Embed Size (px)
Citation preview

Molecular Ecology Notes (2002),
2
, 273–275 doi:10.1046/j.1471-8278 .2002.00220.x
© 2002 Blackwell Science Ltd
Blackwell Science, Ltd
PRIMER NOTE
Polymorphic microsatellite loci for eusocial wasps (Hymenoptera: Vespidae)
D . DALY,* M. E . ARCHER,† P . C . WATTS, M. P . SPEED,‡ M. R . HUGHES,§ F . S . BARKER,* J . JONES, K . ODGAARD* and S . J . KEMP**
Laboratory 1.03 g, Donnan Laboratories, Crown Street, School of Biological Sciences, University of Liverpool, Liverpool. L69 7ZD. UK,
†
College of Ripon and York St John, York. YO31 7EX. UK,
‡
Human & Biological Sciences, Liverpool Hope, Hope Park, Chidwall, Liverpool, L16 9JD. UK,
§
Silicon Genetics, 2601, Spring Street, Redwood City, CA 94063, USA
Abstract
Hymenopterans are an important model for studying the evolution of cooperation in animalsocieties. Here, we characterize 19 microsatellite loci, isolated from the common wasp
Vespulavulgaris
, that can be used to study genetic variation in three genera (seven species) within theVespidae. The number of alleles in
V. vulgaris
was moderate, varying from 2 to 14, with expectedheterozygosity ranging between 0.04 and 0.93. Eleven loci amplified DNA in
V. austriaca
and
Dolichovespula sylvestris
, nine in
V. germanica
, eight in
Vespa crabro
and
V. rufa
, sevenin
D. media
and only five loci could be used for
D. norwegica
.
Keywords
:
Hymenoptera, Microsatellite, Vespidae,
Vespula vulgaris
, VNTR
Received 29 January 2002; revision received 6 March 2002; accepted 6 March 2002
Establishing genetic relationships within and among popula-tions is central to our understanding of how cooperationhas evolved within animal societies. The eusocial Hyme-noptera (ants, bees and wasps) are an important modelsystem for the study of the evolution of cooperation, andone group that has received considerable recent attention isthe Vespidae. In vespine colonies the relatedness of workerswithin colonies is believed to be a key component in deter-mining how the conflict between the queens and workersover male worker production is resolved (see Foster &Ratnieks 1999, 2001). At present, however, only a fewmicrosatellite loci derived from a single species of wasp,
Vespula rufa
(Thorén
et al
. 1995) are available to research thebreeding structure of this group. Here, we describe a panelof polymorphic microsatellite loci that can be used toinvestigate the molecular ecology of the common wasp
Vespula vulgaris
and seven other European vespid species.A size-selected partial genomic library (400–900 bp) was
constructed using DNA isolated from a single worker ofthe common wasp
Vespula vulgaris
