Embed Size (px)
Citation preview

BioEssays Vol. 10, No. 6 - June 1989 179
Plant Viruses: A Tool-Box for Genetic Engineering and Crop Protection T. Michael A. Wilson
Sum ma ry
Traditionally, plant viruses are viewed as harmful, undesirable pathogens. How- ever, their genomes can provide several useful ‘designer functions’ or ‘sequence modules’ with which to tailor future gene vectors for plant or general biotech- nology.
The majority (77%) of known plant viruses have single-stranded RNA of the messenger (protein coding) sense as their genetic material. Over the past 4 years, improved in vitro transcription systems and the construction of partial or full- length DNA copies of several plant RNA viruses have enhanced our ability to manipulate and study their genomes, particularly in the context of their patho- genic interactions with host plants.
Recently, two forms of genetically engineered protection against plant virus infections have been reported. In both, a virus-related ‘ interfering’ molecule was stably introduced into plants via the DNA-transfer mechanism of Agro- bacterium tumefaciens. To date, the choice o f ‘ interfering’ molecule has been guided by empirical jield-observations and each is eflective against only a narrow range of closely-related viruses. As yet , we do not fully understand the molecular mechanism(s) responsible for the ob- served protection.
The ability to manipulate the plant- pathogen relationship is a powerful tool to increase our knowledge and improve
future strategies for unconventional crop- protection by genetic engineering tech- n iques .
Introduction
Plant virology has experienced a sig- nificant revival over the past 4-5 years. From being the ‘poor relation’ of molecular virology, plant virology has developed rapidly into one of the most exciting and innovative areas of con- temporary virus research. This can be attributed to some easily identified technical breakthroughs as well as an upsurge of interest and effort in plant molecular biology, biotechnology and the genetic manipulation of plants for resistance to disease.
We have an urgent need to under- stand and control pathogens of the few ‘core’ crops on which the world depends for the vast majority of its food, without the use of expensive or environmentally toxic chemicals. Plant viruses can cause serious losses in all the major crops of Man. Recent studies on the molecular biology of plant viruses, and the way in which they invade, subvert and exploit the sub-cellular machinery and metabo- lism of their particular host, have suggested new ways to protect plants against these pathogens.
Two novel areas of plant molecular virology which may help directly or indirectly in this goal are described here.
(a) Transplanting a short functional sequence from a plant virus genome into an alien genetic environment - thereby adding some desirable, func- tional module to ‘your favourite’ gene vector (spare-part genetic surgery, with a virus donor).
The converse, inserting a useful foreign gene into a suitable, non- essential site in the virus genome and obtaining highly amplified levels of expression, i.e. making a gene replacement vector - has been achieved successfully with both plant and animal viruses.
(b) Introducing anti-viral ‘inter- fering molecules’ into otherwise sus- ceptible host plants - thereby providing some measure of stable, inheritable protection against normally devastating virus infections.
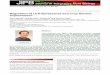
First, to introduce the 632 currently identified plant viruses, the most recent classification scheme based on particle morphology and the nature of the virus genetic material is shown in Fig. 1 (updated from ref. 1). Seventy-seven percent, or 484, of these viruses contain single-stranded (ss) messenger-sense RNA (mRNA) as their genetic material’ and lack any DNA intermediate form during their natural life-cycle. Serious technical limitations in handling ssRNA were a major drawback until 4 years ago when convenient and efficient in vitro transcription systems were devel- oped.3 These systems now permit one to manipulate an artificial DNA-copy of all or part of the RNA virus genome, using routine genetic engineering tech- niques, before running-off an RNA copy to be tested experimentally. The value of this technique should become clear below. In only one case,4 has a DNA copy of part of a multicomponent plant RNA virus been shown to be active in plants.
Subscriptions: BioEssays (ISSN 0265-9247) is published by Cambridge University Press. Two volumes of five parts each will be published in 1989. The subscription price includes air speeded delivery to most countries. Dollar prices apply to USA and Canada. Institutional subscriptions include a volume index. Individuals must order direct from the Press and certify that the journal is for their personal use. One volume €47.50 UK, €52.50 elsewhere ($99.50) for institutions. Two volumes f95.00 UK, €1 05.00 elsewhere ($1 99.00) for institutions, f44.00 (870.00) for individuals. !Single parts f8.00 ($15.00). Orders. which must be accompanied by payment, may be sent to a bookseller, subscription agent or direct to the publishers: Cambridge University Press, The Edinburgh Building, Shaftesbury Road, Cambridge CB2 2RU, UK. Orders from the USA or Canada should be sent to Cambridge University Press, 32 East 57th Street, New York, NY 10022, USA. Second class postage paid at New York, NY, and at additional mailing offices. POSTMASTER: send address changes in USA and Canada to BioEssays, Cambridge University Press, 11 0 Midland Avenue, Port Chester, New York, NY 10573. Back Volumes: Volumes 1-9 are available from Cambridge University Press.

180 BioEssays Vol. 10, No. 6 - June 1989
REVIEW ARTICLES
paq
0 Caulimovirus
0- 0 -1 c3
Geminivirus Reoviridae Cryptovirus
-0 Tobravi rus
Tobamovirus
M U Furovirus
on0 Hordeivirus
] without envelopes
Maize chlorotic dwarf vi.rus Luteovirus Tymovirus Tombusvirus Sobemovirus Necrovirus Carmovirus Marafivirus Parsnip yellow fleck virus
0
uu Co movi rus Fa bavi rus
Potexvirus Nepovirus Pea enation mosaic virus Dianthovirus
1
000 Carlavirus
Cucumovirus Bromovirus llarvirus
L Capillovirus
I - 1
0 000 Potyvi rus
t Alfalfa mosaic virus
Closterovirus
with envelopes
I Rhabdoviridae Tomato spotted wilt virus( 100 nm -
~~
(+), positive-strand genomic RNA, i.e. coding directly for protein; (-), negative-strand genomic RNA. Generously provided by Dr R. I. B. Francki and reproduced with his permission.
genetically transformed (transgenic) plants. The foreign gene and its adjacent CaMV 35s-RNA promoter are stably inherited in a conventional Mendelian fashion.
