Embed Size (px)
Citation preview

J. Cell Sci. 42, 401-415 (1980)Printed in Great Britain © Company of Biologists Limited jg8o
PLASMA MEMBRANE ULTRASTRUCTURE
DURING PLANT PROTOPLAST PLASMOLYSIS,
ISOLATION AND WALL REGENERATION:
A FREEZE-FRACTURE STUDY
M. J. WILKINSON AND D. H. NORTHCOTEDepartment of Biochemistry, University of Cambridge, Tennis Court Road,Cambridge, CBz \QW, U.K.
SUMMARY
The freeze-fracture morphology of the plasma membrane of cells and isolated protoplasts ofplant callus suspensions has been investigated. Plasmolysis of suspension cells leads to theformation of 2 types of hexagonal arrays of intramembrane particles situated on the innerfracture face (PF). These arrays are interpreted as proteins that have ' crystallized' in the planeof the membrane as the area of surrounding lipid bilayer is reduced during protoplast retractionfrom the cell wall. Time-course studies have revealed no positive relationship between thedistribution of hexagonal arrays and the occurrence of microfibrils regenerated around isolatedprotoplasts during periods of culture. No evidence for the specialized transport functionsattributed to hexagonal arrays of plant cells by previous workers has been found.
INTRODUCTION
Cellulose microfibrils of plant cell walls are probably synthesized at the plasmamembrane. The molecular mechanism by which this membrane elaborates an intricatemeshwork of microfibrils, forming a coherent wall with associated matrix poly-saccharides and proteins, is poorly understood.
Freeze-fracture replication is an excellent means of visualizing the macromolecularorganization of biological membranes and the wall architecture of plant cells. Studiesof the unicellular algae Oocystis opiculatum have demonstrated a relationship betweenthe distribution of particles in the plasma membrane and wall microfibrils (Brown &Montezinos, 1976; Montezinos & Brown, 1976). Such clearly defined spatial associa-tions between plasma membrane components and nascent microfibrils of higher plantcells have not yet been identified, though a number of studies on isolated plantprotoplasts undergoing wall regeneration have been conducted (Willison & Cocking,1972, 1975; Grout, 1975; Willison & Grout, 1978). The majority of these investi-gations employed isolated leaf mesophyll protoplasts of tomato and tobacco. Incontrast, we have studied the plasma membrane ultrastructure of protoplasts isolatedfrom fast-growing callus cell suspensions. This material was found to possess areas ofregularly arranged intramembrane particles, a feature limited to the plasma membrane.While this investigation was in progress particle arrays were reported as charac-teristic of the plasma membrane of protoplasts isolated from Skimmia japonica callus

402 M. J. Wilkinson and D. H. Northcote
tissues (Robenek & Peveling, 1977). It was suggested that particle arrays resultedfrom the incorporation of specific proteins from the endoplasmic reticulum into theplasma membrane during wall regeneration. Arrays were interpreted as formingspecialized transport channels for the outward passage of wall precursors. Ourinvestigations of the factors involved in the production of particle arrays and of theirpossible role in wall regeneration leads us to an alternative explanation of theirpresence in the plasma membrane of isolated protoplasts.
MATERIALS AND METHODS
Growth of cells
Callus was initiated from tuber explants of potato (Solatium tiiberosum cv. Record) after themethod of Anstis & Northcote (1973). Solid callus was subcultured every 4-6 weeks on PRL4medium (Gamborg, 1966) containing 6 mg/1. 2,4-dichlorophenoxyacetic acid (2,4-D). Suspen-sion cultures were established by dispersing 2 g of solid callus in 100 ml liquid medium (eitherPRL4 or the medium described by Upadhya, 1975) containing 3 mg/1. 2,4-D, 2 g/1. N.Z.Amine (KW-Revai Chemicals Ltd., London, U.K.) and incubating at 100 rev/min in anorbital shaker at 26 °C. Suspension cultures grew slowly for several weeks but thereafter couldbe maintained by subculturing approximately half the cells into 100 ml of fresh growth medium,every 5-7 days. Cultures comprising large numbers of small, round cells growing in fineclumps were most suitable for protoplast isolation and could be obtained by subculturing at3-day intervals for at least 2 weeks. These fast growing cultures were used throughout thisinvestigation. They reduced the original pH of the growth medium (5-8-6-0) to approximatelypH 5-0 by the end of the 3-day growth period.
Protoplast isolation
Pectinase (Sigma, Poole, Dorset, U.K.) was purified before use (Hanke & Northcote, 1974).Driselase (a mixture of cellulase and pectinase obtained from Kyowa Hokko Kogyo Co.,Tokyo, Japan) was used as supplied. Insoluble components were removed by centrifugation at12000 g for 10 min prior to sterile filtration of enzyme solutions (Millipore filter, O'4S-/tm poresize).