collected from SouthLiverpool during 1998. Ten micrograms of genomic DNA,prepared using the protocol of Strassmann
et al
. (1996), wasdigested with
Sau
IIIA, ligated into dephosphorylated
pUC18 (Boehringer Mannheim) digested with
Bam
HI andthen transformed into
Escherichia coli
competent cells(Stratagene). Clones containing microsatellite sequenceswere detected using a digoxygenin nucleic acid detectionkit (Boehringer Mannheim) with hybridization and detec-tion conditions as described by Estoup & Turgeon. (1996).Probing was performed using three different combinationsof oligonucleotide; (i) (AAG)
7
(AAC)
7
(AAAG)
6
(GATA)
5
and (CA)
10;
(ii) (CAT)
7
(TAG)
7
(AAGG)
6
and (CAG)
7;
and(iii) (AAT)
7
. Plasmids containing positive clones were puri-fied and then cycle sequenced using Big Dye™ chemistry(PE Applied Biosystems) and electrophoresis on anABI377. Primers to amplify microsatellite core repeatswere designed using
primer
3 (Rozen & Skaletsky 1998).Primers were initially tested on
V. vulgaris
workers selectedfrom a variety of locations in south Liverpool. Polymerasechain reaction (PCR) was undertaken in a 5-
µ
L reactionvolume using a PTC-100–96 V thermal cycler (MJ ResearchInc). Each reaction mix contained 2.5
µ
L of Reddy-LoadPCR Master Mix (0.0625 units
Taq
DNA polymerase,75 m
m
Tris-HCl (pH 8.8), 20 m
m
(NH)
4
SO
4
, 1.5 m
m
MgCl
2
,0.01% (v/v) Tween 20 and 0.2 m
m
of each dNTP) (AdvanceBiotechnologies), 0.05
µ
L of forward and reverse primer(20 pmol
/
µ
L) (MWG Biotech) and 1.0
µ
L of template DNA(20 ng/
µ
L). PCR cycle conditions were: (i) 1 min at 95
°
C;(ii) 6 cycles of 30 s at 95
°
C, 30 s at 55
°
C and 45 s at 72
°
C;
Correspondence: Steve Kemp. Fax: 44 (0) 151 7943655 E-mail:[email protected]
men_220.fm Page 273 Friday, August 16, 2002 9:30 AM
274
P R I M E R N O T E
© 2002 Blackwell Science Ltd,
Molecular Ecology
Notes
, 2, 273–275
(iii) 26 cycles of 30 s at 92
°
C, 30 s at 55
°
C and 55 s at 72
°
C;and (iv) 72
°
C for 30 min The forward primer was end-labelledwith [
γ
33
P]-ATP. Allelic variation was determined by verticalgel electrophoresis on a 6% denaturing polyacrylamide gel(Severn Biotech). Gels were run at 90 W for 3.5 h, exposedovernight to a phosphor screen and scanned using aSTORM phosphor imager (Molecular Dynamics Inc).
All polymorphic loci were tested on 16 workers (fromthe same colony) of seven other species of European Vespi-dae:
Dolichovespula media
collected from Liverpool during1999,
V. germanica
,
V. rufa
,
V. austriaca
,
D. sylvestris
and
D. norwegica
collected from the Yorkshire area between 1976and 1979, and
Vespa crabro
caught in Cambridgeshire during1983. PCR conditions were as above but with the annealingtemperature reduced to 50
