Embed Size (px)
Citation preview
212 J. H. TALLIS: [J.L.s.B. LVI
Periodicity of growth in Rhacomitr ium lanuginosum. By J. H. TALLIS. Department of Botany, University College of Wales, Aberystwythl
(With 7 Text-figures)
The remarks I am about to make refer nominally only to a single moss species, Rhuco- mitrium lanuginosum (Hedw.) Brid., but they can be taken as applying in part probably to most mosses. However, certain features of growth are seen at their simplest in Rhaco- mitrium, and an uncritical extension of the conclusions reached to other mosses must be undertaken with care.
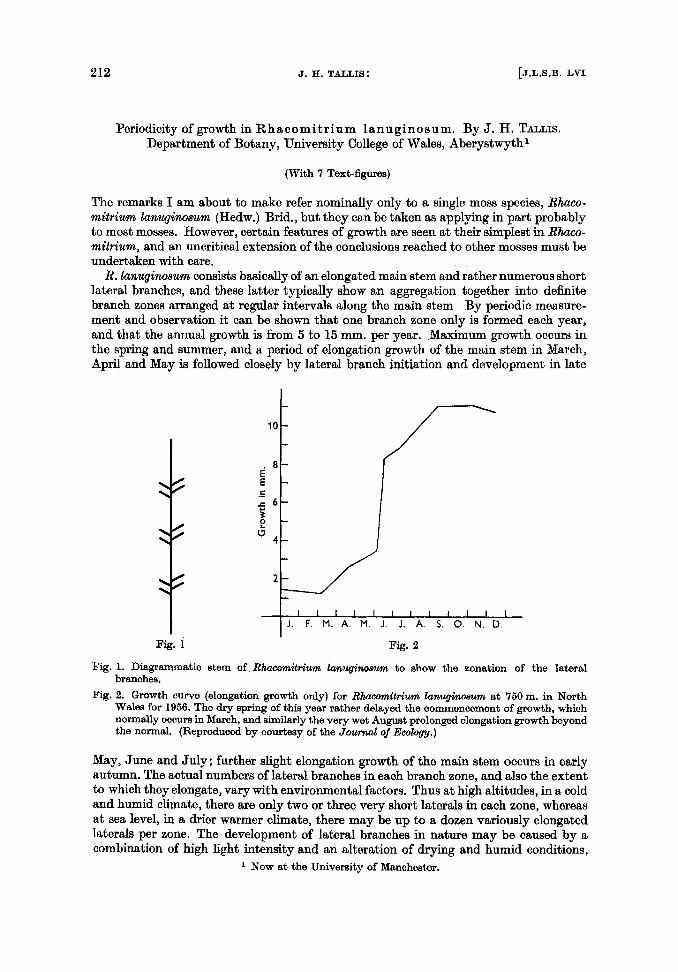
R. lanuginosum consists basically of an elongated main stem and rather numerous short lateral branches, and these latter typically show an aggregation together into definite branch zones arranged at regular intervals along the main stem By periodic measure- ment and observation it can be shown that one branch zone only is formed each year, and that the annual growth is from 5 to 15 mm. per year. Maximum growth occurs in the spring and summer, and a period of elongation growth of the main stem in March, April and May is followed closely by lateral branch initiation and development in late
1 1 1 1 1 1 1 1 1 1 1 1 J . F . M . A . M . J . J . A . S . O . N . D .
Pig. 1 Fig. 2
Fig. 1. Diagrammatic stem of Rfracomitrium lanllgimmvn to show the zonation of the lateral branches.
Fig. 2. Growth cullre (elongation growth only) for Rhacomktrium lanugimwm at 750m. in North Wales for 1956. The dry spring of this year rather delayed the commencement of growth, which normally occurs in March, and similarly the very wet August prolonged elongation growth beyond the normal. (Reproduced by courtesy of the Journal of Ecobgy.)
May, June and July; further slight elongation growth of the main stem occurs in early autumn. The actual numbers of lateral branches in each branch zone, and also the extent to which they elongate, vary with environmental factors. Thus at high altitudes, in a cold and humid climate, there are only two or three very short laterals in each zone, whereas at sea level, in a drier warmer climate, there may be up to a dozen variously elongated laterals per zone. The development of lateral branches in nature may be caused by a combination of high light intensity and an alteration of drying and humid conditions,
Now at the University of Manchester.

