Embed Size (px)
Citation preview

Personality Stability in Late Adulthood: A
Behavioral Genetic Analysis
Wendy Johnson, Matt McGue, and Robert F. Krueger
University of Minnesota–Twin Cities
ABSTRACT A sample of 833 twins from the Minnesota Twin Studyof Adult Development and Aging completed the MultidimensionalPersonality Questionnaire (MPQ; Tellegen, 1982) twice, averaging 59.4(sd5 9.7) years of age at first and 64.4 (sd5 10.2) years of age at secondtesting (average retest interval 5.0 years, sd5 2.36, range 1.0–10.4 years).Both means and standard deviations of scale scores were extremely stablefrom first to second testing. In addition, sample participants tended toretain their rank order on the scales (average r5 .76 across scales).Bivariate biometric analyses showed that the genetic influences on mostof the scale scores were almost perfectly correlated across the two waves(range .95 to 1.00). The nonshared environmental influences were alsohighly correlated across the two waves (range .53 to .73). Models spec-ifying identical variance components at the two time points and fixing thegenetic correlation to 1.00 provided improved fit. The results suggest thatthe high stability of personality in later adulthood has a strong geneticfoundation, supplemented by stability of environmental effects.
A substantial body of evidence has accumulated for the stability ofpersonality in adulthood. Such evidence is especially strong for theconsistency of an individual’s relative position in a group (Roberts &DelVecchio, 2000). Consistency of this type, known as differential
The Minnesota Twin Study of Adult Development and Aging was supported by
National Institute of Aging Grant #AG 06886. Robert F. Krueger was supported by
National Institute of Mental Health Grant #MH 65137.
Correspondence concerning this article should be addressed to Wendy Johnson, De-
partment of Psychology, University of Minnesota, 75 East River Road, Minneapolis,
MN 55455. E-mail [email protected].
Journal of Personality 73:2, April 2005r Blackwell Publishing 2005DOI: 10.1111/j.1467-6494.2005.00319.x

stability (Caspi & Roberts, 2001), is generally measured by a corre-lation coefficient. Such coefficients have been estimated for a varietyof self-report personality questionnaires, ages, and retest intervals(e.g. Lubinski, Schmidt, & Benbow, 1996; McCrae & Costa, 1990;McGue, Bacon, & Lykken, 1993), but also for personality ratings byclinicians, spouses, and acquaintances (e.g. Conley, 1985; Costa &McCrae, 1988). In addition, there is strong evidence for the con-stancy of absolute levels of personality measures over time (Caspi &Roberts, 2001; Costa & McCrae, 1988). Roberts and DelVecchio(2000) concluded that the evidence to date suggests that relative sta-bility of personality over a period of several years increases with age,with correlations going from .31 in childhood to .64 around age 30,and leveling off at .74 for the ages 50 to 70.
Stability of personality also decreases as the time interval betweenobservations increases, yet impressive levels of stability have beenobserved over lengthy time spans in adulthood. Though 30-day re-test reliabilities for well-constructed scales can range from .80 to .90(reflecting the measurement capacity of the instrument; Tellegen,1982), retest stabilities across spans on the order of 25 years can re-main in the range of .60 to .70 (Costa & McCrae, 1992b; Helson &Klohnen, 1998), with higher stabilities (on the order of .65 to .80)reported for shorter time intervals of 6 to 12 years (Costa &McCrae,1988, 1992b; Viken, Rose, & Koskenvuo, 1994). For adults underage 30, comparable stabilities for 6- to 10-year time intervals are onlyslightly lower, ranging from .48 to .60 (McGue et al., 1993; Vikenet al., 1994). The particular aspect of personality measured appearsto have little effect on the extent of stability, nor do sex or method ofmeasurement (Roberts & DelVecchio, 2000).
That genes contribute to personality throughout the lifespan isalso very well established. Loehlin and Nichols’s (1976) study of al-most 800 pairs of adolescent twins and dozens of personality traitshas stood the test of time, having been replicated many times using avariety of personality measurements and sample populations across abroad range of ages. Bouchard and Loehlin (2001) summarized thefindings of 25 years of investigations as showing that 40–60% of in-dividual differences in personality can be attributed to genetic vari-ation. McCrae et al. (2000) have used this consistency of results acrosssamples and many age ranges, construed in light of temperament as aconstitutional predisposition to personality qualities, to argue thatpersonality traits, even at the facet level, are genetically controlled
524 Johnson, McGue, & Krueger

dispositions for which the environment plays no fundamental roleand merely sets the stage for expression. They maintain, therefore,that both continuity and change in personality have their sources ingenetic influences, with genetic influences on basic temperamentcontributing stability and genetic influences on developmental proc-esses contributing to change.
The concept of genetic influences on developmental processes isfamiliar when considering children and adolescents, for whom it isnot difficult to conceive of and even demonstrate psychologicalprocesses that parallel the clear physical manifestations of develop-ment. In considering adults, however, the concept is more foreign.Though adults, too, experience physical changes over time, we tendto think of the physical manifestations of aging as resulting fromgradual and passive decline in function rather than from the kinds ofwell-regulated and dynamic processes that govern change in youth.This may greatly oversimplify the actual situation, as there is grow-ing evidence, from fruit flies as well as mammals, that many of theorganism’s biological responses (manifested by genetic expression)to aging, at least, are very analogous to the sorts of developmentalprocesses that operate in youth (Helfand & Rogina, 2003). That re-sponses to physical aging may be under systematic genetic develop-mental control in such nonhuman organisms hints at a similar rolefor psychological mechanisms such as personality characteristics inhumans. For example, there is evidence for systematic personalitychange with age (Srivastava, John, Gosling, & Potter, 2003), with thegeneral trait of Big Five Extraversion showing some decrease andConscientiousness increasing. These patterns at the level of generaltraits may result from much more specific changes, such as the ob-servation of increased Harm Avoidance with age (Carmichael, 1993).It is possible that this increase could be a genetically controlledadaptive psychological response to increased physical fragility. Thus,McCrae et al.’s (2000) conception of genetic sources of change inadulthood is plausible, though it requires testing.
Roberts and Caspi (2003) have addressed the question of geneticand environmental influences on continuity and change in personalitysomewhat differently. Noting that personality stability is more com-mon than change in personality traits, they point out that, almost bydefinition, trait theories make no explanation for personality change.On the other hand, developmental theories focus primarily onchange, and they tend to present change as change in responses to
Personality Stability 525

circumstances rather than as changes in characteristics intrinsic tothe individual such as personality. To address the gap, they describethe ways in which genetic and environmental influences, psycholog-ical function, and habitual person-environment transactions cancontribute to stability. Very generally stated, these same processescan also contribute to change in the form of genetic influences ondevelopmental processes, responses to environmental circumstancesthat reflect genetic as well as environmental influences, observationallearning, learning generalizations, and learning from others’ descrip-tions of ourselves. They thus see both personality stability and changeas the results of an ongoing series of transactions between genetic andenvironmental influences, which both have elements of stability andchange. This conception also should be tested.
In behavior genetic studies, environmental influences are generallycategorized as either shared or nonshared. Shared environmentalinfluences are those that contribute to similarity among reared-to-gether relatives. Frequently given examples of such influences referto experiences of children, such as growing up with a common set ofparents who provide a certain level of socioeconomic status, valuessuch as religion, and common experiences such as travel, presence ofalcoholism, or provision of a college education. But the concept ofshared environmental influences can be extended to adults as well. Itis easy to conceive of these kinds of experiences as having lifelonginfluences, and there is evidence for the existence of such effects forsome of them. The process of measuring the magnitude of sharedenvironmental influences does nothing to identify the specific influ-ences operating, so conceptual plausibility of this sort is all that isnecessary to justify their quantification in this manner. Environmen-tal influences can be shared by siblings in adulthood as well. Forexample, siblings may continue to associate with each other fre-quently in adulthood, sharing holidays and care for elderly parents,among many other things. Thus, in adults, shared environmentalinfluences include both lifelong effects of shared childhood experi-ences and the effects of currently shared experiences. The concept ofshared environmental influence has had little effect on the concep-tion of personality emergence, stability, and change, however, be-cause most studies have shown little evidence of it for personality(Bouchard & Loehlin, 2001).
Nonshared environmental influences are those that contribute todifferences between siblings, and the same sort of discussion applies.
526 Johnson, McGue, & Krueger