Cells were filtered onto 25-/tm steel mesh and washed once with fresh growth medium.They were mixed with enzyme solution (3 g fresh weight of cells to 10 ml solution) com-prising 5 % (w/v) Driselase, 05 % (w/v) purified pectinase and 0-35 M sorbitol as the plasmo-lysing agent. This was prepared in growth medium (excluding sucrose, N.Z. Amine and2,4-D) buffered with 20 mM 2 iV-morpholino ethanesulphonic acid (MES) at pH 5-7. Themixture was gently agitated on a reciprocal shaker for 2-5-3 h at 25 °C to effect plasmolysis,wall digestion and release of protoplasts. The crude protoplast preparation was then filteredthrough a 7-cm-wide, 25-/«n steel mesh. Isolated protoplasts, with diameters up to approxi-mately 70 /(m passed through the mesh by constriction. They were centrifuged at 80 g for3 min and then washed 3 times with osmoticum. Viability was estimated by staining withfluorescein diacetate (Larkin, 1976).
Protoplast culture
After washing, protoplasts were resuspended to a density of approximately 5 x io5/ml infresh growth medium containing 0-35 M sorbitol and 1 g/1. N.Z. Amine. They were dispensedin o-5-ml aliquots in 50-ml plastic pots (Sterilin Ltd., Teddington, Middlesex, U.K.) in thedark at 25 CC. The presence of cellulose was detected by staining with 0-2 % (w/v) CalcofluorWhite (American Cyanamid Co., New Jersey, U.S.A.).

Plasma membrane of plant protoplasts 403
Digestion of the walls of prefixed cells
Cells were first fixed in 1 % glutaraldehyde prepared in growth medium at pH 50 (i.e. thepH to which it had been conditioned by the rapidly growing cells) for 15 h at 25 °C. Moreglutaraldehyde was then added to bring the concentration to 25 % and the pH readjusted to5-0. Fixation was continued for a further 4 h at 25 °C. Excess fixative was removed by rinsingthe cells 3 times with 100-ml quantities of growth medium. The fixed cells were then treated ina similar manner to that used to isolate plasmolysed protoplasts, except that sorbitol wasomitted from the enzyme solution and wall digestion was continued for 4 h. After cell wallremoval fixed protoplasts retained their original shape and could not be purified by constrictionthrough fine mesh. Wall fragments were removed by several washes in growth medium bycentrifugation at 80 g for 3 min.
Cell plasmolysis
To test the effect of plasmolysis alone on the organization of the plasma membrane, cells(2 g fresh weight) were placed in 10 ml growth medium containing 0-35 M sorbitol and 10 mMMES at pH 50 or 5-7 for 25 h. The effect of using 035 M glucose or a salt mixture (i'7S %KC1, C75 % MgCl2, 05 % KNO3) in place of sorbitol for plasmolysis was also examined.
Freeze-fracture replication and electron microscopy
Frceze-fracture replication was performed using a method developed specifically for thisstudy (Wilkinson, 1978; Wilkinson & Northcote, 1980). Briefly, a small volume of cells orprotoplasts, either alive or prefixed, was sandwiched between copper plates and frozen veryrapidly in liquid propane. The samples were then loaded into a hinge device and fractured byseparating the plates under high vacuum in a Balzers BA360 freeze-etching unit. Replicationwas by tungsten—tantalum evaporation using an electron gun.
Replicas mounted on gold grids were cleaned with 70 % (v/v) H2SO4 for 14-48 h at 35 °Cwith the aid of a glass capillary apparatus. In experiments on wall regeneration etching wasperformed prior to replication on protoplast samples that had been fixed in 2-5 % glutaraldehydefor 2-3 h at 25 °C or overnight at 2 CC followed by washing with distilled water. Etching wascarried out at a pressure of less than 133 /tPa (133 x io"1 N m~2) and at —100 °C for 3 min.
Replicas were examined using an AEI 801 transmission electron microscope operating at60 kV. Optical diffraction patterns were obtained using an optical diffractometer equipped witha laser source (Home & Markham, 1973). All reciprocal lattice points were used for imagereconstruction.
RESULTS
The appearance of the plasma membrane of callus cells before and after plasmolysis
Freeze-fracture replication of turgid suspension cells exposed only a small pro-portion of the total surface area of the plasma membrane. After enzymic digestion ofthe cell wall, however, the fracture characteristics of the remaining fixed protoplastswere similar to those of live plasmolysed protoplasts, with greater areas of plasmamembrane exposed for replication and electron-microscope examination. Theappearance of the plasma membrane of turgid suspension cells was typical of freeze-fractured biological membranes. Intramembrane particles, most prevalent on theinner fracture face (PF; Branton et al. 1975) occurred in an apparently randomdistribution (Fig. 1). The ultrastructure of the plasma membrane of cells sampledfrom growth medium at pH 5-0 was identical to that of cells that had been transferredto medium adjusted to pH 57 (i.e. the pH used for protoplast isolation) for 3 h.