J.L.S.B. LVI] GROWTH IN RHACOMITRIUM LANUGINOSUM 2 13
perhaps with the mean temperature above a certain threshold value, of about 12-14" C. This is suggested chiefly by a comparison of the growth curve with the relevant climatic data. In culture (which is difficult, and has so far been possible only for relatively short periods) under humid shaded conditions, the initiation of lateral branches can be delayed by up to 4 weeks, but apparently not indefinitely. When the stem apex is removed, and the lateral branch next below takes over the function of the main stem (giving sympodial growth), or when regeneration occurs from stem fragments, lateral branches are not formed until after 2-3 years) elongation growth has taken place. However, where, under certain favourable conditions, elongation of certain of the lateral branches occurs in nature in the year subsequent to initiation, lateral branches are formed on these as on the main stem. These results would suggest, perhaps, some active principle formed at the main apex and affecting growth at the proximate lateral apices. In other mosses (e.g. Eurhynchium striaturn (Hedw.) Schp. emend Starmer) it appears from field observa- tions that high humidity and shading may completely suppress lateral branch formation for up to a year.
Fig. 3. Progressively maturing leaves of Rhacmitrium lanugimgzlm, showing the preoooious development of the hair-point.
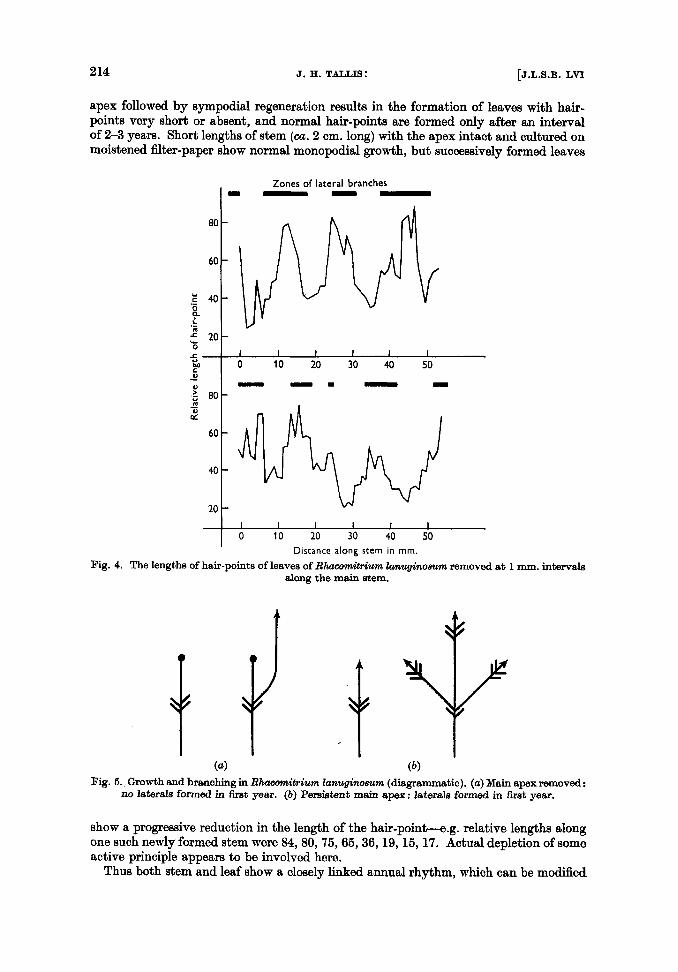
The leaf of R. languinosum consists of a longly triangular green lamina terminating in a hyaline hair-point which may be nearly as long as the lamina. The hair-point is formed early in leaf development, and reaches maturity while the lamina is still relatively small. Hence it should reflect fairly closely the conditions at the growing apex. If leaves are removed at regular short intervals along the main stem and the lengths of the hair- points measured, a cyclical fluctuation in length of the hair-point is found to occur: the peaks occur consistently in the lateral branch zones, whilst leaves along the inter- zone portions of the main stem have rather short hair-points. A similar cyclic fluctuation has been claimed for the length of the green lamina, but I have not as yet been able to confirm this. However, a cyclical fluctuation of lamina length or of the lengthlbreadth ratio does occur in other mosses. As with the lateral branches, length of the hair-point is determined both by internal and by external factors. I am using these two words in a very loose sense, and in the ultimate analysis they may well be found to be inseparable. Light and humidity are probably the most important external factors, but it is difficult to separate their effects: thus plants grown in shade or in very humid conditions have hair-points shortened to 50 % of normal or rarely even almost entirely wanting. I n the summer of 1956 in North Wales at high altitudes (when the weather w a ~ consistently cold, wet, and dull), hair-points formed during that period were reduced to 65 % of nor- mal. However, only rarely do external factors reduce the length of the hair-point to values similar to those at the minima of the cyclic fluctuations. Removal of the stem
2 14 J. H. TALLIS: [J.L.s.B. LVI
20- 0
5 F
2 80-
2
- 0
m - 60
40
20
apex followed by sympodial regeneration results in the formation of leaves with hair- points very short or absent, and normal hair-points are formed only after an interval of 2-3 years. Short lengths of stem (ca. 2 cm. long) with the apex intact and cultured on moistened filter-paper show normal monopodial growth, but successively formed leaves
I I I I I I
0 10 20 30 40 50 - - I - I -
-
- I I I I 1 I 0 10 20 30 40 50
Distance along stem in mm.
