Embed Size (px)
Citation preview

ORIGINAL RESEARCH PAPER
Penicillium sp. mitigates Fusarium-induced biotic stressin sesame plants
Ramalingam Radhakrishnan • Suk-Bok Pae •
Kang-Bo Shim • In-Youl Baek
Received: 14 November 2012 / Accepted: 4 March 2013 / Published online: 12 March 2013
� Springer Science+Business Media Dordrecht 2013
Abstract Fusarium-infected sesame plants have sig-
nificantly higher contents of amino acids (Asp, Thr, Ser,
Asn, Glu, Gly, Ala, Val, Met, Ile, Leu, Tyr, Phe, Lys, His,
Try, Arg, and Pro), compared with their respective levels
in the healthy control. These higher levels of amino acids
induced by Fusarium infection were decreased when
Penicillium was co-inoculated with Fusarium. Compared
with the control, Fusarium-infected plants showed higher
contents of palmitic (8 %), stearic (8 %), oleic (7 %), and
linolenic acids (4 %), and lower contents of oil (4 %) and
linoleic acid (11 %). Co-inoculation with Penicillium
mitigated the Fusarium-induced changes in fatty acids.
The total chlorophyll content was lower in Fusarium- and
Penicillium-infected plants than in the healthy control.
The accumulation of carotenoids and c-amino butyric
acid in Fusarium-infected plants was slightly decreased
by co-inoculation with Penicillium. Sesamin and sesam-
olin contents were higher in Penicillium- and Fusarium-
infected plants than in the control.
Purpose of work: To clarify the mechanism of the
biocontrol effect of Penicillium against Fusarium by
evaluating changes in primary and secondary metab-
olite contents in sesame plants.
Keywords Amino acid � Fatty acids Fusarium �Penicillium � Sesame
Introduction
Sesame plants are affected by numerous pathogenic fungi,
the most common of which are Fusarium species (El-
Shazly et al. 1999). Phytopathogen infection can change
pigment contents and disrupt normal metabolism of
amino acids and fatty acids in plants (Whiteside et al.
2012; Xue et al. 2006; Zafari et al. 2012). Fatty acid
signaling plays an important role in plant defense against
pathogen attack. The plant hormones jasmonic acid and
salicylic acid are fatty-acid derivatives with roles in plant
responses leading to protection against pathogens (Cre-
elman and Mulpuri 2002). c-Aminobutyric acid (GABA),
a non-protein amino acid that is widely distributed in all
living organisms, is involved in the stress response and
also in carbon and nitrogen metabolism (Bouche and
Fromm 2004). Sesamin and sesamolin are unique
secondary metabolites in sesame that prevent oxidative
damage induced by pathogens (Jeng and Hou 2005).
Recently, several studies have focused on the use of
non-pathogenic organisms to prevent phytopathogen
infection. The results of such studies have suggested
that biological organisms could be an alternative to
chemical fungicides, and also provide new insights
into the development of creative management strate-
gies to suppress soil borne diseases (Choudhary 2011).
Previously, we observed the biocontrol activity of
R. Radhakrishnan � S.-B. Pae � K.-B. Shim (&) �I.-Y. Baek
Department of Functional Crop, National Institute of Crop
Science, RDA, Miryang 672-803, Republic of Korea
e-mail: [email protected]
123
Biotechnol Lett (2013) 35:1073–1078
DOI 10.1007/s10529-013-1177-7

Penicillium against Fusarium in a dual-culture system.
In the present study, we analyzed changes in primary
and secondary metabolism in sesame plants infected
with these two fungi. The amino acid, fatty acid, and
photosynthetic pigment compositions in diseased
plants were studied by Trepanier et al. (2005) and
Zafari et al. (2012). However, there are no reports on
the involvement of amino acids, fatty acids, and
lignans in plant defenses against Fusarium infection.
The objective of this study was to determine the effect
of co-inoculation with Penicillium and Fusarium on
sesame plants, based on analyses of amino acids, fatty
acids, photosynthetic pigments, lignans, and GABA.
