Embed Size (px)
Citation preview

MINIREVIEW
Poly(ADP-ribose)
The most elaborate metabolite of NAD+
Alexander Burkle
Department of Biology, University of Konstanz, Germany
Introduction
The life cycle of poly(ADP-ribose)
NAD+ ⁄NADH is among the most versatile biomole-
cules, as it can be used not only as a coenzyme for a
large number of oxidoreduction reactions, but in its
oxidized version can also serve as substrate for several
different of ADP-ribosyl transfer reactions, which are
the overarching theme of this minireview series. The
covalent transfer onto glutamic acid, aspartic acid or
lysine residues of target proteins (‘acceptors’), followed
by successive transfer reactions onto the protein–
mono(ADP-ribosyl) adduct, and subsequently onto the
emerging chain of several covalently linked ADP-ribo-
syl residues is the basis of the formation of poly(ADP-
ribose), which can be regarded the cell’s most elaborate
metabolite of NAD+ [1]. ADP-ribose chains may com-
prise up to 200 ADP-ribose units, coupled via unique
ribose (1¢¢fi2¢) ribose phosphate-phosphate linkages
and display several branching points resulting from the
formation of ribose (1¢¢¢fi2¢¢) ribose linkages (Fig. 1).
Poly(ADP-ribosyl)ation occurs in multicellular organ-
isms including plants and some lower unicellular eu-
karyotes, but is absent in prokaryotes and yeast.
Poly(ADP-ribosyl)ation is catalysed by the family of
poly(ADP-ribose) polymerases (PARPs; Fig. 2), enco-
ded in human cells by a set of 18 different genes [2].
Keywords
PARP; tankyrase; poly(ADP-ribose); DNA
damage; DNA repair; genomic instability;
centrosome; centromere; telomeres; mitotic
spindle
Correspondence
A. Burkle, Department of Biology,
Box X911, University of Konstanz,
D-78457 Konstanz, Germany
Tel: +49 7531 884035
Fax: +49 7531 884033
E-mail: [email protected]
Website: http://gutenberg.biologie.
uni-konstanz.de/
(Received 5 May 2005, accepted 14 July
2005)
doi:10.1111/j.1742-4658.2005.04864.x
One of the most drastic post-translational modification of proteins in eu-
karyotic cells is poly(ADP-ribosyl)ation, catalysed by a family enzymes
termed poly(ADP-ribose) polymerases (PARPs). In the human genome, 18
different genes have been identified that all encode PARP family members.
Poly(ADP-ribose) metabolism plays a role in a wide range of biological
structures and processes, including DNA repair and maintenance of
genomic stability, transcriptional regulation, centromere function and mito-
tic spindle formation, centrosomal function, structure and function of vault
particles, telomere dynamics, trafficking of endosomal vesicles, apoptosis
and necrosis. In this article, the most recent advances in this rapidly grow-
ing field are summarized.
Abbreviations
ANK, ankyrin; BER, base excision repair; BRCA1, breast cancer 1 protein; DBD, DNA-binding domain; HPS, His-Pro-Ser-rich; IRAP, insulin-
responsive amino peptidase; MVP, major vault protein; NuMa, nuclear mitotic apparatus protein; PARG, poly(ADP-ribose) glycohydrolase;
PARP, poly(ADP-ribose) polymerase; RNP, ribonucleoprotein particle; Sir2, silent information regulator 2; TCDD, 2,3,7,8-tetrachlorodibenzo-
p-dioxin; TRF, telomeric-repeat binding factor.
4576 FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS

Apart from the covalent modification of acceptor
proteins, poly(ADP-ribose) has also been shown to
interact noncovalently, yet specifically, with a wide
range of proteins [3]. This interaction is mediated
through a sequence motif displaying a conserved pat-
tern, and it is interesting to note that DNA damage
checkpoint proteins such as p53 and p21 possess such
poly(ADP–ribose) interaction domains. DNA methyl-
transferase-1 (DNMT1) is also able to bind long and
branched ADP-ribose polymers in a noncovalent fash-
ion, which leads to inhibition of DNMT1 activity [4].
Taken together, poly(ADP-ribose) can also affect the
function of proteins that are not direct modification
targets.
Enzymes involved in poly(ADP-ribose)metabolism
PARP-1
The prototypic enzyme of the PARP family is poly
(ADP-ribose) polymerase-1 (PARP-1; EC 2.4.2.30;
Table 1). Its discovery was made four decades ago by
Chambon, Weill and Mandel [5] and marked the birth
of a most interesting and intriguing field originally
occupied by biochemists, but later on also attracting
radiobiologists, toxicologists, geneticists, molecular
biologists, pharmacologists, cell biologists and experts
from other biological disciplines.
Nicotinamide
NAD+
PAR
PAR
n NAD+
n Nicotinamide + n H+
Rib
O
O
PO
O
PO
Rib O-
O-
N
N
NH2
N
NAcceptor protein CH2 C
O
O-
Rib
N
N
NH2
N
N
O
O
O-
PO
O
O-
PO
Rib
Rib
N
N
NH2
N
N
O
O
O-
PO
O
O-
PO
Rib
RibRib
N
N
NH2
N
N
O
O
O-
PO
O
O-
PO
Rib
Rib
N
N
NH2
N
N
O
O
O-
PO
O
O-
PORib Rib
N
N
NH2
N
N
O
O
O-
PO
O
O-
PO
Rib Rib
N+
H2NCO
N
H2NCO
+ H+
Rib
N
N
NH2
N
N
O
O
O-
PO
O
O-
PO
Rib
+
CH2 C
O
O
CH2 C
O
OAcceptor protein
Acceptor protein
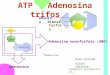
Fig. 1. Poly(ADP-ribose) structure.
Poly(ADP-ribose) polymerases (PARPs)
cleave the glycosidic bond of NAD+
between nicotinamide and ribose followed
by the covalent modification of mainly glu-
tamate residues of acceptor proteins with
an ADP-ribosyl unit. PARPs also catalyse
an adduct elongation, giving rise to linear
polymers with chain lengths of up to about
200 ADP-ribosyl units, characterized by
their unique ribose (1¢¢fi2¢) ribose phos-
phate-phosphate backbone. At least some
of the PARP family members also catalyse
a branching reaction by creating ribose
(1¢¢¢fi2¢¢) ribose linkages. The sites of
hydrolysis catalysed by poly(ADP-ribose)
glycohydrolase (PARG), the major poly(ADP-
ribose)-degrading enzyme, are indicated
by arrows.
A. Burkle Poly(ADP-ribosyl)ation
FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS 4577

PARP-1 is a highly conserved and abundant nuclear
protein, which is catalytically active as a dimer and is
the major acceptor protein in intact cells, via the
so-called automodification reaction. A range of other
nuclear proteins can also serve as acceptor proteins
(see below). PARP-1 displays a characteristic three-
domain structure (Fig. 2), which can be further broken
down into modules A–F [6]. The N-terminal 42 kDa
DNA-binding domain (DBD) also comprises the pro-
tein’s nuclear localization signal and is adjacent to a
central 16 kDa automodification domain. The 55 kDa
catalytic domain, which includes the active site, is
located at the C-terminus.
The DBD of PARP-1 binds to single- or double-
strand breaks with high affinity via two zinc fingers
but has also been reported to be involved in protein–
protein interactions. The first zinc finger is essential for
PARP-1 activation by DNA strand breaks, whereas
the second is essential for PARP-1 activation by DNA
single-strand breaks but not double-strand breaks. In
the absence of DNA breaks, PARP-1 displays a very
low basal enzyme activity. The molecular mechanism
linking DNA binding to catalytic activation is
unknown. Automodification of PARP-1 was reported
to be mostly induced by single-strand breaks, whereas
histone H1 seems to be modified preferentially when
PARP-1 binds to double-strand breaks [7]. Following
up earlier reports [8–10], several groups have provided
recent evidence for alternative activation mechanisms
for PARP-1, independent of DNA strand breakage.
PARP-1 was reported to recognise distortions in the
Table 1. Human enzymes involved in poly(ADP-ribose) formation or degradation and their genes.