M. jf. Wilkinson and D. H. Northcote

Plasma membrane of plant protoplasts 405
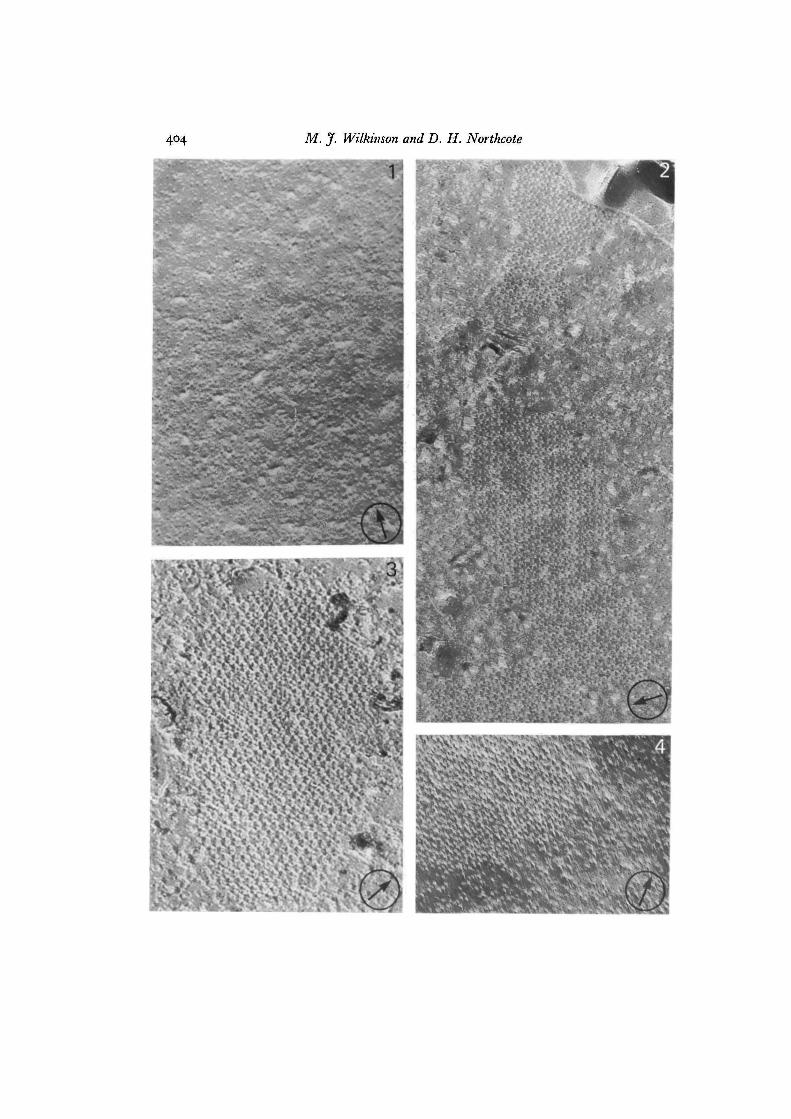
The plasma membrane of cells plasmolysed in growth medium containing 0-35 Msorbitol at pH 5-0 was similar to that of unplasmolysed control samples and exhibitedrandomly spaced particles. When plasmolysis was performed at pH 57, however,particles clustered to form discrete arrays (Fig. 2). These became apparent within30 min of transferring cells to plasmolyticum, that is, by the time plasmolysis wascomplete. The arrays were irregular in outline and comprised rows of particles (approx.10 nm diameter) located on the inner half (PF) of the plasma membrane. Particlearrays were formed regardless of which osmoticum was used to plasmolyse the proto-plasts, i.e. sorbitol, glucose (Fig. 3) or salts. Arrays were evident in freeze-fixedspecimens, where no pretreatment with fixatives and cryoprotectants was used, and inthose fixed with glutaraldehyde and then cryoprotected with 20% glycerol beforefreeze-fracturing (Fig. 4).
The appearance of the plasma membrane of isolated protoplasts
Fast-growing potato callus cells treated simultaneously with a plasmolysing agentand a cellulase-pectinase mixture yielded large numbers of viable protoplasts in 2-3I1(approx. ioB/g fresh weight of cells). The viability of protoplast preparations was90-100%. The appearance of the plasma membrane was unaltered by treating proto-plasts with wall-degrading enzyme at pH 5-7. Particle arrays still characterized theinner fracture face (PF) of the plasma membrane of isolated protoplasts. Isolatedprotoplasts generally provided better quality plasma membrane fracture faces thanplasmolysed cells.