Materials and methods
Plant growth and fungal treatments
Penicillium sp. NICS01 (accession no: JX481980)
isolated from the rhizosphere soil of peanut plants was
cultured in potato/dextrose broth (PDB) at 28 ± 2 �C.
Sesame (Sesamum indicum L. cv. 90 days) seeds were
surface-sterilized with 0.5 % sodium hypochlorite and
washed repeatedly with sterile distilled water. Surface-
sterilized seeds were sown in pots in sterilized yard
manure that had been pretreated with Penicillium sp. The
medium consisted of peat moss (13–18 % w/v), Perlite
(7–11 % w/v), coco-peat (63–68 % w/v), Zeolite (6–8 %
w/v), NH4? (*90 mg kg-1), NO3- (*205 mg kg-1),
P2O5 (*350 mg kg-1), and K2O (*100 mg kg-1). The
control plants were sown in medium treated with sterile
water. The control and Penicillium-inoculated plants
were grown in a greenhouse under a 16-h day/8-h night
cycle at 28/20 �C and 60 % relative humidity. Fifty days
after planting, 10 ml Fusarium culture (1.9 9 104 c.f.u./
ml) was applied to control plants and Penicillium-
inoculated plants under greenhouse conditions. The
experiment had a randomized block design with five
replications, each consisting of four plants per pot. The
aerial parts of the plants were harvested 15 days after first
appearance of wilting symptoms. The collected plants
were stored at -80 �C until analyses.
Amino acids, GABA, fatty acids, photosynthetic
pigments, and lignan analyses
Samples (50 mg) were dissolved in 1 ml H2O and
centrifuged at 13,0009g at 4 �C for 30 min. The
supernatant was filtered through a nylon 66 syringe
filter (0.452 lm) and 0.1 ml was then mixed with
0.9 ml lithium citrate-loading buffer (pH 2.2). Amino
acids and GABA were determined using a Biochrom
30 amino acid analyzer on a single lithium cation-
exchange resin column (4.6 9 200 mm), using nin-
hydrin as the color reactant. Chromatography data
were analyzed using Ezchrom E software. The con-
centrations of individual amino acids and GABA were
calculated by the same procedure by comparison with
a known concentration of a standard solution contain-
ing the amino acid. The proline content was deter-
mined as described by Bates et al. (1973).
Total lipids in plant samples were extracted by the
Soxhlet method using a Buchi B-811 extraction
system (Buchi Labotechnik, AG, Flawil, Switzerland).
Fatty acid methyl esters (FAMEs) were prepared from
total lipids by acid-catalyzed transesterification as
described by Christie (2003). Lipids (1 mg) were re-
suspended in 1 ml toluene, and then 2 ml methanolic
H2SO4 (1 % v/v) was added. The reaction mixture was
heated at 100 �C for 1 h, and 3 ml H2O with 2 ml
hexane was used to extract FAMEs. Fatty acids were
analyzed as FAMEs using a capillary gas chromato-
graph equipped with a HP-FFAP capillary column
(30 m 9 0.318 mm, I.D. 25 lm film). The oven was
kept at 150 �C for 1 min, and increased to 230 �C for
1 min, then to 240 �C for 2 min. Both the injector and
detector were held at 260 �C. N2 carrier gas was at
1 ml/min; the injection volume was 1 ll, and the split
ratio was 1:50. The percentage of fatty acids was
calculated from standard values of peak areas of
C16:0, C18:0, C18:1, C18:2, and C18:3 methyl esters.
To extract photosynthetic pigments, leaf material
was ground in 80 % (v/v) acetone. Chlorophylls and
carotenoids were estimated according to the methods
of Arnon (1949) and Lichtenthaler (1987), respec-
tively. For the lignan analysis, powered sample (1 g)
was extracted with methanol for 24 h and filtered
through a nylon syringe filter (0.45 lm). To identify
lignans, the filtered sample (0.7 ml) was separated
using HPLC equipped with Lichrospher 100 RP 18
column (5 lm) at 30 �C. The mobile phase was 60 %
methanol (0.1 % trifluoroacetic acid) at 0.8 ml/min.
Lignans were detected at 361 nm. The sesamin and
sesamolin contents of Fusarium- and Penicillium-
treated and control samples were calculated by
comparisons with known concentrations of their
standards.
