Embed Size (px)
Citation preview

Positional differences in size, morphology, and in vitro performance of pea axillary buds
K. S. GOULD' A N D E. G. CUTTER Department of Botany, University of Manchester, Manchester, M I 3 9PL United Kingdom
AND
J. P. W. YOUNG John lnnes Institute, Colney Lane, Norwich, NR4 7UH United Kingdorrz
AND
W. A. CHARLTON Department of Botany, University of Manchester, Manchester, M I 3 9PL United Kingdom
Received December 6, 1985
GOULD, K. S., E. G. CUTTER, J . P. W. YOUNG, and W. A. CHARLTON. 1987. Positional differences in size, morphology, and in vitro performance of pea axillary buds. Can. J . Bot. 65: 406-41 1.
Numbers of buds within a leaf axil and of leaf primordia within a bud varied with node of insertion, both in intact pea (Pisurn sativum) seedlings and in cultured axillary shoots. Normally one or more nodes bore no visible buds. At the higher nodes naked buds and aberrant forms were observed. Shoots dissected from the embryo bore five or six leaf primordia and buds were present at the cotyledonary node and at three nodes immediately above. Benzylaminopurine in the nutrient medium promoted vegetative growth of cultured shoots. The height and extent of proliferation of cultured shoots varied both with the parental node from which explants were derived and with benzylaminopurine concentration. Results are discussed in relation to come- lative inhibition.
GOULD, K. S., E. G. CUTTER, J . P. W. YOUNG et W. A. CHARLTON. 1987. Positional differences in size, morphology, and in vitro performance of pea axillary buds. Can. J . Bot. 65 : 406-41 1.
Le nombre de bourgeons prCsents dans l'aisselle d'une feuille et le nombre de primordiums foliaires dans un bourgeon variaient selon le noeud d'insertion chez des semis intacts du pois (Pisum sativum) et chez des pousses axillaires en culture. Normalement, un noeud ou plus ne portaient pas de bourgeons visibles. Aux noeuds supCrieurs, des bourgeons nus et des formes anormales Ctaient presents. Les pousses prClevCes de l'embryon portaient cinq ou six primordiums foliaires et des bour- geons Ctaient presents au noeud cotylCdonnaire et aux trois noeuds immidiatement au-dessus. La prCsence de benzylamino- purine dans le milieu nutritif encourageait la croissance vCgCtative des pousses. La hauteur et le degrC de prolifCration des pousses en culture variaient B la fois selon le noeud parental d'oh provenaient les explants et selon la concentration de benzyla- minopurine. Les auteurs discutent des rCsultats par rapport B I'inhibition comClative.
[Traduit par la revue]
Introduction The in vitro culture of axillary shoots is commonly used for
propagating herbaceous species en masse and as a tool for studying organogenesis. workers frequently omit to indicate the node of insertion from which explants were derived. More- over, Phillips (1975) stated that "all vegetative buds on a plant possess essentially equal developmental potential. " However, differential responses to growth regulators and nutrients have been reported for buds along intact and decapitated shoots. Cutter and Chiu (1975) found that lateral shoots of Hygrophila sp. grew out first at basal nodes of intact shoots on a medium containing benzyladenine and decreased in size acropetally. In decapitated shoots on the same medium lateral shoots of approximately equal size grew out at all nodes. Following work with decapitated pea seedlings Husain and Linck (1966) concluded that "the growth potential of a particular bud seems to be related to its location on the stem." Nodes were numbered acropetally above the seed cotyledons. When plants were decapitated from below node 3 up to below node 5 , the shoot at node 2 invariably dominated. Decapitation at any place above node 6 resulted in the outgrowth of the highest axillary bud. The radioisotope 32P moved preferentially to those lateral shoots with greatest growth potential in decap-
'Present address: Department of Botany and Plant Sciences, University of California, Riverside, CA, U.S.A. 92521.
itated plants. McDaniel (1978, 1984) and McDaniel and Hsu (1976) showed by means of grafting and other experiments that axillary buds in tobacco fell into two categories: those that were close to the tip of the stem, the developmental potential of which could not be modified, and those farther down the stem, which could be caused to produce a greater or lesser number of leaves by altering their position. In this paper we report differences in the growth of buds along both intact plants and isolated axillary shoots of peas.