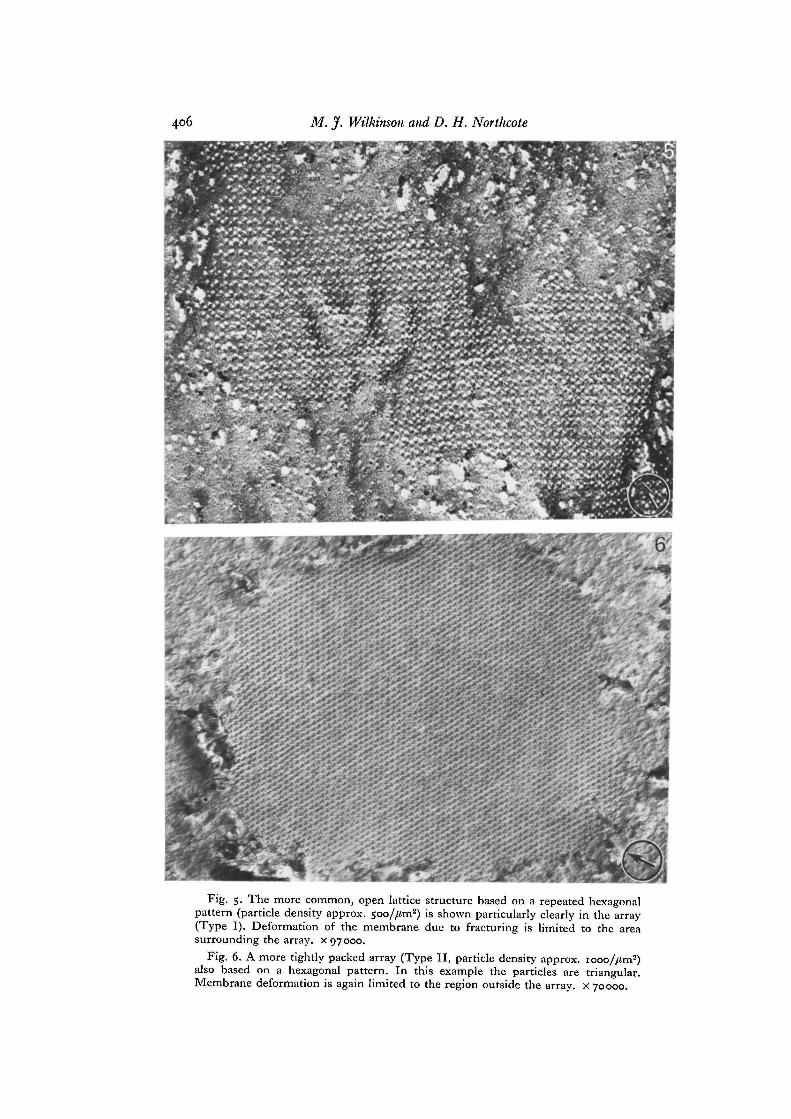
The more common array morphology, seen particularly clearly in isolated proto-plasts, was that of a hexagonal lattice (Type I) (Fig. 5). A more closely packed arrange-ment of particles was occasionally evident (Type II, Fig. 6) and this was also based on ahexagonal pattern. The density of particles in this type of array was approximately
Figs. 1-14. The circled arrow on each photograph incidates the direction of shadow.All specimens (except that shown in Fig. 4) were freeze-fixed, i.e. preserved in culturemedium or water without the addition of glycerol. Figs. 1-7, 9-12 show the plasmamembrane of potato callus cells (Figs. 1-4) and isolated protoplasts (Figs. 5-7, 9-12).Figs. 13 and 14 are of bean callus protoplasts.
Fig. 1. The inner fracture face (PF) of a potato callus suspension cell sampleddirectly from the growth medium (pH S-o). The intramembrane particles exhibit nomarked clustering, x 80000.
Fig. 2. The plasma membrane PF of a suspension cell that was plasmolysed by theaddition of sorbitol (0-35 M) to the growth medium at pH 5-7. A proportion of theintramembrane particles have clustered into discrete arrays. An area of the outersurface (ES) is visible at top right, x 93000.
Fig. 3. An intramembrane particle array present on the plasma membrane, PF, afterplasmolysis at pH 5-7 with 0-35 M glucose instead of sorbitol. x 77200.
Fig. 4. The plasma membrane, PF, of a specimen that was cryoprotected with 20 %glycerol following plasmolysis (035 M sorbitol, pH 57). Randomly spaced particlesin the area surrounding the array are clearly visible as this area has suffered nodeformation (compare with Figs. 5, 6). x 96000.
406 M. jf. Wilkinson and D. H. Northcote
Fig. 5. The more common, open lattice structure based on a repeated hexagonalpattern (particle density approx. 5oo//im2) is shown particularly clearly in the array(Type I). Deformation of the membrane due to fracturing is limited to the areasurrounding the array, x 97000.
Fig. 6. A more tightly packed array (Type II, particle density approx. iooo//im2)also based on a hexagonal pattern. In this example the particles are triangular.Membrane deformation is again limited to the region outside the array, x 70000.

Plasma membrane of plant protoplasts
Fig. 7. Optical diffraction patterns of intramembrane particle arrays. Types I andII (top left and right, x 194000 and 163000, respectively) are shown, together withtheir corresponding filtered image (middle row) obtained using all the reciprocallattice points in the diffraction patterns (bottom row). Averaged lattice constants are267 nm (Type 1) and 18-1 nm (Type II).

408 M. J. Wilkinson and D. H. Northcote

Plasma membrane of plant protoplasts 409
twice that of the Type I arrangement. The optical diffraction patterns of these 2 typesof array together with their reconstructed images are presented in Fig. 7.