In childhood, experiences such as having different schoolteachers,participating in different sports or other leisure activities, and dif-ferential parental treatment would qualify, and these experiencescould have effects that persist into adulthood. In addition, in adult-hood, siblings typically have many different experiences involvingspouses, jobs, and financial security (again among many otherthings). Nonshared environmental influences are rarely measureddirectly. Rather, they are measured as residual variance, whichmeans that the term in which they are included always includes ran-dom error of measurement and often includes systematic methodbias as well. Thus we would tend to expect relatively little relation-ship between nonshared environmental influences over time, due, inpart, to measurement error and, in part, to changes in nonsharedenvironmental circumstances over time. This means that one way toevaluate some of the implications of Roberts and Caspi’s (2003) andMcCrae et al.’s (2000) conceptions of stability and change in per-sonality is to use a longitudinal twin study.
When it is carried out in longitudinal form, the twin study methodthat serves as the cornerstone of behavioral genetic analysis can offerimportant contributions to our understanding of the nature of boththe observed stability of and change in personality over time. Inparticular, such studies can address the extent to which genetic andenvironmental influences change with time by estimating theamounts of variance attributable to genetic and environmental in-fluences at varying time points. They can also address the extent towhich genetic and environmental influences contribute to stabilityand change by estimating the extent to which the same genetic andenvironmental influences remain in place over time and new onescome into play. Loehlin (1992) reviewed longitudinal behavior ge-netic studies of variation in personality from childhood into adult-hood in order to summarize the evidence for various kinds of andchanges in genetic influence. Though most of the studies he revieweddid not address genetic and environmental sources of personalitystability and change directly, he concluded that although there aresignificant sources of genetic influence on personality change inchildhood, after that, genetic influences contribute primarily to per-sonality stability rather than to change. This conclusion may havebeen driven by the relative lack of change in adulthood to evaluate.
Genetic and environmental sources of stability of or change inpersonality in adulthood have been addressed specifically in just a
Personality Stability 527

few longitudinal studies. These studies have primarily focused on thetraits of Extraversion and Neuroticism as defined by Eysenck (Eys-enck & Eysenck, 1985). Viken et al. (1994) found that the same genescontributed to personality variance over time, based on a sample ofnearly 15,000 male and female Finnish twins ranging in age from 18to 53 at baseline and tested on average 6 years later. They also ob-served that heritabilities decreased with age, primarily resulting fromincreasing variance associated with nonshared environmental influ-ences at the two time points, and that the correlations between non-shared environmental influences at the two time points increasedwith age from about .30 at age 18 to about .55 at age 53. Pedersenand Reynolds (1998) also conducted a longitudinal study, involving2,209 participants of the Swedish Adoption/Twin Study of Agingwho averaged 60 years of age at baseline and reported over a 9-yearperiod. Like Viken, they concluded that the genetic influences onExtraversion, Neuroticism, and the Five-Factor Model trait ofOpenness were stable. Despite the stability of both mean levels ofthe personality traits and genetically influenced variance, they notedboth generally increasing nonshared environmental variance and in-creasing change with time. Neither study showed evidence of newgenetic contributions after age 30. Very similar results were obtainedfor Positive and Negative Emotionality and for Constraint from theMultidimensional Personality Questionnaire (MPQ; Tellegen, 1982)in a Minnesota sample of 127 younger twin pairs averaging 20 yearsof age at baseline and 30 at retest (McGue et al., 1993). In general,the data suggested higher levels of genetic influence in early adult-hood, declining to about 40% over the age 20–30 period, with littleevidence of new genetic contributions during the period observed.Similar results were also obtained by Angleitner and Spinath (2002)using Five-Factor Model NEO-PI-R (Costa & McCrae, 1992a) self-and peer reports in a German sample spanning a large adult agerange.
The studies discussed above have focused either on the very broadpersonality constructs of Extraversion and Neuroticism or on earlyadulthood. They have also focused primarily on the extent to whichgenetic factors contribute to personality stability and/or change.They have established evidence for a strong genetic foundation in-fluencing stability in these broad traits across the adult lifespan, butwe are not aware of any study that has investigated the extent towhich these findings apply to more specific personality traits in late
528 Johnson, McGue, & Krueger

adulthood, nor are we aware of any study that has investigated spe-cifically the question of environmental as well as genetic influenceson stability over time. It is important to examine the stability ofmore specific personality traits as well as the broad traits such asExtraversion and Neuroticism because these broad traits are man-ifested in various specific ways, and stability at the level of the broadtraits could mask change at the specific level. For example, bothsociability and adventurousness are often considered to be aspects ofExtraversion. A person, however, could change from being adven-turous but not particularly sociable to being sociable but not par-ticularly adventurous without changing in overall level ofExtraversion. It is important to estimate the extent of environmen-tal as well as genetic influences on stability of personality becausethey can help us to distinguish between McCrae et al.’s (2000) con-ception of both stability of and change in personality as primarilygenetic in origin and Roberts and Caspi’s (2003) perspective of alarger role for the environment in personality stability.
Thus, the purpose of the current study was to investigate the ex-tent and sources of stability of and change in personality as meas-ured by the primary trait scales of the MPQ in late adulthood. Wemade use of the MPQ because it was designed to measure 11 specificpersonality traits with no item overlap among its primary scales(Tellegen, 1982). It therefore measures specific aspects of personalitywith high fidelity. Nevertheless, the primary scales of the MPQ covera broad range of traits, showing good bandwidth. Thus, the MPQcombines fidelity and bandwidth in the measurement of personality,making it especially well suited to a comprehensive investigation ofthe genetic and environmental sources of personality stability.
The MPQ is a self-report personality inventory that was con-structed by an exploratory factor-analytic process with the goal ofdeveloping relatively pure indexes of trait dispositions within normalpersonality. The result was 11 relatively independent primary traitscales. In factor analyses, the primary scales load on three higher-order factors, Positive Emotionality, Negative Emotionality, andConstraint, which relate to neurobiological mechanisms and maponto constructs of emotion and temperament. That is, Positive andNegative Emotionality incorporate temperamental dispositions to-ward those two emotions and are linked to brain motive systemsunderlying the appetitive-approach and defensive-withdrawal be-haviors (Derryberry & Reed, 1994; Lang, 1995; Sutton & Davidson,
Personality Stability 529

1997). Constraint incorporates traits related to behavioral restraintand reversed impulsivity, which has been characterized in termsof low brain arousal and reduced conditionability (Zuckerman,1991). The primary MPQ scales have high internal consistency and30-day retest reliabilities, and they cover a range of relatively psy-chometrically pure traits encompassing temperament, behavioralregulation, and interpersonal and imaginative styles. The MPQ’scombination of high reliability, construct independence, broad cov-erage of the personality domain, and neurobiological referents makeit a particularly good choice for investigation of the extent andsources of personality stability. Table 1 briefly characterizes theMPQ scales.
Positive Emotionality reflects the tendency to be actively andpleasurably engaged with one’s social and work environments; it issimilar to the Extraversion dimension of the Eysenck PersonalityQuestionnaire (EPQ; Eysenck, 1960) and the Five-Factor Model(FFM; McCrae & Costa, 1997) and to the Ascendance factor of theCalifornia Psychological Inventory (CPI; Gough & Bradley, 1996).The primary scales loading on positive emotionality reflect agencyand communion; they are Wellbeing, Social Potency, Achievement,and Social Closeness. Negative Emotionality is characterized byperceptions of the world as threatening, problematic, and distress-ing; it is similar to Neuroticism as defined by many personalitymeasures. Its primary scales reflect neurosis, estrangement, andconfrontation; they are Stress Reactivity, Alienation, and Aggres-sion. Constraint is marked by the tendency to be restrained (impul-siveness reversed), conventional, and careful (sensation-seekingreversed). It is related to the Conscientiousness factor in theFFM (Church, 1994). The primary MPQ scales loading on Con-straint are Control, Harm Avoidance, and Traditionalism. The 11thprimary MPQ trait scale, Absorption, reflects emotional responsive-ness to sensory input and loads on both Positive and NegativeEmotionality.
Specifically, in this study, we addressed three issues in a longitu-dinal, population-based Minnesota sample of primarily older-adulttwins. First, we assessed the extent of personality stability andchange, given measurements at two time points. Second, we esti-mated the genetic and environmental influences on the personalityscale scores at both time points. Third, we investigated the extent towhich the same genetic and environmental influences were involved
530 Johnson, McGue, & Krueger
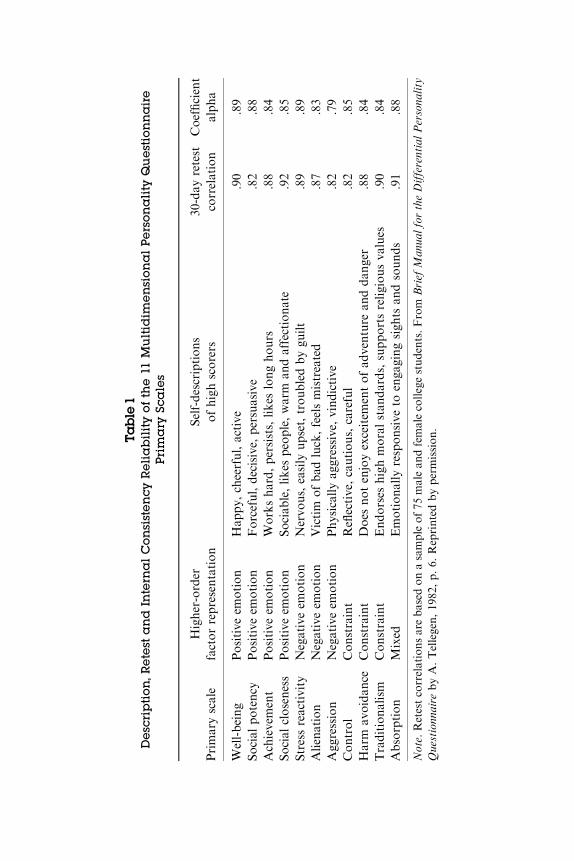
Table
1Description,R
etest
andInternalConsistencyReliabilityofthe11
Multidim
ensionalPersonality
Questionnaire
Primary
Scales
Primaryscale
Higher-order
factorrepresentation
Self-descriptions
ofhighscorers
30-day
retest
correlation
Coefficient
alpha
Well-being
Positive
emotion
Hap
py,
cheerful,active
.90
.89
Social
potency
Positive
emotion
Forceful,decisive,
persuasive
.82
.88
Achievement
Positive
emotion
Workshard,persists,likes
longhours
.88
.84
Social
closeness
Positive
emotion
Sociab
le,likes
people,warm
andaffectionate
.92
.85
Stressreactivity
Negativeem
otion
Nervo
us,easily
upset,troubledbygu
ilt
.89
.89
Alienation
Negativeem
otion
Victim
ofbad
luck,feelsmistreated
.87
.83
Aggression
Negativeem
otion
Physically
aggressive,vindictive
.82
.79
Control
Constraint
Reflective,cautious,careful
.82
.85
Harm
avoidan
ceConstraint
Does
notenjoyexceitem
entofad
venture
anddan
ger
.88
.84
Traditionalism
Constraint
Endorses
highmoralstan
dards,supportsreligiousvalues
.90
.84
Absorption
Mixed
Emotionally
responsive
toengagingsigh
tsan
dsounds
.91
.88
Note.Retestcorrelationsarebased
onasampleof75
malean
dfemalecollegestudents.From
Brief
Man
ualfortheDifferentialPersona
lity
Questionn
aire
byA.Tellegen,19
82,p.6.
Reprintedbypermission.

