Embed Size (px)
Citation preview

POLLEN MORPHOLOGY AND ULTRASTRUCTURE
OF CALYCERACEAE
Melanie L. DeVore,1 Zaiming Zhao,2 Robert K. Jansen,2 and John J. Skvarla3
1Department of Biology and Environmental Sciences, Georgia College and State University, Milledgeville, GA 310612Section of Integrative Biology and Plant Resources Center, University of Texas at Austin, Austin, TX 78712
3Department of Botany and Microbiology, University of Oklahoma, Norman, OK 73019
Abstract: Pollen morphology of 13 species from all six genera of Calyceraceae(Acicarpha, Boopis, Calycera, Gamocarpha, Moschopsis, and Nastanthus) and represen-tatives of the Campanulaceae and Goodeniaceae is examined with light (LM), scanning(SEM), and transmission (TEM) electron microscopy. Acicarpha, Calycera, andNastanthus pollen grains are distinguished by angulaperturate apertures, colpar ledgesand surface depressions between colpi known as intercolpar concavities (IC). Pollen ofGamocarpha and Moschopsis is tricolporate rather than angulaperturate and without anIC. Some species of Boopis are similar to the preceding genera (e.g., B. graminea), whileothers (e.g., B. gracilis) are angulaperturate with ICs. Structural features derived fromfractured pollen in SEM and sections in TEM show pollen walls composed of prominentcolumellae ca. 0.55–1.1 mm high and ,0.25 mm wide. The columellae terminate distallyinto a complex of shortened columellae ca. 1.5 mm in length and are separated by an ill-defined irregular internal tectum layer. This structural complex is well known in severaltribes of the Asteraceae and referred to as the Anthemoid type. In those grains with anIC, the structure consists of essentially short (ca. 1 mm), unbranched columellae, similarto those found within the Asteraceae subfamily Barnadesioideae (Dasyphyllum andSchlechtendalia). Goodeniaceae (including Brunonia) pollen has angulaperturateapertures, spinules (i.e., minute spines), problematic IC and some structural similarityto Calyceraceae pollen. The tendency within Calyceraceae to develop colpar ledges,ektexine bridges, and ICs may be a synapomorphy uniting the family withGoodeniaceae. If the ancestral pollen type for the Calyceraceae, Asteraceae, andGoodeniaceae clade is the Gamocarpha type (convex intercolpar regions; no colparledges and no ektexine bridges), then the appearance of these structures within eachfamily may be a synapomorphy supporting their close phylogenetic relationshipsuggested by molecular analyses.
Keywords: Asteraceae, Barnadesioideae, Calyceraceae, Goodeniaceae, Asterales, inter-colpar concavities (IC), pollen.
Calyceraceae consist of six genera withapproximately 50 species. Some members ofthe family, especially Acicarpha, have capit-ula strikingly similar to Asteraceae, but theylack fully fused anthers and are characterizedby a single pendulous ovule per achene. Thefirst published evidence of a close relation-ship between Asteraceae and Calyceraceaewas provided by an extensive pollen survey(Skvarla et al., 1977). One of the outcomesof this study was the suggestion thatCalyceraceae were most closely related toAsteraceae based on the ultrastructuralsimilarity of Nastanthus (Calyceraceae) andDasyphyllum (Asteraceae, subfamily Barna-desioideae). Molecular phylogenetic analyses
(Jansen and Kim, 1996; Gustaffson et al.,1996; Lundberg and Bremer, 2003) supportthe monophyly of a clade that includesGoodeniaceae, Calyceraceae, and Asteraceae,and pollen data continue to provide insightsinto relationships among these families andother members of the Asterales (Polevova,2006). Hansen (1992) published a survey ofCalyceraceae pollen. One question Hansenraised was whether the intercolpar concav-ities (IC) present in some members ofBarnadesioideae and Calyceraceae representa synapomorphy.
There is strong evidence based onmolecular (Jansen and Kim, 1996; Gustaff-son et al., 1996; Lundberg and Bremer,
32 LUNDELLIA DECEMBER, 2007
LUNDELLIA 10:32–48. 2007

2003), morphological (DeVore, 1994; De-Vore and Stuessy, 1995; Pesecreta et al.,1994; Carlquist and DeVore, 1998) andphytochemical (Bohm and Stuessy, 1995;Bohm et al., 1995) data that Calyceraceae aresister to Asteraceae. Pollen data have pro-vided some of the strongest support fora close relationship between these families(Skvarla et al., 1977) and additional palyno-logical surveys of Calyceraceae are valuablebecause they could elucidate morphologicalevolution within these families. In particu-lar, these data are needed to reconstruct theancestral features of Asteraceae.
In the present study, pollen morphology(Erdtman, 1960) of 13 species from all sixgenera of Calyceraceae and two representa-tives of Campanulaceae and Goodeniaceae isexamined with light (LM), scanning (SEM)and transmission (TEM) microscopy. Inaddition to providing a palynologic over-view of the Calyceraceae, which bothcomplements and extends the original pollenmorphologic data of Hansen (1992), we alsocompare pollen morphology of Calyceraceaewith other families of Asterales.
MATERIALS AND METHODS
Pollen was obtained from herbariumspecimens of 13 species representing all sixgenera of Calyceraceae. Also included forcomparative purposes was one species fromtwo other families of Asterales, Goodenia-ceae (Goodenia ovata Sm.), and Campanu-laceae (Campanula rotundiflora L.). Table 1shows collection information for all taxaexamined as well as SEM and TEM figurecitations.
All pollen samples were initially acet-olyzed (Erdtman, 1960) to enhance viewingof the exine surfaces in (LM) and (SEM).For LM, pollen grains were stained andmounted on glass slides according to Nair(1970), and then examined with transmittedlight using a Leitz Wetzlar microscope. Theslides are housed in the reference collectionat The University of Texas at Austin (TEX).
For SEM, pollen was screened with finewire mesh to remove undigested coarseplant fragments (Skvarla, 1966) and placedon sucrose pads to remove finer particles(Chissoe and Skvarla, 1974). Whole, intact
TABLE 1. Calyceraceae, Goodeniaceae and Campanulaceae pollen examined and collection data.
Taxon Locality Collector Herbarium Figures
Acicarpha tribuloides Juss Argentina Burkart & Troncoso28056
TEX 1a–d
Boopis gracilis Phil. Argentina DeVore 1747 TEX 1e–h, 6aB. graminea Phil. Chile DeVore 1249 TEX 1i–LCalycera calcitrapa Griseb. Argentina Cabrera 18119 TEX 2a–dC. eryngioides Remy Chile DeVore 1303 TEX 2e–gC. herbacea Cav. Chile DeVore 1216 TEX 2h–k, 6bC. involucrata Phil. Argentina DeVore 1737 TEX 2L,3a–cC. leucanthema Kuntze Chile DeVore 1143 TEX 3d–fC. spinulosa Gill. ex Miers Argentina DeVore 1730 TEX 3g–jGamocarpha alpina (Poepp. ex
Less.) H.V. HansenChile DeVore 1250 TEX 3k, L, 4a,
b, 6cMoschopsis sp. Argentina Moore 1966 US 4c–e, 6dNastanthus andina (Miers. H.V.