Official gene
symbol Name [aliases]
Chromosomal
location
Gene
ID Protein size
PARP1 Poly(ADP-ribose) polymerase family, member 1
[PARP-1, ADPRT, ADPRT1, PARP, PPOL, pADPRT-1]
1q41-q42 142 1014 aa (113 kDa)
PARP2 Poly(ADP-ribose) polymerase family, member 2
[PARP-2, NAD+ poly(ADP-ribose) polymerase-2]
14q11.2-q12 10038 583 aa (66 kDa)
PARP3 Poly(ADP-ribose) polymerase family, member 3
[PARP-3, NAD+ poly(ADP-ribose) polymerase-3]
3p22.2-p21.1 10039 532 aa (60 kDa);
splice variant
539 aa (60.8 kDa)
PARP4 Poly(ADP-ribose) polymerase family, member 4
[PARP4, ADPRTL1, PARPL, PH5P, VAULT3, VPARP, p193]
13q11 143 1724 aa (192.8 kDa)
TNKS Tankyrase, TRF1-interacting ankyrin-related ADP-ribose
polymerase [PARP-5a, PARP5A, PARPL, TIN1, TINF1, TNKS1]
8p23.1 8658 1327 aa (142 kDa)
TNKS2 Tankyrase-2, TRF1-interacting ankyrin-related
ADP-ribose polymerase 2 [PARP-5b, PARP-5c,
PARP5B, PARP5C, TANK2, TNKL]
10q23.3 80351 1166 aa (126.9 kDa)
TIPARP TCDD-inducible poly(ADP-ribose) polymerase
[PARP-7]
3q25.31 25976 657 aa (75 kDa)
PARP10 Poly (ADP-ribose) polymerase family, member 10
[PARP-10]
8q24.3 84875 1025 aa (150 kDa)
PARG Poly(ADP-ribose) glycohydrolase
[PARG]
10q11.23 8505 Three splice variants
976 aa (111 kDa);
893 aa (102 kDa);
866 aa (99 kDa)
PARP-1
PARP-2
PARP-3
PARP-4(VPARP)
Tankyrase (PARP-5a)
Tankyrase-2 (PARP-5b)
N BRCTNLSZFI ZFII
Module A B C E D F
DNA-binding domain (DBD) NAD+-binding domain
Automodification domain
Active site
DBD
NLS
BRCT
SA24 ankyrinHPS
SA24 ankyrin
VIT TM VWA SH3 MVP-BD
C
Fig. 2. Structural organization of some PARP protein family mem-
bers. BRCT, BRCA1 C-terminus; DBD, DNA-binding domain; HPS,
His-Pro-Ser-rich domain; MVP-BD major vault binding domain; NLS,
nuclear localization signal; SAM, sterile a-module; SH3, src homo-
logy region; TM, transmembrane domain; VIT, vault protein inter-
alpha-trypsin domain; VWA, von Willebrand factor type A domain;
ZF; zinc finger.
Poly(ADP-ribosyl)ation A. Burkle
4578 FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS

DNA helical backbone and to bind to three- and four-
way junctions as well as to stably unpaired regions in
double-stranded DNA [11]. Such PARP-1 interactions
with non-B DNA structures led to catalytic activation
of the enzyme in the absence of free DNA ends. DNA
hairpins, cruciforms and stably unpaired regions are
all effective coactivators of PARP-1 automodification
and poly(ADP-ribosyl)ation of histone H1. These data
suggest a link between PARP-1 binding to non-B
DNA structures in the genome and its function in the
dynamics of local modulation of chromatin structure
in normal cellular physiology (see below). Binding to
and activation by unbroken DNA in the context of
chromatin has also been reported to play a crucial role
in the NAD+-dependent modulation of chromatin
structure and transcriptional regulation, mediated by
PARP-1 [12]. Along the same lines it was suggested
that fast and transient decondensation of chromatin
structure by poly(ADP-ribosyl)ation occurring in Aply-
sia neurones in the absence of DNA strand breaks
enables the transcriptional regulation needed to form
long-term memory in this organism [13].
The automodification domain of PARP-1 is rich in
glutamic acid residues, consistent with the fact that
poly(ADP-ribosy)lation occurs on such residues. This
domain also comprises a breast cancer 1 protein
(BRCA1) C-terminus (BRCT) motif that is present in
many DNA damage repair and cell-cycle checkpoint
proteins.
The C-terminal 55 kDa catalytic domain contains the
residues essential for NAD+-binding, ADP-ribosyl
transfer and branching reactions. The crystal structure
of the C-terminal catalytic fragment revealed a striking
homology with bacterial toxins that act as mono(ADP-
ribosyl) transferases [14].
While PARP-1 is constitutively expressed, its charac-
teristic ability of being activated by DNA strand
breaks makes poly(ADP-ribosyl)ation an immediate
and drastic cellular response to DNA damage as
induced by ionizing radiation, alkylating agents and
oxidants. In the absence of DNA single and double
strand breaks, poly(ADP-ribosyl)ation seems to be a
very rare event in live cells, but it can increase over
100-fold upon DNA damage [15]. Under these condi-
tions about 90% of poly(ADP-ribose) is synthesized
by PARP-1 [16].
Among many identified interaction partners of
PARP-1 are also other members of the PARP-family,
such as PARP-2 [17–19] and PARP-3 [20]. As men-
tioned above, the most prominent target protein
(acceptor) of this poly(ADP-ribosyl)ation reaction is
PARP-1 itself but many other acceptor proteins have
been described, including p53 [21], both subunits of
NF-jB [22], histones, DNA-topoisomerases and the
catalytic subunit of DNA-dependent protein kinase
(DNA-PKcs). Due to the high negative charge of the
polymer, this modification significantly alters the phys-
ical and biochemical properties of the modified pro-
teins, such as their DNA-binding affinity, and it is
likely that such alteration will have a regulatory func-
tion concerning the interaction with other proteins
[23].
Three different PARP-1 knockout mouse models
(Parp1– ⁄ –) have been created independently [24–26],
lacking the PARP-1 protein. Parp1– ⁄ – mice are viable
and fertile, but show hypersensitivity to alkylation
treatment or ionizing radiation and loss of genomic
stability. In stark contrast, however, they display pro-
tection against various pathophysiological phenomena,
such as lipopolysaccharide-induced septic shock or
streptozotocin-induced diabetes [27].
PARP-2
Apart from PARP-1, PARP-2 is the only other PARP
isoform known to be activated by DNA strand breaks
[28] (Table 1; Fig. 2). This enzyme displays automodifi-
cation properties similar to PARP-1. Its DBD, how-
ever, is different from that of PARP-1, consisting of
only 64 amino acids and lacking any obvious DNA-
binding motif. The crystal structure of the catalytic
fragment of murine PARP-2 has recently been solved,
thus providing a basis for the development of isoform-
specific inhibitors by rational drug design [29]. Despite
major structural differences between PARP-1 and
PARP-2, including size and the absence of zinc fingers
or the BRCT motif, they are both targeted to the nuc-
leus, and they bind to and become activated by DNase
I-treated DNA. Both PARP-2 and PARP-1 can homo-
and heterodimerise, and both are involved in the base
excision repair (BER) pathway where they form a
complex with X-ray cross-complementing factor 1
(XRCC1) [18]. Parp1– ⁄ – ⁄Parp2– ⁄ – double mutant mice
are nonviable and die at the onset of gastrulation,
highlighting that the expression of both PARP-1
and PARP-2 and ⁄or DNA strand break-dependent
poly(ADP-ribosyl)ation is essential during early embryo-
genesis.
PARP-3
The protein domain structure of PARP-3 is very
similar to PARP-2, featuring a small DNA-binding
domain consisting of only 54 amino acids and compri-
sing a centrosome-targeting motif [20]. Overexpression
of PARP-3 or its N-terminal domain in HeLa cells
A. Burkle Poly(ADP-ribosyl)ation
FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS 4579

interfered with the G1 ⁄S cell cycle transition. PARP-3
catalyses the synthesis of poly(ADP-ribose) in vitro
and in purified centrosome preparations, and forms a
stable complex with PARP-1, in agreement with other
reports of PARP-1 localization at the centrosome
[30,31].
PARP-4
Vault particles are cytoplasmic ribonucleoprotein parti-
cles (RNPs) composed of several small untranslated
RNA molecules and three proteins of 100, 193 and
240 kDa. With a total mass of 13 MDa, vaults are the
largest RNP complexes found in the cytoplasm of
mammalian cells. The 193 kDa vault protein was iden-
tified as a novel PARP [32,33] and is now termed
PARP-4 (Table 1, Fig. 2). PARP-4 poly(ADP-ribo-
syl)ates the p100 subunit (major vault protein; MVP)
within the vault particle and to a lesser extent itself.
The N-terminal region of PARP-4 contains a BRCT
domain similar to the automodification domain of
PARP-1, suggestive of a related function.
Tankyrase
This protein was initially identified through its inter-
action with the telomeric-repeat binding factor 1
(TRF1), which is another negative regulator of telo-
mere length [34]. The N-terminus of tankyrase contains
a so-called His-Pro-Ser-rich (HPS) domain consisting
of stretches of consecutive histidine, proline and serine
residues, followed by 24 ankyrin (ANK) repeats, which
is a structural feature only found in tankyrase and
tankyrase-2 (see below) within the known members of
the PARP family. Adjacent to the ANK domain is
another protein interaction motif, the sterile alpha-
module. The C-terminus of tankyrase displays homo-
logy to the PARP-1 catalytic region. Consistent with
the absence of any DNA-binding domain, tankyrase
activity does not depend on the presence of DNA
strand breaks but seems to be regulated by the phos-
phorylation state of the protein [35]. About 10% of
cellular tankyrase protein is recruited to telomeres
through binding of its ANK domain to TRF1. Thus
the binding of TRF1 to telomeric DNA controls telo-
mere length in cis by inhibiting the action of telo-
merase at the ends of individual telomeres [36].