1074 Biotechnol Lett (2013) 35:1073–1078
123
Statistical analysis
The effects of the fungal treatments on primary and
secondary metabolite contents in sesame plants were
analyzed by comparison of means according to
Duncan’s multiple-range test (DMRT) at a signifi-
cance level of P \ 0.05.
Results and discussion
Biological control of phytophathogen infection in
plants is an effective and environmental friendly
method for sustainable management of soil-borne
plant diseases. In the present study, sesame plant
tissues contained detectable levels of 18 amino acids
(Asp, Thr, Ser, Asn, Glu, Gly, Ala, Val, Met, Ile, Leu,
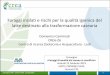
Tyr, Phe, Lys, His, Try, Arg, and Pro; Fig. 1). Among
these, Asp, Glu, and Arg were most abundant in both
infected and uninfected plants. Mycorrhizal coloniza-
tion strongly influences amino acid uptake by plants
(Whiteside et al. 2012). In the present study, Fusarium
infection significantly increased the concentrations of
18 amino acids in sesame plants compared with their
respective concentrations in the control. However, co-
inoculation of Penicillium with Fusarium inhibited
plant damage by mitigating the increase in amino acid
levels (Fig. 1). In plants, higher concentrations of free
amino acids under stress conditions are concomitant
with symptoms of damage (Silveira et al. 2001). The
mechanisms responsible for biotic stress-mediated
regulation of individual amino acids are still poorly
understood. However, asparagine plays a role in
certain physiological changes, including the accumu-
lation of secondary metabolites under adverse condi-
tions (Jia et al. 2001). The activities of some enzymes
involved in amino acid metabolism change after
pathogen infection. For example, aspartate amino-
transferase (AspAT), which can catalyze the bidirec-
tional conversion of Asn into Gln, was expressed
differently after infection by Botrytis cinerea, Phy-
tophthora infestans, and Pseudomonas syringae (Hruz
et al. 2008). A previous study hypothesized that
glutamate consumption pathways were inhibited by
the increased activity of AspAT to maintain normal
glutamate levels. In addition, glutamate might be
converted to other stress-related amino acids such as
proline, GABA, and arginine (Brauc et al. 2011).
All samples contained lower concentrations of Gly
than Ser. But both Gly and Ser were higher in
Fusarium-infected plants than in the control (Fig. 1).
Brauc et al. (2011) reported that a decrease in Pro
content in infected plants was associated with greater
susceptibility to B. cinerea infection. In contrast,