HansenArgentina DeVore 1723 TEX 4f–h
N. spathulatus Miers Chile DeVore 1427 TEX 4i–LCampanula rotundiflora L.
(Campanulaceae)United States Powell 321110 TEX 5a, b
Goodenia ovata Smith(Goodeniaceae)
Australia Vasek 680929-10 TEX 5c–f
NUMBER 10 DEVORE ET AL.: CALYCERACEAE POLLEN 33

pollen grains were dehydrated using 5 minwashes in graded ethanol (EtOH) solutions,three 5 min washes in absolute EtOH, andtwo 5 min treatments in 100% hexamethyl-disilazane (Chissoe et al., 1994). Afterdehydration, all samples received a sputter-coating with gold for 4–5 min. Finally,pollen grains were examined with a JEOLJSM 880 scanning electron microscope.
Freeze-sectioned pollen grains were pre-pared following the method described byVezey et al. (1994). Sections were thensputter-coated and examined as describedabove.
TEM sample preparation followed themethod described by Skvarla (1966, 1973).Examination and photography were per-formed with a JEOL 2000-FX TEM.
SEM and TEM negatives were scannedand final plates prepared using AdobePhotoShop 7.
RESULTS
Pollen Descriptions. All examined pol-len samples are described below. All quan-titative numbers in pollen description arethe average of five random measurements.
CalyceraceaeAcicarpha—A genus of annuals to
perennials distributed from the altiplano ofBolivia and Peru to southern Brazil andnorthern Argentina. Acicarpha tribuloidesJuss. has also been collected in New Zealandand eastern North America where it wasintroduced in ballast dumps and neversurvived and reproduced past the year itwas introduced (DeVore, 1991). Acicarpha isthe genus within Calyceraceae that mostresembles Asteraceae in capitular morphol-ogy. Like Asteraceae, members of the genuspossess capitula that develop centripetally.Other distinctive features of the genusinclude lateral fusion of the filaments abovenectaries and a prominent flared connectivebase (Pesecreta et al., 1994).
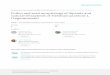
Acicarpha tribuloides Juss.—Figs. 1a–d.Grains spheroidal-rhombohedral, 13.0 mm
3 12.5 mm.
Tricolporate and angulaperturate; inter-colpar regions strongly concaved; intercol-par ledges present; bridge over pore pro-nounced; colpi 10.4 mm 3 0.9 mm, furrownearly same width with pointed ends;endocolpium 2.5 mm 3 6.0 mm, lalongate,lateral ends acute.
Exine 1.1 mm at equator; foot layer-endexine 0.4 mm; columellae distally branched,0.19 mm wide, 0.55–0.90 mm high; tectumthickness 0.15 mm; surface spinulate, 0.15 mmhigh, 0.4 mm wide.
Boopis. A genus of annuals and peren-nials ranging from central Chile and north-ern Argentina south to Tierra del Fuego.Capitula consist of cymose subunits andachenes characterized by calycine lobes ofequal length, and indicated here and else-where (i.e., Patel, 1976; Hellwig, 2007) thegenus has dimorphic pollen.
Boopis gracilis Phil.—Figs. 1e–h, 6a.Grains spheroidal, 18.9 mm 3 18.8 mm.Tricolporate and angulaperturate; inter-
colpar regions flat; intercolpar ledges pres-ent; bridge over pore; colpi 14.2 mm 3
1.0 mm, furrow nearly same width withpointed ends; endocolpium 1.8 mm 3
5.2 mm, lalongate, narrowly lenticulate.Exine 1.2 mm at equator; foot layer-
endexine 0.48 mm; columellae distallybranched, 0.2 mm wide, 0.52–0.90 mm high;tectum thickness 0.2 mm; surface spinulate,0.2 mm high, 0.4 mm wide.
Boopis graminea Phil.—Figures 1i–L.Grains subspheroidal, 22.3 mm 3 18.3
mm.Tricolporate; intercolpar regions and
ledges absent; bridge over pore absent; colpi16.3 mm 3 3.0 mm, furrow nearly samewidth with rounded ends; endocolpium2.0 mm 3 8.0 mm, lalongate, narrowlylenticulate.
Exine 1.7 mm at equator; foot layer-endexine 0.44 mm; columellae distallybranched, 0.3 mm wide, 1.1 mm high; tectumthickness 0.16 mm; surface spinulate,0.23 mm high, 0.4 mm wide.
Calycera. A genus of annuals andperennials with a principal distribution
34 LUNDELLIA DECEMBER, 2007
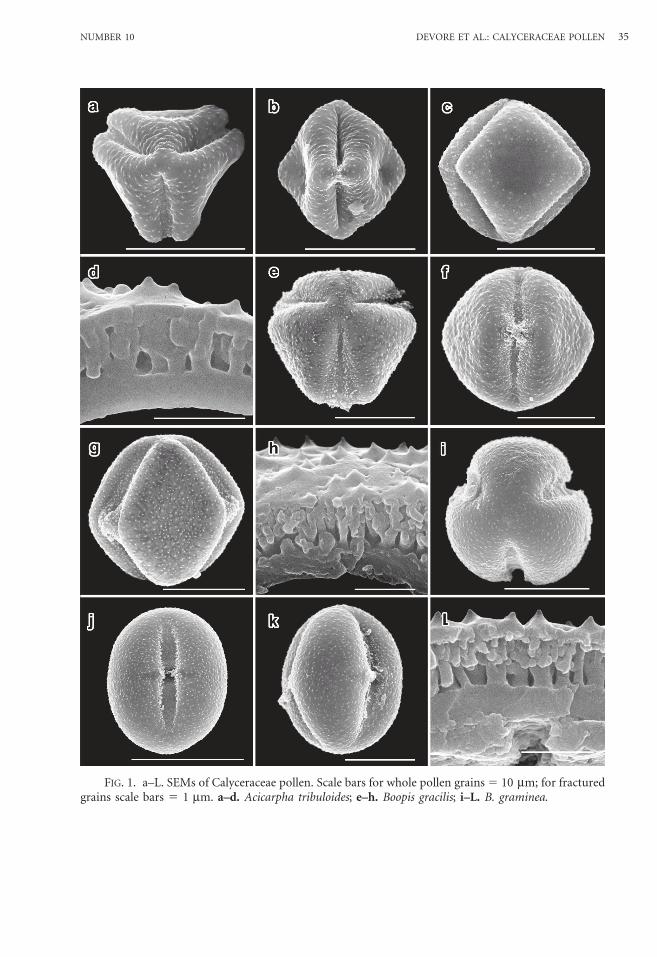
FIG. 1. a–L. SEMs of Calyceraceae pollen. Scale bars for whole pollen grains 5 10 mm; for fracturedgrains scale bars 5 1 mm. a–d. Acicarpha tribuloides; e–h. Boopis gracilis; i–L. B. graminea.
NUMBER 10 DEVORE ET AL.: CALYCERACEAE POLLEN 35

along the Andes of Argentina and Chile.Calycera consists of two distinct groups ofspecies: Argentinean taxa (with the excep-tion of C. herbacea Cav.) with chromosomenumbers of n 5 21 and anthers lacking tails;Chilean species (with the exception of C.pulvinata Remy) consisting of taxa withvariable chromosome numbers and antherswith distinct tails (DeVore, 1994). Capitulaconsist of cymose subunits. The genus hasbeen delimited from other genera withinCalyceraceae based on the presence ofdimorphic achenes (achenes with differentcalycine lobe lengths).