Though the tonoplast contained numerous intramembrane particles no arrays wereever observed in this membrane either before or after plasmolysis (Fig. 8).
Plasma membrane ultrastructure during protoplast wall regeneration
When isolated protoplasts were returned to growth medium containing 0-35 Msorbitol they rapidly began to regenerate a new wall. Cell shape resumed within2 days and protoplasts then stained positively with Calcofluor White. Buddingoccurred during the first 2 weeks of culture and this was followed by the onset of celldivision. After 3-4 weeks small aggregates of cells developed. Particle arrays in theplasma membrane were most common in freshly isolated protoplasts and in a numberof preparations were detectable on almost every inner fracture face (PF) examined.They continued to be a regular feature of the plasma membrane for up to 48 h andaccounted for up to 19% of the surface area of plasma membrane (PF) visible inreplicas, accommodating up to approximately 45 % of all particles. As protoplast wallregeneration continued the fracture characteristics of the material remained constant.The pH of the protoplast culture medium did not vary significantly. After 3 to 4 days,however, particle arrays became far less common, with only about 1 in 25 fracturefaces (PF) showing evidence of such particle arrangements. After 7 days of protoplastculture arrays were rarely seen and they did not reappear during further periods ofwall regeneration and cell growth up to 3 weeks, when cell aggregates developed.
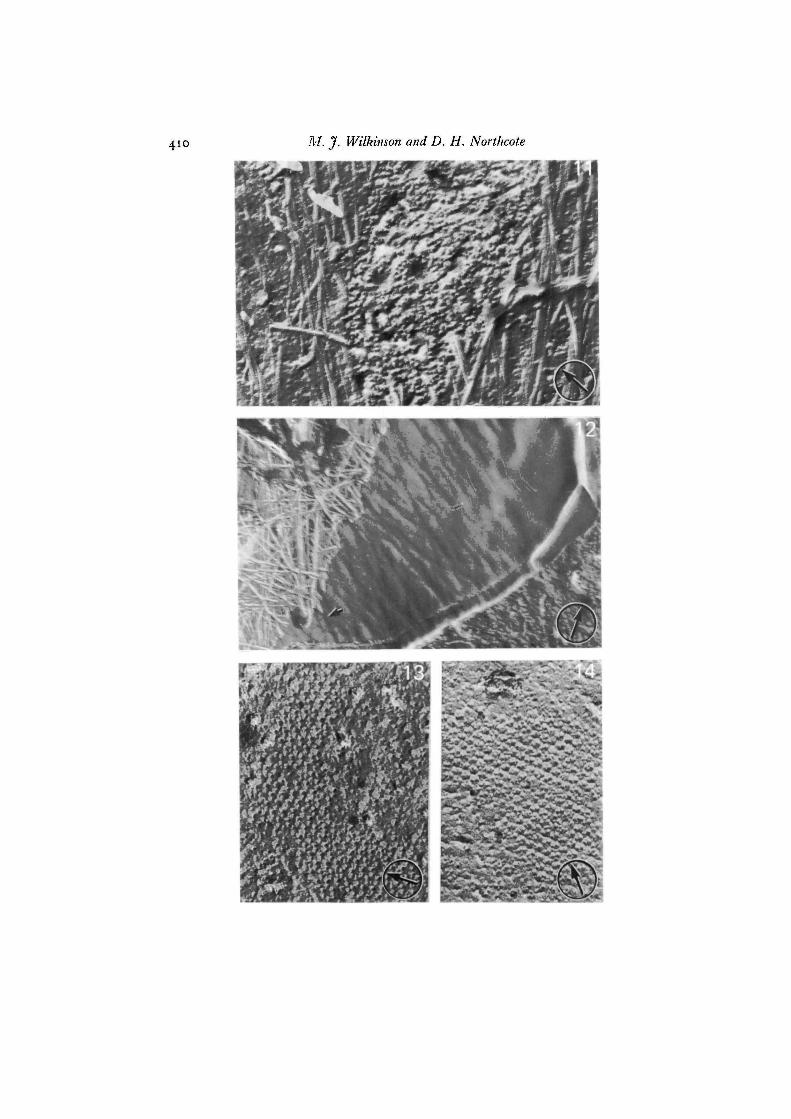
The progressive development of a network of microfibrils, probably of cellulose,over the plasma membrane surface during wall regeneration was visualized by etchingof glutaraldehyde-fixed samples that had been resuspended in water before freeze-fracturing. Freshly isolated protoplasts were naked and smooth (Fig. 9). Microfibrilsbegan to appear within 30 min of transferring protoplasts to culture medium. Theproduction of microfibrils on the plasma membrane surface (ES) did not appear todepend upon the presence of particle arrays, as judged by inspection of adjacentregions of membrane where the inner fracture face (PF) was revealed. Where particlearrays did occur in conjunction with newly formed microfibrils the orientation of
Fig. 8. Randomly distributed intramembrane particles on the inner fracture face(PF) of the tonoplast of a potato protoplast; such particles were never observed toaggregate, even after plasmolysis. A region of the outer surface is visible (top left),x 60000.