Tankyrase-2
This enzyme was originally described as a tumour anti-
gen that elicited antibody responses in certain tumour
patients (Table 1, Fig. 2) [37,38]. Later, tankyrase-2
was reported to interact with several other proteins
such as TRF1 [39], insulin-responsive amino peptidase
(IRAP) [40], or Grb14, an SH2 domain-containing
adaptor protein that binds to the insulin and fibroblast
growth factor receptors [41]. Despite being encoded by
a separate gene, tankyrase-2 displays a domain struc-
ture that is strikingly similar to tankyrase except for
the N-terminal HPS domain, which is missing in
tankyrase-2 (Fig. 2) [39,41]. Tankyrase and tankyrase-
2 also show a significant functional overlap: both pro-
teins possess PARP activity and poly(ADP-ribosyl)ate
some of their interaction partners (IRAP, TAB182 and
TRF1, but not TRF2) as well as themselves, whereas
tankyrase-2 displays preferential automodification
activity. Strikingly, overexpression of tankyrase-2, but
not of tankyrase, caused rapid poly(ADP-ribo-
syl)ation-dependent cell death [39]. Both tankyrases
can self-associate via the sterile alpha-module domain
to form high-molecular-mass complexes, indicative of
a function as master scaffolding molecules in organ-
izing protein complexes [42].
PARG
While there are 18 genes currently known or assumed
to encode different PARP isoforms [2] there is only a
single gene known to encode an enzyme catalysing the
hydrolysis of ADP-ribose polymers to free ADP-
ribose. This is the gene encoding poly(ADP-ribose)
glycohydrolase (PARG; EC 3.2.1.143; Table 1) [43–45].
The human PARG gene shares a 470 bp bidirectional
promoter with the gene encoding the translocase of the
inner mitochondrial membrane 23 (TIM23). Promoter
activity is several fold higher for TIM23 than for
PARG [46]. Three splice variants of the human PARG
have been described, giving rise to PARG isoforms tar-
geted either to the nucleus or to the cytoplasm [47].
Overexpression studies revealed that the largest iso-
form of PARG is targeted to the nucleus while the two
smaller isoforms show mostly cytoplasmic localization.
PARG is an enzyme that possesses both endoglyco-
sidic and exoglycosidic activity and is the only protein
known to catalyse the hydrolysis of ADP-ribose poly-
mers to free ADP-ribose (Fig. 1) [43]. Its products are
free poly(ADP-ribose) and monomeric ADP-ribose,
the latter being a potent protein-glycating carbohy-
drate capable of causing protein damage [48]. Interest-
ingly, an ADPR pyrophosphatase has been described
that converts ADPR to AMP and ribose 5-phosphate,
thus decreasing the risk of nonenzymatic protein gly-
cation [49]. As a consequence of the combined action
of PARPs and PARG, poly(ADP-ribose) undergoes a
dynamic turnover in live cells.
Poly(ADP-ribosyl)ation A. Burkle
4580 FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS

Biological functions of poly(ADP-ribosyl)ation
DNA repair and maintenance of genomic stability
Mechanistic aspects
A plethora of studies have firmly established that DNA
damage-induced poly(ADP-ribosyl)ation contributes to
cellular recovery from cytotoxicity in proliferating cells
inflicted with low or moderate levels of DNA damage
by alkylation, oxidation or ionizing radiation. PARP-1
as well as the related protein PARP-2 are those members
of the PARP family that are responsive to DNA damage
and play important roles in DNA repair and mainten-
ance of genomic integrity, thus behaving as ‘survival fac-
tors’ [50,51]. Specifically, PARP-1 and PARP-2 have
been shown to play a crucial role in the BER pathway.
In mechanistic terms, no clear picture has yet emerged,
despite intense research conducted by many groups.
Attractive scenarios developed over the last couple of
years are (a) the localized relaxation of chromatin at the
site of DNA damage, mediated either by direct modifi-
cation of histones or noncovalent interaction of histones
with poly(ADP-ribose) present as automodification
on PARP-1 or -2; (b) a damage signalling function [3];
or (c) recruitment of specific DNA repair proteins
to the site of damage via noncovalent interaction with
poly(ADP-ribose). These are but a few of the current
hypotheses.
Recent work has added some new interesting
aspects. PARP-1 is a known interaction partner of the
Werner syndrome protein, a protein involved in DNA
repair, maintenance of genomic stability and the pre-
vention of premature ageing. Recently PARP-1 was
shown to regulate both the exonuclease and helicase
activities of the Werner syndrome protein, suggesting a
possible mechanism of action of PARP-1 [52]. Another
scenario that has been put forward is the provision of
ATP from pyrophosphorolytic cleavage of poly(ADP-
ribose). Such ATP could be used for the DNA ligation
step in BER [53]. Accordingly, the decision between
the short-patch and the long-patch pathway in BER in
living cells appears to be dependent on the availability
of ATP. The long-patch pathway would be pre-
ferred under conditions of energy shortage, as this
pathway might lead to increased generation of ATP
from poly(ADP-ribose) via increased provision of
pyrophosphate from deoxynucleotide incorporation
into DNA [54].
On the other hand, recent data clearly show that
DNA-damage induced poly(ADP-ribosyl)ation has
roles to play beyond classical BER. For instance, it
accelerates the repair of oxidative base damage as well
as of UV-induced pyrimidine dimers in a pathway
dependent on Cockayne syndrome B protein [55]. Fur-
thermore, PARP-1 plays a role in regulating double-
strand break repair, independently of p53 [56]. In this
context, a novel route for DNA double-strand breaks
rejoining seems to involve PARP-1 and XRCC1 ⁄DNA
ligase III, i.e. proteins classically associated with
BER [57]. A particular type of DNA damage is
represented by stalled DNA topoisomerase I mole-
cules. Poly(ADP-ribose) reactivates stalled DNA topo-
isomerase I and induces resealing of the DNA strand
breaks caused by topoisomerase stalling, which may be
viewed as a direct repair activity of PARP-1 [58].
Another function of poly(ADP-ribosyl)ation in the
absence of any exogenous DNA damage was highligh-
ted recently. During spermiogenesis, spermatid differ-
entiation is marked by dramatic changes in chromatin
density and composition. The extreme condensation of
the spermatid nucleus is characterized by a shift from
histones to transition proteins and then to protamines
as the major nuclear proteins. Recently poly(ADP-
ribose) formation driven by endogenous DNA strand
breaks was discovered in spermatids of steps 11–14 of
spermiogenesis, i.e. those steps that immediately pre-
cede the most pronounced phase of chromatin con-
densation in spermiogenesis. Transient ADP-ribose
polymer formation may therefore facilitate the process-
ing of DNA strand breaks arising endogenously during
the chromatin remodelling steps of sperm cell matur-
ation [59].
It is interesting to note that there is apparently some
specificity in the downstream consequences of PARP-1
deficiency in cells surviving a genotoxic attack, in that
an increase of deletion mutations and insertions ⁄ rear-rangements was recorded in vivo after treatment with
an alkylating agent [60]. The importance of a proficient
poly(ADP-ribosyl)ation system for the maintenance of
genomic stability and thus the prevention of cancer is
also mirrored in the finding that the V762A genetic
variant of PARP-1 [61,62], which is associated with
diminished enzyme activity, contributes to prostate
cancer susceptibility [63]. This is perfectly in line with
the diminished PARP activity previously recorded in
normal peripheral blood lymphocytes of laryngeal
cancer patients [64].
Apart from experiments aiming at inhibition of cellu-
lar poly(ADP-ribosyl)ation, experimental systems have
also been established to raise cellular poly(ADP-ribose)
levels above normal [65,66]. Supranormal levels of
cellular poly(ADP-ribose) achieved by overexpression
of PARP-1 in cultured cells proved to block genomic
instability, assessed as sister-chromatid exchange,
A. Burkle Poly(ADP-ribosyl)ation
FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS 4581

induced by DNA damage [65], thus yielding the mirror
image of what experiments with PARP inhibition have
shown. Viewed together, the data demonstrate that
poly(ADP-ribose) acts as a negative regulator of DNA-
damage induced genomic instability [67].
PARP inhibitors in cancer treatment
Proficient DNA repair is pivotal to the survival and
maintenance of genomic stability of cells and organ-
isms, given the relentless attack by endogenous and
exogenous DNA-damaging agents. In the setting of
cancer therapy with cytotoxic agents, however, DNA
repair in tumour cells will counteract the desirable cell
killing effect of the treatment. Accordingly, pharmaco-
logical PARP inhibitors, which interfere with DNA
repair pathways, have long been considered a useful
addition to current cancer chemotherapy ⁄ radiotherapyprotocols, drawing on the cocytotoxic effect of
PARP inhibition under conditions of DNA damage
[68]. Very recently, the specific killing of BRCA2-defici-
ent tumour cells by the sole administration of
PARP inhibitors, i.e. in the absence of any exogenous
DNA-damaging treatment, was demonstrated [69,70].