‘Gene Shears‘- Self-Cleaving RNA Isolates of some plant RNA viruses often contain a small (approx. 330-360 nucleotide-long) ‘parasitic’ or ‘hitch- hiker’ RNA molecule called a satellite RNA.*”vb These satellite RNAs require the supporting virus for multiplication (replication) and become packaged in the coat protein of the ‘helper’ virus.
Satellite RNA replication involves complex, repetitive head-to-tail RNA structures - concatameric satellite RNAs - which perform a unique, aston- ishing and unexpected function. The concatameric RNA spontaneously and precisely cuts itself into shorter pieces, monomeric satellite units of 330-360 nucleotides, without requiring any other biological catalyst (i.e. an enzyme). Only magnesium ions are required for this autolytic processing. No protein is
This is thought to be a very ancient process, probably one of the earliest reactions of nucleic acids to occur before ‘Life’ evolved as we recognize it.
To cleave itself, the satellite RNA forms an unusual folded structure de- scribed in two-dimensions as a ‘ham- merhead ’, in which the site of cleavage (or substrate) is positioned precisely next to the active part of the RNA molecule (the so-called ‘ribozyme’). The minimum functional seuences required to achieve this positioning and autolytic
Designer Functions Derived from Plant Viruses Plant-Active Promoter Sequence To date, probably the most widely exploited plant virus function has been the 35s-RNA promoter sequence from cauliflower mosaic virus (CaMV) - an unusual plant virus, in that the virions contain double-stranded DNA.l
As shown in Fig. 2, the 35s promoter of CaMV normally serves to make a more-than-full-length RNA copy of CaMV DNA inside the nuclei of in- fected cells5 using the native plant enzyme : DNA-dependent RNA poly-
merase 11. The CaMV 35s promoter was thought to be constitutively ex- pressed in plants, however recent evi- dence suggest that in roots and in cultured callus tissue the 35s promoter is less active (S. N. Covey, personal communication). This promoter and many foreign genes of interest have been transferred into chromosomal DNA in the plant cell nucleus using the natural DNA-transfer and integration process of the soil bacterium Agro- bacterium tumefaciens. High levels of foreign gene expression are then ‘driven’ by the CaMV 35s promoter in
19s H 3 5 ~ promoter \\ \ 35s -RNA
CaMV transcription
Fig. 2. Schematic diagram highlighting the tran- scriptional activities of the dsDNA of cauliflower mosaic virus (CaMV), shown as the inner thin double lines. Transcripts (ssRNA) are depicted as bold outer lines with arrowheads showing the direction of synthesis by the plant enzyme, DNA- dependent RNA polymerase 11. Both 35s- and 19s-RNA molecules share a common termination Ipoly(A)] signal. Diagram kindly provided by Dr S. N. Covey.

BioEssays Vol. 10, No. 6 -June 1989 181
REVIEW ARTICLES
Substrate A I 5’ x x x x x x x x x x x x x x x x x x x x x x x x x x x 3 ‘ / . ‘ * t t W T L 2
3‘ k x x x C
5’
Fig. 3. Generic model for the design of ‘gene shears’ (or ribozyme) based on viral satellite RNA sequences. Three structural domains (A-C) are boxed. A, minimum conserved sequence for the RNA substrate next to the cleavage site (arrowed) ; B, highly conserved sequence of the catalytic module (ribozyme); C, flanking complementary RNA sequences, forming base-paired double helices to juxtapose domains A and B. Reprinted by permission from Nurure, vol. 334, p. 587 and Dr J. Haseloff. . - Copyright 0 1988 Macmillan Magazines i t d .
activity are depicted in Fig. 3 and have been called ‘gene shears’. Recently the ‘gene shears’ have been introduced into other, foreign RNA molecules.1° Clearly the effect of ‘gene shears’, if genetically engineered into or targetted against a natural mRNA, would be to destroy its activity and thereby depress specific gene expression. Now that this has been shown to work in the test-tube,’O no doubt it will soon be demonstrated to function in intact cells and organisms.
The ability to switch-off harmful genes, selectively, is a long-standing goal of molecular biology. The ‘gene shears’, taken from a plant virus satellite system, may be one way to achieve this goal.
RNA - Packaging Signals - ‘A Molecular Suitcase Apart from some mutant strains of plant viruses and some sub-viral plant pathogens known as viroids,”, all vir- uses encapsidate their genetic material in a protective shell of virus-coded protein - the coat or capsid protein. The coat proteins of many animal viruses interact with complex and speci- fic host cell receptor(s). In contrast, plant virus capsid proteins appear to serve only a protective function during virus spread within a plant or between plants in the field. Environmental spread of plant viruses relies on natural agents such as insects, soil fungi, nema- tode worms, or even Man’s agricultural implements. There is no compelling evidence for any virus-specific receptor on or in any plant cell.
The assembly mechanism(s) of plant viruses are surprisingly poorly under- stood. Most is known about rod-shaped viruses with helical capsids, in particular tobacco mosaic virus (TMV). Here, the length of the virus particle is determined by, and directly proportional to, the size of its genomic RNA. In theory, an infinitely long virus-like structure is possible. In contrast, spherical or poly- hedral viruses have a limited capacity to fit nucleic acid inside their hollow protein shells.