Arabidopsis plants infected with P. syringae showed a
higher Pro content than that in uninfected controls
(Fabro et al. 2004). We also observed a significantly
elevated Pro content in Fusarium-infected sesame
plants, compared with that in the control (Fig. 1).
Quantitative and qualitative changes in fatty acids
modulate the plant response to a variety of biotic and
abiotic stresses. In sesame plants, the oil and fatty
Fig. 1 Changes in amino acid contents in sesame plants infected with Fusarium and Penicillium. Bars represent means plus standard
error (n = 3). Means followed by the same letter are not significantly different (P \ 0.05) as determined by DMRT
Biotechnol Lett (2013) 35:1073–1078 1075
123
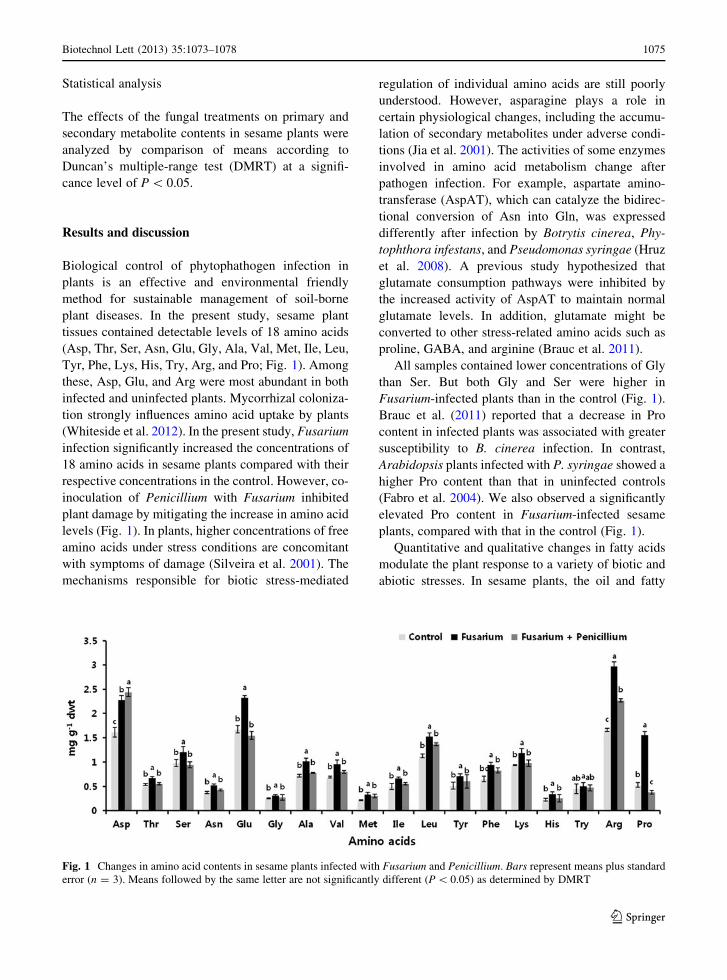
acids contents changed in response to fungal infection
(Fig. 2). Those infected with Fusarium had lower oil
and C18:2 contents, while co-inoculation with Peni-
cillium treatment alleviated these stress effects by
increasing oil and C18:2 contents (Fig. 2). Fusarium-
infected plants showed increased concentrations of
C16:0, C18:0, C18:1, and C18:3 compared with their
respective levels in the control, while co-inoculation
with Penicillium with Fusarium significantly
decreased the levels of these fatty acids (Fig. 2).
There have been few studies on changes in fatty
acid metabolism in plants under biotic stress. Different
plant species show different patterns of changes in
fatty acids in response to various stresses. For
example, the endophytic fungus Paecilomyces formo-
sus positively influenced plant growth and physiolog-
ical characters of cucumber, and infected plants
showed decreased C16:0 content (Khan et al. 2012).
Xue et al. (2006) reported increased C18:0 content in
soybean seeds infected by the seed-borne pathogen
Diaporthe phaseolorum. Fungal seed colonization was
related to the oleic (C18:1) and linoleic (C18:2) acid
contents. Higher linoleic acid and lower oleic acid
contents in soybean increased the seed colonization of
D. phaseolorum. Similarly, higher levels of oleic acid
promoted the Cercospora kikuchii association with
soybean seeds. In plant–pathogen interactions, the
ratio of oleic and linoleic acids differs depending on
the plant and the causal agent (Xue et al. 2006). Our
results showed that Penicillium co-infection increased
the levels of C18:2 and oil, compared with the levels in
plants infected with only Fusarium. The regulation of
other saturated and unsaturated fatty acids could be
due to the protective effect of Penicillium against
Fusarium infection.