Zones of lateral branches I - - - -
i P
(a) (b ) Fig. 6 . , Growth and branching in R k m i t r i u m knuqimwm (diagrammatic). (a) Main apex removed:
no laterals formed in first year. (b) Persiatent main apex : laterals formed in first year.
show a progressive reduction in the length of the hair-p0int-e.g. relative lengths along one such newly formed stem were 84,80,75,65,36,19,16,17. Actual depletion of some active principle appears to be involved here.
Thus both stem and leaf show a closely linked annual rhythm, which can be modified

J.L.S.B. LVI] OROWTH IN REACOMITRIUM LANUOINOSUM 2 15
by external factors to some extent ; however, there is in addition an underlying internal control (with the proviso mentioned above) associated apparently with a substance or substances produced at the stem apex and passing into the newly formedlateral branches; this substance appears to be produced chiefly when elongation growth of the main stem is slow, but when conditions for assimilation are favourable (maximum assimilation occurs experimentally at 12-15' C.), and its presence results in lateral branch formation and well-developed hair-points to the leaves. Alternatively, these effects could be associated with the absence of a substance which is produced only during active elonga- tion growth under relatively unfavourable conditions. The evidence so far does not per- mit of a definite conclusion either way.
J
Fig. 6. Growth types in pleurocarpous mosses. On the left two stems of Hypnum cupessiforme Hedw. var. ericetomm B. & S . to show zonation of lateral branches; in the centre, portion of the stem of HybcorniumspZendem (Hedw.) B. & S. to show regular arrangement of lateral branches of differing lengths; and on the right, one stem of RhytidiadeZphua triquetrua (Hedw.) Warnst. showing 'irregularly' arranged laterals. The arrows indicate the presumed positions of successive annual increments; the lack of lateral branches along one increment is the result of submergence of the stem during this period below other stems.
A similar annual rhythm, affecting stem or leaf or both, probably occurs in most mosses. This phenomenon of periodicity is well known in acrocarpous mosses from the studies of Hagerup (1935), Lackner (1939), and Jendralski (1955). Here apical growth is ter- minated by the formation of sex organs, and growth is contained by a branch arising a short distance below the apex. Even when sex organs are not formed, annual increments can generally be recognized by seasonal differences in the size and shape of leaves. Periodicity has been less studied in pleurocarpous mosses, where the sex organs develop on lateral branches and where, consequently, any rhythms are shown chiefly in the dis- position and length of the lateral branches. The definite zonation of laterals seen in R. lanuginosum occurs in several other mosses (though in a less distinct form) such as Hypnum cupressiforme Hedw. , Acrocladium cuspidatwm (Hedw.) Lindb. and Campto- thecium lutescens (Hedw.) Brid. Several pleurocarpous mosses have lateral branches regularly arranged along the whole length of the main stem, but typically these laterals are of differing lengths, such that each annual increment has an approximately elliptical outline. Hylocmium splendem (Hedw.) B. & S. and Cratoneuron cmmututum (Hedw.) Roth are good examples. Again, several pleurocarpous mosses, such as Rhytidiadelphus, appear to have an irregular distribution of laterals along the main stem; however, usually it can be shown that the distribution of the laterals is not statistically random,