To initiate the spontaneous self- assembly mechanism of TMV requires o d y a very short sequence of RNA,12“,” single stem-loop structure containing about 75 nucleotides, or 1.2% of the genome. This sequence is located intern- ally about 85 o/o of the way from the 5’- end of the viral genome, within the previously identified and longer origin- of-assembly sequence (OAS; Fig. 4).13 Provided the OAS (or the subsequence) was present, we predicted that any covalently attached ssRNA molecule should become encapsidated to create a virus-like particle ; a ‘ pseudovirus ’14 (Fig. 5). By packaging mRNA coding for a foreign reporter enzyme, we have also been able to quantify the protective effect of coat protein on RNA expressed during subsequent leaf inoculation. l5 ‘ Pseudovirus ’ particles are also useful probes with which to study the earliest structural events of plant cell infection - the site(s) and mechanism(s) of cell entry, virus particle disassembly and early (transient) gene expression.’6
Recently we genetically engineered a
DNA copy of the TMV OAS into the chromosomal DNA of tobacco plants. During systemic infection with TMV, reporter gene mRNAs exported from the plant cell nucleus were packaged and inactivated. In addition, we could recover large amounts of the particular mRNA in virus-like particles.” This is the first example of selective inhibition of plant nuclear gene expression and DNA transcript-rescue by tricking a virus into packaging plant mRNA.
Viral RNA Leader Sequences -
Plant RNA viruses adopt a number of strategies to achieve complete expres- sion of their genetic information within the infected cell. Full-length TMV RNA has long been known to be a remarkably efficient mRNA. At least in part, this must be due to its leader sequence which avidly recruits those host cell factors and ribosomal subunits involved in protein synthesis. We constructed a DNA copy of this untranslated 5’4eader sequence, attached it to various reporter genes and made hybrid mRNAs in the test-tube.ls-zo With all genes tested, and in almost every plant, animal (or even bacterial) cell system or cell-free extract we chose to use, the TMV RNA leader sequence significantly enhanced expres- sion of the attached reporter gene. The utility of this enhancer sequence is now being tested in about twenty labora- tories worldwide, in many different genetic engineering projects, where in- creased gene expression is required.
Groups in Harvard, Madison and at Monsanto (St Louis) have been per- forming analogous experiments with the 5’4eader sequence from an efficient short mRNA (RNA 4) from alfalfa mosaic virus (AlMV). Their results and conclusions,21.22 are similar to ours. However, in direct comparative experi- ments, we found that the TMV leader sequence was substantially more stimu- latory than that from AlMV RNA 4.
To summarize many months of work, it appears that the TMV leader is uniquely efficient because it remains single-stranded and accessible to recruit ribosomal subunits, no matter what gene sequence is placed downstream of itz3 (Fig. 6).
Translational Frameshifting - Viral Gene Fusing Sequence Another interesting ‘module’ from a plant RNA virus, which might prove useful in the future, is a sequence which ‘ splices’ two different protein-coding regions together and avoids a stop codon by causing ribosomes to frame-
Translational Enhancers ‘

182 BioEssays Vol. 10, No. 6 - June 1989
REVIEW ARTICLES
i i I \ I \ I
I
Fig. 4. Schematic representation of the structure and gene organization of tobacco mosaic virus (TMV) RNA (6395-nucleotides; bold line). TWO of the three possible reading frames are used, with corresponding gene products [sizes in kilodaltons (kDa)] depicted above or below the RNA. The 183 kDa protein arises by suppression or readthrough of the 'leaky' UAG stop codon terminating the 126 kDa polypeptide. The nucleotide sequence of the 5'-untranslated leader, including the start codon (AUG) for the 126 kDa protein, is shown expanded above. This can be released as a unique. ribonuclease T1-resistant fragment called 'omega' (Q). To initiate the bi-directional self-assembly of TMV, coat protein (the 17.5 kDa gene product) interacts with the so-called 'origin-of-assembly sequence' (OAS) on the RNA. As shown here, the OAS is located internally in the 30 kDa protein gene, is approx. 200-nucleotides long and can fold into 3 hairpin-loop structures. The minimum sequence (75 nucleotides) required for assembly is located to the 3' (right-hand) end of the longer @AS (ref. 12 a, b). The untranslated 3' terminal 204 nucleotides can be folded by complex 'pseudo-knotting', into a histidine-accepting transfer-RNA (tRNA)-like structure.
shift (-1 position) during protein syn- beet western yellows virus.zQn*b Hence, thesis. Such regions have been known under different translation conditions for several years in animal retroviruses in plant cells one could obtain either a (a group which contains some tumour short or an extended (fused-frame) viruses and the AIDS virus) and have protein, each with potentially different now been reported in two plant luteo- biological properties. viruses - barley yellow dwarf virus and
neo mRNA 'pseudovirions'
nm
Fig. 5. Laboratory-made. 'pseudovirus particles' resembling TMV but containing the messenger RNA (mRNA) for the bacterial reporter enzyme, neomycin phosphototransferase (neo). Neo mRNA was transcribed in uifro using an SP6-promoter plasmid (see refs. 14-23) and then packaged in TMV coat protein as described (ref. 14). Samples of rodlets were viewed in the electron microscope (left-side panel) by K. A. Plaskitt. A transcript approx. 1650-nucleotides long produces rodlets 77 nm in length ( I nm = ID-' m) as confirmed by the rod length-frequency distribution histogram on the right. Data kindly provided by Dr D . E. Sleal (Ph.D. Thesis, University of Liverpool).