Fig. 9. The etched outer surface (ES) of a freshly isolated potato callus protoplastshowing the complete absence of wall material. An area of PF is visible (top left),x 58000.
Fig. 10. The etched surface of a potato callus protoplast cultured for 2-5 h. A fibril(single arrow) runs across the outer surface (ES). The orientation of particle rows inthe array present on the adjacent fracture face (PF) does not coincide with the directionin which the fibril has been deposited. A second fibril (double arrows) runs in asimilar direction to the first and has been pulled away from the surface and henceis thickened with carbon, x 66000.
27 CEL42
410 M. J. Wilkinson and D. H. Northcote

Plasma membrane of plant protoplasts 411
particle rows did not necessarily coincide with the direction in which microfibrils weredeposited (Figs. 10, 11). The surface of the plasma membrane lying beneath the net-work of microfibrils was only rarely displayed. It appeared relatively smooth exceptfor depressions left by the original microfibrils which had fractured away (Fig. 12).
We have also observed hexagonal particle arrays on the PF of the plasma membraneof protoplasts isolated from bean (JPhaseolus vulgaris L.) callus suspensions. Thesesuspensions were originally obtained from stem callus and protoplasts were isolatedby the method used for potato callus suspensions. Two types of array, strikinglysimilar to Types I and II present in potato protoplasts, were evident (Figs. 13, 14).
DISCUSSION
Using freeze-fracture replication we have demonstrated the presence of hexagonallyarranged intramembrane particles on the inner half (PF) of the plasma membrane ofplasmolysed potato callus cells. These arrays occurred in glutaraldehyde-fixedspecimens. It is therefore unlikely that they arose by lateral displacement of macro-molecules due to lipid solidification during freezing (Bullivant, 1977). The presenceof arrays in samples frozen live without pretreatment showed that chemical fixationand glycerination were not the cause of particle clustering. Particle arrays maytherefore be taken as indicative of the distribution of macromolecules, probablyintegral proteins, within the plasma membrane in vivo. The lack of marked particle-pitcomplementarity between matching halves of fractured plasma membrane (Wilkinson& Northcote, 1980) suggested that the particles were composed of proteins rather thanlipids (Verkleij & Ververgaert, 1978).
Array formation depended on plasmolysis rather than the plasmolysing agent(sorbitol, glucose or salts). Plasmolysis has 2 major consequences for the plant cellplasma membrane. It breaks any links between the outer surface of the membraneand the wall and simultaneously reduces the surface area of lipid bilayer available to
Fig. 11. A partially regenerated wall formed after 3 days of protoplast culture.Bundles of microfibrils, closely applied to the etched surface (ES) of the plasmamembrane, are being produced at a significant rate at this time, yet the frequency ofparticle arrays is decreasing, so that areas of PF below the microfibrils (centre) oftenlack evidence of particle arrays, x 75000.
Fig. 12. A rare view of the plasma membrane ES with all microfibrils fracturedaway. The membrane surface appears smooth except for depressions left by theoriginal structures. In some cases these depressions can be traced back to the re-maining microfibrils (arrow). The PF is exposed on the right of the picture, x 42200.
Fig. 13. An intramembrane particle array present on the inner fracture face (PF)of a freshly isolated bean protoplast obtained from a callus suspension. The patternis very similar to the Type I arrays seen in potato callus protoplasts (compare withF'g- 5)- x 112000.
Fig. 14. A second type of intramembrane particle array evident on the PF of theplasma membrane of a freshly isolated bean callus protoplast. This more closelyresembles the Type II arrays of potato callus protoplasts (compare with Fig. 6).x 125000.
27-2

412 M. J. Wilkinson and D. H. Northcote
accommodate proteins. Breaking membrane-wall links may tend to set membraneproteins adrift in the lipid bilayer and so facilitate mutual interaction by increasingtheir lateral mobility. A concomitant reduction in membrane surface area wouldperhaps reinforce this effect by increasing the concentration of proteins per unit areaof bilayer. We envisage the net result of these 2 processes to be the partial' crystalliza-tion' of proteins into the 2-dimensional arrays observed in plasmolysed material. Whyonly a proportion, up to approximately 45%, of intramembrane particles form arraysis unclear. There is at present no detailed information on the chemical identity of theintramembrane particles which might provide clues as to their capacity for aggregation.
Plasma membrane-wall connexions would be broken by mechanical stress duringplasmolysis but the fate of any transmembrane connexions on the inner face of themembrane, perhaps involving microtubules, is more difficult to determine. Micro-tubules are certainly present in close proximity to the inner plasma membrane face,probably appressed to it, in both turgid cells and plasmolysed protoplasts. Linearstructures lying parallel to each other and interpreted as microtubule impressions werecommonly visible on the plasma membrane fracture faces which exhibited arrays(Wilkinson & Northcote, 1980). These impressions did not, however, relate in anyobvious way to the ordering of particles within arrays or to the grouping of arrays.