2 16 J. R. TALLIS: [J.L.S.B. LVI
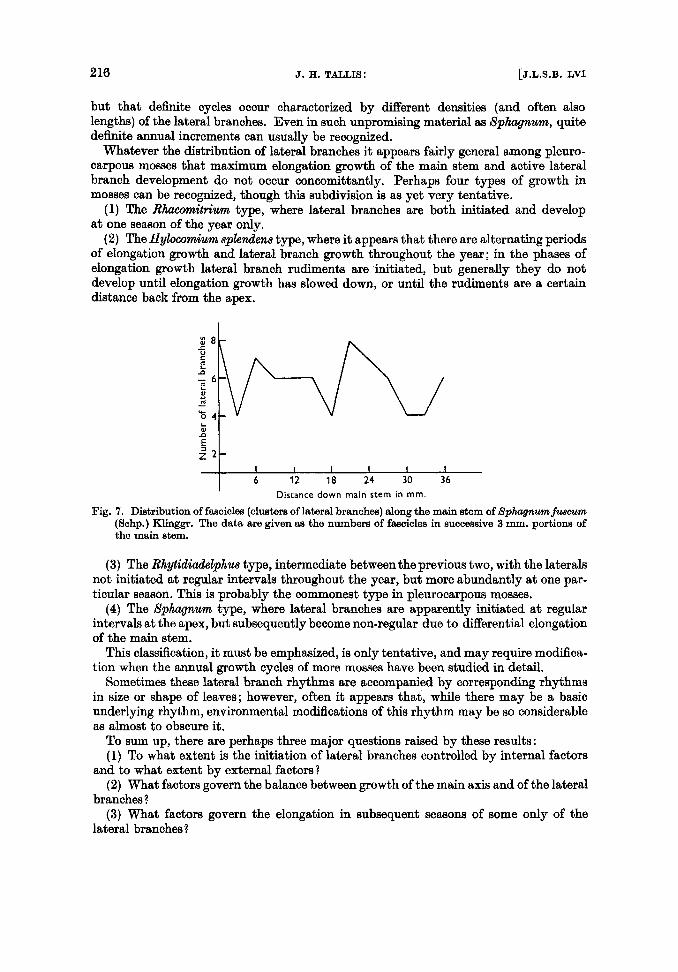
but that definite cycles occur characterized by different densities (and often also lengths) of the lateral branches. Even in such unpromising material as Sphagnum, quite definite annual increments can usually be recognized.
Whatever the distribution of lateral branches it appears fairly general among pleuro- carpous mosses that maximum elongation growth of the main stem and active lateral branch development do not occur concomittantly. Perhaps four types of growth in mosses can be recognized, though this subdivision is w yet very tentative.
(1) The Rhacmnitrium type, where lateral branches are both initiated and develop at one sewon of the year only.
( 2 ) The Hylocomium splendens type, where it appears that there are alternating periods of elongation growth and lateral branch growth throughout the year; in the phases of elongation growth lateral branch rudiments are initiated, but generally they do not develop until elongation growth has slowed down, or until the rudiments are a certain distance back from the apex.
I I I I I I I 6 12 18 24 30 36
Distance down main stern in rnrn.
Fig. 7. Distribution of fascicles (clusters of lateral branches) along the main stem of Sphagnurnfusoum (Schp.) Klinggr. The data are given as the numbers of fascicles in successive 3 mm. portions of the main stem.
(3) The RhytidiadeLphus type, intermediate between the previous two, with the laterals not initiated at regular intervals throughout the year, but more abundantly at one par- ticular season. This is probably the commonest type in pleurocarpous mosses.
(4) The Sphagnum type, where lateral branches are apparently initiated at regular intervals at the apex, but subsequently become non-regular due to differential elongation of the main stem.
This classification, it must be emphasized, is only tentative, and may require modifica- tion when the annual growth cycles of more mosses have been studied in detail.
Sometimes these lateral branch rhythms are accompanied by corresponding rhythms in size or shape of leaves; however, often it appears that, while there may be a basic underlying rhythm, environmental modifications of this rhythm may be so considerable as almost to obscure it.
To sum up, there are perhaps three major questions raised by these results : (1) To what extent is the initiation of lateral branches controlled by internal factors
( 2 ) What factors govern the balance between growth of the main axis and ofthe lateral
(3) What factors govern the elongation in subsequent seasons of some only of the
and to what extent by external factors?
branches ?
lateral branches?

J.L.S.B. LVI] GROWTH IN RHACOMI!L%ITJM LANUGINOSUM 217
REFERENCES
Further details of the Rhcomitrium data will be found in:
TALLIS, J. H. (1957). A study of the biology and ecology of Rhawminitrium lanuginosum Brid., Ph.D.
TALLIS, J. H. (1959). Studies in the biology and ecology of Rhacomitrium ~ n u g i n o s u ~ Brid. 11. thesis, University of Wales. (Unpublished.)
Growth, reproduction, and physiology. J. Ecol. 47, 325-360.
For details of periodicity in mosses in general, consult:
HAGERUP, 0. (1935). Zur Periodizitat im Laubwechsel der Moose. K . Damke Videmk Selsk. Biol.
JENDRALSPI, U. (1955). Die Jahresperiodizitiit in der Entwicklung der Laubmoose im Rheinlasde.
LACPNER, L. (1939). fjber die Jahresperiodizitiit in der Entwicklung der Laubmoose. Planta, 29,
M a . 11.
Decheniana, 108, 105-163
534-616.
10 JOURN. LINN. S0C.-BOTANY, VOL. LVI





