Apparently this phenomenon is related with the defici-
ency of BRCA2-deficient cells to perform homologous
recombination, a pathway most prominently used by
cells lacking PARP-1 activity. These findings should
allow targeting of the DNA repair defect in BRCA-
mutant cells as a new therapeutic strategy for some
forms of human cancer.
Regulation of transcription
It has long been postulated that poly(ADP-ribo-
syl)ation could influence the regulation of gene expres-
sion via regulation of chromatin remodelling [23,71].
Indeed, numerous physical and functional interactions
of PARP-1 with transcription factors have been des-
cribed [72]. PARP-1, for example, plays a pivotal role
in NF-jB-dependent gene expression, which makes it
an important cofactor in immune and inflammatory
responses [73–75]. Furthermore, recent data reveal
functions of PARP-1 in the CaM kinase IId-dependentneurogenic gene activation pathway [76], in the
NAD+-dependent modulation of chromatin structure
and transcription mediated by nucleosome binding of
PARP-1 [12], and in the determination of specificity in
a retinoid signalling pathway via direct modulation of
Mediator [77]. The involvement of poly(ADP-ribo-
syl)ation in long-term potentiation in Aplysia neurones,
occurring in the absence of DNA strand breaks, has
also been linked with transcriptional effects [13].
Implications for apoptosis
The specific cleavage of PARP-1 by caspase-3 ⁄ -7within the nuclear location signal of PARP-1 generates
a 24 kDa and an 89 kDa fragment, and this phenom-
enon has been used extensively as a biochemical mar-
ker of apoptosis. Caspase-mediated PARP-1 cleavage
is thought to cause a loss of stimulation of the cata-
lytic PARP-1 activity in the presence of DNA strand
breaks. Recently a Parp1 knock-in mouse model
(PARP-1(KI ⁄KI)) was reported, in which the caspase
cleavage site of PARP-1 has been mutated so as to
render the protein resistant to caspases during apopto-
sis [78]. Perhaps surprisingly the mice developed nor-
mally. They also proved highly resistant to endotoxic
shock and ischaemia–reperfusion damage, which was
associated with reduced inflammatory responses in the
target tissues and cells, due to the reduced production
of specific inflammatory mediators. Despite normal
binding of NF-jB to DNA, NF-jB-mediated tran-
scription activity was impaired in these knock-in mice,
which explains the above phenotype and creates a new
and unexpected link between PARP-1 cleavage typical
of apoptosis and the regulation of NF-jB, a master
switch in inflammation.
Despite the above-mentioned PARP-1 cleavage,
massive formation of poly(ADP-ribose) can be
observed during the early stages of apoptosis indica-
ting that PARP family proteins are involved in this
process [79,80]. Furthermore it could be shown that
PARP-1 activation is required for the translocation of
apoptosis-inducing factor from the mitochondria to
the nucleus and that apoptosis-inducing factor is neces-
sary for PARP-1-dependent cell death [81].
NAD+ depletion, necrotic cell death and
pathological conditions
Twenty years ago, a mechanism of cell death depend-
ing on the overactivation of PARP-1, and on severe
and irreversible depletion of its substrate NAD+, was
proposed for the first time [82]. Subsequently this para-
digm was confirmed experimentally in mammalian sys-
tems and also in plants [83]. In recent years this
mechanism has been demonstrated to play a major
role in a wide variety of pathophysiological conditions,
including ischaemia–reperfusion damage and a wide
range of inflammatory conditions (reviewed in [84,85]).
Mitochondrial dysfunction triggered by PARP-1 over-
activation seems to play a critical role for the ensuing
cell death [86]. Based on this mechanism, PARP-inhibi-
tory compounds are currently being developed as novel
therapeutics to treat such kind of diseases.
Poly(ADP-ribosyl)ation A. Burkle
4582 FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS

Another intriguing scenario of how the decline of
NAD+ and the rise of nicotinamide triggered by
extensive PARP-1 activation might impact on cellular
physiology is the possible down-regulation of the activ-
ity of silent information regulator 2 (Sir2)-like mam-
malian proteins (sirtuins), a class of NAD+-dependent
deacetylases [87]. Sir2 activity depends on high concen-
tration of NAD+ and is inhibited by nicotinamide.
Sirtuins have been implicated in mediating gene silen-
cing, longevity of organisms and genome stability. It
was proposed that poly(ADP-ribosyl)ation by PARP-
1, which is induced by DNA damage, could modulate
protein deacetylation by Sir2 via the NAD+ ⁄nicotina-mide connection [87].
Relevance of poly(ADP-ribose) catabolism
A PARG loss-of-function mutant was described in
Drosophila melanogaster, lacking the conserved cata-
lytic domain of PARG. This mutant exhibits lethality
in the larval stages at the normal developmental tem-
perature of 25 �C [88]. However, about a quarter of
the mutant fly population progressed to the adult stage
at 29 �C but then displayed progressive neurodegenera-
tion with reduced locomotor activity and a shortened
lifespan. This phenotype was accompanied by extensive
accumulation of poly(ADP-ribose) in the central ner-
vous system. These results suggest that undisturbed
poly(ADP-ribose) metabolism is required for mainten-
ance of the normal function of neuronal cells.
Recently, mouse models with mutant PARG gene
have also been created. The complete loss of PARG
activity induced by disruption of exon 1 led to the fail-
ure of cells to degrade poly(ADP-ribose) and caused
increased sensitivity to cytotoxicity and early embry-
onic lethality [89]. By contrast, combined disruption of
exons 2 and 3 led to the selective depletion of the
110 kDa isoform of the enzyme. This intervention was
compatible with survival and fertility of mice and led
to increased sensitivity to genotoxic and endotoxic
stress in vivo [90]. Viewed together a picture emerges
that loss of PARG activity may produce biological
effects that depend very much on the cellular compart-
ment(s) affected by such loss. This conclusion is rather
sobering, but not implausible in view of the multiple
cellular sites where poly(ADP-ribosyl)ation has been
detected recently (see below).
Centromere function and mitotic spindle
formation
PARP-2 together with PARP-1 has been detected
at centromeres [19,91] where they both interact with
constitutive and transient centromeric proteins [17,92],
indicating that poly(ADP-ribosyl)ation might act as a
regulator of both constitutive kinetochore proteins and
those involved in spindle checkpoint control. Whereas
PARP-2 localization is discrete at the centromere,
PARP-1 shows a broader centromeric and pericentro-
meric distribution [17]. The absence of any drastic
centromeric phenotype in Parp1 knockout mice is sug-
gestive of some functional redundancy for PARP-1 at
the centromere [19]. On the other hand, an increase in
centromeric chromatid breaks observed in Parp2
knockout mice exposed to c-irradiation has been
reported. Furthermore female-specific lethality associ-
ated with X-chromosome instability has been observed
in Parp1+ ⁄ – ⁄Parp2– ⁄ – mice [19]. These data are sug-
gestive of diverse roles of PARP-2 and PARP-1 in
modulating the structure and checkpoint functions of
the mammalian centromere, in particular during radi-
ation-induced DNA damage.
In addition, poly(ADP-ribose) was recently identified
as a new component of the spindle, in addition to the
known major spindle components including micro-
tubules, microtubule-associated proteins and motors
consisting of proteins and DNA [93]. The presence of
poly(ADP-ribose) is required for bipolar spindle
assembly and function.
Centrosomal function
The regulation of centrosome function is crucial to the
accurate transmission of chromosomes to the daughter
cells in mitosis. Both PARP-1 and PARP-3 (Table 1;
Fig. 2) have been identified at centrosomes where they
form a stable complex [20] and poly(ADP-ribosyl)ate
p53 [31]. p53, in turn, has also been shown to localize
at centrosomes and to control centrosome duplication
[94]. Thus both PARP-1 and PARP-3 seem to be
involved in centrosome duplication by modulating p53
activity via poly(ADP-ribosyl)ation. In particular,
PARP-3 localizes preferentially to the daughter centri-
ole throughout the cell cycle [20]. An attractive hypo-
thesis is that the presence of both PARP-1 and
PARP-3 at the centrosome may link the DNA damage
surveillance network to the mitotic fidelity checkpoint.
Tankyrase (Table 1, Fig. 2) is another member of
the PARP family that localizes to the centrosome in a
cell cycle-dependent manner. During mitosis, tankyrase
colocalizes with nuclear mitotic apparatus protein
(NuMa) [95], with which it was shown to form a stable
complex at the centrosome [40]. When NuMa returns
to the nucleus after mitosis this colocalization termin-
ates and tankyrase associates with GLUT4 vesicles
that coalesce around centrosomes [35] and function in
A. Burkle Poly(ADP-ribosyl)ation
FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS 4583

insulin-dependent glucose utilization. Thus, spindle
poles and Golgi apparatus alternately contain most of
cellular tankyrase, whereas only a small fraction of
tankyrase functions at telomeres (see below).