Internal Sequences for Generating Subgenomic RNAs - Gene Repfacement Vectors Many plant RNA viruses have internal sequences which serve to promote the synthesis of a short, efficient sub- genomicmRNA during infectiomZ5 This process usually activates an otherwise silent, internal coding region. In at least two cases (with T M V and bromegrass mosaic virusz7), a foreign reporter gene (encoding chloramphenicol acetyltran- ferase) was engineered into a sub- genomic viral message position, re- placing a non-essential virus gene. The remainder of the virus still functioned normally to propagate and cleave-off the foreign gene message. An example of using a plant RNA virus as a self- amplifying, cytoplasmic gene-replace- ment vector.
As yet we do not know if such autonomous cytoplasmic RNA vectors will have any practical value. Due to a high error-frequency in the RNA-RNA replication process, there is uncertainty about the stability and durability of such modified RNA replicons. In partic- ular, there is unlikely to be a strong selection pressure to retain the foreign gene insert. To overcome this problem, ssDNA and dsDNA plant viruses - geminiviruses and caulimoviruses, re-

BioEssuys Vol. 10, No. 6 -June 1989 183
REVIEW ARTICLES
how switch-on an array of natural genes to alleviate the stress or to protect against infection. Some of these endo- genous gene products have a broad spectrum of activity and could restrict the spread of either a viral, bacterial or fungal invader. Since most plants are susceptible to only a few viruses, they must either have very effective natural defences, or fail to provide a suitable environment for the majority of patho- gens to multiply or spread. This is an area about which we still know very little in molecular terms, but which is attracting much attention in research laboratories throughout the world. Introducing these natural ‘defence ’ or ‘immunity ’ genes into crop plants which are at risk through intensive breeding programmes and growing conditions, is difficult. Not least because we still do not know what many of these genes are, where they are located on the chromo- somes, or how they are regulated.
An alternative approach, analogous to antibiotic therapy, is to introduce a specific anti-pathogen (or anti-virus) molecule into the plant itself. However, unlike bacteria or fungi, viruses use the host cell’s metabolism, therefore any attempt to kill viruses by anti-metabo- lites would also kill the cell. Clearly some unique feature of the virus life- cycle must be identified and ‘interfered with’ or inhibited. In practice this has mean that any genetically engineered plant resistance or protection is active against only a fairly narrow range of closely related viruses. Two successful examples of engineered protection, de- scribed below, have exploited classical, empirical field observations. As yet we do not fully understand how the ob- served resistance to a challenge virus infection works in molecular terms, but, by manipulating these model systems in subtle ways, we hope to reveal the molecular events involved.
Satellite Protection - RNA Interference As mentioned above, isolates of several plant RNA viruses contain a small satellite RNA.6a-b In some cases, the presence of the satellite reduces or ameliorates the symptoms caused by the associated ‘helper’ virus alone. The satellite may reduce the amount of damaging virus produced in infected cells although, in some combinations, less direct ‘ symptom-intereference ’ phenomena may also be involved.
For example, in China, a cucumber mosaic virus (CMV) isolate containing a benign natural satellite has been used to protect valuable tomato and pepper
B
A
TMV RNA 110
tl
E
AlMV-CAT
Fig. 6. Computer-predictions of the minimum free energy folding of chimaeridc mRNAs containing the genomic leader sequence (‘omega’) from TMV RNA (panels B, C), a random sequence devoid of G- residues of the same length as ‘omega’ (panel D), or the sub-genomic leader from alfalfa mosaic virus RNA 4 (panel E; see refs. 19, 21, 22). The uniquely unstructured nature of the TMV leader exists no matter what protein-coding sequence (mRNA) is placed downstream (3’) of it. Shown are those encoding neomycin phosphotransferase (NPTII ; panel B) or bacterial chloramphenicol acetyl- transferase (CAT; panel C). The position of the start codon (AUG) in each sequence is marked with a bold arrow. Nucleotides are number sequentially from the 5’-end and the equivalent predicted structure of the first 240 nucleotides of natural TMV RNA is shown for comparison (panel A). Modified from data published in ref. 23.
spectively - have recently been used as gene-replacement v e ~ t o r s . ~ ~ - ~ ’ Replica- tion of DNA has a much lower error- frequency because of enzymic proof- reading mechanisms. Successive genera- tions of mRNA, copied from the viral DNA, are therefore more likely to remain unchanged. Similar arguments apply to transgenic plants containing stable DNA copies of parts of RNA viruses (or any other gene).
Protein Engineering and Foreign Peptide Rescue Using Virus Coat Protein as a Carrier The last example of a ‘designer func- tion’ from the plant virus tool-box illustrates the power and ingenuity of genetic engineering to exploit a partic- ular desirable feature.
Under conditions of low pH, the coat protein of TMV self-assembles (without packaging RNA) into very long helical rods which can be purified easily. Recently, protein engineers used this self-assembling property to prepare
large amounts of an immunogenic pep- tide made in a bacterial culture.31
First, a synthetic TMV coat protein gene was constructed, to optimize the genetic code used in bacteria. This gene also encoded an additional, short (10 amino acid), carboxy-terminal se- quence from one of the coat proteins (VPI) of poliovirus Type 3. Large amounts of the hybrid protein were synthesized in a bacterial culture and later purified from crude cell extracts by encouraging the TMV coat protein portion to self-assemble at low pH. The concentrated hybrid protein was then used to immunize rats against intact, infectious poliovirus. The efficiency of the rats’ immune response was directly related to the size of the protein complex used to raise the polio-neutralizing antibodies.