We describe a number of simple observations showing clear positional differences in size and number of axillary buds along intact shoots (i.e., first order) and axillary buds along cultured explants (i.e., second order relative to the seedling axis). Furthermore, such differences can markedly affect the response of isolated nodal segments in culture to the cytokinin 6-benzylaminopurine (BAP), which is commonly added to nutrient media to promote shoot growth (Hussey 1978).
Materials and methods
Plant material As part of a larger study, three isogenic lines with contrasting leaf
morphology have been compared: the conventional genotype, Af TI, bearing leaves with the normal assemblage of pairs of proximal leaflets and distal tendrils; the "afila" genotype, af TI, in which ten- drils occupy normal leaflet sites (Kujala 1953; Goldenberg 1965); and the "tendril-less" genotype, Af t l , in which leaflets occupy normal tendril sites (Vilmorin and Bateson 1912; White 1917).
Printed in Canada / lmpnmt au Canada
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F W
YO
MIN
G o
n 09
/20/
13Fo
r pe
rson
al u
se o
nly.

GOULD ET AL.
Seeds from the isogenic lines JI 1194 (conventional), JI 1195 (afila), and JI 1197 (tendril-less), derived from a programme of back- crossing by Dr. G. A. Marx, New York State Agricultural Experi- ment Station, Geneva, NY, U.S.A., were provided by John Innes Institute.
Axillary buds in intact plants Seeds were sown in vermiculite, saturated and subinigated with
deionized water, and grown for 21 days in a controlled-environment room maintained at 22°C with an inadiance of 25 W m-' at plant level and an 18-h photoperiod. Shoots harvested at this stage typically bore seven mature leaves, one expanding leaf, and three or four leaf prim- ordia. Nodes were numbered acropetally above the cotyledons. Scale . - leaves occurred at nodes 1 and 2; younger leaves, whi;h were foliage leaves and progressively more complex up the shoot, have been des- cribed in detail elsewhere (Gould et al. 1986). Grown in the absence of exogenous nutrients, internode elongation and rates of leaf initi- ation were negligible by the 21-day stage.
The number of buds, and the number of leaf primordia within each bud, in the axils of the cotyledons and leaves at nodes 1 to 8 were recorded, using a dissecting microscope, for 10 plants of each genotype.
In vitro technique Surface-sterilised seed was sown in autoclaved vermiculite mois-
tened with sterile water, in 100-mL polystyrene screw-topped, presterilised jars (Richardsons Ltd, Leicester). After 10 days growth in a controlled-environment room under the conditions described above, all stipules, leaves, and the shoot apex of each plant were removed. Nodal segments were excised and inserted in 20 mL nutrient agar in fresh polystyrene jars. The medium contained the basic salts of Murashige and Skoog (1962) as modified by Ziv et al. (1970), Hussey and Gunn (1984), and Gould et al. (1986). The standard medium contained the cytokinin at 0.75 mg L", introduced prior to autoclaving.
Explants thus obtained were examined after 6 weeks in the con- trolled-environment room, following the cessation of vegetative growth of the terminal, first-order shoot but prior to the outgrowth of second-order lateral buds. Each explant typically bore 9- 12 mature leaves above a basal callus. The oldest leaf was a scale leaf; younger leaves were foliage leaves and progressively more complex up the shoot (Gould et al. 1986). Nodes were numbered acropetally above the basal callus. The number of axillary buds at each of the first eight nodes and the number of leaf primordia within each bud were recorded for 10 explants per genotype (derived from nodes 1 and 2 of five parent seedlings).