at the two time points. We expected considerable stability of per-sonality, as well as a substantial genetic contribution to stabilityacross time, as anticipated in the conceptions of the sources of per-sonality stability and change of both McCrae et al. (2000) andRoberts and Caspi (2003). To the extent that we also observed sub-stantial environmental contributions to stability across time, our re-sults would tend to provide greater support for Roberts and Caspi’sconception.
METHOD
Sample
The study sample was drawn from participants in the Minnesota TwinStudy of Adult Development and Aging, a population-based registry in-cluding 901 intact monozygotic and same-sex dizygotic twin pairs born inthe state of Minnesota. Most were born between 1904 and 1934, but 29%were born between 1936 and 1955, and one pair was born in 1897. Thisstudy has been described previously (Finkel & McGue, 1994). Briefly,participants were identified from Minnesota State Health Departmentbirth records and recruited using state marriage and motor vehiclerecords; the initial participation rate was approximately 60%, likely dueto the 4–5-hour length of the in-home assessment, which included a seriesof cognitive, personality, and lifestyle measures. Zygosity was determinedusing a five-item questionnaire that has been shown to provide over 95%accuracy (Lykken, Bouchard, McGue, & Tellegen, 1990). Twins wereassigned as Twin 1 and Twin 2 by order of birth.
A total of 1,123 participants (including 378 members of complete maletwin pairs, 556 members of complete female twin pairs, and 189 twinswhose co-twins did not provide data) completed the MPQ in the early1990’s. In addition, 833 of these individuals (118 members of completemale MZ twin pairs, 106 members of complete male DZ twin pairs, 266members of complete female MZ twin pairs, 168 members of completefemale DZ twin pairs, and 90 male twins and 85 female twins whose co-twins did not provide data) completed the MPQ again, on average 5.0years later (SD 2.4, range 1.0–10.4). Participants averaged 59.4 years ofage (SD 9.7, range 27.0–92.0) at first and 64.4 (SD 10.2, range 32.2–98.7)at second administration. Though the full age range was rather large, themajority of participants were between ages 50 and 70 at first testing; lessthan 18% of participants were under age 50, and less than 10% were overage 70.
532 Johnson, McGue, & Krueger

Analysis of Mean-Level Personality Stability
To assess mean-level stability of our personality measures, we fit a hier-archical linear model (Bryk & Raudenbush, 1992), with overall meanscale score and retest interval as fixed effects and twin pair and initialscore as random effects, to each set of MPQ scale scores. The use of in-terval of testing was necessary because the testing interval was reasonablyevenly distributed over the range from 1.5 to 9 years. We made use of thehierarchical linear modeling technique rather than analysis of variance sothat the estimates of statistical precision resulting from our models wouldbe correctly adjusted both for the variable retest interval and for thecorrelations between scale scores within twin pairs.
Age and Sex Adjustment
Twins share a common age and, in our sample, a common sex. Thus ageand sex effects act to increase twin similarity (McGue & Bouchard, 1984).To correct for these effects in the biometric analysis of the twin data, weadjusted the personality scores at the two time points by subtracting thesex-appropriate mean from each individual’s score and regressing out thelinear component of age at each time point. In doing so, we did not normthe residuals to unit variance as we wished to be able to identify factorsthat contribute to variability in whatever personality change we observed.
Biometric Modeling
Our quantitative genetic model is based on the assumption that the ob-served phenotypic variance (Vp) is a linear additive function of genetic(A) and shared (C) and nonshared (E) environmental variance, respec-tively. Symbolically,
Vp! A" C" E:
Under this model, the nonshared environmental variance representsresidual variance not explained by either of the other two sources. Thenonshared environmental component also includes variance attributableto measurement error. Genetic variance can be additive in the sense that ifmultiple genes influence the trait, they do so independently of each other.It can also be nonadditive, reflecting dominance and other polygenic ef-fects. We note that there is evidence for nonadditive effects on person-ality, but little evidence for shared environmental effects (Bouchard &Loehlin, 2001; Finkel & McGue, 1997; Tellegen et al., 1988). For additivegenetic effects, the expected covariance between any two members of atwin pair as a function of the variance components given above can be
Personality Stability 533

specified as,
COV#MZ$! A" C
COV#DZ$ !:5%A" C:
Heritability estimates of this form are based on several assumptions.The first is that twins (both MZ and DZ) are representative of the pop-ulation as a whole for the trait in question. For personality traits, inparticular as measured by the MPQ, this appears to be the case ( Johnson,Krueger, Bouchard, & McGue, 2002). Second, we assume that MZ twinsshare trait-relevant environmental influences to the same degree as DZtwins. Numerous attempts have been made to uncover circumstances inwhich this assumption does not hold, with generally negative results (e.g.,Borkenau, Riemann, Angleitner, & Spinath, 2002; Loehlin & Nichols,1976). Another assumption is that there is no assortative mating (meaningthat the parents of the twins were not similar) for the traits in question.Because there is some marital resemblance (with coefficients ranging from.10 to .20) for most personality-related traits (Price & Vandenberg, 1980),including those measured by the MPQ (Lykken & Tellegen, 1993), thisassumption generally has the effect of understating heritability estimates.Such relatively small values, however, are unlikely to reduce estimates ofgenetic influence substantially.
In models of this form, we also assume that there are no gene-envi-ronment interactions or correlations. It is possible to develop models as-sessing the effects of such correlations and interactions (Purcell, 2002),but it is necessary to specify and measure the environmental moderators.When left unmodeled, as we have done here, the presence of gene-envi-ronment interactions or correlations do not render the resulting estimatesof genetic and environmental influences meaningless. Rather, they statethem at overall population levels. Though this could mask differences inunderlying subgroups within the data, our goals in this study involvedclarification of such overall population level effects. This is an importantstep in the long-term process of developing conceptualizations of theoperation of gene-environment correlations or interactions in a mannercoherent and specific enough that they can be tested.
The final assumption is that the portion of variance attributable togenetic influence is additive. Violations of this assumption could maskshared environmental effects in reared-together twin studies, but withoutreared-apart twins, we have no way of addressing the extent to which theassumption holds in the current study. We note that there have been re-ports of significant nonadditive effects on personality, particularly asmeasured by the MPQ (though the particular effects do not generally
534 Johnson, McGue, & Krueger