Crop-Protection via Engineering Virus-Specific Interfering Molecules
When stressed or wounded, plants some-

184 BioEssays Vol. 10, No. 6 ~ June 1989
REVIEW ARTICLES
plants against severe strains of CMV. A similar phenomenon can be demon- strated in the laboratory in tobacco plants.
Two years ago, at the Plant Breeding Institute in Cambridge, a dimeric DNA copy of an attenuating strain of CMV satellite RNA was genetically engin- eered into tobacco cells.32 Subsequent infection with CMV or a closely-related virus, tomato aspermy virus (TAV), greatly amplified the endogenous level of satellite RNA which then sub- stantially reduced the symptoms caused by the same ‘helper’ virus. Control plants which lacked the genetically engineered satellite became severely dis- eased. The molecular mechanism for this observation is still largely unknown but the ability to achieve this form of plant protection opens up exciting re- search possibilities.
Parallel studies in California (G. Bruening, personal communication) with genetically engineered walnut trees expressing the satellite RNA of tobacco ringspot virus (STobRV) have shown a significant level of protection against a devastating disease of this important crop caused by cherry leafroll virus (CLRV) which is transmitted by nema- tode worms in infested soils.
Unfortunately, a satellite RNA which ameliorates the symptoms of one virus may exacerbate those of another virus in the same crop. Relatively few viruses in nature have associated satellites. Also, different strains of a satellite can cause more or less severe symptoms with a particular virus. Given the high mutation rate of RNA-RNA repli- cating systems and the risk of field- infection by another virus, one with which the endogenous satellite could cause a more severe infection, there are clearly major environmental drawbacks to the widespread growth of transgenic plants expressing a complete satellite RNA. However, work is underway to identify a small part of the satellite sequence which could be used for crop-protection without these attendant risks.
Engineered Resistance Via Viral Coat Protein In 1929, M ~ K i n n e y ~ ~ demonstrated that primary infection with a mild strain of a virus could protect a plant against later challenge with a more severe strain of the same or a closely related virus. The phenomenon was called ‘cross-protec- tion’ and can also be shown to apply to viroids (autonomous, small, naked cir- cular RNAs) and coat protein-free mutants of viruses. This suggests that
‘interference ’ between pathogens can operate at several levels.
In the last 10 years, cross-protection between intact viruses has been shown to depend upon the relatedness of the coat proteins of the two viruses and the fact that the second, challenge inoculum must contain virus and not naked viral RNA. To date, the most likely ex- planation for this is that the coat protein of the first (mild) strain prevents par- ticles of the severe, challenge virus strain from uncoating after they have entered the plant cell. Using our laboratory made ‘pseudovirus ’ particles, we are currently investigating this hypothesis.
Two years ago, genetically engineered tobacco plants expressing suitably high levels of TMV coat protein were pro- duced by Professor R. N. Beachy in St Louis.34 While these plants exhibited significant resistance to challenge inocu- lations with TMV particles, naked TMV RNA was infectious. Because an early structural event during virus infection appears to be inhibited,35*36 we are collaborating with Beachy to discover why their plants are protected against TMV infection. One curious recent observation is that these coat protein transgenic tobacco plants contain low concentrations of ribonucleoprotein rodlets prior to virus inoculation (Fig. 7). The significance and nature of these structures is currently under investi- gation.
Since 1986, several groups in America and Holland have published analogous results with transgenic tobacco plants expressing one of several different virus coat proteins. In all, five different coat- protein-virus-protection systems have now been No doubt many more are on the way!
Last year, Beachy and his colleagues at Monsanto were the first to apply this technology in the field.43 Transgenic tomato plants expressing the TMV coat protein gene were field-tested in Illinois and found to be resistant to TMV, tomato mosaic virus (ToMV) and two strains of ToMV which usually over- come a natural virus resistance gene (Tm-2) bred into commercial tomato lines. The agronomic characteristics (leaf weight, fruit yield, etc) of the transgenic plants were the same as control plants. However, after challenge virus inoculation the fruit yield of the controls fell by 25-35 %, whereas the transgenic plants were unaffected.
Clearly research on engineered virus resistance has an immediate practical application. Unlike satellite virus RNA, expression of a viral coat protein gene at relatively low levels in crop plants
creates minimal environmental risk. Doubtless we all ingest plant viruses and their proteins regularly in our fruits and vegetables without any ill-effects. Even so, Beachy’s group are currently searching for a sub-fragment of the TMV coat protein gene which might still confer protection against virus infection and have even less capacity to recombine or otherwise interact un- desirably with an invading virus.
Other Routes To Crop-Protection Using Viral Sequences For the future, other ‘interfering’ mole- cules which could be engineered into plants include those called anti-sense RNA - complementary RNA copies of viral sequences required for replica- t i ~ n ~ ~ (i.e. the termini of the viral RNA). Arguably, the anti-sense RNA mole- cules should bind to an incoming virus genome, forming a short RNA:RNA duplex and block any further virus function/multiplication. However, one drawback of this approach is that large numbers of virus particles enter wounded plant cells and soon begin replicating. Any such protection strat- egy must ensure the inhibition of every incoming virus genome. If applied to animal cells, the resulting double-stran- ded RNA fragment might than act to induce the natural defence mechanisms involving interferon.
Another strategy would be to saturate the cells with positive-sense competitor R N A s ~ ~ to remove an essential virus- coded enzyme (e.g. the replicase). Fur- ther more, the competitor RNA could be designed to be similar to a satellite RNA, in that it became highly amplified only when the particular ‘ helper’ virus, which it attenuated, had invaded the cell.