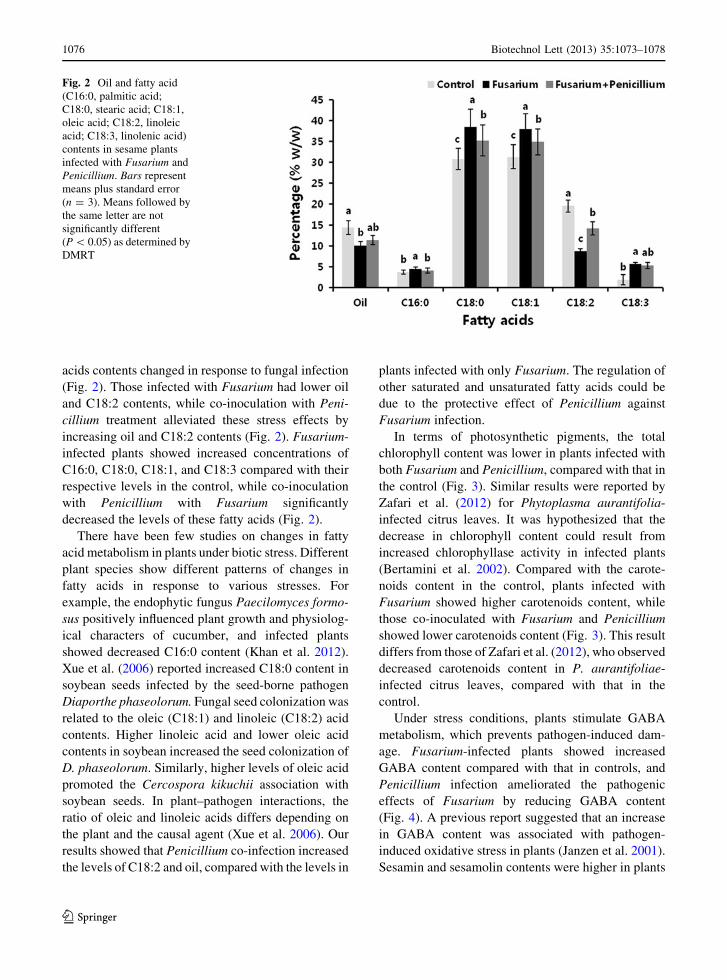
In terms of photosynthetic pigments, the total
chlorophyll content was lower in plants infected with
both Fusarium and Penicillium, compared with that in
the control (Fig. 3). Similar results were reported by
Zafari et al. (2012) for Phytoplasma aurantifolia-
infected citrus leaves. It was hypothesized that the
decrease in chlorophyll content could result from
increased chlorophyllase activity in infected plants
(Bertamini et al. 2002). Compared with the carote-
noids content in the control, plants infected with
Fusarium showed higher carotenoids content, while
those co-inoculated with Fusarium and Penicillium
showed lower carotenoids content (Fig. 3). This result
differs from those of Zafari et al. (2012), who observed
decreased carotenoids content in P. aurantifoliae-
infected citrus leaves, compared with that in the
control.
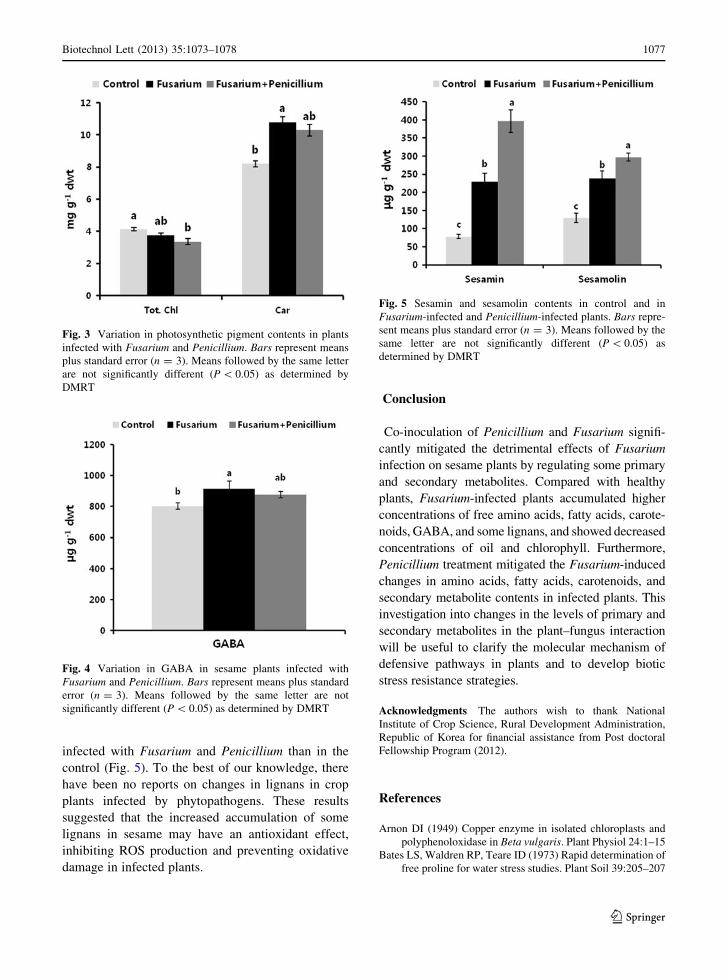
Under stress conditions, plants stimulate GABA
metabolism, which prevents pathogen-induced dam-
age. Fusarium-infected plants showed increased
GABA content compared with that in controls, and
Penicillium infection ameliorated the pathogenic
effects of Fusarium by reducing GABA content
(Fig. 4). A previous report suggested that an increase
in GABA content was associated with pathogen-
induced oxidative stress in plants (Janzen et al. 2001).