Calycera calcitrapa Griseb.—Figs. 2a–d.Grains spheroidal-rhombohedral, 16.0 mm
3 16.5 mm.Tricolporate and angulaperturate; inter-
colpar regions flat-slightly concave; inter-colpar ledges present; bridge over pore; colpi16.3 mm 3 3.0 mm, furrow nearly samewidth with pointed ends; endocolpium2.0 mm 3 5.5 mm, lalongate, lateral endsacute.
Exine 1.5 mm at equator; foot layer-endexine 0.4 mm; columellae distally branched,0.18 mm wide, 0.9 mm high; tectum thickness0.2 mm; surface spinulate, 0.15 mm high,0.3 mm wide.
Calycera eryngioides Remy—Figs. 2e–g.Grains spheroidal-rhombohedral, 21.6 mm
3 28.0 mm.Tricolporate and angulaperturate; inter-
colpar regions strongly concave; intercolparledges present and pronounced; bridge overpore; colpi 18.0 mm 3 2.6 mm, furrownearly same width with pointed ends;endocolpium 2.8 mm 3 6.0 mm, lalongate,lateral ends rounded.
Exine 2.1 mm at equator; foot layer-endexine 0.82 mm; columellae distallybranched, 0.2 mm wide, 1.1 mm high; tectumthickness 0.2 mm; surface spinulate, 0.05 mmhigh, 0.15 mm wide.
Calycera herbacea Cav.—Figs. 2h–k, 6b.Grains spheroidal-rhombohedral, 17.6 mm
3 20.8 mm.Tricolporate and angulaperturate; inter-
colpar regions flat-slightly concave; inter-
colpar ledges present; bridge over pore; colpi15.6 mm 3 3.6 mm, furrow nearly samewidth with pointed ends; endocolpium2.2 mm 3 7.2 mm, lalongate, lateral endsacute.
Exine 1.5 mm at equator; foot layer-endex-ine 0.44 mm; columellae distally branched,0.24 mm wide, 0.9 mm high; tectum thickness0.16 mm; surface spinulate, 0.2 mm high,0.35 mm wide.
Calycera involucrata Phil.—Figs. 2L,3a–c.
Grains spheroidal-rhombohedral, 17.6 mm3 19.8 mm.
Tricolporate and angulaperturate; inter-colpar regions flat-slightly concave; inter-colpar ledges present; bridge over pore; colpi15.2 mm 3 3.8 mm, furrow nearly samewidth with rounded ends; endocolpium2.0 mm 3 6.0 mm, lalongate, lateral endsacute.
Exine 1.4 mm at equator; foot layer-endexine 0.5 mm; columellae distally branched,0.22 mm wide, 0.75 mm high; tectum thickness0.15 mm; surface spinulate, 0.15 mm high,0.35 mm wide.
Calycera leucanthema Kuntze—Figs.3d–f.
Grains spheroidal-rhombohedral, 20.0 mm3 23.0 mm.
Tricolporate and angulaperturate; inter-colpar regions strongly concave; intercolparledges present and pronounced; bridge overpore pronounced; colpi 17.7 mm 3 2.4 mm,furrow nearly same width with pointed ends;endocolpium 3.1 mm 3 7.5 mm, lalongate,lateral ends rounded.
Exine 0.9 mm at equator; foot layer-endexine 0.25 mm; columellae distallybranched, 0.17 mm wide, 0.5 mm high;tectum thickness 0.15 mm; surface spinulate,0.1 mm high, 0.3 mm wide.
Calycera spinulosa Gilles ex Miers.—Figs. 3g–j.
Grains spheroidal-rhombohedral, 20.0 mm3 22.7 mm.
Tricolporate and angulaperturate; inter-colpar regions flat-slightly concave; inter-colpar ledges present; bridge over pore; colpi
36 LUNDELLIA DECEMBER, 2007
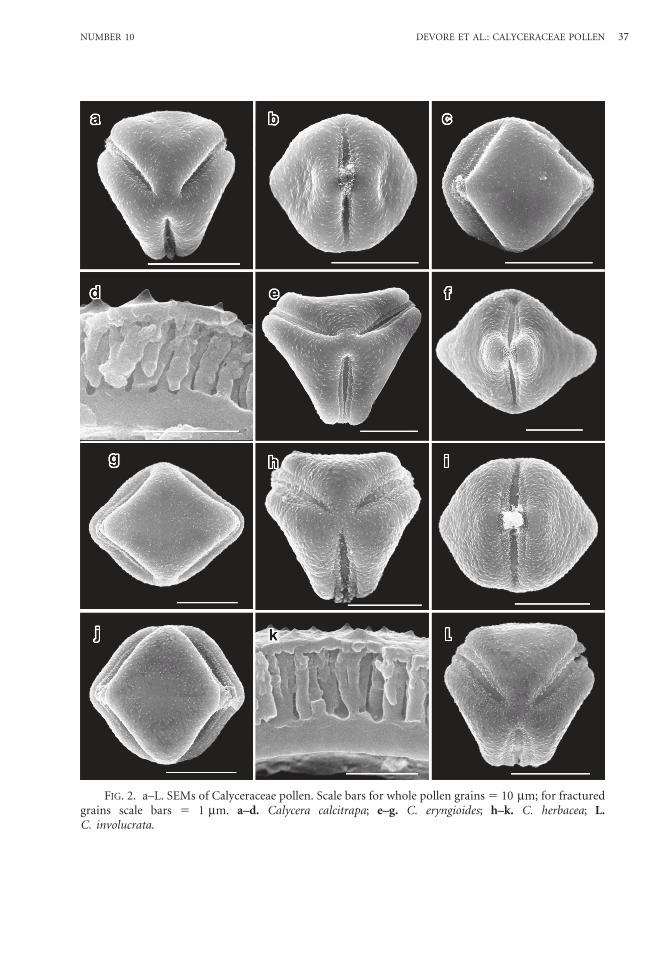
FIG. 2. a–L. SEMs of Calyceraceae pollen. Scale bars for whole pollen grains 5 10 mm; for fracturedgrains scale bars 5 1 mm. a–d. Calycera calcitrapa; e–g. C. eryngioides; h–k. C. herbacea; L.C. involucrata.
NUMBER 10 DEVORE ET AL.: CALYCERACEAE POLLEN 37
FIG. 3. a–L. SEMs of Calyceraceae pollen. Scale bars for whole pollen grains 5 10 mm; for fracturedgrains scale bars 5 1 mm. a–c. Calycera involucrata; d–f. C. leucanthema; g–j. C. spinulosa; k, L.Gamocarpha alpina.
38 LUNDELLIA DECEMBER, 2007

17.3 mm 3 3.6 mm, furrow nearly samewidth with rounded ends; endocolpium1.8 mm 3 7.2 mm, lalongate, lateral endsacute.
Exine 1.52 mm at equator; foot layer-endexine 0.4 mm; columellae distally branched,0.2 mm wide, 0.9 mm high; tectum thickness0.22 mm; surface spinulate, 0.18 um high,0.35 mm wide.
Gamocarpha. A genus of rhizomatousperennials of Andean regions of southernArgentina and Chile. Capitula are composedof heads consisting of cymes surrounded byfused pales.