Structure and function of vault particles
As mentioned above, vault particles are large cytoplas-
mic RNPs. Although the cellular function of vaults is
unknown, their subcellular localization and distinct
morphology point to a role in intracellular transport,
particularly nucleo–cytoplasmic transport [96]. It was
reported that vault particles may also be involved in
intracellular detoxification, as all three vault proteins
display increased expression in many multidrug resist-
ant human cell lines examined [97]. PARP-4 is identi-
cal with the 193 kDa vault protein (Table 1, Fig. 2)
and poly(ADP-ribosyl)ates the p100 subunit (major
vault protein; MVP) within the vault particle and to a
lesser extent itself. Immunofluorescence and biochemi-
cal data show that PARP-4 is not exclusively associ-
ated with the vault particle but can also localize to the
nucleolus, the nuclear spindle and to nuclear pores
[32,96]. The enzyme has also been found in association
with mammalian telomerase but is dispensable for
telomerase function and vault structure in vivo [98].
Telomere dynamics
Parp1 knockout mice were reported to have shorter
telomeres than wild-type mice [99]. This observation is
indicative of a role of PARP-1 in maintaining telomere
length and is compatible with the positive correlation
between cellular poly(ADP-ribosyl)ation capacity (lar-
gely reflecting PARP-1 activity) and lifespan of mam-
malian species [100]. Apparently, the differences in
enzyme activity may be due, at least in part, to chan-
ges the primary structure of PARP-1 that arose during
evolution [101].
A functional role of PARP-2 in the maintenance of
telomere integrity is supported by the colocalization of
PARP-2 and telomeric-repeat binding factor 2 (TRF2),
which is a negative regulator of telomere length.
PARP-2 activity regulates the DNA-binding activity of
TRF2 via poly(ADP-ribosyl)ation of the dimerization
domain of TRF2 as well as via noncovalent binding of
poly(ADP-ribose) to the myb domain of TRF2 [102].
This protein interaction may well be involved in modu-
lating t-loop formation in response to DNA damage.
Tankyrase is another negative regulator of telomere
length [34], but surprisingly only about 10% of cellular
tankyrase protein is recruited to telomeres, and
its binding to telomeres is mediated through TRF1.
Poly(ADP-ribosyl)ation of TRF1 by tankyrase inhibits
binding of TRF1 to telomeric DNA and so contributes
to telomere length regulation, by reversing the negative
effect of TRF1 on telomere length [103]. The catalytic
activity of tankyrase is crucial for this effect, because
nuclear overexpression of tankyrase, but not of a
PARP-deficient mutant, causes the lengthening of
telomeres [104]. Additional proteins, however, are also
involved in TRF1 regulation, such as TRF1-interacting
nuclear protein 2 (TIN2), which was reported to pro-
tect TRF1 from poly(ADP-ribosyl)ation by tankyrase
via formation of a ternary complex with TRF1 and
tankyrase, yet without affecting tankyrase automodifi-
cation [105].
As mentioned above, the domain structure of tanky-
rase-2 is very similar to that of tankyrase except for
the N-terminal HPS domain, which is missing in
tankyrase-2 (Fig. 2) [39,41]. Likewise the two enzymes
share several interaction partners including IRAP,
TAB182 and TRF1. Overexpression of either tanky-
rase or tankyrase-2 in the nucleus released endogenous
TRF1 from the telomere, suggesting that the function
of the two enzymes may partially be redundant
[103,104].
Taken together, at least four members of the PARP
family have been implicated in telomere regulation. A
full understanding of their distinct functions at telo-
meres, their regulation and possible functional cooper-
ation will require a substantial amount of additional
research work.
Trafficking of endosomal vesicles
Despite the effect of tankyrase on telomere dynamics,
it should be noted that most of this protein is found
outside the cell nucleus. As mentioned above, it can
either be detected at centrosomes, where it seems to
interact with NuMa [40,95], or in association with
nuclear pore complexes [103] or at Golgi-associated
GLUT4 vesicles [35]. Tankyrase was also reported to
interact with a tankyrase-binding protein of 182 kDa
(TAB182), displaying a heterochromatin-like staining
pattern in the nucleus and colocalizing with cortical
actin in the cytoplasm [106]. Endocytotic vesicles in
myocytes and adipocytes contain the glucose transpor-
ter GLUT4 as well as IRAP. The reversible transloca-
tion of GLUT4 between these GLUT4 vesicles in the
Golgi and the plasma membrane allows insulin to
regulate glucose utilization. Tankyrase appears to be
an important insulin-signalling target, as the protein
not only interacts with IRAP located to GLUT4 stor-
age vesicles in the Golgi, but is also phosphorylated by
mitogen-activated protein kinase (MAPK) upon insulin
Poly(ADP-ribosyl)ation A. Burkle
4584 FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS

stimulation. Tankyrase poly(ADP-ribosyl)ates IRAP,
as well as itself, and this activity is enhanced
by MAPK-mediated phosphorylation indicating that
tankyrase may mediate the long-term regulation of
GLUT4 vesicles by the MAPK cascade [35,107].
Emerging new PARP family members
TIPARP (PARP7)
2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) is a pro-
totype substance from the class of dioxins and causes
pleiotropic effects in mammalian species through
modulating gene expression. A novel, TCDD-inducible
member of the PARP family (TiPARP or PARP7 [2])
was recently identified and characterized (Table 1)
[108,109]. TiPARP mRNA is expressed in a broad
range of mouse tissues and can poly(ADP-ribosyl)ate
histones. Genetic analyses revealed that induction
depends on the aromatic hydrocarbon receptor (AhR)
and on the aromatic hydrocarbon receptor nuclear
translocator (Arnt).
PARP10
Recently a novel member of the PARP family (PARP-
10) has been characterized at the biochemical
level, which is a novel c-myc-interacting protein with
poly(ADP-ribose) polymerase activity (Table 1) [110].
PARP-10 is a 150 kDa protein that interacts with Myc
and possesses a domain with homology to RNA recog-
nition motifs. PARP-10 can poly(ADP-ribosyl)ate itself
and core histones, but neither Myc nor Max, a well-
known c-myc interactor. PARP-10 is localized to the
nuclear and cytoplasmic compartments under the con-
trol of a nuclear export sequence, which also seems to
be relevant for the inhibitory effect of PARP-10 con-
cerning c-Myc- and E1A-mediated cotransformation of
rat embryo fibroblasts.
Conclusion and outlook
The field of poly(ADP-ribosyl)ation is currently a very
exciting one and it has widened in every respect.
Whereas until a couple of years ago a single enzyme
was looked at (PARP-1), we are now dealing with over
a dozen. Whereas DNA strand breakage used to be
considered the only trigger of poly(ADP-ribose) syn-
thesis and consequently the only relevant cellular
condition for poly(ADP-ribose) function, several
additional activation conditions and cellular pheno-
mena related with poly(ADP-ribosyl)ation have been
described. As a consequence, the range of specialists
interested in poly(ADP-ribosyl)ation has broadened
enormously. Obtaining a comprehensive picture of the
biological functions of poly(ADP-ribosyl)ation and the
underlying molecular mechanisms is highly desirable
and would further accelerate the transfer of basic sci-
entific information on this subject to the medical appli-
cation. Hopes that this may be achieved in the not too
distant future are increasing in view of the growing
interest of the scientific community in this field.
References
1 Burkle A (2004) Poly(ADP-ribosyl)ation. Landes Bio-
science, Georgetown, TX, USA.
2 Ame JC, Spenlehauer C & de Murcia G (2004) The
PARP superfamily. Bioessays 26, 882–893.
3 Pleschke JM, Kleczkowska HE, Strohm M & Althaus
FR (2000) Poly(ADP-ribose) binds to specific domains
in DNA damage checkpoint proteins. J Biol Chem 275,
40974–40980.
4 Reale A, Matteis GD, Galleazzi G, Zampieri M &
Caiafa P (2005) Modulation of DNMT1 activity by
ADP-ribose polymers. Oncogene 24, 13–19.
5 Chambon P, Weill JD & Mandel P (1963) Nicotina-
mide mononucleotide activation of new DNA-depen-
dent polyadenylic acid synthesizing nuclear enzyme.
Biochem Biophys Res Commun 11, 39–43.
6 de Murcia G & Menissier de Murcia J (1994)
Poly(ADP-ribose) polymerase: a molecular nick-
sensor. Trends Biochem Sci 19, 172–176.
7 Kun E, Kirsten E & Ordahl CP (2002) Coenzymatic
activity of randomly broken or intact double-stranded
DNAs in auto and histone H1 trans-poly(ADP-ribosy-
lation), catalyzed by poly(ADP-ribose) polymerase
(PARP I). J Biol Chem 277, 39066–39069.