Embryonic shoots Seeds of the conventional genotype were soaked in water for 2 h;
then the embryonic shoots were removed and fixed in 2.5% glutaraldehyde in 0.025 M phosphate buffer, pH 6.8, overnight at room temperature (Glauert 1975). The material was dehydrated through a methoxyethanol -ethanol -propano1 -n-butanol series (Feder and O'Brien 1968) and infiltrated and embedded in glycol methacrylate ("JB4," Polysciences Ltd). Serial longitudinal sections, 3 pm thick, were taken with glass knives in the plane of the distichous leaf insertion and stained with 0.5 % (wlv) toluidine blue in 0.5 % (wlv) borax solution. The number of leaf primordia and the occur- rence of buds within the leaf axils were recorded for five embryonic shoots.
Abenant axillary structures were processed for sectioning by an identical procedure.
Response to cytokinin Nodal segments of plants grown from seed under sterile conditions
were transferred to the nutrient medium containing BAP at a concen- tration of 0, 0.75, 5.0, 10.0, or 50.0 mg L-'. The morphology of eight explants derived from two parent seedlings (taken from the cotyledonary node, and from nodes 1 -3) per genotype was examined for each concentration of BAP after 5 weeks in the controlled- environment room.
Buds Pr imordia
I
Node o f insert ion
Node of insertion
FIGS. 1 and 2. Mean number of axillary buds and of leaf primordia within the largest bud at consecutive nodes of insertion. Fig. 1. Intact seedlings. Fig. 2. Cultured shoots. Bars show 95% confidence inter- vals (from Student's Q range).
In addition, nodal segments bearing second-order buds taken from 10 explants per genotype that had been cultured in the standard medium for 6 weeks were subcultured on nutrient medium containing 10 mg L-' BAP. The response of explants derived from the first four nodes of each parent explant was observed after 4 weeks.
Results Axillary buds
In intact shoots two buds were invariably associated with each of the cotyledons. There were no significant differences among genotypes in the number of buds present at each node, or in the number of leaf primordia within each bud (analysis of variance; p > 0.05), and data from the three genotypes have been pooled. The mean number of buds per axil rose to a maxi- mum at node 2 in intact seedlings and thereafter decreased at successively higher nodes (Fig. 1). The maximum recorded number of buds in a leaf axil was four. Where more than one bud was present, invariably one bud was larger than the remainder; it was impossible to determine from superficial examination alone whether all buds were independent or whether the smaller buds were, in fact, subsidiaries of the largest.
In the cultured shoots the maximum mean number of axillary buds occurred at node 1 (which subtends the only scale leaf). The mean number of buds per axil decreased through suc- cessive nodes to a minumum at node 5 and thereafter increased slightly (Fig. 2).
There was a positive and highly significant correlation, both in intact seedlings and in cultured shoots, between the number of buds present per axil and the number of leaf primordia borne on the largest bud (Figs. 1 and 2) (r = 0.98; p < 0.001). Normally one or more nodes were present at which no axillary bud was visible (Figs. 3a and 4) and the minimum mean number of buds per axil fell below unity both in intact
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F W
YO
MIN
G o
n 09
/20/
13Fo
r pe
rson
al u
se o
nly.