Gamocarpha alpina (Poepp. ex Less.) H.V. Hansen—Figs. 3k, L; 4a, b; 6c.
Grains spheroidal, 20.0 mm 3 18.5 mm.Tricolporate; intercolpar regions and
ledges absent; bridge over pore absent; colpi14.6 mm 3 4.2 mm, furrow ends pointed;endocolpium 4.0 mm 3 8.0 mm, lalongate,lateral ends acute.
Exine 1.9 mm at equator; foot layer-endexine 0.5 mm; columellae distally branched,0.3 mm wide, 1.3 mm high; tectum thickness0.1 mm; surface spinulate, 0.1 mm high,0.2 mm wide.
Moschopsis. A genus of perennial herbsextending from the Andes of Argentina andChile to Tierra del Fuego. The only characterthat delimits the genus is the absence ofa clearly defined involucre.
Moschopsis sp.—Figs. 4c–e, 6d.Grains spheroidal, 17.5 mm 3 17.3 mm.Tricolporate; intercolpar regions and
ledges absent; bridge over pore absent; colpi14.2 mm 3 2.2 mm, furrow nearly samewidth with rounded ends; endocolpium2.9 mm 3 7.2 mm, lalongate, broadly lenti-culate.
Exine 1.6 mm at equator; foot layer-endexine 0.4 mm; columellae distally branched,0.26 mm wide, 1.0 mm high; tectum thickness0.2 mm; surface spinulate, 0.1 mm high, 0.2 mmwide.
Nastanthus. This unusual genus, oncedescribed as appearing like a cauliflower(Miers, 1860), extends from the northern tosouthern Andean ranges of Chile and
Argentina. The generic delimitation betweenNastanthus and Boopis has been unclear. Theonly noted difference between the twogenera has been the size of the receptacle.
Nastanthus andina (Miers) H. V. Han-sen—Figs. 4f–h.
Grains spheroidal-rhombohedral, 23.5 mm3 25.5 mm.
Tricolporate and angulaperturate; inter-colpar regions flat to slightly concave;intercolpar ledges present; bridge over pore;colpi 20.2 mm 3 4.1 mm, furrow nearlysame width with pointed ends; endocolpium3.1 mm 3 7.0 mm, lalongate, lateral endsacute.
Exine 1.85 mm at equator; foot layer-endexine 0.41 mm; columellae distally branched,0.25 mm wide, 1.25 mm high; tectum thickness0.19 mm; surface spinulate, 0.18 mm high,0.32 mm wide.
Nastanthus spathulatus Miers.—Figs. 4i–L.Grains spheroidal-rhombohedral, 20.0 mm
3 19.0 mm.Tricolporate and angulaperturate; inter-
colpar regions flat slightly concave; inter-colpar ledges present; bridge over pore; colpi15.5 mm 3 5.2 mm, furrow nearly samewidth with pointed ends; endocolpium3.0 mm 3 7.5 mm, lalongate, lateral endsacute.
Exine 1.2 mm at equator; foot layer-endexine 0.5 mm; columellae distally branched,0.22 mm wide, 0.6 mm high; tectum thickness0.1 mm; surface spinulate, 0.1 mm high,0.2 mm wide.
CampanulaceaeCampanula rotundiflora L.—Figs. 5a–b.Grains spheroidal, 32.2 mm 3 30.6 mm.Porate with at least 4 rounded pores;
exine 0.67 mm; highly reduced columellae;surface entirely spinate, spines 2.2 mm highand 0.9 mm wide.
GoodeniaceaeGoodenia ovata Sm.—Figs. 5c–f.Grains spheroidal, 23.4 mm 3 24.7 mm.Tricolporate and angulaperturate; inter-
colpar regions convex; intercolpar ledgesabsent; bridge over pore absent; colpi16.8 mm 3 2.6 mm, furrow ends rounded;
NUMBER 10 DEVORE ET AL.: CALYCERACEAE POLLEN 39
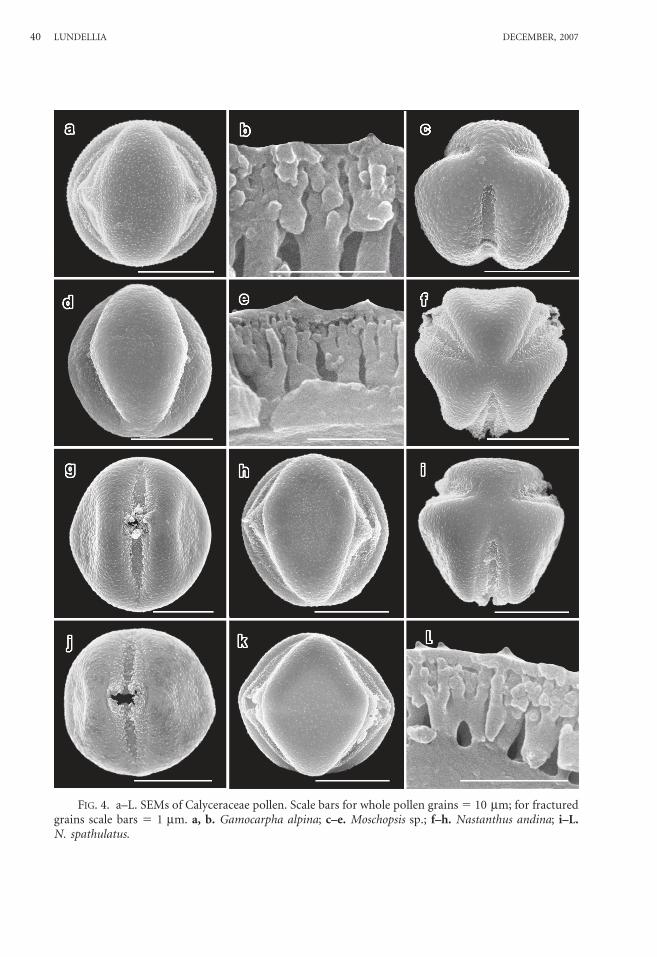
FIG. 4. a–L. SEMs of Calyceraceae pollen. Scale bars for whole pollen grains 5 10 mm; for fracturedgrains scale bars 5 1 mm. a, b. Gamocarpha alpina; c–e. Moschopsis sp.; f–h. Nastanthus andina; i–L.N. spathulatus.