replicate), but little evidence for shared environmental effects (Tellegenet al., 1988; Bouchard & Loehlin, 2001).
We began our biometric analyses by using Mx (Neale, 1997) to fitbivariate Cholesky models (ACE models) estimating additive genetic(termed the ‘‘A’’ parameter), and shared or common (termed the ‘‘C’’parameter) and nonshared (termed the ‘‘E’’ parameter) environmentalcontributions to variance in and covariance between the age- and sex-adjusted MPQ personality scale scores at the two time points (referred toas Time 1 and Time 2). Because we had a substantial number of brokentwin pairs in our sample, we made use of the raw data option in Mx sothat the unmatched twins could contribute information about the vari-ances at the two time points. The path coefficients in the Cholesky modelswe fit were partial regression estimates, with the genetic and environ-mental path estimates to Time 1 stated directly. The effects of genetic andenvironmental influences on Time 2 personality, however, were decom-posed into those specific to Time 2 and those in common with Time 1. Weexplicitly made accommodation for the length of the retest interval byallowing it to moderate the decomposition of the genetic and environ-mental influences on Time 2 personality (a copy of the Mx script we usedto do so can be obtained from the first author upon request).
We derived two pieces of information from the Cholesky decomposi-tion. Figure 1 depicts the information we computed. First, we estimatedthe correlation between the genetic effects on personality at Time 1 andthe genetic effects on personality at Time 2. This is known as the geneticcorrelation. The genetic correlation is defined as the square root of theproportion of the total heritability of the Time 2 scale score that can beexplained by genetic effects at Time 1. Thus, when the genetic correlationis 1.00, all of the genetic influences are common to the two times. Thisalso means that there is no room for genetic influence on whatever changehas taken place over the interval. When the genetic correlation is 0.00,completely different sets of genetic influences act to affect individual dif-ferences at the two ages, and change could be genetic or environmental inorigin to any degree. In other words, the genetic correlation reflects theextent to which the genetic effects overlap at the two times, regardless oftheir relative contribution to phenotypic variance. A similar interpreta-tion can be given to correlations of environmental influences, and wemade the analogous calculation to estimate the nonshared environmentalcorrelation between scale scores at Times 1 and 2. Thus, if stability inpersonality were primarily genetic in origin, we would expect high geneticcorrelations and very low environmental correlations. If stability in per-sonality reflected transactions between genes and environments, we wouldexpect substantial correlations at both the genetic and environmentallevels. The shared environmental influences on the personality scale
Personality Stability 535
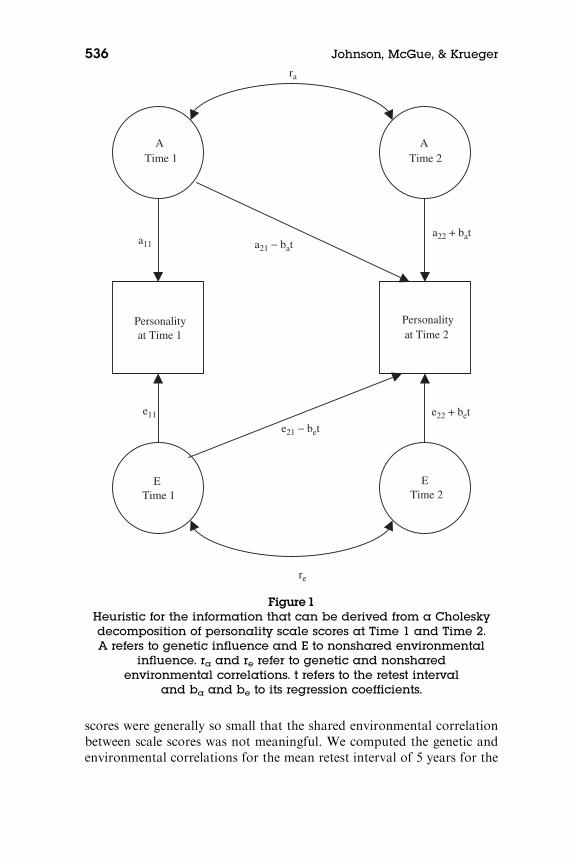
scores were generally so small that the shared environmental correlationbetween scale scores was not meaningful. We computed the genetic andenvironmental correlations for the mean retest interval of 5 years for the
Personalityat Time 1
Personalityat Time 2
ATime 1
ETime 1
ETime 2
ra
re
a22 + bat
e22 + bet
a11
e11
e21 ! bet
a21 ! bat
ATime 2
Figure1Heuristic for the information that can be derived from a Choleskydecomposition of personality scale scores at Time 1 and Time 2.A refers to genetic influence and E to nonshared environmental
influence. ra and re refer to genetic and nonsharedenvironmental correlations. t refers to the retest interval
and ba and be to its regression coefficients.
536 Johnson, McGue, & Krueger