°
C for some locus-species com-binations (see Table 2). The forward primers for this phasewere labelled with either IRD800 or IRD700 fluorescentdye (MWG Biotech, Germany) and the PCR products visu-alized by electrophoresis through an 8% denaturing poly-acrylamide gel on a LiCor 4200 DNA sequencer.
Nineteen polymorphic microsatellite loci were isolatedin
V. vulgaris
(Table 1). Three pairs of loci are situated along
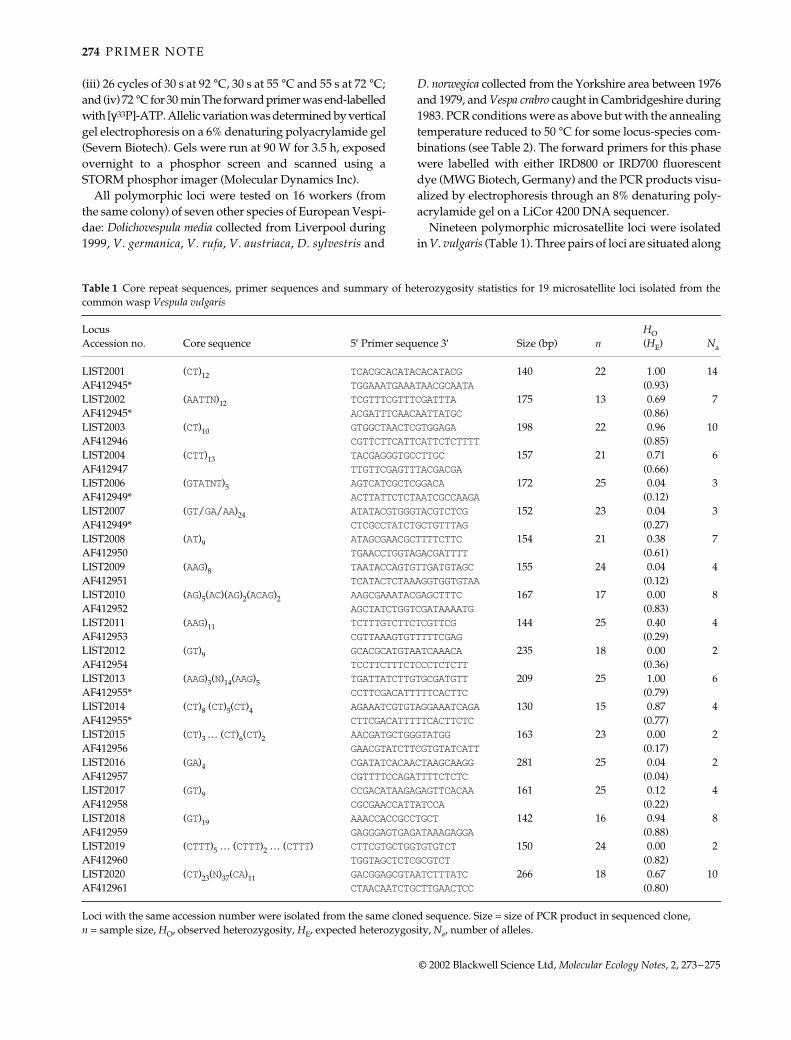
Table 1 Core repeat sequences, primer sequences and summary of heterozygosity statistics for 19 microsatellite loci isolated from thecommon wasp Vespula vulgaris
Locus Accession no. Core sequence 5′ Primer sequence 3′ Size (bp) n
HO (HE) Na
LIST2001 (CT)12 TCACGCACATACACATACG 140 22 1.00 14AF412945* TGGAAATGAAATAACGCAATA (0.93)LIST2002 (AATTN)12 TCGTTTCGTTTCGATTTA 175 13 0.69 7AF412945* ACGATTTCAACAATTATGC (0.86)LIST2003 (CT)10 GTGGCTAACTCGTGGAGA 198 22 0.96 10AF412946 CGTTCTTCATTCATTCTCTTTT (0.85)LIST2004 (CTT)13 TACGAGGGTGCCTTGC 157 21 0.71 6AF412947 TTGTTCGAGTTTACGACGA (0.66)LIST2006 (GTATNT)5 AGTCATCGCTCGGACA 172 25 0.04 3AF412949* ACTTATTCTCTAATCGCCAAGA (0.12)LIST2007 (GT/GA/AA)24 ATATACGTGGGTACGTCTCG 152 23 0.04 3AF412949* CTCGCCTATCTGCTGTTTAG (0.27)LIST2008 (AT)9 ATAGCGAACGCTTTTCTTC 154 21 0.38 7AF412950 TGAACCTGGTAGACGATTTT (0.61)LIST2009 (AAG)8 TAATACCAGTGTTGATGTAGC 155 24 0.04 4AF412951 TCATACTCTAAAGGTGGTGTAA (0.12)LIST2010 (AG)5(AC)(AG)2(ACAG)2 AAGCGAAATACGAGCTTTC 167 17 0.00 8AF412952 AGCTATCTGGTCGATAAAATG (0.83)LIST2011 (AAG)11 TCTTTGTCTTCTCGTTCG 144 25 0.40 4AF412953 CGTTAAAGTGTTTTTCGAG (0.29)LIST2012 (GT)9 GCACGCATGTAATCAAACA 235 18 0.00 2AF412954 TCCTTCTTTCTCCCTCTCTT (0.36)LIST2013 (AAG)3(N)14(AAG)5 TGATTATCTTGTGCGATGTT 209 25 1.00 6AF412955* CCTTCGACATTTTTCACTTC (0.79)LIST2014 (CT)8 (CT)5(CT)4 AGAAATCGTGTAGGAAATCAGA 130 15 0.87 4AF412955* CTTCGACATTTTTCACTTCTC (0.77)LIST2015 (CT)3 … (CT)6(CT)2 AACGATGCTGGGTATGG 163 23 0.00 2AF412956 GAACGTATCTTCGTGTATCATT (0.17)LIST2016 (GA)4 CGATATCACAACTAAGCAAGG 281 25 0.04 2AF412957 CGTTTTCCAGATTTTCTCTC (0.04)LIST2017 (GT)9 CCGACATAAGAGAGTTCACAA 161 25 0.12 4AF412958 CGCGAACCATTATCCA (0.22)LIST2018 (GT)19 AAACCACCGCCTGCT 142 16 0.94 8AF412959 GAGGGAGTGAGATAAAGAGGA (0.88)LIST2019 (CTTT)5 … (CTTT)2 … (CTTT) CTTCGTGCTGGTGTGTCT 150 24 0.00 2AF412960 TGGTAGCTCTCGCGTCT (0.82)LIST2020 (CT)23(N)37(CA)11 GACGGAGCGTAATCTTTATC 266 18 0.67 10AF412961 CTAACAATCTGCTTGAACTCC (0.80)
Loci with the same accession number were isolated from the same cloned sequence. Size = size of PCR product in sequenced clone, n = sample size, HO, observed heterozygosity, HE, expected heterozygosity, Na, number of alleles.