8 Gradwohl G, Mazen A & de Murcia G (1987)
Poly(ADP-ribose) polymerase forms loops with DNA.
Biochem Biophys Res Commun 148, 913–919.
9 Sastry SS & Kun E (1990) The interaction of adenosine
diphosphoribosyl transferase (ADPRT) with a cruciform
DNA. Biochem Biophys Res Commun 167, 842–847.
10 Oei SL, Herzog H, Hirsch-Kauffmann M, Schneider
R, Auer B & Schweiger M (1994) Transcriptional regu-
lation and autoregulation of the human gene for ADP-
ribosyltransferase. Mol Cell Biochem 138, 99–104.
11 Lonskaya I, Potaman VN, Shlyakhtenko LS, Oussat-
cheva EA, Lyubchenko YL & Soldatenkov VA (2005)
Regulation of poly(ADP-ribose) polymerase-1 by DNA
structure-specific binding. J Biol Chem 280, 17076–
17083.
12 Kim MY, Mauro S, Gevry N, Lis JT & Kraus WL
(2004) NAD+-dependent modulation of chromatin
structure and transcription by nucleosome binding
properties of PARP-1. Cell 119, 803–814.
A. Burkle Poly(ADP-ribosyl)ation
FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS 4585

13 Cohen-Armon M, Visochek L, Katzoff A, Levitan D,
Susswein AJ, Klein R, Valbrun M & Schwartz JH
(2004) Long-term memory requires polyADP-ribosyla-
tion. Science 304, 1820–1822.
14 Ruf A, Mennissier de Murcia J, de Murcia G &
Schulz GE (1996) Structure of the catalytic fragment of
poly(ADP-ribose) polymerase from chicken. Proc Natl
Acad Sci USA 93, 7481–7485.
15 Juarez-Salinas H, Sims JL & Jacobson MK (1979)
Poly(ADP-ribose) levels in carcinogen-treated cells.
Nature 282, 740–741.
16 Shieh WM, Ame JC, Wilson MV, Wang ZQ, Koh
DW, Jacobson MK & Jacobson EL (1998) Poly
(ADP-ribose) polymerase null mouse cells synthesize
ADP-ribose polymers. J Biol Chem 273, 30069–
30072.
17 Saxena A, Wong LH, Kalitsis P, Earle E, Shaffer LG
& Choo KH (2002) Poly(ADP-ribose) polymerase 2
localizes to mammalian active centromeres and inter-
acts with PARP-1, Cenpa, Cenpb and Bub3, but not
Cenpc. Hum Mol Genet 11, 2319–2329.
18 Schreiber V, Ame JC, Dolle P, Schultz I, Rinaldi B,
Fraulob V et al. (2002) Poly(ADP-ribose) polymerase-2
(PARP-2) is required for efficient base excision DNA
repair in association with PARP-1 and XRCC1. J Biol
Chem 277, 23028–23036.
19 Menissier de Murcia J, Ricoul M, Tartier L, Nieder-
gang C, Huber A, Dantzer F et al. (2003) Functional
interaction between PARP-1 and PARP-2 in chromo-
some stability and embryonic development in mouse.
Embo J 22, 2255–2263.
20 Augustin A, Spenlehauer C, Dumond H, Menissier-De
Murcia J, Piel M, Schmit AC et al. (2003) PARP-3
localizes preferentially to the daughter centriole and
interferes with the G1 ⁄ S cell cycle progression. J Cell
Sci 116, 1551–1562.
21 Mendoza-Alvarez H & Alvarez-Gonzalez R (2001)
Regulation of p53 sequence-specific DNA-binding by
covalent poly (ADP-ribosyl) ation. J Biol Chem 276,
36425–36430.
22 Kameoka M, Ota K, Tetsuka T, Tanaka Y, Itaya A,
Okamoto T & Yoshihara K (2000) Evidence for regu-
lation of NF-kappaB by poly(ADP-ribose) polymerase.
Biochem J 346 Part 3, 641–649.
23 Rouleau M, Aubin RA & Poirier GG (2004) Poly(ADP-
ribosyl)ated chromatin domains: access granted. J Cell
Sci 117, 815–825.
24 Wang ZQ, Auer B, Stingl L, Berghammer H, Haida-
cher D, Schweiger M & Wagner EF (1995) Mice lack-
ing ADPRT and poly(ADP-ribosyl)ation develop
normally but are susceptible to skin disease. Genes Dev
9, 509–520.
25 de Murcia JM, Niedergang C, Trucco C, Ricoul M,
Dutrillaux B, Mark M et al. (1997) Requirement of
poly(ADP-ribose) polymerase in recovery from DNA
damage in mice and in cells. Proc Natl Acad Sci USA
94, 7303–7307.
26 Masutani M, Nozaki T, Nishiyama E, Shimokawa T,
Tachi Y, Suzuki H et al. (1999) Function of poly(ADP-
ribose) polymerase in response to DNA damage:
gene-disruption study in mice.Mol Cell Biochem 193,
149–152.
27 Shall S & de Murcia G (2000) Poly(ADP-ribose) poly-
merase-1: what have we learned from the deficient
mouse model? Mutat Res 460, 1–15.
28 Ame JC, Rolli V, Schreiber V, Niedergang C, Apiou
F, Decker P et al. (1999) PARP-2, A novel mammalian
DNA damage-dependent poly(ADP-ribose) poly-
merase. J Biol Chem 274, 17860–17868.
29 Oliver AW, Ame JC, Roe SM, Good V, de Murcia G
& Pearl LH (2004) Crystal structure of the catalytic
fragment of murine poly(ADP-ribose) polymerase-2.
Nucleic Acids Res 32, 456–464.
30 Kanai M, Uchida M, Hanai S, Uematsu N, Uchida K
& Miwa M (2000) Poly(ADP-ribose) polymerase loca-
lizes to the centrosomes and chromosomes. Biochem
Biophys Res Commun 278, 385–389.
31 Kanai M, Tong WM, Sugihara E, Wang ZQ, Fuka-
sawa K & Miwa M (2003) Involvement of poly (ADP-
Ribose) polymerase 1 and poly (ADP-Ribosyl) ation in
regulation of centrosome function. Mol Cell Biol 23,
2451–2462.
32 Kickhoefer VA, Siva AC, Kedersha NL, Inman EM,
Ruland C, Streuli M & Rome LH (1999) The 193-kD
vault protein, VPARP, is a novel poly(ADP-ribose)
polymerase. J Cell Biol 146, 917–928.
33 Still IH, Vince P & Cowell JK (1999) Identification of a
novel gene (ADPRTL1) encoding a potential poly(ADP-
ribosyl) transferase protein. Genomics 62, 533–536.
34 Smith S, Giriat I, Schmitt A & de Lange T (1998) Tan-
kyrase, a poly(ADP-ribose) polymerase at human telo-
meres. Science 282, 1484–1487.
35 Chi NW & Lodish HF (2000) Tankyrase is a golgi-
associated mitogen-activated protein kinase substrate
that interacts with IRAP in GLUT4 vesicles. J Biol
Chem 275, 38437–38444.
36 van Steensel B & de Lange T (1997) Control of telo-
mere length by the human telomeric protein TRF1.
Nature 385, 740–743.
37 Monz D, Munnia A, Comtesse N, Fischer U, Steudel
WI, Feiden W, Glass B & Meese EU (2001) Novel tan-
kyrase-related gene detected with meningioma-specific
sera. Clin Cancer Res 7, 113–119.
38 Kuimov AN, Kuprash DV, Petrov VN, Vdovichenko
KK, Scanlan MJ, Jongeneel CV et al. (2001) Cloning
and characterization of TNKL, a member of tankyrase
gene family. Genes Immun 2, 52–55.
39 Kaminker PG, Kim SH, Taylor RD, Zebarjadian Y,
Funk WD, Morin GB, Yaswen P & Campisi J (2001)
TANK2, a new TRF1-associated poly (ADP-ribose)
Poly(ADP-ribosyl)ation A. Burkle
4586 FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS

polymerase, causes rapid induction of cell death upon
overexpression. J Biol Chem 276, 35891–35899.
40 Sbodio JI & Chi NW (2002) Identification of a tankyr-
ase-binding motif shared by IRAP, TAB182, and
human TRF1 but not mouse TRF1. NuMA contains
this RXXPDG motif and is a novel tankyrase partner.
J Biol Chem 277, 31887–31892.
41 Lyons RJ, Deane R, Lynch DKYeZS, Sanderson GM,
Eyre HJ, Sutherland GR & Daly RJ (2001) Identifica-
tion of a novel human tankyrase through its interac-
tion with the adaptor protein Grb14. J Biol Chem 276,
17172–17180.
42 De Rycker M, Venkatesan RN, Wei C & Price CM
(2003) Vertebrate tankyrase domain structure and ster-
ile alpha motif (SAM) -mediated multimerization. Bio-
chem J 372, 87–96.