40 LUNDELLIA DECEMBER, 2007
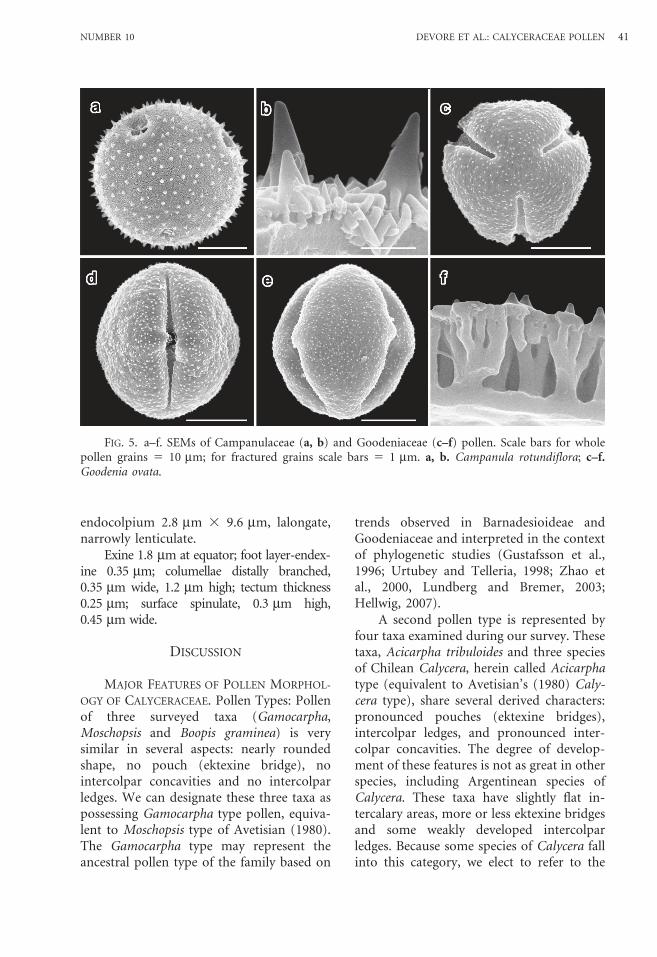
endocolpium 2.8 mm 3 9.6 mm, lalongate,narrowly lenticulate.
Exine 1.8 mm at equator; foot layer-endex-ine 0.35 mm; columellae distally branched,0.35 mm wide, 1.2 mm high; tectum thickness0.25 mm; surface spinulate, 0.3 mm high,0.45 mm wide.
DISCUSSION
MAJOR FEATURES OF POLLEN MORPHOL-
OGY OF CALYCERACEAE. Pollen Types: Pollenof three surveyed taxa (Gamocarpha,Moschopsis and Boopis graminea) is verysimilar in several aspects: nearly roundedshape, no pouch (ektexine bridge), nointercolpar concavities and no intercolparledges. We can designate these three taxa aspossessing Gamocarpha type pollen, equiva-lent to Moschopsis type of Avetisian (1980).The Gamocarpha type may represent theancestral pollen type of the family based on
trends observed in Barnadesioideae andGoodeniaceae and interpreted in the contextof phylogenetic studies (Gustafsson et al.,1996; Urtubey and Telleria, 1998; Zhao etal., 2000, Lundberg and Bremer, 2003;Hellwig, 2007).
A second pollen type is represented byfour taxa examined during our survey. Thesetaxa, Acicarpha tribuloides and three speciesof Chilean Calycera, herein called Acicarphatype (equivalent to Avetisian’s (1980) Caly-cera type), share several derived characters:pronounced pouches (ektexine bridges),intercolpar ledges, and pronounced inter-colpar concavities. The degree of develop-ment of these features is not as great in otherspecies, including Argentinean species ofCalycera. These taxa have slightly flat in-tercalary areas, more or less ektexine bridgesand some weakly developed intercolparledges. Because some species of Calycera fallinto this category, we elect to refer to the
FIG. 5. a–f. SEMs of Campanulaceae (a, b) and Goodeniaceae (c–f) pollen. Scale bars for wholepollen grains 5 10 mm; for fractured grains scale bars 5 1 mm. a, b. Campanula rotundiflora; c–f.Goodenia ovata.
NUMBER 10 DEVORE ET AL.: CALYCERACEAE POLLEN 41
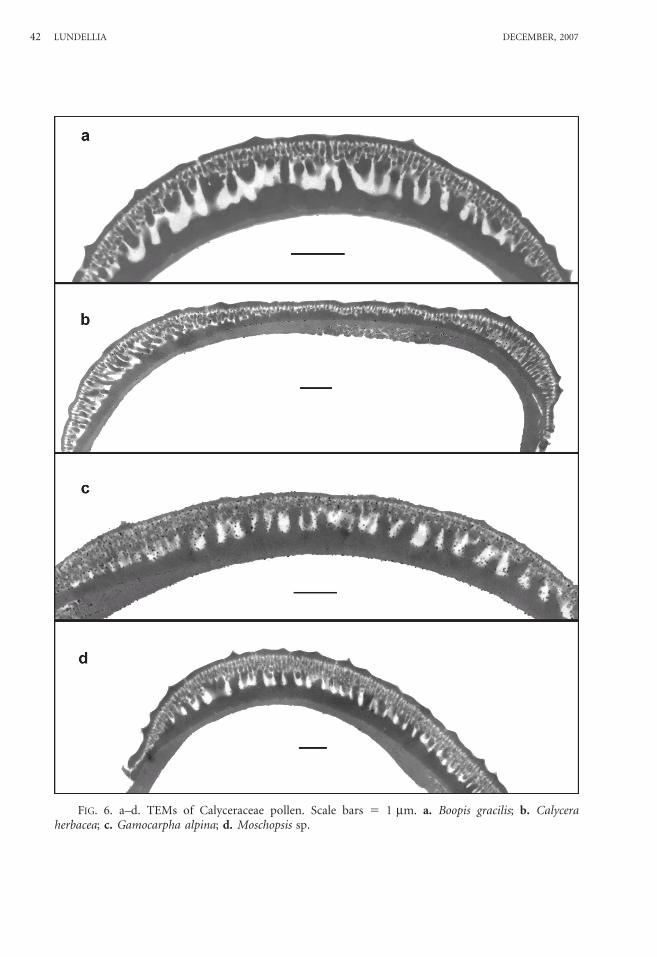
FIG. 6. a–d. TEMs of Calyceraceae pollen. Scale bars 5 1 mm. a. Boopis gracilis; b. Calyceraherbacea; c. Gamocarpha alpina; d. Moschopsis sp.
42 LUNDELLIA DECEMBER, 2007

pollen type with strongly developed, derivedcharacters, as the Acicarpha type.
Intercolpar concavities: Skvarla et al.(1977) first noted that Asteraceae andCalyceraceae possess intercolpar concavities,and this is one of the major reasons that theysuggested a close relationship between thetwo families. Intercolpar concavities werefirst described by Wodehouse (1928) in sixspecies of Chuquiraga. His original descrip-tion of these species was that they ‘‘… areunique in the possession of three distinctconcavities appearing as the impressions onecould make with the thumb in a ball of softdough. Since these impressions are betweenfurrows, I shall call them intercolpar con-cavities, and their position on the equatorsuggests the further designation of equato-rial concavities. They have not been foundelsewhere in the grains of either theMutisieae or any other tribes of the Com-positae except Schlectendalia.’’ These con-cavities vary from simple concavities ofWodehouse to prominent depressions ofUrtubey and Telleria (1998). Contemporarystudies have shown that intercolpar concav-ities characterize Dasyphyllum, Schlectendalia,and some species of Arnaldoa (Skvarla et al.,1977; Urtubey and Telleria, 1998; Zhao et al.,2000; Telleria and Katinas, 2005). Theterminology of Wodehouse (1928) has neverfound its way into any pollen glossaries ordictionaries, including the glossary in Wode-house’s 1935 book, which explained mostpalynological terms at that time. This may bethe reason why few pollen studies ofCalyceraceae did not specifically recognizethese features (Erdtman, 1952; Auer et al.,1955), although they appeared in some LMs(Markgraf and D’Antoni, 1978; Wingenrothand Heusser, 1984). Perhaps unaware ofWodehouse’s early description, Heusser(1971) described them for Calycera eryn-gioides as being ‘‘pinched in the middle toappear in the form of a bow.’’ Heusser (1971)also presented clear photos of intercolparconcavities in Boopis leucanthema Poepp. inLess. (5 Calycera leucanthema (Poepp. inLess.) Kuntze) and B. pusilla Phil. In contrast,
his descriptions of Moschopsis monocephalaReiche, Gamocarpha poeppigii DC. and Nas-tanthus scapigera Miers did not indicate theirpresence. The most definitive work on thesestructures is credited to Hansen (1992).Examining unacetolyzed pollen with SEMhe recorded intercolpar concavities in fourspecies of Acicarpha, three of seven species ofBoopis, five of eight species of Calycera, andtwo of three species of Nastanthus. Gamocar-pha was the only taxon uniformly lackingthem. Most recently, intercolpar concavitieswere recognized as mesocolpal depressions inSEMs of Boopis anthemoides Juss. andMoschopsis rosulata (N. E. Br.) Dusen, severalmembers of Goodeniaceae (Goodenia scapi-gera R. Br., G. ovata Sm., Pentaptilon,Scaevola, Selliera, Velleia, Verreauxia) (Gus-tafsson et al., 1997), and in Dasyphyllum(Urtubey and Telleria, 1998; Zhao et al.,2000).