men_220.fm Page 274 Friday, August 16, 2002 9:30 AM
P R I M E R N O T E 275
© 2002 Blackwell Science Ltd, Molecular Ecology Notes, 2, 273–275
the same sequence: LIST2001 & LIST2002; LIST2006 &LIST2007; LIST2013 & LIST20014 (Table 1), and may beparticularly useful in providing haplotypes when determin-ing parentage. The longest uninterrupted core sequencesare generally provided in Table 1 for brevity, althoughit should be noted that the majority of the loci had runs ofinterrupted or compound repeat sequences flanking thecore microsatellite region (see sequences in GenBankdata for further information). Eight loci have observedheterozygosities over 0.66 and seven loci had observedheterozygosities between 0.04 and 0.38 (Table 1). For someloci there were large discrepancies between observedand expected heterozygosities, with 5 loci having multiplealleles but no heterozygotes (Table 1). This discrepancymay be explained by the collection of individual waspsfrom a number of unrelated nests. Overall, the numberof alleles at each locus was moderate, varying between2 and 14.
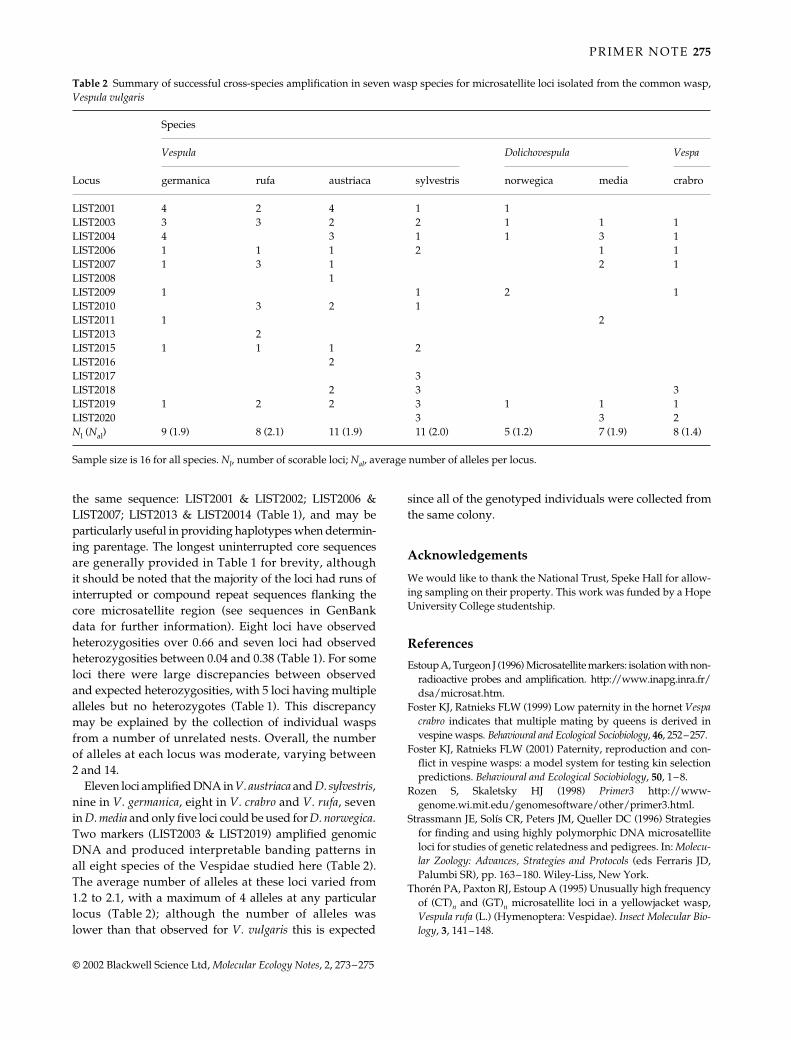
Eleven loci amplified DNA in V. austriaca and D. sylvestris,nine in V. germanica, eight in V. crabro and V. rufa, sevenin D. media and only five loci could be used for D. norwegica.Two markers (LIST2003 & LIST2019) amplified genomicDNA and produced interpretable banding patterns inall eight species of the Vespidae studied here (Table 2).The average number of alleles at these loci varied from1.2 to 2.1, with a maximum of 4 alleles at any particularlocus (Table 2); although the number of alleles waslower than that observed for V. vulgaris this is expected
since all of the genotyped individuals were collected fromthe same colony.
Acknowledgements
We would like to thank the National Trust, Speke Hall for allow-ing sampling on their property. This work was funded by a HopeUniversity College studentship.