Following the elegant work on ‘gene shears’ described above,“’ it seems certain that several groups will utilize this approach to transform and protect plants against invasion by viruses. Tar- getting a ‘ribozyme’ to some essential part of the pathogenic genome.
Concluding Remarks
From the examples of ‘designer func- tions’ and anti-virus ‘interfering mole- cules’ described above, the flexibility and ingenuity of genetic engineering and its application to unconventional crop protection should have become clear. Molecular plant virology is ad- vancing rapidly by this approach. As we have seen, viruses have many efficient and useful biological functions or mod-

BioEssays Vol. 10, No. 6 - June 1989 185
REVIEW ARTICLES
TMV CP antiserum-coated grids
Fig. 7. Selected electron micrographs of endogenous ‘pseudovirions’ isolated from the sap of uninoculuted transgenic tobacco leaves which express the coat protein gene of TMV (transformed tobacco lin 3404 (8), described in ref. 34). Leaves were crushed in buffer at pH 6.5 (upper panels) or pH 8.0 (lower panels). centrifuged briefly ( 5 min at 10000 g) and the supernatmts allowed to react with polyclonal rabbit anti-TMV coat protein antiserum-coated carbon films on EM grids for several hours. At pH 8.0, TMV protein-only helices would not survive extraction. Thus we believe the stable rodlets shown contain RNA(s) of yet unknown identity. In the lowest panels, the identity of the protein component is confirmed by sequential labelling with a monoclonal antibody (MAb) which reacts with an epitope present only in ribonucleoprotein helices of TMV coat protein (a generous gift from M. H. V. Van Regenmortel, Strasbourg). MAb binding was then visualized by reaction with a gold- conjugated second antibody.
ules to offer, in contrast to their less- desirable pathogenic effects on plants.
These are exciting times for plant biotechnology in general, and for plant virology in particular - almost every week some major technical innovation or new insight into plant-pathogen interactions is reported. Only by in- creasing our knowledge in this way can we hope to understand, control and perhaps even exploit virus diseases in the future.
REFER ENCES 1 FRANCKI, R. 1. B., MILNE, R. & HAITA, T. (1985). Atlus oJ Plant Viruses, vols. 1 and 2. CRC Press, Boca Raton, Fla.
2 ZAITLIN, M. & HULL, R. (1987). Plant virus-host interactions. Ann. Rev. Plant Physiol. 38. 291-315. 3 MELTON, D. A. KRIEG, P. A,, REBAGLIATI, M. R., MANIATIS, T., ZINN, K. & GREEN, M. R. (1984). Efficient in uifro synthesis of biologically active RNA hybridisation probes from plasmids containing a bacteriophage SP6 promoter, Nucl. Acid.7 Res. 12, 7035-7056. 4 DORE, J.-M. & PINCK, L. (1988). Plasmid DNA containing a copy of RNA3 can substitute for RNA3 in alfalfa mosaic virus RNA inocula. J. G m . Virol 69, 1331-1338. 5 HULL, R. & COVEY, S. N. (1985). Cauliflower mosaic virus: pathways of infection. EioEssay.~ 3, 160-163. 6 ( a ) FRANCKI, R. 1. B. (1985). Plant virus satellites. Annu. Reu. Microhiol. 39, 151--174. (h ) SIMON, A. E. (1988). Satellite RNAs of plant viruses. Plant Molec. Biol. Reporter 6, 24CL252. 7 BUZAYAN, J. M., GFRLACH, W. L. & BRUENING, G. (1986). Non-enzymatic cleavage and ligation of RNAs complementary to a phnt virus satellite RNA. Nature 323, 349-353. 8 BUZAYAN, J. M., GERLACH, W. L. & BRUENING, G. (1986). Satellite tobacco ringspot virus RNA: A subset
of the RNA sequence is sufficient for autolytic processing. Proc. Natl. Acud. Sci. USA. 83, 8859-8862. 9 FORSTER, A. C. & SYMONS, R. H. (1987). Self-cleavage of plus and minus RNAs of a virusoid and a structural model for the active sites. Cell 49, 21 1-220. 10 HASELOFF, J. & GERLACH, W. L. (1988). Simple RNA enzymes with new and highly specific endoribonuclease activities. Nuturr 334, 585-591. 11 DIENER, T. 0. (1987). The Viroids. In The Viruws (series eds. H. Fraenkel-Conrat & R. R. Wagner). Plenum Press, New York. 12(a) TURNER, D. R. (1987). Protein-RNA interactions in tobacco mosaic virus assembly. Ph.D. Thesis, Uni- versity of Cambridge. 12(h) TURNER, D. R., JOYCE, L. E. & BUTLER, P. J. G. (1988). The tobacco mosaic virus assembly origin. Functional characteristics defined by directed muta- genesis 1. Mol. Biol. 203, 531-547. 13 ZIMMERN, D. (1977). The nucleotide sequence at the origin for assembly on tobacco mosaic virus RNA. Cell 11, 463482. 14 SLEAT, D. E., TURNER, P. C., FINCH, J. T., BUTLER, P. J. G. & WILSON, T. M. A. (1986). Packaging of recombinant RNA molecules into pseudovirus particles directed by the origin-of-assembly sequence from tobacco mosaic virus RNA. Virology 155. 