Ektexine bridges: An ektexine bridge isan extension of the exine that projects overthe pore. This feature has been observed inseveral groups of Asteridae including Ly-thraceae (Graham et al., 1985) and Boragi-naceae (Nowicke and Skvarla 1974). Thisfeature, which is a later stage of a commontrend found within Asteridae, is likelyderived within Calyceraceae, and there isa trend from slight (Gamocarpha) to welldeveloped ektexine bridges within the Chi-lean species of Calycera and Acicarpha.
Colpar ledges: Colpar ledges are ridgesof exine that line the inner margin of thecolpi. This feature is found in bothCalyceraceae and Goodeniaceae. The func-tion of colpar ledges may be related toharmomegathy. Wodehouse (1935) showedthat colpi function in accommodatingvolume changes as pollen passes out ofthe humid anther to the dry air. Bolick(1978) hypothesized that the evenly spacedcolpi enable round pollen grains to ap-proach a triangular perimeter followingwater loss. This triangular cross sectionwould be the most stable cross section andwould prevent further collapse of pollen.Prominent colpar ledges might prevent
NUMBER 10 DEVORE ET AL.: CALYCERACEAE POLLEN 43

additional water loss by sealing off the colpibefore shrinkage reduces the diameter ofthe pollen grain to the extent that thecolpar ledges are in contact.
GENERAL POLLEN CHARACTERISTICS OF
GOODENIACEAE, CAMPANULACEAE AND SIM-
ILARITIES WITH CALYCERACEAE: Goodenia-ceae consist of about 14 genera and 300species and exhibit a wide range of pollenvariation (Patel, 1976; Skvarla et al., 1977;Gustafsson et al., 1997; Polevova, 2006).Duigan (1961) surveyed 39 species from sixgenera (Brunonia, Dampiera, Goodenia,Scaevola, Selliera, and Velleia) within thefamily using light microscopy with Erdtman(1952) and Avetisian (1973) providingadditional but more limited LM observa-tions. Skvarla et al. (1977) selected ninespecies from six genera (Anthotium, Bruno-nia, Dampiera, Goodenia, Scaevola, andVelleia) for observation using SEM andTEM. Our summary of the overall pollenmorphology for use in comparison withCalyceraceae is based on these two studiesand the new data generated in this paper.
Most pollen of Goodeniaceae is tricol-porate, however grains with two, four, ormore colpi, as well as syncolpate grains, arefound within the family (Duigan, 1961).When grains have more than three colpi,Duigan (1961) noted that these pollengrains were present within species withpolyploid races that also produce largergrains. One genus, Leschenaultia, has tetradpollen grains (Skvarla et al., 1977). Basedon the overall pollen morphology, Duigan(1961) recognized three pollen types: 1)Brunonia; 2) Dampiera; and 3) Goodenia.Brunonia-type pollen is distinct in havingcolpar ledges (Duigan, 1961). These struc-tures are layers of exine that extend alongthe edges of the colpi. Endocolpia aresublanceolate lalongate. In polar view,Brunonia-type pollen appears triangular inshape, while in equatorial view, the grainsare prolate-spheroidal in shape. The surfaceis microspinulose. Dampiera-type grainsare characterized by non-elongated colpiwith irregular outlines. In equatorial view,
the grains appear circular or elliptical,while in polar view the grains are oblateto subprolate in shape with straight,slightly convex, circular, or lobed inter-colpar regions. The surface is rugulate. Incontrast to the Dampiera-type grains,Goodenia-type pollen has lalongate endo-colpia with acute or rounded lateral ends.Goodenia-type grains, like those of theDampiera-type, are triangular when viewedat the polar position and exhibit straight-sided colpi. Equatorial views show thegrains to be circular-elliptical in shape.Unlike the other pollen types, some specieswith Goodenia-type pollen have colpimargins modified to form bridges overthe endoaperture (e.g., Scaevola ramosis-sima (Sm.) K. Krause).
Observations from SEM and TEM(Patel, 1976; Skvarla et al., 1977; Polevova,2006) were used to determine pollensurface and ultrastructural characteristicsof Goodeniaceae pollen grains. Most gen-era within the family have spinulatesurfaces, with two genera (Dampiera andAnthotium) being characterized by thepresence of striae and rugulae. TEMmicrographs document a relatively thinendoexine and a thick footwall with large,distally bifurcating columellae (see Good-enia, Fig. 5f). This bifurcating pattern canbe simple (e.g., Dampiera) or of a higherorder (e.g., Brunonia). In Scaevola, thehigher order columellae are fused. Morerecent LM/SEM/TEM studies in the Good-eniaceae (Gustafsson et al., 1997) sup-ported the three groups based on pollenmorphology originally established by Dui-gan (1961) but with an expanded list oftaxa. Their Scaevola-Goodenia group(5Brunonia and Goodenia groups of Dui-gan, 1961), with tricolporate, microspinu-lose grains having distally branched colu-mellae was indicated by Gustaffson et al.(1997) as showing similarity to Asteraceaeand Calyceraceae. Finally, Carolin’s review(2007) of Goodeniaceae further supportsthe pollen data of Duigan (1961) andGustafsson (1997).
44 LUNDELLIA DECEMBER, 2007

Based on the available pollen data(Duigan, 1961; Skvarla et al., 1977; Gustafs-son et al., 1997), as well as phylogeneticanalyses based on morphology of Good-eniaceae (Carolin, 1978) and a molecularphylogeny of Asterales that included 12species of Goodeniaceae (Gustafsson et al.,1996), one can begin to elucidate sometrends in pollen morphology of Goodenia-ceae. The Goodeniaceae contains two largegenera with more than eighty species (Good-enia and Scaevola) that have not been fullysurveyed for pollen variation. Therefore, theextent of pollen variation within thesegenera is unknown. In Gustafsson et al.(1996), Goodenia ovata groups with Sellieraradicans, and G. scapigera forms a clade withVerreuxia reinwardtii.