Sesamin and sesamolin contents were higher in plants
Fig. 2 Oil and fatty acid
(C16:0, palmitic acid;
C18:0, stearic acid; C18:1,
oleic acid; C18:2, linoleic
acid; C18:3, linolenic acid)
contents in sesame plants
infected with Fusarium and
Penicillium. Bars represent
means plus standard error
(n = 3). Means followed by
the same letter are not
significantly different
(P \ 0.05) as determined by
DMRT
1076 Biotechnol Lett (2013) 35:1073–1078
123
infected with Fusarium and Penicillium than in the
control (Fig. 5). To the best of our knowledge, there
have been no reports on changes in lignans in crop
plants infected by phytopathogens. These results
suggested that the increased accumulation of some
lignans in sesame may have an antioxidant effect,
inhibiting ROS production and preventing oxidative
damage in infected plants.
Conclusion
Co-inoculation of Penicillium and Fusarium signifi-
cantly mitigated the detrimental effects of Fusarium
infection on sesame plants by regulating some primary
and secondary metabolites. Compared with healthy
plants, Fusarium-infected plants accumulated higher
concentrations of free amino acids, fatty acids, carote-
noids, GABA, and some lignans, and showed decreased
concentrations of oil and chlorophyll. Furthermore,
Penicillium treatment mitigated the Fusarium-induced
changes in amino acids, fatty acids, carotenoids, and
secondary metabolite contents in infected plants. This
investigation into changes in the levels of primary and
secondary metabolites in the plant–fungus interaction
will be useful to clarify the molecular mechanism of
defensive pathways in plants and to develop biotic
stress resistance strategies.
Acknowledgments The authors wish to thank National
Institute of Crop Science, Rural Development Administration,
Republic of Korea for financial assistance from Post doctoral
Fellowship Program (2012).
References
Arnon DI (1949) Copper enzyme in isolated chloroplasts and
polyphenoloxidase in Beta vulgaris. Plant Physiol 24:1–15
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of
free proline for water stress studies. Plant Soil 39:205–207
Fig. 3 Variation in photosynthetic pigment contents in plants
infected with Fusarium and Penicillium. Bars represent means
plus standard error (n = 3). Means followed by the same letter
are not significantly different (P \ 0.05) as determined by
DMRT
Fig. 4 Variation in GABA in sesame plants infected with
Fusarium and Penicillium. Bars represent means plus standard
error (n = 3). Means followed by the same letter are not
significantly different (P \ 0.05) as determined by DMRT
Fig. 5 Sesamin and sesamolin contents in control and in
Fusarium-infected and Penicillium-infected plants. Bars repre-
sent means plus standard error (n = 3). Means followed by the
same letter are not significantly different (P \ 0.05) as
determined by DMRT
Biotechnol Lett (2013) 35:1073–1078 1077
123

Bertamini M, Nedunchezhian N, Tomasi F, Grando S (2002)
Phytoplasma [Stolbur subgroup (Bois Noir-BN)] infection
inhibits photosynthetic pigments, ribulose-1, 5-biphos-
phate carboxylase and photosynthetic activities in field
grown grapevine (Vitis vinifera L. cv. Chardonnay) leaves.
Physiol Mol Plant Pathol 61:357–366
Bouche N, Fromm H (2004) GABA in plants: just a metabolite?