299-308. 15 GALLIE, D. R., SLEAT, D. E., WATTS, J. W., TURNER, P. C. & WILSON, T. M. A. (1987). In uiuo uncoating and efficient expression of foreign mRNAs packaged in TMV-like particles. Science 236, 1122-1 124. 16 PLASKITT, K. A., WATKINS, P. A. C., SLFAT, D. E., GALLIE, D. R., SHAW, J. G. & WILSON, T. M. A. (1988). Immunogold labeling locates the site of disassembly and transient gene expression of tobacco mosaic virus-like pseudovirus particles in uit’o. Molecular Plnnf-Microhe Inrernctions 1, 10- 16. 17 SLEAT, D. E., GALLIE, D. R., WATTS, J. W., DEOM. C. M., TURNER, P. C., BEACHY, R. N. & WILSON, T. M. A. (1988). Selective recovery of foreign gene transcripts as virus-like particles in TMV-infected transgenic tobaccos. Nucl. Acids Res. 16, 3127-3140. 18 GALLIE, D. R., SLEAT, D. E., WATTS, J. W., TIJRNI.R, P. C. & WILSON, T. M. A. (1987). The 5’-leader sequence of tobacco mosaic virus RNA enhances the expression of foreign gene transcripts in oitro and in r i m Nucl. Arid.7 Res. 15, 3257-3273. 19 G A L L I ~ , D. R., SLEAT, D. E., WATTS, J. W., TURNLR, P. C . & WILSON. T. M. A. (1987). A comparison of eukaryotic viral Yleader sequences as enhancers of mRNA expression in riro. Nucl. Acids Re.\. 15, 8693-871 I . 20 SLEAT, D. E., GALLIE, D. R., JEFFERSON. R. A,, BEVAN, M. W., TURNER. P. C. & WILSON, T. M. A. (1987). ChardCterisdllOn of the 5’-leader sequence of tobacco mosaic virus RNA as a gencral enhancer of translation in iirro. Gene 60, 211-225. 21 JORLING, S. A. & GEHRKI., L. (1987). Enhanced translation of chimaeric messenger RNAs containing a plant viral leader sequence. Nuture 325, 622 625. 22 JOBLING, S. A,, CUTHB~RT, C. M., ROGERS, S. G., FRALEY, R. T. & OLHRKI-., L. (1988). In I i fro transcription and translational efficiency of chimeric SP6 messenger RNAs devoid of 5’-vector nucleotides. Nucl. Acirls Rr.v. 16, 44834498. 23 SLEAT. D. E.. HULL, R., TURNER, P. C. & WIIXJN, T. M. A. (1988). Studies on the mechanism of translational enhancement by the 5’-leader sequence of tobacco mosaic virus RNA. Eur. J . Biochem. 175, 75-86. 24 (u ) MILLER, W. A,, WATERHOUSE, P. M. & GERLACH, W. L. (1988). Sequence and organization of barley yellow dwarf virus genomic RNA. Nucl. Acids Res. 16, 6097411 1. (h) VEIDT, I., LOT, H., LEISER, M., SCHEI- DCCKER, D.. GUILLFV, H., RICHARDS, K. & JONARD, C . (1988). Nucleotide sequence of beet western yellows virus RNA. Nucl. Acids Rex 16, 991 7-9932. 25 FRENCH. R. & AHLQUIST, P. (1988). Characterization and engineering of sequences controlling in oiao synthesis of brome mosaic virus subgenomic RNA. J . Virol. 62. 24 I 1-2420. 26 TAKAMATSU, N., ISHIKAWA, M., MESHI, T. & OKADA. Y. (1987). Expression of bacterial chloramphenicol acetyltransferase gene in tobacco plants mediated by TMV-RNA. EMBO J . 6, 307-311. 27 FRENCH, R., JANDA, M. & AHLQUIST, P. (1986). Bacterial gene inserted in an engineered RNA virus: Efficient expression in monocotyledonous plant cells. Science 231, 1294- 1297. 28 WARD, A,, ETESSAMI, P. & STANLFY, J. (1988). Expression of a bacterial gene in plants mediated by infectious geminivirus DNA. EMBO J . 7. 1583- 1587.

186 BioEssays Vol. 10, No. 6 -June 1989
REVIEW ARTICLES
29 HAYES, R. J., PETTY, I. T. D., COUTTS, R. H. A. & BUCK, K. W. (1988). Gene amplification and expression in plants by a replicating geminivirus vector. Nature 334, 179-182. 30 BRISSON, N., PASZKOWSKI, J . , PENSWICK, J . R., GRONENRORN, B., POTRYKUS, I . & HOHN, T. (1984). Expression of a bacterial gene in plants by using a viral vector. Nature 310, 51 1-514. 31 HAYNES, J. R., CUNNINGHAM, J., VON SEEPRIED, A., LENNICK, M., GARVIN, R . T . & SHEN, S.-H. (1986). Development of a genetically-engineered, candidate polio vaccine employing the self-assembling properties of the tobacco mosaic virus coat protein. BiolTechnology 4, 637-641. 32 HARRISON, B. D., MAYO, M. A., & BAULCOMRE, D. C. (1987). Virus resistance in transgenic plants that express cucumber mosaic virus satellite RNA. Nulure 328, 799-802. 33 MCKINNEY, H. H. (1929). Mosaic diseases in the Canary Islands, West Africa and Gibralter. J. Agrirult. Res. 39, 551-578. 34 POWELL AREL, P., NELSON, R. S., DE, B., HOPFMANN, N., RWERS, S. G., FRALEY, R. T. & BEACHY, R . N. (1986). Delay of disease development in transgenic plants that express the tobacco mosaic virus coat protein gene. Science 232, 738-743. 3 5 NELSON, R. S.. PowrLL ABEI., P. & BCACHY, R. N. (1987). Lesions and virus accumulation in inoculated transgenic tobacco plants expressing the coat protein gene of tobacco mosaic virus. Virology 158, 126-132.
