism. Science 225, 1493-1495, 1984. BROWN, W., MARSH, J. and LARUE, A. Exponential electrophysiological aging: P300 latency. Electroenceph. Clin. Neurophysiol. 55, 211.-285, 1983. BUCHSBAUM, M. S. Average evoked response and stimulus intensity in identical and fraternal twins. Physiol. Psychol. 2, 365-370, 1974. COURCHESNE, E. Cognitive components of the event-related brain potential: changes associated with development. In Tutorials in ERP Research: Endogenous Components, A. W. K. GAILI.ARD and W. RITTER (Editors), pp. 329-344. North-Holland, Amsterdam, 1984. DONCHIN, E. Surprise! Surprise? Psychophysiology 18, 493-513, 1981. DONTHIN, E., RITTER, W. and MCCALLUM, C. Cognitive psychophysiology: the endogenous components of the ERP. In Brain Event-Related Potentials in Man, E. CALLAWAY, P. TUETING and S. KOSLOW (Editors), pp. 349411. Academic Press, New York, 1978. ELMASIAN, R., NEVILLE, H., WOODS, D., SCHUCKIT, M. and BLOOM, F. E. Event-related brain potentials are different in individuals at high and low risk for developing alcoholism. Proc. mtn. Acad. Sci., U.S.A. 79, 7900 7903, 1982. FINLEY, W. W., FAUX, S. F., HUTCHESON, J. and AMSTUTZ, L. Long-latency event-related potentials in the evaluation of cognitive function in children. Neural. 35. 323 -327. 1985. GOODIN, D., SQUIRES, K., HENDERSON, B. and STARR. A. Age-related variations in evoked potentials to auditory stimuli in normal human subjects. Elrcfroenceph. C/in. Neurophysiol. 44, 447478, 1978. HALGREN, E., SQUIRES, N., WILSON, C., ROHRBAUGH, J., BARR, T. and CRANDALL, P. Endogenous potentials generated in the human hippocampal formation and amygdala by infrequent events. Science 210, 803 -805, 1980. HOWARD, L. and POLICH, J. P300 latency and memory span development. Dec. Psychol. 21, 293-~289, 1985. KARIS, D., FABIAN], M. and DONC‘HIN, E. ‘P300’ and memory: individual differences in the von Restorff effect. Cbyn. Psychol. 16, 177-216, 1984. LAI. J. A., BROWN. W. S., MARSH, J. T. and LARUE, A. Covariation of P3 latency and mini-mental state scores in geriatric patients. Psychophysiology, 20, 455, 1983. LEWIS, E. G., DUSTMAN, R. E. and BECK, E. C. Evoked response similarity in monozygotic, dizygotic and unrelated individuals: a comprehensive study. Electroenceph. Clin. Neurophysiol. 23, 309-316, 1972. NEVILLE, H. J. and SCHMIDT, A. L. Event-related brain potentials in subiects at risk for alcoholism. In Proceedings ofthe NIAAA Workshop on Early Identifcation-qj’ Alcohol Abuse. N. C. CHANG and H. M. CHAO
(Editors), Monograph No. 17, pp. 228 239. US. Department of Health and Human Services, Washington, D.C., 1985. OKADA, Y. C., KAUFMAN, L. and WILLIAMSON. S. J. The hippocampal formation as a source of the slow endogenous potentials. Elrctroenceph. Clin. Netrrophysiol. 55, 417 426. 1983. PAPANICOLAOU, A. C., LORING, D. w., RAZ, N. and E~SENRERG, H. M. Relationship between stimulus intensity and the P300. Psychophysiology. 22, 326-329, 1985. PFEFFERBAUM, A., WENEGRAT, B. G., FORD. J. M., ROTH, W. T. and KOPELL, B. S. Clinical application ofthe P3 component of event-related potentials. II. Dementia, depression and schizophrenia. Electroenceph. Clin. Nrurophysiol. 59, 104~ 124. 1984. PI(.TON, T. W., STUSS, D. T., CHAMPAGNE, S. C. and NELSON, R. F. The effects of age on human event-related potentials. Psychoph~lsiology 21, 312 -326, 1984. POLICH, J. Normal variation of P3OO from auditory stimuli. Electroenceph. Clin. NeurophJsiol. 65, 236~240. POLICH, J. P300 latency reflects personal drinking historv. Psvchoohvsioloav 21. 592-593. 1984. POLISH, J. Attention, probability, and iask demands as determinants of Pjbo latency from auditory stimuli. Electroencrph. Clin. Nrurophysiol. 63, 251-259, 1986. POLICH. J. and BLOOM. F. E. P300 reflects the degree of cognitive decline from the residual effects of alcohol consumption m normals and individuals at risk for alcoholism-a preliminary report. Progress in Neurops_vchopharmacoloqy: Biological Psychiatry 10, 201-210, 1986.
24. POLI(.H, J., HOWAKI), L. and STARR, A. P300 latency correlates with digit span. Psychophysiology 20,665~669, 1983.
25. POLICH, J., HOWARD, L. and STARR, A. Effects of aging on the P300 component of the event-related potential from auditory stimulii: peak definition variation and measurement. J. Gerontol. 40, 721-726, 1985.

304 NOTE
26. POLICH, J., HOWARD, L. and STARR, A. Stimulus frequency and masking as determinants of P300 latency in event-related potentials from auditory stimuli. Biol. Psych&. 21, 309-318, 1985.
27. POLICH, J., EHLERS, C. L., OTIS, S., MANDELL, A. J. and BLOOM, F. E. P300 latency reflects the degree of cognitive decline in dementing illness. Electroenceph. C/in. Neurophysiol. 63, 138 144, 1986.
28. RUST, J. Genetic effects in the cortical auditory evoked potential: a twin study. Electroenceph. Chn. Neurophysiol. 39, 321L327, 1975.
29. SKLARE, D. A. and LYNN, G. E. Latency of the P3 event-related potential: normative aspects and within-subject variability. Electroenceph. C/in. Neurophysiol. 59, 420424, 1984.
30. SMITH, M. E., STAPLETON, J. M. and HALGREN, H. Human medial temporal lobe potentials in memory and language tasks. Electroenceph. C[in. Neurophysiol., 63, 145-t 59, 1986.
31. SQUIRES, N., GALBRAITH, G. and AIRE, C. Event-related potential assessment of sensory and cognitive deficits in the mentally retarded. In Human Evoked Potentials. D. LEHMANN and E. CALLAWAY (Editors), pp. 397413. Plenum Press, New York, 1979.
32. SURWILLO, W. W. Cortical evoked potentials in monozygotic twins and unrelated subjects: comparisons of exogenous and endogenous components. Behav. &net. 10, 201-209, 1980.











![A Study of Identical Twins Palmprints 7Therefore, identical twins have the same genetic expressions. The frequency of identical twins is about 0.4% across different populations [3]](https://img.pdfslide.net/doc/110x75/5f8521a5e957fd4bd8723534/a-study-of-identical-twins-palmprints-7-therefore-identical-twins-have-the-same.jpg)
![A Study of Identical Twins’ Palmprints for Personal ... · identical twins have the same DNA. The frequency of identical twins is about 0.4% across different populations [2]. Some](https://img.pdfslide.net/doc/110x75/5f03fe9e7e708231d40bcb63/a-study-of-identical-twinsa-palmprints-for-personal-identical-twins-have-the.jpg)
