Trends Plant Sci 9:110–115
Brauc S, Vooght ED, Claeys M, Hofte M, Angenon G (2011)
Influence of over-expression of cytosolic aspartate ami-
notransferase on amino acid metabolism and defense
responses against Botrytis cinerea infection in Arabidopsisthaliana. J Plant Physiol 168:1813–1819
Choudhary DK (2011) Plant growth promotion (PGP) activities
and molecular characterization of rhizobacterial strains
isolated from soybean (Glycine max L. Merril) plants
against charcoal rot pathogen, Macrophomina phaseolina.
Biotechnol Lett 33:2287–2295
Christie WW (2003) Preparation of derivates of fatty acids.
Lipid analysis: isolation, separation and structural analysis
of lipids, 3rd edn. PJ Barnes and Associates, Bridgwater,
pp 205–225
Creelman RA, Mulpuri R (2002) The oxylipin pathway in
Arabidopsis. The Arabidopsis book. American Society of
Plant Biologists, Rockville, pp 1–24
El-Shazly MS, Abdul-Wahid OA, El-Ashry MA, Ammar SM,
El-Barmawy MA (1999) Evaluation of resistance to
Fusarium wilt disease in sesame germplasm. Int J Pest
Manag 45:207–210
Fabro G, Kovacs I, Pavet V, Szabados L, Alvarez ME (2004)
Proline accumulation and AtP5CS2 gene activation are
induced by plant-pathogen incompatible interactions in
Arabidopsis. Mol Plant Microbe Interact 17:343–350
Hruz T, Laule O, Szabo G, Wessendorp F, Bleuler S, Oertle
Widmayer P, Gruissem W, Zimmermann PL (2008)
Genevestigator V3: a reference expression database for the
meta-analysis of transcriptomes. Adv Bioinformatics. doi:
10.1155/2008/420747
Janzen DJ, Allen LJ, Macgregor KB, Bown AW (2001) Cyto-
solic acidification and c-aminobutyric acid synthesis dur-
ing the oxidative burst in isolated Asparagus sprengerimesophyll cells. Can J Bot 79:438–443
Jeng KCG, Hou RCW (2005) Sesamin and Sesamolin: nature’s
therapeutic lignans. Curr Enzym Inhib 1:11–20
Jia M, Keutgen N, Matsuhashi S, Mitzuniwa C, Ito T, Fujimura
T, Hashimoto S (2001) Ion chromatographic analysis of
selected free amino acids and cations to investigate the
change of nitrogen metabolism by herbicide stress in soy-
bean (Glycine max). J Agric Food Chem 49:276–280
Khan AL, Hamayun M, Radhakrishnan R, Waqas M, Kang SM,
Kim YH, Shin JH, Choo YS, Kim JG, Lee IJ (2012)
Mutualistic association of Paecilomyces formosus LHL10
offers thermotolerance to Cucumis sativus. Antonie Van
Leeuwenhoek 101:267–279
Lichtenthaler HK (1987) Chlorophylls and carotenoids: pigment
of photosynthetic biomembranes. Method Enzymol 148:
350–382
Silveira JAG, Melo ARB, Viegas RA, Oliveira JTA (2001)
Salinity-induced effects on nitrogen assimilation related to
growth in cowpea plants. Environ Exp Bot 46:171–179
Trepanier M, Becard G, Moutoglis P, Willemot C, Gagne S,
Avis TJ, Rioux JA (2005) Dependence of arbuscular-
mycorrhizal fungi on their plant host for palmitic acid
synthesis. Appl Environ Microbiol 71:5341–5347
Whiteside MD, Garcia MO, Treseder KK (2012) Amino acid
uptake in arbuscular mycorrhizal plants. PLoS One 7(10):
e47643
Xue HQ, Upchurch RG, Kwanyuen P (2006) Ergosterol as a
quantifiable biomass marker for Diaporthe phaseolorumand Cercospora kikuchii. Plant Dis 90:1395–1398
Zafari S, Niknam V, Musetti R, Noorbakhsh SN (2012) Effect of
phytoplasma infection on metabolite content and antioxi-
dant enzyme activity in lime (Citrus aurantifolia). Acta
Physiol Plant 34:561–568
1078 Biotechnol Lett (2013) 35:1073–1078
123