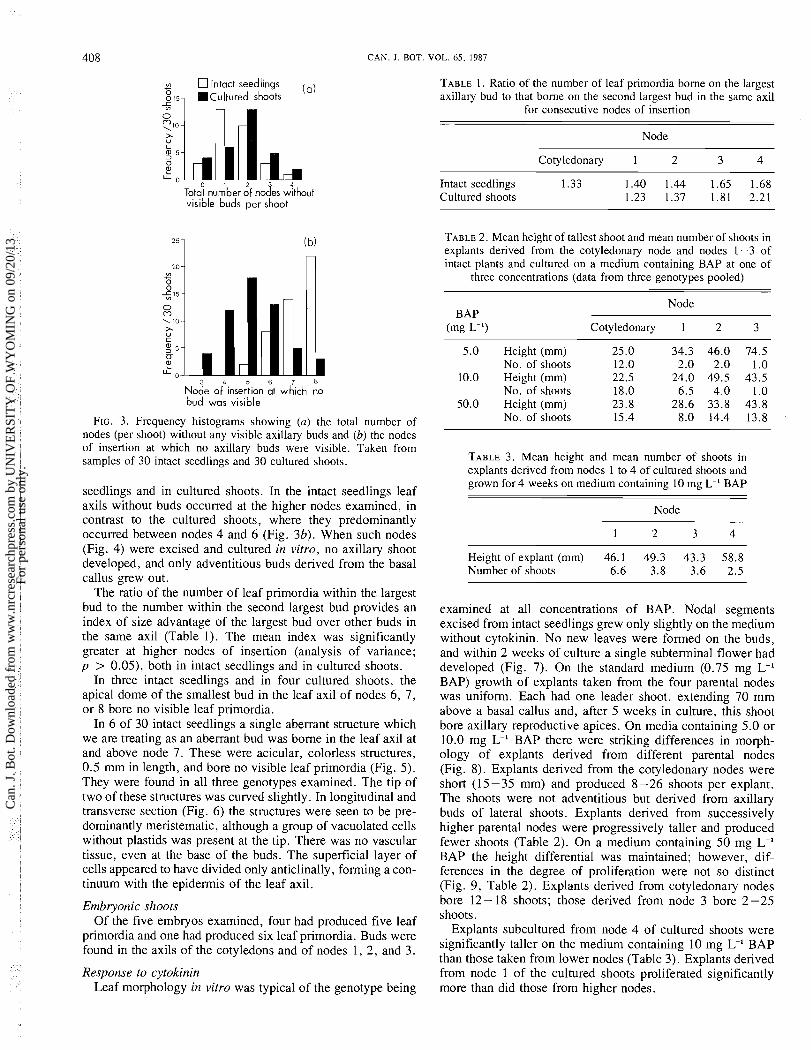
CAN. J. BOT. VOL. 65, 1987
m Intact seedlings - 815, .Cultured shoots
( 0 )
1
u
L L 0 0 1 2 3 4 Total number of nodes without visible buds per shoot
3 4 5 6 7 8
Node of insertion at which no bud was visible
FIG. 3. Frequency histograms showing (a) the total number of nodes (per shoot) without any visible axillary buds and (b) the nodes of insertion at which no axillary buds were visible. Taken from samples of 30 intact seedlings and 30 cultured shoots.
seedlings and in cultured shoots. In the intact seedlings leaf axils without buds occurred at the higher nodes examined, in contrast to the cultured shoots, where they predominantly occurred between nodes 4 and 6 (Fig. 3b). When such nodes (Fig. 4) were excised and cultured in vitro, no axillary shoot developed, and only adventitious buds derived from the basal callus grew out.
The ratio of the number of leaf primordia within the largest bud to the number within the second largest bud provides an index of size advantage of the largest bud over other buds in the same axil (Table 1). The mean index was significantly greater at higher nodes of insertion (analysis of variance; p > 0.05), both in intact seedlings and in cultured shoots.
In three intact seedlings and in four cultured shoots, the apical dome of the smallest bud in the leaf axil of nodes 6 , 7, or 8 bore no visible leaf primordia.
In 6 of 30 intact seedlings a single aberrant structure which we are treating as an aberrant bud was bome in the leaf axil at and above node 7. These were acicular, colorless structures, 0.5 mm in length, and bore no visible leaf primordia (Fig. 5). They were found in all three genotypes examined. The tip of two of these structures was curved slightly. In longitudinal and transverse section (Fig. 6) the structures were seen to be pre- dominantly meristematic, although a group of vacuolated cells without plastids was present at the tip. There was no vascular tissue, even at the base of the buds. The superficial layer of cells appeared to have divided only anticlinally, forming a con- tinuum with the epidermis of the leaf axil.