Particle arrays were common in protoplasts plasmolysed at pH 5-7, the pH usedfor optimum yield and viability during protoplast isolation, but could not be detectedin identical cell batches plasmolysed at pH 5-0. It was not possible to obtain goodquality protoplasts at pH 5-0 as bursting occurred upon their release into the enzymesolution, possibly due to alterations in the permeability properties of the plasmamembrane or to the toxicity of wall-degrading enzymes at this lower pH.
Time-course experiments demonstrated the progressive loss of arrays duringprotoplast culture. After 7 days arrays were only rarely detected. During this periodthe effect of protoplast growth and expansion which occur despite the presence ofosmoticum, would be to increase the surface area of the plasma membrane. The resultof this increase may be to reduce the number of proteins per unit area of lipid bilayer.This may tend to reverse the original effect of plasmolysis and encourage the disso-lution of 'crystalline' proteins back into their lipid 'solvent' by diffusion.
All freeze-fractured biological membranes exhibit randomly spaced intramembraneparticles. The occurrence of such particles in geometric arrays is often taken toindicate their role in some specialized transport function, such as in the case of arrayspresent in the junction between axons of crayfish ganglia (Peracchia, 1974). Hexagonalarrays very similar to the Type I arrays described here were found in the plasmamembrane (PF) of cryoprotected transfer cells of clover root nodules (Briarty, 1973).It is particularly relevant that glutaraldehyde-fixed transfer cells that were sub-sequently glycerated did not exhibit particle arrays. Only those cells placed in 20 to30% glycerol without prior fixation, and thereby suffering plasmolysis, possessedevidence of arrays. Briarty argued that aldehyde fixation might disrupt any naturallyoccurring arrays which were possibly involved in the specialized transport functionsof transfer cells. We have found that glutaraldehyde fixation has no detrimental effecton the morphology of particle arrays present in potato callus. It seems more probable,

Plasma membrane of plant protoplasts 413
therefore, that glycerol-mediated plasmolysis of unfixed cells created the arrays andthat they do not occur under normal conditions in the plasma membrane of clovertransfer cells.
The presence of particle arrays in the plasma membrane of protoplasts isolated fromSkimmia japonica callus has been reported (Robonek & Peveling, 1977). It was sug-gested that the arrays represent specific proteins incorporated into the plasmamembrane from the highly active endoplasmic reticulum (which characterized proto-plasts in culture), so forming specialized channels for the passage of cellulose pre-cursors during wall regeneration. In the case of Skimmia protoplasts, intramembraneparticles began to form arrays after 24 h of protoplast isolation and particles inhexagonal arrangements became more clearly defined after some 48-72 h of culture.This contrasts with our observations that hexagonal particle arrays are apparent assoon as protoplasts are plasmolysed, well before there has been any opportunity forthe activation of endoplasmic reticulum. Furthermore, observations of thin sections(unpublished results) show that the endoplasmic reticulum also appears to proliferatein isolated potato callus protoplasts during periods of culture when arrays arebecoming less rather than more frequent. From the observations on potato and beanprotoplasts we suggest that there is no need to implicate the endoplasmic reticulumand the incorporation of specific proteins into the plasma membrane to explain theformation of particle arrays. In the absence of positive evidence of protein incor-poration the simplest explanation is that plasmolysis is the essential trigger that bringsabout the clustering of intramembrane particles.
Though particle arrays have been viewed here as a product of plasmolysis ratherthan as specialized regions of plasma membrane involved in wall precursor transport,we nevertheless considered the possibility that the individual macromolecules com-prising the arrays function in wall production and that their geometrical arrangementwould facilitate the investigation of such a role. Freeze-etching of protoplasts atvarious stages in the process of wall regeneration was performed to uncover anysimilarities in orientation between nascent microfibrils and particles aligned withinarrays. A number of time courses were performed. The synthesis and orientation ofmicrofibrils always seemed independent of particle arrays. Even when occurring inclose proximity to each other, the arrangement of particles within arrays was notnecessarily reflected in the orientation of newly formed microfibrils. At present,therefore, there seems to be no definite indication of a role for hexagonal particlearrays in any aspect of wall regeneration in higher plant cells. What is clear, however,is that plasmolysis may markedly influence the ultrastructural appearance of the plasmamembrane of plant cells. Whether plasmolysis has been intentional (e.g. when con-ferring osmotic stability on isolated protoplasts) or whether it has been due to the useof a cryoprotectant such as glycerol without prior chemical fixation, its potential forcausing alterations in plasma membrane organization should encourage caution wheninterpreting the results of ultrastructural studies on plasmolysed cells and isolatedprotoplasts.
During the completion of this manuscript, Davy & Mathias (1979) reported thathexagonal particle arrays in the plasma membrane of isolated higher plant protoplasts

414 M.J. Wilkinson and D. H. Northcote
were essentially an artefact of 20% glycerol treatment rather than the direct result of
plasmolysis. In contrast to our results, they did not detect any arrays in protoplasts
which had been only plasmolysed by the osmoticum during isolation. In the absence
of evidence from control experiments where the extensive plasmolysis of the glycerol
treatment (i.e. additional to that of the osmoticum) is paralleled by, for example,
exposing protoplasts to an increased concentration of osmoticum, we view any possible
effects of glycerol on array formation as being secondary to the macromolecular
rearrangements related directly to the retraction of the plasma membrane and its
significant reduction in surface area during plasmolysis.