References
Estoup A, Turgeon J (1996) Microsatellite markers: isolation with non-radioactive probes and amplification. http://www.inapg.inra.fr/dsa/microsat.htm.
Foster KJ, Ratnieks FLW (1999) Low paternity in the hornet Vespacrabro indicates that multiple mating by queens is derived invespine wasps. Behavioural and Ecological Sociobiology, 46, 252–257.
Foster KJ, Ratnieks FLW (2001) Paternity, reproduction and con-flict in vespine wasps: a model system for testing kin selectionpredictions. Behavioural and Ecological Sociobiology, 50, 1–8.
Rozen S, Skaletsky HJ (1998) Primer3 http://www-genome.wi.mit.edu/genomesoftware/other/primer3.html.
Strassmann JE, Solís CR, Peters JM, Queller DC (1996) Strategiesfor finding and using highly polymorphic DNA microsatelliteloci for studies of genetic relatedness and pedigrees. In: Molecu-lar Zoology: Advances, Strategies and Protocols (eds Ferraris JD,Palumbi SR), pp. 163–180. Wiley-Liss, New York.
Thorén PA, Paxton RJ, Estoup A (1995) Unusually high frequencyof (CT)n and (GT)n microsatellite loci in a yellowjacket wasp,Vespula rufa (L.) (Hymenoptera: Vespidae). Insect Molecular Bio-logy, 3, 141–148.
Table 2 Summary of successful cross-species amplification in seven wasp species for microsatellite loci isolated from the common wasp,Vespula vulgaris
Locus
Species
Vespula Dolichovespula Vespa
germanica rufa austriaca sylvestris norwegica media crabro
LIST2001 4 2 4 1 1LIST2003 3 3 2 2 1 1 1LIST2004 4 3 1 1 3 1LIST2006 1 1 1 2 1 1LIST2007 1 3 1 2 1LIST2008 1LIST2009 1 1 2 1LIST2010 3 2 1LIST2011 1 2LIST2013 2LIST2015 1 1 1 2LIST2016 2LIST2017 3LIST2018 2 3 3LIST2019 1 2 2 3 1 1 1LIST2020 3 3 2Nl (Nal) 9 (1.9) 8 (2.1) 11 (1.9) 11 (2.0) 5 (1.2) 7 (1.9) 8 (1.4)
Sample size is 16 for all species. Nl, number of scorable loci; Nal, average number of alleles per locus.
men_220.fm Page 275 Friday, August 16, 2002 9:30 AM





