Embryonic shoots Of the five embryos examined, four had produced five leaf
primordia and one had produced six leaf primordia. Buds were found in the axils of the cotyledons and of nodes 1, 2, and 3.
Response to cytokinin Leaf morphology in vitro was typical of the genotype being
TABLE 1. Ratio of the number of leaf primordia bome on the largest axillary bud to that borne on the second largest bud in the same axil
for consecutive nodes of insertion
Node
Cotyledonary 1 2 3 4
Intact seedlings 1.33 1.40 1.44 1.65 1.68 Cultured shoots - 1.23 1.37 1.81 2.21
TABLE 2. Mean height of tallest shoot and mean number of shoots in explants derived from the cotyledonary node and nodes 1-3 of intact plants and cultured on a medium containing BAP at one of
three concentrations (data from three genotypes pooled)
Node BAP
(mg L-'1 Cotyledonary 1 2 3
5.0 Height (mm) 25.0 34.3 46.0 74.5 No. of shoots 12.0 2.0 2.0 1.0
10.0 Height (mm) 22.5 24.0 49.5 43.5 No. of shoots 18.0 6.5 4.0 1.0
50.0 Height (mm) 23.8 28.6 33.8 43.8 No. of shoots 15.4 8.0 14.4 13.8
TABLE 3. Mean height and mean number of shoots in explants derived from nodes 1 to 4 of cultured shoots and grown for 4 weeks on medium containing 10 mg L-' BAP
Node
Height of explant (mm) 46.1 49.3 43.3 58.8 Number of shoots 6.6 3.8 3.6 2.5
examined at all concentrations of BAP. Nodal segments excised from intact seedlings grew only slightly on the medium without cytokinin. No new leaves were formed on the buds, and within 2 weeks of culture a single subterminal flower had developed (Fig. 7). On the standard medium (0.75 mg L-' BAP) growth of explants taken from the four parental nodes was uniform. Each had one leader shoot, extending 70 mm above a basal callus and, after 5 weeks in culture, this shoot bore axillary reproductive apices. On media containing 5.0 or 10.0 mg L-' BAP there were striking differences in morph- ology of explants derived from different parental nodes (Fig. 8). Explants derived from the cotyledonary nodes were short (15-35 mm) and produced 8-26 shoots per explant. The shoots were not adventitious but derived from axillary buds of lateral shoots. Explants derived from successively higher parental nodes were progressively taller and produced fewer shoots (Table 2). On a medium containing 50 mg L-I BAP the height differential was maintained; however, dif- ferences in the degree of proliferation were not so distinct (Fig. 9, Table 2). Explants derived from cotyledonary nodes bore 12 - 18 shoots; those derived from node 3 bore 2 - 25 shoots.
Explants subcultured from node 4 of cultured shoots were significantly taller on the medium containing 10 mg L-I BAP than those taken from lower nodes (Table 3). Explants derived from node 1 of the cultured shoots proliferated significantly more than did those from higher nodes.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F W
YO
MIN
G o
n 09
/20/
13Fo
r pe
rson
al u
se o
nly.