Dr K. Roberts of the John Innes Institute kindly performed the optical-diffraction analysisshown in Fig. 7. We thank Mr D. Knights for assistance in operating the freeze-fracture unitand Mr L. Jewitt for help with photography. M. J.W. is grateful to the Potato Marketing Boardfor a studentship during the tenure of which this work was performed.
REFERENCES
ANSTIS, P. J. P. & NORTHCOTE, D. H. (1973). The initiation, growth and characteristics of atissue culture from potato tubers. J. exp. Bot. 24, 425-441.
BRANTON, D., BULLIVANT, S., GILULA, N. B., KARNOVSKY, M. J., MOORE, H., MOHLETHALER,
K., NORTHCOTE, D. H., PACKER, L., SATIR, B., SATIR, P., SPETH, V., STAEHLIN, L. A.,
STEERE, R. L. & WEINSTEIN, R. S. (1975). Freeze-etching nomenclature. Science, N.Y. 190,54-50-
BRIARTY, L. G. (1973). Repeating particles associated with membranes of transfer cells.Planta 113, 373-377-
BROWN, R. M. JR. & MONTEZINOS, D. (1976). Cellulose microflbrils: Visualisation of bio-synthetic and orienting complexes in association with the plasma membrane. Proc. natn.Acad. Sci. U.S.A. 73, 143-147.
BULLIVANT, S. (1977). Evaluation of membrane structure facts and artefacts produced duringfreeze-fracturing. J. Microscopy 111, 101-116.
DAVY, M. R. & MATHIAS, R. J. (1979). Close-packing of plasma membrane particles duringwall regeneration by isolated higher plant protoplasts - fact or artefact? Protoplasma 100,85-99-
GAMBORG, O. (1966). Aromatic metabolism in plants. II . Enzymes of the shikimate pathway insuspension cultures of plant cells. Can. J. Biochem. 44, 791-799.
GROUT, B. W. W. (1975). Cellulose microfibril deposition at the plasmalemma surface ofregenerating tobacco mesophyll protoplasts: a deep etch study. Planta 123, 275-282.
HANKE, D. E. & NORTHCOTE, D. N. (1974). Cell wall formation by soyabean callus protoplasts.J. Cell Sci. 14, 29-50.
HORNE, R. W. & MARKHAM, R. (1973). Applications of optical diffraction and image recon-struction techniques to electron micrographs. In Practical Methods in Electron Microscopy,vol. 11 (ed. A. M. Glauert), pp. 327-440. Amsterdam: North-Holland.
LARKIN, P. J. (1976). Purification and viability determinations of plant protoplasts. Planta 128,213-216.
MONTEZINOS, D. & BROWN, R. M. JR (1976). Surface architecture of the plant cell: biogenesisof the cell wall with special emphasis on the role of the plasma membrane in cellulose bio-synthesis. J. supramolec. Struct. 5, 277-290.
PERACCHIA, C. (1974). Excitable membrane ultrastructure. I. Freeze-fracture of crayfish axons.J. Cell Biol. 61, 107-122.
ROBENEK, H. & PEVELING, E. (1977). Ultrastructure of the cell wall of isolated protoplasts ofSkimmia japonica Thunb. Planta 136, 135-145.
UPADHYA, M. D. (1975). Isolation and culture of mesophyll protoplasts of potato (Solonumtuberosum L.). Potato Res. 18, 438-445.

Plasma membrane of plant protoplasts 415
VEKKLEIJ, A. J. & VERVERGAERT, P. H. J. TH. (1978). Freeze-fracture morphology of bio-logical membranes. Biochim. biophys. Ada 515, 303-327.
WILKINSON, M. J. (1978). Studies on Tissue Cultures and Isolated Protoplasts of Potato. Ph.D.thesis, University of Cambridge, U.K.
WILKINSON, M. J. & NORTHCOTE, D. H. (1980). A reliable method for obtaining matchedreplicas of freeze-fractured cell suspensions. J. Cell Sci. 42, 389-400.
WILLISON, J. H. M. & COCKING, E. C. (1972). The production of microfibrils at the surface ofisolated tomato-fruit protoplasts. Protoplasma 75, 397-403.
WILLISON, J. H. M. & COCKING, E. C. (1975). Microfibrils synthesis at the surface of tobaccomesophyll protoplasts, a freeze-etch study. Protoplasma 84, 147-159.
WILLISON, J. H. M. & GROUT, B. W. W. (1978). Further observations on cell-wall formationaround isolated protoplasts of tobacco and tomato. Planta 140, 53-58.
{Received 14 August 1979)






