sample. Where the length of the retest interval had a significant effect(as measured at p5 .02 or less; to recognize that, we applied 14 separatetests), we also computed the range over which these correlations wouldfall for the retest interval range of 1.0–10.4 years.
RESULTSAttrition Analysis
There were small differences between those who completed the MPQat Time 1 but who did not complete it again at Time 2 and those whocompleted it at both times. Uncorrected for the effects of multipletesting, the significant differences included the following: Those whocompleted the MPQ only at Time 1 were slightly younger (M5 57.7vs. M5 59.4 for those completing it at both times, p5 .01, standard-ized mean difference & .17) and had marginally lower Social Close-ness scale scores (M5 15.1 vs. M5 15.9 for those completing it atboth times, p5 .02, standardized mean difference & .17), higher Al-ienation scale scores (M5 2.3 vs. M5 1.9 for those completing it atboth times, p5 .02, standardized mean difference .16), higher Aggres-sion scale scores (M5 3.2 vs. M5 2.7 for those completing it at bothtimes, p5 .00, standardized mean difference .20), and lower HarmAvoidance scale scores (M5 21.3 vs. M5 22.6 for those completingit at both times, p5 .00, standardized mean difference & .27).All of these differences would generally be considered small (Cohen,1988).
Analysis of Mean Level Change
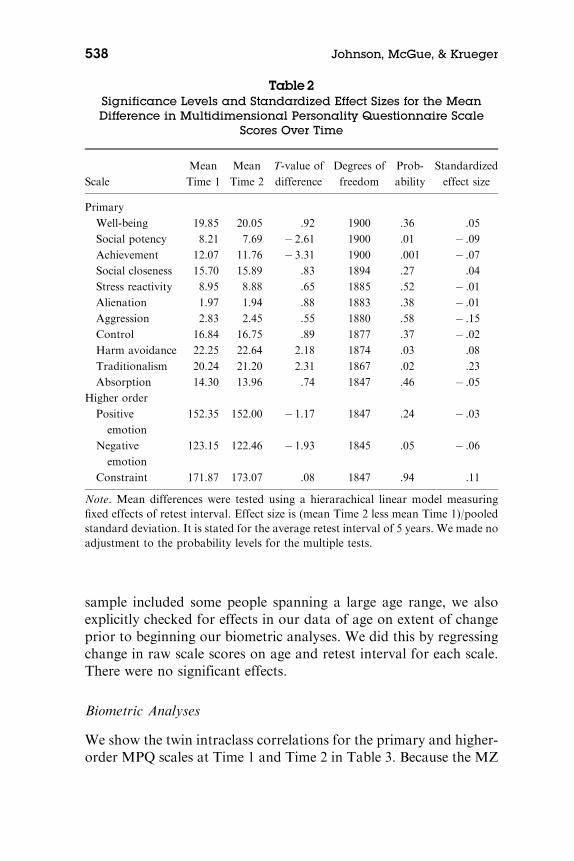
Table 2 gives the results of the hierarchical linear model analysesof Time 1 and Time 2 scale scores, conducted to assess the signif-icance of changes in mean level while correcting for the correlatednature of the twin observations. We show the means at the two timepoints, t-statistics for the estimated parameters for time, and the as-sociated probabilities. To put the mean differences in perspective, wealso show their effect sizes (standardized mean differences). Therewas a clearly significant mean difference only for Achievement.There were small (Cohen, 1988) mean effect size differences of & .15for Aggression and .23 for Traditionalism, though the mean differ-ence was not significant for these scales after adjustment for thevarying retest interval and correlated twin observations. Because our
Personality Stability 537
sample included some people spanning a large age range, we alsoexplicitly checked for effects in our data of age on extent of changeprior to beginning our biometric analyses. We did this by regressingchange in raw scale scores on age and retest interval for each scale.There were no significant effects.
Biometric Analyses
We show the twin intraclass correlations for the primary and higher-order MPQ scales at Time 1 and Time 2 in Table 3. Because the MZ
Table2Significance Levels and Standardized Effect Sizes for the MeanDifference in Multidimensional Personality Questionnaire Scale
Scores Over Time
Scale
Mean
Time 1
Mean
Time 2
T-value of
difference
Degrees of
freedom
Prob-
ability
Standardized
effect size
Primary
Well-being 19.85 20.05 .92 1900 .36 .05
Social potency 8.21 7.69 & 2.61 1900 .01 & .09
Achievement 12.07 11.76 & 3.31 1900 .001 & .07
Social closeness 15.70 15.89 .83 1894 .27 .04
Stress reactivity 8.95 8.88 .65 1885 .52 & .01
Alienation 1.97 1.94 .88 1883 .38 & .01
Aggression 2.83 2.45 .55 1880 .58 & .15
Control 16.84 16.75 .89 1877 .37 & .02
Harm avoidance 22.25 22.64 2.18 1874 .03 .08
Traditionalism 20.24 21.20 2.31 1867 .02 .23
Absorption 14.30 13.96 .74 1847 .46 & .05
Higher order
Positive
emotion
152.35 152.00 & 1.17 1847 .24 & .03
Negative
emotion
123.15 122.46 & 1.93 1845 .05 & .06
Constraint 171.87 173.07 .08 1847 .94 .11
Note. Mean differences were tested using a hierarachical linear model measuringfixed effects of retest interval. Effect size is (mean Time 2 less mean Time 1)/pooledstandard deviation. It is stated for the average retest interval of 5 years. We made noadjustment to the probability levels for the multiple tests.
538 Johnson, McGue, & Krueger
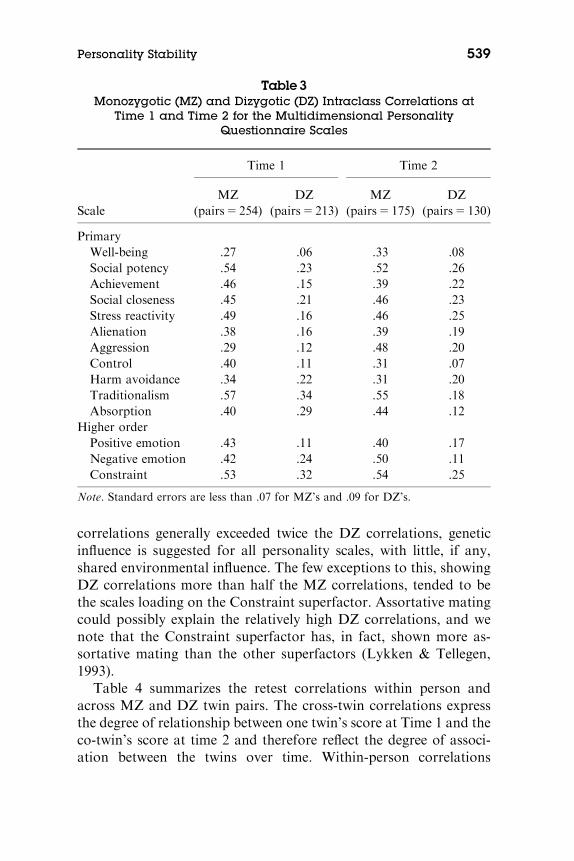
correlations generally exceeded twice the DZ correlations, geneticinfluence is suggested for all personality scales, with little, if any,shared environmental influence. The few exceptions to this, showingDZ correlations more than half the MZ correlations, tended to bethe scales loading on the Constraint superfactor. Assortative matingcould possibly explain the relatively high DZ correlations, and wenote that the Constraint superfactor has, in fact, shown more as-sortative mating than the other superfactors (Lykken & Tellegen,1993).
Table 4 summarizes the retest correlations within person andacross MZ and DZ twin pairs. The cross-twin correlations expressthe degree of relationship between one twin’s score at Time 1 and theco-twin’s score at time 2 and therefore reflect the degree of associ-ation between the twins over time. Within-person correlations
Table 3Monozygotic (MZ) and Dizygotic (DZ) Intraclass Correlations at
Time 1 and Time 2 for the Multidimensional PersonalityQuestionnaire Scales
Scale
Time 1 Time 2
MZ(pairs5 254)
DZ(pairs5 213)
MZ(pairs5 175)
DZ(pairs5 130)
PrimaryWell-being .27 .06 .33 .08Social potency .54 .23 .52 .26Achievement .46 .15 .39 .22Social closeness .45 .21 .46 .23Stress reactivity .49 .16 .46 .25Alienation .38 .16 .39 .19Aggression .29 .12 .48 .20Control .40 .11 .31 .07Harm avoidance .34 .22 .31 .20Traditionalism .57 .34 .55 .18Absorption .40 .29 .44 .12
Higher orderPositive emotion .43 .11 .40 .17Negative emotion .42 .24 .50 .11Constraint .53 .32 .54 .25
Note. Standard errors are less than .07 for MZ’s and .09 for DZ’s.
Personality Stability 539
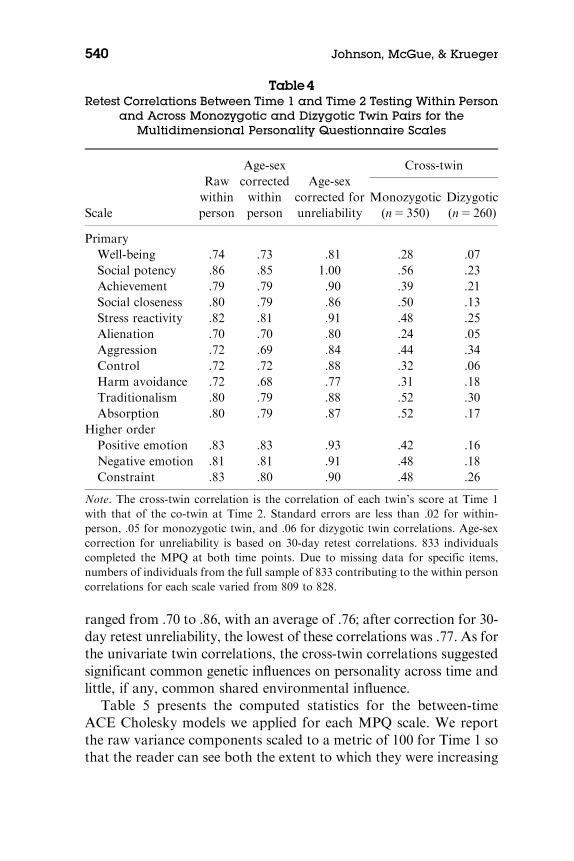
ranged from .70 to .86, with an average of .76; after correction for 30-day retest unreliability, the lowest of these correlations was .77. As forthe univariate twin correlations, the cross-twin correlations suggestedsignificant common genetic influences on personality across time andlittle, if any, common shared environmental influence.
Table 5 presents the computed statistics for the between-timeACE Cholesky models we applied for each MPQ scale. We reportthe raw variance components scaled to a metric of 100 for Time 1 sothat the reader can see both the extent to which they were increasing
Table4Retest Correlations Between Time 1 and Time 2 Testing Within Person
and Across Monozygotic and Dizygotic Twin Pairs for theMultidimensional Personality Questionnaire Scales
Scale
Rawwithinperson
Age-sexcorrectedwithinperson
Age-sexcorrected forunreliability
Cross-twin
Monozygotic(n5 350)
Dizygotic(n5 260)
PrimaryWell-being .74 .73 .81 .28 .07Social potency .86 .85 1.00 .56 .23Achievement .79 .79 .90 .39 .21Social closeness .80 .79 .86 .50 .13Stress reactivity .82 .81 .91 .48 .25Alienation .70 .70 .80 .24 .05Aggression .72 .69 .84 .44 .34Control .72 .72 .88 .32 .06Harm avoidance .72 .68 .77 .31 .18Traditionalism .80 .79 .88 .52 .30Absorption .80 .79 .87 .52 .17
Higher orderPositive emotion .83 .83 .93 .42 .16Negative emotion .81 .81 .91 .48 .18Constraint .83 .80 .90 .48 .26
Note. The cross-twin correlation is the correlation of each twin’s score at Time 1with that of the co-twin at Time 2. Standard errors are less than .02 for within-person, .05 for monozygotic twin, and .06 for dizygotic twin correlations. Age-sexcorrection for unreliability is based on 30-day retest correlations. 833 individualscompleted the MPQ at both time points. Due to missing data for specific items,numbers of individuals from the full sample of 833 contributing to the within personcorrelations for each scale varied from 809 to 828.
540 Johnson, McGue, & Krueger
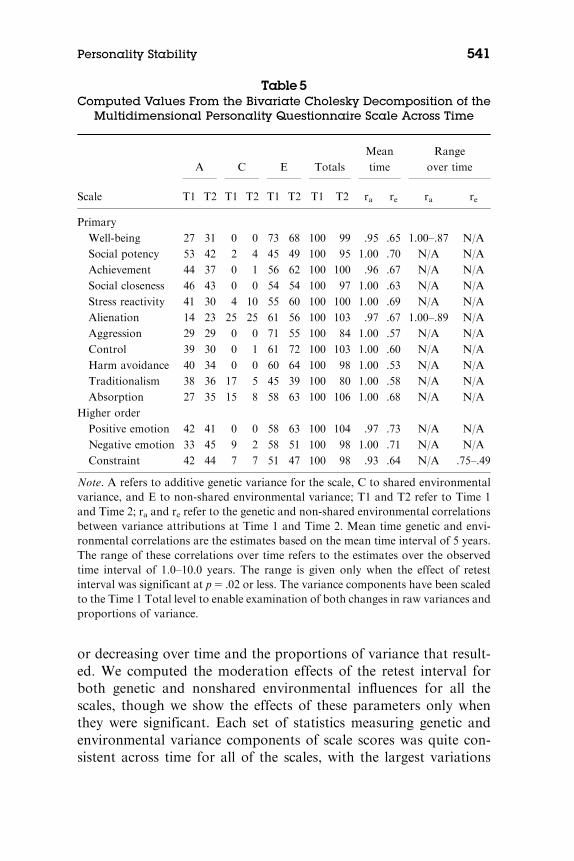
or decreasing over time and the proportions of variance that result-ed. We computed the moderation effects of the retest interval forboth genetic and nonshared environmental influences for all thescales, though we show the effects of these parameters only whenthey were significant. Each set of statistics measuring genetic andenvironmental variance components of scale scores was quite con-sistent across time for all of the scales, with the largest variations
Table 5Computed Values From the Bivariate Cholesky Decomposition of the
Multidimensional Personality Questionnaire Scale Across Time
Scale
A C E Totals
Mean
time
Range
over time
T1 T2 T1 T2 T1 T2 T1 T2 ra re ra re
Primary
Well-being 27 31 0 0 73 68 100 99 .95 .65 1.00–.87 N/A
Social potency 53 42 2 4 45 49 100 95 1.00 .70 N/A N/A
Achievement 44 37 0 1 56 62 100 100 .96 .67 N/A N/A
Social closeness 46 43 0 0 54 54 100 97 1.00 .63 N/A N/A
Stress reactivity 41 30 4 10 55 60 100 100 1.00 .69 N/A N/A
Alienation 14 23 25 25 61 56 100 103 .97 .67 1.00–.89 N/A
Aggression 29 29 0 0 71 55 100 84 1.00 .57 N/A N/A
Control 39 30 0 1 61 72 100 103 1.00 .60 N/A N/A
Harm avoidance 40 34 0 0 60 64 100 98 1.00 .53 N/A N/A
Traditionalism 38 36 17 5 45 39 100 80 1.00 .58 N/A N/A
Absorption 27 35 15 8 58 63 100 106 1.00 .68 N/A N/A
Higher order
Positive emotion 42 41 0 0 58 63 100 104 .97 .73 N/A N/A
Negative emotion 33 45 9 2 58 51 100 98 1.00 .71 N/A N/A
Constraint 42 44 7 7 51 47 100 98 .93 .64 N/A .75–.49
Note. A refers to additive genetic variance for the scale, C to shared environmentalvariance, and E to non-shared environmental variance; T1 and T2 refer to Time 1and Time 2; ra and re refer to the genetic and non-shared environmental correlationsbetween variance attributions at Time 1 and Time 2. Mean time genetic and envi-ronmental correlations are the estimates based on the mean time interval of 5 years.The range of these correlations over time refers to the estimates over the observedtime interval of 1.0–10.0 years. The range is given only when the effect of retestinterval was significant at p5 .02 or less. The variance components have been scaledto the Time 1 Total level to enable examination of both changes in raw variances andproportions of variance.
Personality Stability 541