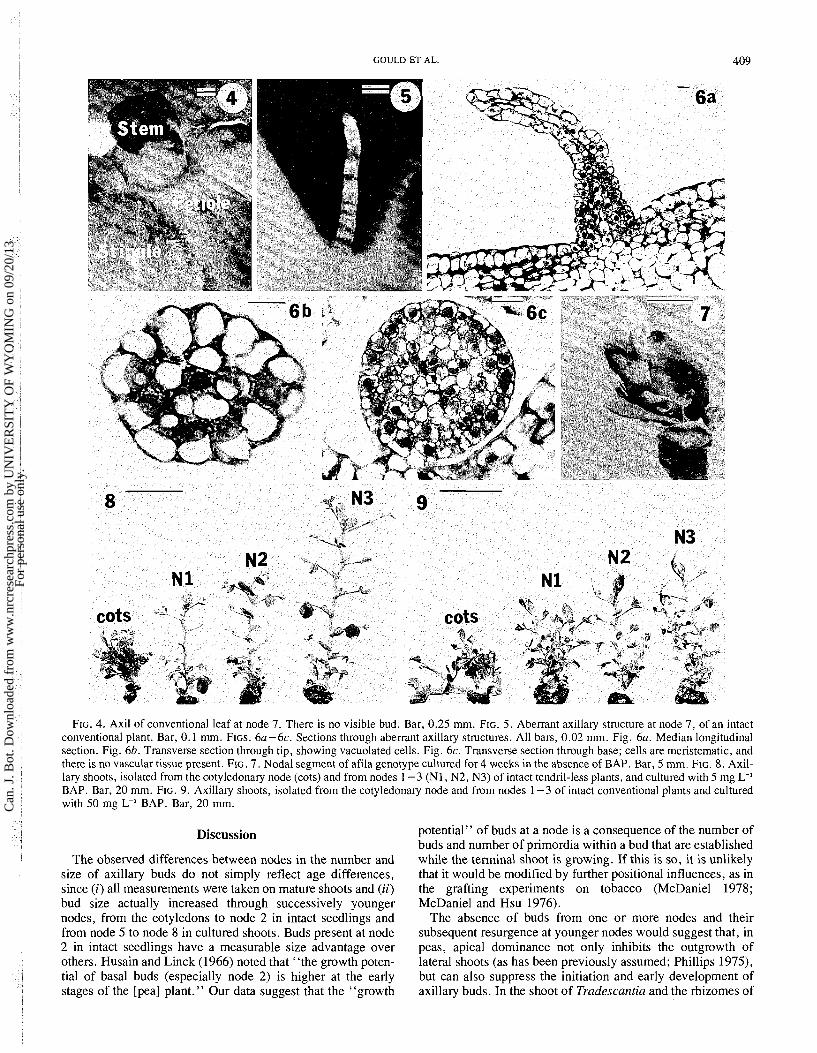
GOULD ET AL. 409
FIG. 4. Axil of conventional leaf at node 7. There is no visible bud. Bar, 0.25 mm. FIG. 5. Aberrant axillary structure at node 7, of an intact conventional plant. Bar, 0.1 mm. FIGS. 6a-6c. Sections through aberrant axillary structures. All bars, 0.02 mm. Fig. 6a. Median longitudinal section. Fig. 6b. Transverse section through tip, showing vacuolated cells. Fig. 6c. Transverse section through base; cells are meristematic, and there is no vascular tissue present. FIG. 7. Nodal segment of afila genotype cultured for 4 weeks in the absence of BAP. Bar, 5 mm. FIG. 8. Axil- lary shoots, isolated from the cotyledonary node (cots) and from nodes 1 -3 (N1 , N2, N3) of intact tendril-less plants, and cultured with 5 mg L-' BAP. Bar, 20 mm. FIG. 9. Axillary shoots, isolated from the cotyledonary node and from nodes 1-3 of intact conventional plants and cultured with 50 mg L-' BAP. Bar, 20 mm.
Discussion
The observed differences between nodes in the number and size of axillary buds do not simply reflect age differences, since (i) all measurements were taken on mature shoots and (ii) bud size actually increased through successively younger nodes, from the cotyledons to node 2 in intact seedlings and from node 5 to node 8 in cultured shoots. Buds present at node 2 in intact seedlings have a measurable size advantage over others. Husain and Linck (1966) noted that "the growth poten- tial of basal buds (especially node 2) is higher at the early stages of the [pea] plant." Our data suggest that the "growth
potential" of buds at a node is a consequence of the number of buds and number of primordia within a bud that are established while the terminal shoot is growing. If this is so, it is unlikely that it would be modified by further positional influences, as in the grafting experiments on tobacco (McDaniel 1978; McDaniel and Hsu 1976).