43 Davidovic L, Vodenicharov M, Affar EB & Poirier
GG (2001) Importance of poly (ADP-ribose) glycohy-
drolase in the control of poly(ADP-ribose) metabolism.
Exp Cell Res 268, 7–13.
44 Shimokawa T, Masutani M, Nozaki T, Nakagama H,
Araki S, Aoki Y & Sugimura T (1998) The human
poly(ADP-ribose) glycohydrolase maps to chromosome
10q11.23–21.1 by fluorescence in situ hybridization.
Hum Cell 11, 243–246.
45 Ame JC, Apiou F, Jacobson EL & Jacobson MK
(1999) Assignment of the poly(ADP-ribose) glycohy-
drolase gene (PARG) to human chromosome 10q11.23
and mouse chromosome 14B by in situ hybridization.
Cytogenet Cell Genet 85, 269–270.
46 Meyer RG, Meyer-Ficca ML, Jacobson EL & Jacob-
son MK (2003) Human poly(ADP-ribose) glycohydro-
lase (PARG) gene and the common promoter sequence
it shares with inner mitochondrial membrane translo-
case 23 (TIM23). Gene 314, 181–190.
47 Meyer-Ficca ML, Meyer RG, Coyle DL, Jacobson EL
& Jacobson MK (2004) Human poly(ADP-ribose) gly-
cohydrolase is expressed in alternative splice variants
yielding isoforms that localize to different cell compart-
ments. Exp Cell Res 297, 521–532.
48 Cervantes-Laurean D, Jacobson EL & Jacobson MK
(1996) Glycation and glycoxidation of histones by
ADP-ribose. J Biol Chem 271, 10461–10469.
49 Bernet D, Pinto RM, Costas MJ, Canales J & Cameselle
JC (1994) Rat liver mitochondrial ADP-ribose pyrophos-
phatase in the matrix space with low Km for free ADP-
ribose. Biochem J 299, 679–682.
50 Bouchard VJ, Rouleau M & Poirier GG (2003) PARP-
1, a determinant of cell survival in response to DNA
damage. Exp Hematol 31, 446–454.
51 Ishizuka S, Martin K, Booth C, Potten CS, de
Murcia G, Burkle A & Kirkwood TB (2003) Poly
(ADP-ribose) polymerase-1 is a survival factor for
radiation-exposed intestinal epithelial stem cells in
vivo. Nucleic Acids Res 31, 6198–6205.
52 von Kobbe C, Harrigan JA, Schreiber V, Stiegler P,
Piotrowski J, Dawut L & Bohr VA (2004) Poly
(ADP-ribose) polymerase 1 regulates both the exonu-
clease and helicase activities of the Werner syndrome
protein. Nucleic Acids Res 32, 4003–4014.
53 Oei SL & Ziegler M (2000) ATP for the DNA ligation
step in base excision repair is generated from poly
(ADP-ribose). J Biol Chem 275, 23234–23239.
54 Petermann E, Ziegler M & Oei SL (2003) ATP-depen-
dent selection between single nucleotide and long patch
base excision repair. DNA Repair (Amst) 2, 1101–
1114.
55 Flohr C, Burkle A, Radicella JP & Epe B (2003)
Poly(ADP-ribosyl) ation accelerates DNA repair in a
pathway dependent on Cockayne syndrome B protein.
Nucleic Acids Res 31, 5332–5337.
56 Susse S, Scholz CJ, Burkle A & Wiesmuller L (2004)
Poly(ADP-ribose) polymerase (PARP-1) and p53 inde-
pendently function in regulating double-strand break
repair in primate cells. Nucl Acids Res 32, 669–680.
57 Audebert M, Salles B & Calsou P (2004) Involvement
of poly(ADP-ribose) polymerase-1 and XRCC1 ⁄DNA
ligase III in an alternative route for DNA double-
strand breaks rejoining. J Biol Chem 279, 55117–55126.
58 Malanga M & Althaus FR (2004) Poly(ADP-ribose)
reactivates stalled DNA topoisomerase I and Induces
DNA strand break resealing. J Biol Chem 279, 5244–
5248.
59 Meyer-Ficca ML, Scherthan H, Burkle A & Meyer RG
(2005) Poly(ADP-ribosyl) ation during chromatin
remodeling steps in rat spermiogenesis. Chromosoma
114, 67–74.
60 Shibata A, Kamada N, Masumura K, Nohmi T,
Kobayashi S, Teraoka H et al. (2005) Parp-1 deficiency
causes an increase of deletion mutations and inser-
tions ⁄ rearrangements in vivo after treatment with an
alkylating agent. Oncogene 24, 1328–1337.
61 Van Gool L, Meyer R, Tobiasch E, Cziepluch C,
Jauniaux JC, Mincheva A et al. (1997) Overexpression
of human poly(ADP-ribose) polymerase in transfected
hamster cells leads to increased poly(ADP-ribosyl)ation
and cellular sensitization to gamma irradiation. Eur J
Biochem 244, 15–20.
62 Cottet F, Blanche H, Verasdonck P, Le Gall I, Schach-
ter F, Burkle A & Muiras ML (2000) New polymorph-
isms in the human poly(ADP-ribose) polymerase-1
coding sequence: lack of association with longevity or
with increased cellular poly(ADP-ribosyl)ation capa-
city. J Mol Med 78, 431–440.
63 Lockett KL, Hall MC, Xu J, Zheng SL, Berwick M,
Chuang SC et al. (2004) The ADPRT V762A genetic
variant contributes to prostate cancer susceptibility and
deficient enzyme function. Cancer Res 64, 6344–6348.
64 Rajaee-Behbahani N, Schmezer P, Ramroth H, Burkle
A, Bartsch H, Dietz A & Becher H (2002) Reduced
A. Burkle Poly(ADP-ribosyl)ation
FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS 4587

poly(ADP-ribosyl)ation in lymphocytes of laryngeal
cancer patients: results of a case-control study. Int J
Cancer 98, 780–784.
65 Meyer R, Muller M, Beneke S, Kupper JH & Burkle
A (2000) Negative regulation of alkylation-induced sis-
ter-chromatid exchange by poly(ADP-ribose) polymer-
ase-1 activity. Int J Cancer 88, 351–355.
66 Brabeck C, Pfeiffer R, Leake A, Beneke S, Meyer R &
Burkle A (2003) 1-selegiline potentiates the cellular
poly(ADP-ribosyl)ation response to ionizing radiation.
J Pharmacol Exp Ther 306, 973–979.
67 Burkle A (2001) Poly(APD-ribosyl)ation, a DNA dam-
age-driven protein modification and regulator of geno-
mic instability. Cancer Lett 163, 1–5.
68 Beneke S, Diefenbach J & Burkle A (2004) Poly(ADP-
ribosyl)ation inhibitors: promising drug candidates for
a wide variety of pathophysiologic conditions. Int J
Cancer 111, 813–818.
69 Bryant HE, Schultz N, Thomas HD, Parker KM,
Flower D, Lopez E et al. (2005) Specific killing of
BRCA2-deficient tumours with inhibitors of poly(ADP-
ribose) polymerase. Nature 434, 913–917.
70 Farmer H, McCabe N, Lord CJ, Tutt AN, Johnson
DA, Richardson TB et al. (2005) Targeting the DNA
repair defect in BRCA mutant cells as a therapeutic
strategy. Nature 434, 917–921.
71 Tulin A & Spradling A (2003) Chromatin loosening by
poly (ADP) -ribose polymerase (PARP) at Drosophila
puff loci. Science 299, 560–562.
72 Kraus WL & Lis JT (2003) PARP goes transcription.
Cell 113, 677–683.
73 Hassa PO & Hottiger MO (2002) The functional role
of poly(ADP-ribose) polymerase 1 as novel coactivator
of NF-kappaB in inflammatory disorders. Cell Mol
Life Sci 59, 1534–1553.
74 Hassa PO, Buerki C, Lombardi C, Imhof R & Hottiger
MO (2003) Transcriptional coactivation of nuclear fac-
tor-kappaB-dependent gene expression by p300 is regu-
lated by poly(ADP)-ribose polymerase-1. J Biol Chem
278, 45145–45153.
75 Carrillo A, Monreal Y, Ramirez P, Marin L, Parrilla
P, Oliver FJ & Yelamos J (2004) Transcription regula-
tion of TNF-alpha-early response genes by poly(ADP-
ribose) polymerase-1 in murine heart endothelial cells.
Nucleic Acids Res 32, 757–766.
76 Ju BG, Solum D, Song EJ, Lee KJ, Rose DW, Glass CK
& Rosenfeld MG (2004) Activating the PARP-1 sensor
component of the groucho ⁄ TLE1 corepressor complex
mediates a CaMKinase IIdelta-dependent neurogenic
gene activation pathway. Cell 119, 815–829.