being on the order of 10%. The magnitudes of the proportional in-fluences on the personality scale scores were very comparable tothose reported in other studies (Bouchard & Loehlin, 2001), exceptfor the genetic influence on Alienation. That is, excluding Alienationand restating the raw variance components as proportions of thetotals, the estimated genetic influences ranged from 27% to 53%,and estimated shared environmental influences were generally neg-ligible, though Alienation, Traditionalism, and Absorption, in par-ticular, showed some shared environmental influence. We believethat the unusually low estimate of genetic influence on Alienationresulted because the variance in that scale was notably smaller thanthose of the other scales with the exception of Aggression, resultingin depression of the correlations due to restriction of range. Thefreely estimated genetic correlations (ra) for the mean retest intervalof 5.0 years ranged from .93 to 1.00. The estimated nonshared en-vironmental correlations (re) were also both significant and substan-tial; they ranged from .53 to .73. For the range of significant retestintervals included in the sample, the nonshared environmental cor-relations did not drop below .49.
The high genetic correlations and small amounts of mean levelchange in the data implied that there were no substantive geneticinfluences on change in personality over the time interval. It waspossible, however, that the small mean level changes masked sub-stantial changes at the individual level that could have either geneticor environmental influences. To evaluate this possibility, we com-puted the MZ and DZ twin correlations for change in raw scalescores over time. The largest correlation was .24, and it was the onlyone that was significant. Thus, consistent with the genetic correla-tions we computed, there was little or no evidence for genetic influ-ence on change in personality over time for this sample of primarilyolder adults.
In our tables, we present the results for the ACE models, in spiteof the fact that, for some of the models, including a parameter es-timating nonadditive genetic influence instead of a parameter esti-mating shared environmental influence fit better. There is substantialevidence for the existence of such nonadditive genetic influences onMPQ scales (Finkel & McGue, 1997; Tellegen et al., 1988). Theirestimation, however, is fraught with difficulty as they cannot be es-timated simultaneously with both additive and shared environmentaleffects, and the power to estimate them is very weak (Martin, Eaves,
542 Johnson, McGue, & Krueger

Kearsey, & Davies, 1978). Probably because of this, when they areestimated, their existence seems to vary considerably by sample,personality measurement scale, and over time (Bouchard & McGue,2003). This was true in our data as well, as nonadditive genetic in-fluences were estimated to be present at Time 1 but absent at Time 2,and vice versa, for several scales. Though we fit models that includednonadditive genetic variance, and they provided some improvementin fit for Wellbeing, Control, and Positive Emotionality, we decidedto present the estimates from the ACE model because we believedthat model to provide the most consistent estimates from the mostgeneral perspective regarding possible sources of genetic influence onpersonality. Though model fit could be improved by eliminating theC parameter, the relationships among model fit statistics for thetypes of models we applied remained the same whether includingor excluding the C parameter, and these relationships were the realfocus of our investigation.
DISCUSSION
This study replicates prior findings of substantial stability of per-sonality in adulthood (see Roberts & DelVecchio, 2000, for a recentreview), and extends those results by focusing primarily on the levelof stability of personality in late adulthood. In addition, the studyextends previous research by measuring personality facets. Our per-sonality measures, even at the primary scale level, were highly stableand showed genetic influences very similar to those from other per-sonality studies (Bouchard & Loehlin, 2001). For all practical pur-poses, all of the genetic influences on personality at Time 1 were stillpresent at Time 2 for each of the primary scales measured, as well asfor the three higher-order factors, and there was no evidence thatnew genetic influences had been introduced. This could be viewedfrom the perspective of substantive genetic contributions to a rangeof stable latent personality traits, given that the genetic variancecomponents were also substantive. At the same time, our resultssuggested that the nonshared environment also played a substantialrole in the stability we observed, as the nonshared environmentalcorrelations and variance components were also high for all of thescales. This, too, could be viewed from the perspective of a range ofstable latent personality traits. In fact, our results indicated that thegenetic and nonshared environmental influences on the latent stable
Personality Stability 543

personality traits were roughly equal. This indicates that, thoughMcCrae et al. (2000) appear to be correct that genetic influencescontribute importantly to the stability of personality in late adult-hood, the contribution of the environment is somewhat more im-portant than their conceptualization would suggest. We think itlikely that the nonshared environment makes the contributions tostability that we observed through transactions over time betweengenes and environment of the sort articulated by Roberts and Caspi(2003).
Increase or decrease in variance over time is one of the key issuesin studies of aging. Increasing variance was addressed at some lengthin Pedersen and Reynolds’s (1998) longitudinal personality study,and Viken et al. (1994) also noted increases in variance with age. Wepresented our findings in such a way that this issue could be eval-uated directly here as well. Still, we found no systematic changes invariances over time. In fact, we found that we could improve modelfit by constraining the variances at our two time points to be equalfor all scales except Traditionalism. Pedersen and Reynolds inter-preted the variance increases they observed in the context of terminaldrop associated with impending death in the oldest members of theirsample. Though our sample consisted primarily of older adults, 90%were under 70 years old at first testing and generally would not beconsidered elderly or likely to be facing terminal drop. Perhaps thiscontributed to the lack of increasing variability in our sample,though it would do nothing to explain the difference in this areabetween our results and those of Viken et al.
We presented raw variance components in our results, leading tothe observation that genetic and nonshared environmental contri-butions to personality were about equal. The high degree of stabilityin the variances over time, however, makes median summary valuesexpressing proportions of variance meaningful as a way of furtherexploring the contribution of nonshared environmental influences tothe personality stability we observed. That is, corrected for meas-urement error, the median nonshared environmental correlation weobserved was .65, suggesting that stable, nonshared environmentalinfluences play a large role, accounting for over 40% of variance.That 40% of individual differences in personality can be attributedto stable, nonshared environmental influences is a striking finding. Itis also a finding that is not unique to this study (Angleitner & Spi-nath, 2002; Pedersen & Reynolds, 1998; Viken et al., 1994), though
544 Johnson, McGue, & Krueger