Trends exist within Goodeniaceae thatare shared with Calyceraceae. The mostevident of these is the presence of colparledges in Brunonia. Within Calyceraceae,colpar ledges are found in Acicarpha,Calycera, and Nastanthus. Ektexine bridgesalso have evolved within both Calyceraceaeand Goodeniaceae, as well as within otherfamilies (e.g., Primulaceae, Nowicke andSkvarla, 1977; Fabaceae, Endo and Ohashi,1996). Within Calyceraceae, a trend is seenfrom slight (Gamocarpha) to well developedektexine bridges within the Chilean speciesof Calycera and Acicarpha. We hypothesizethat such an extension over the pore mayfunction in diminishing water loss. Accord-ing to Ficks law, evaporation rate dependson different resistances to diffusion (Salis-bury and Ross, 1985). Because water mole-cules diffusing from a pore can travel in anydirection, the boundary layer is very thinand the concentration gradient is very steep.The presence of an ektexine bridge may beinstrumental in deterring water loss from thepore.
Nowicke and Skvarla (1979) proposedthat congruity in wall architecture is themanifestation of long-standing structuralsimilarity dictated by common ancestry.Based on this premise, we suggest particularpollen features (e.g., colpar ledges in both
Calyceraceae and Goodeniaceae), mightfollow a pathway dictated by commonancestry. In other words, taxa sharinga common ancestry would obtain featuresvia similar, if not nearly identical, series ofstructural modifications of one particularshared pollen type. Utilizing a trend inpollen morphology as a phylogenetic char-acter proved insightful in previous studies(DeVore et al., 2000) and supports thehypothesis of Nowicke and Skvarla (1979).The question still remains regarding whetherintercolpar concavities in Calyceraceae andAsteraceae are a parallelism or a synapomor-phy for these families.
A recent survey of pollen morphologyand ultrastructure of families closely alliedwith the Asteraceae, Calyceraceae, andGoodeniaceae clade (Polevova, 2006) indi-cates that the sister group to this clade,Menyanthaceae and Campanulaceae, sharepollen features that are unlike those presentwithin Asteraceae, Calyceraceae, and Good-eniaceae. An examination of Figures 5a andb clearly show that the pollen of Campanu-laceae and Menyanthaceae (not figured inthe present paper) are distinctive from thatof Calyceraceae by having large, unbranchedcolmellae in the upper exine covered bya thick tectum. Clearly the Campanulaceae-Menyanthaceae pollen type is not compara-ble to any type present within Calyceraceaeor within its closest related families Aster-aceae and Goodeniaceae.
Clarifying the trends in pollen morphol-ogy for Calyceraceae is valuable because thefamily contains two distinct pollen types, theless derived Gamocarpha–type and thehighly derived Acicarpha type, as well asintermediate forms between these two types.In essence, the entire pathway of modifica-tions of the pollen ground plan for theGoodeniaceae, Calyceraceae, and Asteraceaeclade appears to be present within Calycer-aceae (Table 2). Trends in pollen morphol-ogy within Calyceraceae also are useful inhelping to provide a search image forpaleopalynologists seeking to document thefirst appearance of Asteraceae in the fossil
NUMBER 10 DEVORE ET AL.: CALYCERACEAE POLLEN 45

record using light microscopy. One wouldpredict that the ancestral lineage potentiallypossessed grains similar to what we describeas the Gamocarpha type (nearly roundedshape, no ektexine bridge, no intercolparconcavities and no intercolpar ledges.
ACKNOWLEDGEMENTS
This work was supported by a NationalScience Foundation grant (DEB – 9318279)and the Sidney F. and Doris Blake Centen-nial Professorship in Systematic Botany toRKJ. We are grateful to the staff of the PlantResources Center for use of their facility andto the following herbaria for permission toremove pollen from specimens: TEX, SHST,and US. We also thank William F. Chissoe ofthe Samuel Roberts Noble Electron Micros-copy Laboratory, University of Oklahoma,for technical assistance with the SEM andTEM work and two anonymous reviewersfor valuable comments on an earlier draft of
the manuscript. This paper represents a por-tion of the Ph. D. dissertation of ZZpresented to the Graduate School of theUniversity of Texas.
LITERATURE CITED
Auer, V., Salmi, M., and K. Salminen. 1955. Pollenand spore types of Fuego–Patagonia. Ann. Acad.Sci. Fenn., Ser. A 43: 1–14.
Avetisian, E. M. 1973. Palynology of the orderCampanulales s.l. In: Pollen and Spores of recentplants (in Russian). Proc. 3rd Int. Conf. Palyn.Conf. Acad. Sci. USSR, 90–93, 2pls.
———. 1980. Pollen morphology of the familyCalyceraceae (in Russian). In: Sistematika I Evo-lyutsiya Vysshikh Rastenii. Leningrad: Nauka,pp. 57–64.
Bohm, B. A. and T. F. Stuessy. 1995. Flavonoidchemistry of Barnadesioideae (Asteraceae). Syst.Bot. 20: 22–27.
———, A. Reid, M. L. DeVore, and T. F. Stuessy.1995. Flavonoid chemistry of Calyceraceae. Canad.J. Bot. 73: 162–165.
Bolick, M. R. 1978. Taxonomic, evolutionary, andfunctional considerations of Compositae pollen
TABLE 2. Summary of pollen features and trends of Goodeniaceae, Calyceraceae andBarnadesioideae. Data from Duigan (1961), Patel (1976), Skvarla et al. (1977), Avetisian (1980),Hansen (1992), Gustafsson et al. (1997), Urtubey and Telleria (1998), Zhao et al. (2000), andcurrent study.
Goodeniaceae Calyceraceae Barnadesioideae
Exine Thickened columellaebifurcating distally,do not extendthrough entire exine,thick footwallreticulate, spongynetwork absent
Thickened columellaebifurcating distally,do not extendthrough entire exine,thick footwallreticulate, spongynetwork absent
Thickened columellaebifurcating distally,some extend throughentire exine, thinfootwall; reticulate,spongy networkpresent
Colpar ledges absent topresent
Present only inBrunonia
Present in Acicarpha,Calycera, Nastanthus,and some membersof Boopis
Absent
Ektexine bridges absentto slight to welldeveloped
Present within family Present in Acicarpha,some members ofBoopis, Calycera, andNastanthus
Absent
Intercolpar concavitiesconvex to flattened(mesocolpardepressions) topresent
Present in severalmembers ofGoodeniaceae
Present in Acicarphaand some membersof Boopis, Calycera,and Nastanthus
Present in Dasyphylumand Schlectendalia,paraporal depressionsin Arnaldoa andSchlectendalia
46 LUNDELLIA DECEMBER, 2007

ultrastructure and sculpture. Pl. Syst. Evol. 130:
209–218.
Carlquist, S. and M. L. DeVore. 1998. Wood anatomy
of Calyceraceae with reference to ecology, habit,
and systematic relationships. Aliso 17: 63–76.
Carolin, R. 1978. The systematic relationships of
Brunonia. Brunonia 1: 9–29.
———. 2007. Goodeniaceae. Pp. 589–598 in The
Families and Genera of Vascular Plants, Eudicots,
Asterales. Vol. 8, J. W. Kadereit and C. Jeffrey, eds.
Heidelberg: Springer.
Chissoe, W. F. and J. J. Skvarla. 1974. Sucrose density
pads for concentration and purification of pollen
grains. Stain Technol. 49: 123–124.
———, E. L. Vezey, and J. J. Skvarla. 1994.