36 REGISTER, J . C. & BEACHY', R. N. (1988). Resistance to TMV in transgenic plants results from interference with an early event in infection. Virology 166, 524-532. 37 TUMEX, N. E., OCONNELL, K. M., NELSON, R. S., SANDERS, P. R., BEACHY, R. N., FRALEY, R. T. & SHAH, D. M. (1987). Expression of alfalfa mosaic virus coat protein gene confers cross-protection in transgenic tobacco and tomato plants. EMBO J . 6, 1181-1189. 38 LOESCH-FRIES, L. S., MERLO, D., ZINNEN, T., BUR- HOP, L., HILL, K., KRAHN, K. , JARVIS, N., NELSON, S. & HALK, E. (1987). Expression of alfalfa mosaic virus RNA4 in transgenic plants confers virus resistance. EMBO J. 6, 1845-1851. 39 VAN DUN, C. M. P., BOL, J. F. & VAN VLOTEN- DOTING, L. (1987). Expression of alfalfa mosaic virus and tobacco rattle virus coat protein genes in transgenic tobacco plants. Virology 159, 299-305 40 VAN DUN, C. M . P., OVERDUIN, B.. VAN VLOTEN- D o ~ G , L. & B ~ L , J. F. (192%). Transgenic tobacco expressing tobacco streak virus or mutated alfalfa mosaic virus coat protein does not cross-protect against alfalfa mosaic virus infection. Virology 164, 383-389. 41 Cuozzo, M. , OCONNELL, K. M., KANIEWSKI, W., FANG, R.-X., CHUA, N.-H. &TUMER, N. E. (1988). Viral protection in transgenic tobacco plants expressing the cucumber mosaic virus coat protein or its antisense RNA. BiolTechnology 6, 549-551. 42 HEMENWAY, C., FANG, R.-X., KANIEWSKI, W. K., CHUA, N.-H. & TUMER, N. E. (1988). Analysis of the mechanism of protection in transgenic plants expressing
the potato virus X coat protein or its antisense RNA. EMBO J. 7 , 1273-1280. 43 NELSON, R. S., MCCORMICK. S. M., DELANNAY. X., DURE, P., LAYTON, J., ANDERSON, E. J., KANIEWSKA, M., PROKSCH, R. K., HORSCH. R. B., ROGERS, S. G. , FRALEY, R. T. & BEACHY, R. N. (1988). Virus tolerance, plant growth, and field performance of transgenic tomato plants expressing coat protein from tobacco mosaic virus. BiolTechnology 6, 403-409. 44 CRUM, C., JOHNSON, J. D., NELSON, A. & ROTH, D. ( I 988). Complementary oligodeoxynucleotide mediated inhibition of tobacco mosaic virus RNA translation in vitro. Nucl. Acids Res. 16, 45694581. 45 MORCH, M. D., JOSHI, R. L., DENIAL, T. M. & HAENNI, A. L. (1987) A new 'sense' RNA approach to block viral RNA replication in uitro. Nucl. A d s Res. 15, 41234130.
T M I C H A E L A WILSON isat the Department of Virus Research, John InneJ Institute and AFRC Institute of Plant Science Research, Colney Lane, Norwzrh NR4 l U H , U.K
The Carbonic Anhydrases: Widening Perspectives on Their Evolution, Expression and Function Richard E. Tashian
Sum ma ry
Now, some 55 years after its discooery in bovine red cells, carbonic anhydrase (CA), in all its variedforms, continues to challenge and intrigue physiologists, hio- chemists and molecular geneticists. This is so because oJ an increasing awareness of the many appurently dicerse,functions of the diflerent CA isozymes encoded by this large multigene family, the con- tinuing discovery of new CA, or CA- related, genes, and the extensive variation in their hormonal control, cellular ex- pression and subcellular localization.
Introduction
The carbonic anhydrases (CA) are zinc metalloenzymes that catalyze the simple interconversion of CO, and HC0,- (CO, + 0, $ HC0,- + H+). They are found in almost all organisms, and are notable for the extremely high turnover numbers (some exceeding 1 x lofi sec-') of the high-activity forms, ranking them along the most efficient enzymes known. They are also distinctive because of the
extensive diversity in both the cellular distribution and in the putative or established biological functions of the seven CA isozymes now known to occur in higher vertebrates. Some of the CA genes are expressed in certain cells of nearly all tissues, whereas others appear to be more limited in their distribution. In view of such diversity in expression and function, the CA isozyme system is an excellent model for the study of molecular processes and patterns under- lying the evolution and expression of genes derived from a common ancestor. (for recent accounts, see papers in refs. 1 and 2.)
Evolutionary Patterns
Since the reaction catalyzed by carbonic anhydrase may well have been of wide- spread adaptive value to the earliest organisms, it is possible that it was among the first enzymes to appear. Since what seem to be evolutionarily related (homologous) forms of carbonic anhydrase are found in certain bacteria and algae,3 the original gene probably
arose before the divergence of pro- karyotes and eukaryotes more than one and a half billion years ago. And during their long evolutionary history the CA genes have undergone many rounds of duplications resulting in new genes, the products of which have been implicated in such diverse processes as calcification, photosynthesis, respiration, acid-base homeostasis, bone resorption, forma- tion of aqueous humor and gastric juice, and the synthesis of urea, glucose and lipids.lI2
The Genes In terrestrial vertebrates (i.e. amniotes) the carbonic anhydrase system com- prises at least 7 genes (Table I). Four of these (coding for CA I, CA 11, CA III and CA VII) have now be fully or partially characterized,"R and with the exception of human CA I, which has a large intron in the 5' untranslated region,* they all appear to have seven exons, with the six introns at the same positions, and range in size from 9.8 kb (human CA VII) to 17 kb (chicken and