these other studies have drawn little attention to it. It suggests that asubstantial portion of an individual’s personality cannot be predict-ed by knowing his or her genetic background, nor by knowing theenvironment he or she shared with a co-twin. Rather, in keeping withRoberts and Caspi’s (2003) conception, it can be predicted by his orher prior personality and the individual circumstances in which he orshe lives, including the consequences of prior life events.
For example, the environmental circumstances of people’s livestend to be relatively stable in the 50 to 70 age range in which 73% ofour sample fell at first testing (e.g., Schacter & Kuenzi, 2002). This isnot to say that change does not take place in this age range: childrenmay be leaving the home, elderly parents may be in need of care, jobroles may change due to downsizing, early retirement, or promotion,etc. It is merely to say that, relative to other life periods such as earlyadulthood, people’s lives tend to be subject to less change duringlater adulthood. This is true in general, but more importantly, it wascertainly true for this sample in this geographic region at the par-ticular point in history in which they were assessed, given the cul-tural-level environmental influences to which they were exposed.That is, in the mid-1990s, when this sample was assessed, Minnesota,where they resided, experienced no major catastrophic events such aswars, earthquakes, hurricanes, large terrorist attacks, major eco-nomic upheavals, or other disasters that might tend to cause majorupheavals in the lives of many.
The environmental stability for particular individuals within thisage range results from the highly individualized combination of cir-cumstances people have chosen over time as a result of their genet-ically influenced natures and relatively stable circumstances thathave emerged for a variety of primarily nonshared environmentallydetermined reasons. Thus, the environmental stability we have notedhere contributes to a gene-environment correlation and/or interac-tion of the sort behavioral genetic studies have found very difficult toisolate and evaluate (Plomin, DeFries, McClearn, & Rutter, 1997),as there are many different ways in which it can occur. Particularly inthis age range, for example, it is very common for people to live formany years in the same community (even in the same house), to bemarried to the same spouse or not married, to hold the same job, andto experience no changes in the people living in their home, to namejust a few sources of possible environmental stability that would notgenerally be shared with their co-twins. Though change can of course
Personality Stability 545

occur in this age range, for many, much of it is a period after childrenhave left home and menopause for women has taken place and be-fore retirement, major deteriorations in health and mobility, andspousal bereavement take place. With respect to marriage as onesource of this kind of stability, Caspi and Herbener (1992, 1993)have presented evidence that spouses do not become more similar inpersonality over time, suggesting rather that the shared spousal en-vironment plays a significant role in maintaining each spouse’s par-ticular personality. This probably takes place through theestablishment of the stable individual habits and compromises inlifestyle unique to each married couple. We interpret these environ-mental sources of stability as contributing directly to the substantialnonshared environmental correlations as well as the raw nonsharedvariance components we observed.
Since Plomin and Daniels’s (1987) review of behavior geneticfindings, the nonshared environment has tended to be conceptual-ized as a combination of measurement error and experiences uniqueto each individual that are associated with personality change. Theresulting searches (e.g., Krueger, Markon, & Bouchard, 2003; Reiss,Neiderhiser, Hetherington, & Plomin, 2000; Turkheimer &Waldron,2000) for measured, nonshared environmental variables that can ac-count for the substantial portions of the nonshared environmentalinfluences on differences among individuals have revealed the elusivenature of these influences. The substantial component of stable per-sonality that our results and those of others attribute to stable non-shared environmental influences suggests that the identification ofsuch stable influences may be a far more promising research ap-proach to the explanation of environmental influences on behavior.Individual experiences are always unique to the individual and thushighly idiosyncratic and difficult to measure. But individual experi-ences can be highly stable over time, and this stability should makethem and their consequences more systematic and thus more readilymeasurable. This would mean collecting highly specific data on lifecircumstances in such areas as work, family and marital relation-ships, place of residence, community involvement, etc. and compar-ing the degree and sources of the absolute value of personalitychange in those who had experienced change in specific classes ofcircumstances to those who had not. Though our data included somemeasures of life circumstances such as marital status, type of resi-dence, and occupation, many of the questions were not asked at both
546 Johnson, McGue, & Krueger

time points, nor were they asked in a manner specific enough toevaluate to what extent change may have taken place. In addition,this sample would probably not be the best one in which to study theeffects of changing life circumstances on personality change in thismanner, as there was very little change in personality to explain.
This should not be taken to imply that we would expect the rel-ative proportions of stable genetic and environmental influences tobe constant over the lifespan or constant for people of a given ageliving in very different environmental circumstances. The very highlevel of stability of personality across time in our sample may havehad an effect on the relative magnitude of the genetic and environ-mental influences on the latent personality phenotypes we modeled,as well as on the bivariate genetic and environmental influences weestimated. Specifically, for people of younger ages who might beexpected to be more subject to developmental influences and to livein less stable circumstances, it seems possible that the relative pro-portions of genetic influences on these phenotypes would be largerand the stable nonshared environmental influences smaller.
The sample size used in this study is modest for a twin study, andconfined to a regional cohort living in relatively stable economic andphysical conditions when measured on a historical scale. Thus, careshould be taken in drawing generalizations to other samples living inother circumstances. In addition, though the personality measure weused has good psychometric properties, it is only a single (self-re-port) measure and subject to the limitations of all such measures.This is true as well of the other longitudinal studies of personality(McGue, Bacon, & Lykken, 1993; Pedersen & Reynolds, 1998; Vikenet al., 1994). Thus, the question of consistency of stability estimatesacross reporters and other methods of observation should be inves-tigated as well. The question of genetic and environmental influenceson personality stability and change over time should be investigatedin other samples, using other personality measures at the facet leveland other approaches such as adoption studies. Alternative ap-proaches to evaluate the validity of the assumptions underlying thebiometric models we and others have applied should also be pur-sued. Nonetheless, we believe that our results add substantially tothe growing body of evidence for the conception of adult personalityas an outgrowth of the transaction between a stable underlying ge-netic base and a set of environmental influences largely unique to theindividual that reflect individual life events and their consequences
Personality Stability 547

over time. Thus, we believe it fair to speculate that adult personalitystability results from a combination of genetic and continuingenvironmental influences, while adult personality change resultsboth from life events beyond the control of the individual andfrom new individual life choices. We intend to pursue this conceptionin future research.
REFERENCES
Akaike, H. (1983). Information measures and model selection. Bulletin of theInternational Statistical Institute, 50, 277–290.
Angleitner, A., & Spinath, F. M. (2002). Personality in adulthood: Findings fromthe Bielefeld Longitudinal Study of Adult Twins. 11th European Conferenceon Personality, Jena, Germany.
Borkenau, P., Riemann, R., Angleitner, A., & Spinath, F. M. (2002). Similarityof childhood experiences and personality resemblance in monozygotic anddizygotic twins: A test of the equal environments assumption. Personalityand Individual Differences, 33, 261–269.
Bouchard, T. J., & Loehlin, J. C. (2001). Genes, evolution, and personality. Be-havior Genetics, 31 (3), 243–273.
Bouchard, T. J., & McGue, M. (2003). Genetic and environmental influences onhuman psychological differences. Journal of Neuroscience, in press.
Browne, M., & Cudeck, R. (1992). Alternative methods of assessing model fit.Sociological Methods & Research, 21, 230–258.
Bryk, A. S., & Raudenbush, S. W. (1992). Hierarchical Linear Models: Applica-tions and Data Analysis Methods. Newbury Park, CA: Sage.
Carmichael, C. M. (1993). A cross-sectional examination of twin similarity inpersonality during adulthood. Dissertation Abstracts, International, 54 (6-B),3358, US: Univ. Microfilms International.
Caspi, A., & Herbener, E. S. (1992). Shared experiences and the similarity ofpersonalities: A longitudinal study of married couples. Journal of Personalityand Social Psychology, 62, 281–291.
Caspi, A., & Herbener, E. S. (1993). Marital and phenotypic convergence: Lon-gitudinal evidence. Social Biology, 40 (1–2), 48–60.
Caspi, A., & Roberts, B. W. (2001). Personality development across the lifecourse: The argument for change and continuity. Psychological Inquiry, 12 (2),49–66.
Church, A. T. (1994). Relating the Tellegen and Five-Factor models of personalitystructure. Journal of Personality and Social Psychology, 67, 898–909.
Cohen, J. (1988). Statistical power analysis (2nd ed.). Hillsdale, NJ: Erlbaum.Conley, J. J. (1985). Longitudinal stability of personality traits: A multitrait-mul-
tidimensional-multioccasional analysis. Journal of Personality and Social Psy-chology, 49, 1266–1282.
Costa, P. T., & McCrae, R. R. (1988). Personality in adulthood: A six-year lon-gitudinal study of self-reports and spouse ratings of the NEO personalityinventory. Journal of Personality and Social Psychology, 54, 853–863.
548 Johnson, McGue, & Krueger