Hexamethyldisilazane as a drying agent for pollen
scanning electron microscopy. Biotechnic. Histo-
chem. 69: 192–198.
DeVore, M. L. 1991. The occurrence of Acicarpha
tribuloides (Calyceraceae) in eastern North Amer-
ica. Rhodora 93: 26–35.
———. 1994. Systematic studies of Calyceraceae.
Ph.D. dissertation. Columbus, OH: Ohio State
University.
——— and T. F. Stuessy. 1995. The place and time of
origin of the Asteraceae, with additional comments
on the Calyceraceae and Goodeniaceae. Pp. 23–40
in Advances in Compositae Systematics. Kew: Royal
Botanic Garden.
———, Z. Zhao, R. K. Jansen, and J. Skvarla. 2000.
Utility of trends in pollen morphology for phylo-
genetic analyses: An example using subfamilies
Barnadesioideae and Cichorioideae (Asteraceae).
Pp. 1–14 Proceedings of Pollen and Spores 1998:
Morphology and Biology. Kew: Royal Botanic
Garden.
Duigan, S. L. 1961. Studies of the pollen grains of
plants native to Victoria, Australia. 1. Goodeniaceae
(including Brunonia ). Proc. Roy. Soc. Victoria 74:
87–109.
Endo, Y. and H. Ohashi. 1996. The pollen morphology
of Vicia (Leguminosae) and its systematic implica-
tion. J. Pl. Res. 108: 17–24.
Erdtman, G. 1952. Pollen morphology and plant
taxonomy. Angiosperms. Stockholm: Almqvist &
Wiksell.
———. 1960. The acetolysis method. A revised
description. Svensk Bot. Tidskr. 39: 561–564.
Graham, A., J. Nowicke, J. J. Skvarla, S. A. Graham,V. Patel, and S. Lee. 1985. Palynology and
systematics of the Lythraceae. I. Introduction and
genera Adenaria through Ginoria. Amer. J. Bot. 72:
1012–1031.
Gustafsson, Mats H. G., A. Backlund, and B. Bremer.1996. Phylogeny of the Asterales sensu lato based
on rbcL sequences with particular reference to
the Goodeniaceae. Pl. Syst. Evol. 199: 217–
242.
———, E. Grafstrom, and S. Nilsson. 1997. Pollenmorphology of the Goodeniaceae and comparisonswith related families. Grana 36: 185–207.
Hansen, H. V. 1992. Studies in the Calyceraceae witha discussion of its relationship to Compositae.Nordic J. Bot. 12: 63–75.
Hellwig, E. H. 2007. Calyceraceae. Pp. 19–25 in TheFamilies and Genera of Vascular Plants. Vol. 8, ed.,Flowering Plants, K. Kubitzki, J. W. Kadereit, & C.Jeffrey, eds. Springer.
Heusser, C. J. 1971. Pollen and Spores of Chile. ModernTypes of the Pteridophyta, Gymnospermae, andAngiospermae. Tucson: The University of ArizonaPress.
Jansen, R. K. and K.-J. Kim. 1996. Implications ofchloroplast DNA data for the classification andphylogeny of the Asteraceae. Pp. 317–319 inCompositae: Systematics. Vol. I, D. J. Hind and H.Beentje, eds, Proc. International Compositae Con-ference, Kew, 1994. Kew: Royal Botanic Garden.
Lundberg, J. and K. Bremer. 2003. A phylogeneticstudy of the order Asterales using one largemorphological and three molecular data sets. Int.J. Pl. Sci. 164: 553–578.
Markgraf, V. and H. L. D’Antoni. 1978. Pollen flora ofArgentina. Tucson: The University of Arizona Press.
Miers, J. 1860. Nastanthus. Ann. Mag. Nat. Hist. 6: 184.
Nair, P. K. K. 1970. Pp. 1–160 in Pollen morphology ofangiosperms. Lucknow: Scholar Publishing House.
Nowicke, J. W. and J. J. Skvarla. 1974. A palynologicalinvestigation of the genus Tournefortia (Boragina-ceae). Amer. J. Bot. 61: 1020–1036.
——— and ———. 1977. Pollen morphology and therelationship of the Plumbaginaceae, Polygonaceae,and Primulaceae to the order Centrospermae.Smithsonian Contr. Bot. 37: 1–64.
——— and ———. 1979. Pollen morphology: thepotential influence on higher order systematics.Annals Missouri Botanical Garden 66: 633–700.
Patel, V. C. 1976. Pollen morphology of Valerianaceae,Goodeniaceae, Brunoniaceae, Stylidiaceae and Ca-lyceraceae. M. S. thesis. University of Oklahoma.Norman, Oklahoma., 1–126.
Pesecreta, T. C., V. I. Sullivan, and M. L. DeVore.1994. The connective base and filament of Acicar-pha tribuloides (Calyceraceae). Amer. J. Bot. 81:753–759.
Polevova, S. V. 2006. Review of the sporodermultrastructure of members of the Asterales. Paleon-tol. J. 40: S656–663.
Salisbury, F. B. and C. W. Ross. 1985. PlantPhysiology. Belmont, CA: Wadsworth PublishingCompany.
Skvarla, J. J. 1966. Techniques of pollen and electronmicroscopy. Part I. Staining, dehydration andembedding. Oklahoma Geol. Notes 26: 179–186.
———. 1973. Pollen. Pp. 456–459 in Encyclopedia ofMicroscopy and Micro-techniques, P. Gray, ed. NewYork: Van Norstrand Reinhold, Inc.
NUMBER 10 DEVORE ET AL.: CALYCERACEAE POLLEN 47

———, B. L. Turner, V. C. Patel, and A. S. Tomb.1977. Pollen morphology in the Compositae and inmorphologically related families. Pp. 141–248 inThe Biology and Chemistry of the Compositae, V. H.Heywood et al, ed. London: Academic Press.
Telleria, M. C. and L. Katinas. 2005. The unusualoccurrence of tricolpate pollen within Mutisieae(Asteraceae). Grana 44: 91–97.
Urtubey, E. and M. C. Telleria. 1998. Pollenmorphology of the subfamily Barnadesioideae(Asteraceae) and its phyletic and taxonomicsignificance. Rev. Palaeobot. Palynol. 104: 19–37.
Vezey, E. L., L. E. Watson, J. J. Skvarla, and J. R.Estes. 1994. Pleisiomorphic and apomorphic pollen
structure characteristics of Anthemideae (Asteroi-deae: Asteraceae). Amer. J. Bot. 81: 648–657.
Wigenroth, M. and C. J. Heusser. 1984. Pollen of theHigh Andean Flora. Quebrada Benjamin Matienzo,Province of Mendoza Argentina. Mendoza: InstitutoArgentino de Nivologia y Glaciologia.
Wodehouse, R. P. 1928. Pollen grains in the identifica-tion and classification of plants. II. Barnadesia. 1991.Bull. Torrey Bot. Club 55: 449–462.
———. 1935. Pollen Grains. New York: McGraw-Hill.Zhao, Z., J. J. Skvarla, R. Jansen, and M. DeVore.
2000. Phylogenetic implications of pollen morphol-ogy and ultrastructure in the Barnadesioideae(Asteraceae). Lundellia 3: 26–40.
48 LUNDELLIA DECEMBER, 2007