77 Pavri R, Lewis B, Kim TK, Dilworth FJ, Erdjument-
Bromage H, Tempst P et al. (2005) PARP-1 determines
specificity in a retinoid signaling pathway via direct
modulation of mediator. Mol Cell 18, 83–96.
78 Petrilli V, Herceg Z, Hassa PO, Patel NS, Di Paola R,
Cortes U et al. (2004) Noncleavable poly(ADP-ribose)
polymerase-1 regulates the inflammation response in
mice. J Clin Invest 114, 1072–1081.
79 Negri C, Donzelli M, Bernardi R, Rossi L, Burkle A &
Scovassi AI (1997) Multiparametric staining to identify
apoptotic human cells. Exp Cell Res 234, 174–177.
80 Simbulan-Rosenthal CM, Rosenthal DS, Iyer S,
Boulares AH & SmulsonME (1998) Transient poly(ADP-
ribosyl) ation of nuclear proteins and role of poly(ADP-
ribose) polymerase in the early stages of apoptosis.
J Biol Chem 273, 13703–13712.
81 Yu SW, Wang H, Poitras MF, Coombs C, Bowers
WJ, Federoff HJ et al. (2002) Mediation of poly(ADP-
ribose) polymerase-1-dependent cell death by apopto-
sis-inducing factor. Science 297, 259–263.
82 Berger NA (1985) Poly(ADP-ribose) in the cellular
response to DNA damage. Radiat Res 101, 4–15.
83 De Block M, Verduyn C, De Brouwer D & Cornelissen
M (2005) Poly(ADP-ribose) polymerase in plants
affects energy homeostasis, cell death and stress toler-
ance. Plant J 41, 95–106.
84 Virag L & Szabo C (2002) The therapeutic potential of
poly(ADP-ribose) polymerase inhibitors. Pharmacol
Rev 54, 375–429.
85 Meli E, Pangallo M, Baronti R, Chiarugi A, Cozzi A,
Pellegrini-Giampietro DE & Moroni F (2003)
Poly(ADP-ribose) polymerase as a key player in excito-
toxicity and post-ischemic brain damage. Toxicol Lett
139, 153–162.
86 Cipriani G, Rapizzi E, Vannacci A, Rizzuto R, Moroni
F & Chiarugi A (2005) Nuclear poly (ADP-ribose)
polymerase-1 radpidly triggers mitochondrial dysfunc-
tion. J Biol Chem.
87 Zhang J (2003) Are poly (ADP-ribosyl) ation by
PARP-1 and deacetylation by Sir2 linked? Bioessays
25, 808–814.
88 Hanai S, Kanai M, Ohashi S, Okamoto K, Yamada
M, Takahashi H & Miwa M (2004) Loss of poly
(ADP-ribose) glycohydrolase causes progressive neuro-
degeneration in Drosophila melanogaster. Proc Natl
Acad Sci USA 101, 82–86.
89 Koh DW, Lawler AM, Poitras MF, Sasaki M, Wattler
S, Nehls MC et al. (2004) Failure to degrade poly
(ADP-ribose) causes increased sensitivity to cytotoxici-
ty and early embryonic lethality. Proc Natl Acad Sci
USA 101, 17699–17704.
90 Cortes U, Tong WM, Coyle DL, Meyer-Ficca ML,
Meyer RG, Petrilli V et al. (2004) Depletion of the
110-kilodalton isoform of poly (ADP-ribose) glyco-
hydrolase increases sensitivity to genotoxic and endo-
toxic stress in mice. Mol Cell Biol 24, 7163–7178.
91 Earle E, Saxena A, MacDonald A, Hudson DF, Shaf-
fer LG, Saffery R et al. (2000) Poly (ADP-ribose) poly-
Poly(ADP-ribosyl)ation A. Burkle
4588 FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS

merase at active centromeres and neocentromeres at
metaphase. Hum Mol Genet 9, 187–194.
92 Saxena A, Saffery R, Wong LH, Kalitsis P & Choo
KH (2002) Centromere proteins Cenpa, Cenpb, and
Bub3 interact with poly(ADP-ribose) polymerase-1
protein and are poly(ADP-ribosyl)ated. J Biol Chem
277, 26921–26926.
93 Chang P, Jacobson MK & Mitchison TJ (2004) Poly
(ADP-ribose) is required for spindle assembly and
structure. Nature 432, 645–649.
94 Tarapore P, Tokuyama Y, Horn HF & Fukasawa K
(2001) Difference in the centrosome duplication regula-
tory activity among p53 ‘hot spot’ mutants: potential
role of Series 315 phosphorylation-dependent centro-
some binding of p53. Oncogene 20, 6851–6863.
95 Smith S & de Lange T (1999) Cell cycle dependent
localization of the telomeric PARP, tankyrase, to
nuclear pore complexes and centrosomes. J Cell Sci
112 (21), 3649–3656.
96 van Zon A, Mossink MH, Schoester M, Houtsmuller
AB, Scheffer GL, Scheper RJ et al. (2003) The forma-
tion of vault-tubes: a dynamic interaction between
vaults and vault PARP. J Cell Sci 116, 4391–4400.
97 Siva AC, Raval-Fernandes S, Stephen AG, LaFemina
MJ, Scheper RJ, Kickhoefer VA & Rome LH (2001)
Up-regulation of vaults may be necessary but not suffi-
cient for multidrug resistance. Int J Cancer 92, 195–
202.
98 Liu Y, Snow BE, Kickhoefer VA, Erdmann N, Zhou
W, Wakeham A et al. (2004) Vault poly(ADP-ribose)
polymerase is associated with mammalian telomerase
and is dispensable for telomerase function and vault
structure in vivo. Mol Cell Biol 24, 5314–5323.
99 d’Adda di Fagagna F, Hande MP, Tong WM, Lansdorp
PM, Wang ZQ & Jackson SP (1999) Functions of
poly(ADP-ribose) polymerase in controlling telomere
length and chromosomal stability. Nat Genet 23, 76–80.
100 Grube K & Burkle A (1992) Poly(ADP-ribose) poly-
merase activity in mononuclear leukocytes of 13 mam-
malian species correlates with species-specific life span.
Proc Natl Acad Sci USA 89, 11759–11763.
101 Beneke S, Alvarez-Gonzalez R & Burkle A (2000)
Comparative characterisation of poly(ADP-ribose)
polymerase-1 from two mammalian species with differ-
ent life span. Exp Gerontol 35, 989–1002.
102 Dantzer F, Giraud-Panis MJ, Jaco I, Ame JC, Schultz
I, Blasco M et al. (2004) Functional interaction
between poly(ADP-ribose) polymerase 2 (PARP-2) and
TRF2: PARP activity negatively regulates TRF2. Mol
Cell Biol 24, 1595–1607.
103 Smith S & de Lange T (2000) Tankyrase promotes telo-
mere elongation in human cells. Curr Biol 10, 1299–
1302.
104 Cook BD, Dynek JN, Chang W, Shostak G & Smith S
(2002) Role for the related poly(ADP-ribose) poly-
merases tankyrase 1 and 2 at human telomeres. Mol
Cell Biol 22, 332–342.
105 YeJZ & de Lange T (2004) TIN2 is a tankyrase 1
PARP modulator in the TRF1 telomere length control
complex. Nat Genet 36, 618–623.
106 Seimiya H & Smith S (2002) The telomeric poly(ADP-
ribose) polymerase, tankyrase 1, contains multiple
binding sites for telomeric repeat binding factor 1
(TRF1) and a novel acceptor, 182-kDa tankyrase-bind-
ing protein (TAB182). J Biol Chem 277, 14116–14126.
107 Sbodio JI, Lodish HF & Chi NW (2002) Tankyrase-2
oligomerizes with tankyrase-1 and binds to both TRF1
(telomere-repeat-binding factor 1) and IRAP (insulin-
responsive aminopeptidase). Biochem J 361, 451–459.
108 Ma Q, Baldwin KT, Renzelli AJ, McDaniel A &
Dong L (2001) TCDD-inducible poly(ADP-ribose)
polymerase: a novel response to 2,3,7,8-tetrachloro-
dibenzo-p-dioxin. Biochem Biophys Res Commun 289,
499–506.
109 Ma Q (2002) Induction and superinduction of
2,3,7,8-tetrachlorodibenzo-rho-dioxin-inducible
poly(ADP-ribose) polymerase: role of the aryl hydro-
carbon receptor ⁄ aryl hydrocarbon receptor nuclear
translocator transcription activation domains and a
labile transcription repressor. Arch Biochem Biophys
404, 309–316.
110 Yu M, Schreek S, Cerni C, Schamberger C, Lesniewicz
K, Poreba E et al. (2005) PARP-10, a novel Myc-
interacting protein with poly(ADP-ribose) polymerase
activity, inhibits transformation. Oncogene 24,
1982–1993.
A. Burkle Poly(ADP-ribosyl)ation
FEBS Journal 272 (2005) 4576–4589 ª 2005 FEBS 4589