Costa, P. T., & McCrae, R. R. (1992a). The revised NEO Personality Inventory(NEO-PI-R) and the NEO Five Factor Inventory (NEO-FFI) professionalmanual. Odessa, FL: Psychological Assessment Resources.
Costa, P. T., & McCrae, R. R. (1992b). Trait psychology comes of age. In T. B.Sonderegger (Ed.), Nebraska Symposium on Motivation: Psychology and Aging(pp. 169–204). Lincoln, NE: University of Nebraska Press.
Derryberry, D., & Reed, M. A. (1994). Temperament and attention: Orientingtoward and away positive and negative signals. Journal of Personality andSocial Psychology, 66, 1128–1139.
Eaves, L. J., Eysenck, H. J., & Martin, N. G. (1989). Genes, culture, and person-ality: An empirical approach. San Diego, CA: Academic Press.
Eysenck, H. J. (1960). The structure of human personality. London: Methuen.Eysenck, H. J., & Eysenck, M. W. (1985). Personality and individual differences.
New York: Plenum.Finkel, D., & McGue, M. (1994). The origins of individual differences in memory
among the elderly: A behavior genetic analysis. Psychology and Aging, 8 (4),527–537.
Finkel, D., & McGue, M. (1997). Sex differences and nonadditivity in heritabilityof the Multidimensional Personality Questionnaire Scales. Journal of Person-ality and Social Psychology, 72 (4), 929–938.
Floderus-Myrhed, B., Pedersen, N., & Rasmuson, I. (1980). Assessment of her-itability for personality, based on a short form of the Eysenck PersonalityInventory: A study of 12,898 twin pairs. Behavior Genetics, 10, 153–162.
Gough, H. G., & Bradley, P. (1996). CPI Manual (3rd Ed.). Palo Alto, CA: Con-sulting Psychologists Press, Inc.
Helfand, S. L., & Rogina, B. (2003). Genetics of aging in the fruit fly, Drosophilamelanogaster. Annual Review of Genetics, 37, 329–348.
Helson, R., & Klohnen, E. C. (1998). Affective coloring of personality fromyoung adulthood to midlife. Personality and Social Psychology Bulletin, 24,241–252.
Johnson, W., Krueger, R. F., Bouchard, T. J., & McGue, M. (2002). The per-sonalities of twins: Just ordinary folks. Twin Research, 5 (2), 125–131.
Krueger, R. F., Markon, K. E., & Bouchard, T. J. (2003). The extended genotype:The heritability of personality accounts for the heritability of recalled familyenvironments in twins reared apart. Journal of Personality, 71, 809–833.
Lang, P. J. (1995). The emotion probe: Studies of motivation and attention.American Psychologist, 50, 372–385.
Loehlin, J. C. (1992). Genes and environment in personality development. NewburyPark, CA: Sage.
Loehlin, J. C., & Nichols, J. (1976).Heredity, environment and personality. Austin:University of Texas.
Lubinski, D., Schmidt, D. B., & Benbow, C. P. (1996). A 20-year stability analysisof the study of values for intellectually gifted individuals from adolescence toadulthood. Journal of Applied Psychology, 81, 443–451.
Lykken, D. T., Bouchard, T. J., McGue, M., & Tellegen, A. (1990). The Min-nesota Twin Family Registry: Some initial findings. Acta Genetica MedicaGemellol, 39, 35–70.
Personality Stability 549

Lykken, D. T., & Tellegen, A. (1993). Is human mating adventitious or the resultof lawful choice? A twin study of mate selection. Journal of Personality andSocial Psychology, 65 (1), 56–68.
Martin, N. G., Eaves, L. J., Kearsey, M. J., & Davies, P. (1978). The power of theclassical twin study. Heredity, 40 (1), 97–116.
McCrae, R. R., & Costa, P. T. (1990). Personality in adulthood. New York: Guil-ford.
McCrae, R. R., & Costa, P. T. (1997). Personality trait structure as a humanuniversal. American Psychologist, 52, 509–516.
McCrae, R. R., Costa, P. T., Ostendorf, F., Angleitner, A., Hrebickova, M., &Avia, M. D., et al. (2000). Nature over nurture: Temperament, personality,and life span development. Journal of Personality and Social Psychology, 78,173–186.
McGue, M., Bacon, S., & Lykken, D. T. (1993). Personality stability and changein early adulthood: A behavioral genetic analysis. Developmental Psychology,29, 96–109.
McGue, M., & Bouchard, T. J. Jr. (1984). Adjustment of twin data for the effectsof age and sex. Behavior Genetics, 14, 325–343.
Neale, M. C. (1997).Mx: Statistical modeling. Richmond, VA: Medical College ofVirginia, Department of Psychiatry.
Neale, M. C., & Cardon, L. R. (1992).Methodology for genetic studies of twins andfamilies. Dordrecht, Nederlands: Kluwer Academic.
Pedersen, N. L., & Reynolds, C. A. (1998). Stability and change in adult person-ality: Genetic and environmental components. European Journal of Personal-ity, 12, 365–386.
Plomin, R., & Daniels, D. (1987). Why are children in the same family so differentfrom one another? Behavioral and Brain Sciences, 10, 1–60.
Plomin, R., DeFries, J. C., & McClearn, G. E. (1988). Nature and nurture duringinfancy and early childhood. New York: Cambridge University Press.
Plomin, R., DeFries, J. C., McClearn, G. E., & Rutter, M. (1997). Behavioralgenetics. New York: W. H. Freeman & Company.
Price, R. A., & Vandenberg, S. G. (1980). Spouse similarity in American andSwedish couples. Behavior Genetics, 10, 59–71.
Purcell, S. (2002). Variance components models for gene-environment interactionin twin analysis. Twin Research, 5, 554–571.
Reiss, D., Neiderhiser, J. M., Hetherington, E. M., & Plomin, R. (2000).The relationship code: Deciphering genetic and social influences on adolescentdevelopment. Washington, DC: George Washington University MedicalCenter.
Roberts, B. W., & Caspi, A. (2003). The cumulative continuity model of person-ality development: Striking a balance between continuity and change in per-sonality traits across the life course. In U. M. Staudinger & U. Lindenberger(Eds.) Understanding human development: Dialogues with lifespan psychology(pp. 183–214). Dordrecht, Netherlands: Kluwer Academic Publishers.
Roberts, B. W., & DelVecchio, W. F. (2000). The rank-order consistency of per-sonality from childhood to old age: A quantitative review of longitudinalstudies. Psychological Bulletin, 126, 3–25.
550 Johnson, McGue, & Krueger

Schachter, J. P., & Kuenzi, J. J. (2002). Seasonality of moves and the duration andtenure of residence: 1996. Washington, DC: Population Division, U. S. CensusBureau.
Srivastava, S., John, O. P., Gosling, S. D., & Potter, J. (2003). Development ofpersonality in early and middle adulthood: Set like plaster or persistentchange? Journal of Personality and Social Psychology, 84, 1041–1053.
Sutton, S. K., & Davidson, R. J. (1997). Prefrontal brain asymmetry: A biologicalsubstrate of the behavioral approach and inhibition systems. PsychologicalScience, 8, 204–210.
Tellegen, A. (1982). Brief manual for the Differential Personality Questionnaire.Minneapolis.
Tellegen, A. T., Bouchard, J., Jr., Wilcox, K. J., Segal, N. L., Lykken, D. T., &Rich, S. (1988). Personality similarity in twins reared apart and together.Journal of Personality and Social Psychology, 54 (6), 1031–1039.
Turkheimer, E., & Waldron, M. (2000). Nonshared environment: A theoretical,methodological, and quantitative review. Psychological Bulletin, 126 (1),78–108.
Viken, R. J., Rose, R. J., & Koskenvuo, M. (1994). A developmental-geneticanalysis of adult personality: Extraversion and neuroticism from 18 to 59.Journal of Personality and Social Psychology, 67, 722–730.
Zuckerman, M. (1991). Psychobiology of personality. Cambridge: CambridgeUniversity Press.
Personality Stability 551


![Epigenetic Upregulation of Corticotrophin-Releasing ......behavioral parameters in the adulthood [1]. Early postnatal ... tergic strength and synaptic plasticity, and modulated the](https://img.pdfslide.net/doc/110x75/61243ae65192913b7d1c78a7/epigenetic-upregulation-of-corticotrophin-releasing-behavioral-parameters.jpg)



