The absence of buds from one or more nodes and their subsequent resurgence at younger nodes would suggest that, in peas, apical dominance not only inhibits the outgrowth of lateral shoots (as has been previously assumed; Phillips 1975), but can also suppress the initiation and early development of axillary buds. In the shoot of Tradescantia and the rhizomes of
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F W
YO
MIN
G o
n 09
/20/
13Fo
r pe
rson
al u
se o
nly.

410 CAN. J . BOT. VOL. 65, 1987
Matteuccia struthiopteris and Onoclea sensibilis the develop- ment of lateral buds is arrested at a very early stage (Booker and Dwivedi 1972; Naylor 1958; Wardlaw 1943a, 1943b); it appears that in peas the initiation of lateral buds may be sup- pressed completely.
The stage at which the ontogeny of an axillary bud is cor- relatively arrested may be a function of the developmental stage and the physiological activity of the terminal shoot apex. Buds borne at the cotyledonary nodes and at nodes 1 to 3 are formed within the developing seed, at which stage correlative inhibition might well be minimal. Successive axillary buds formed within the seed would be larger as a result of the increase in size and organization of the embryonic shoot. Development of buds borne at node 3 would be arrested with the onset of seed dormancy. Upon germination and the emergence of the plumule, apical dominance would be rapidly established, the correlative inhibition (levels of auxin?) increasing with age of the plant and resulting in progressively fewer leaf primordia developing on the buds, fewer buds being initiated, and finally the absence of buds from leaf axils.
We can offer two explanations for the absence of an initial rise in the size and number of buds along cultured shoots. Firstly, the second-order buds borne on cultured shoots have not experienced the internal environment of the developing seed. Secondly, size and number of buds may relate to the morphology of subtending leaves, since in both intact seed- lings and cultured shoots bud size was maximal within the axil of the scale leaf which preceded the first compound leaf (cul- tured shoots produce only one scale leaf, which may equate with node 2 of intact seedlings). There was no indication of any subsequent dependence on the morphology of subtending leaves, since leaf morphology invariably increases in complex- ity through successively higher nodes (Gould et al. 1986) yet bud size decreases above node 2 and then increases above node 5 in cultured shoots.
Correlative inhibition declines with the maturation of the terminal pea shoot (Husain and Linck 1966). According to our hypothesis, any decline in apical dominance would lead to a resurgence of axillary buds at the highest nodes. The fact that such a resurgence was observed more frequently in cultured shoots than in intact plants probably reflects the shortened life cycle of cultured shoots. when plants were harvested, all cul- tured shoots bore reproductive apices, whereas growth of intact plants remained vegetative.
It is difficult to be certain of the identity of the aberrant structures. By virtue of their position in the leaf axil they may be modified buds, although they do not exhibit a cellular organization typical of angiosperm shoot apices, and no inter- mediate types were observed. They are derived from more than the superficial layer of cells and are not,'therefore, trichomes. The curved tip of some may indicate that they are potential tendrils; axillary buds modified as tendrils are common among Cucurbitaceae (Goebel 1905), although not in legumes. There are some similarities between the anatomy of the aberrant structures and the early development of buds as thorns in Ulex europaeus (Bieniek and Millington 1967). However, since peas have been widely studied and since, to our knowledge, these aberrant structures have not been reported previously, it is probable that their developmental i n intact remains so limited that they never emerge from the leaf axil. It would be interesting to isolate and culture the aberrant struc- tures in vitro.
The size advantage of the largest bud over adjacent buds in
the axil of a single leaf, which increases at higher nodes of insertion (Table I), reflects potential growth advantage of the largest bud in cultured nodes (Fig. 9). Moreover, the greater the size advantage of the largest bud, the higher was the threshold level of cytokinin required to induce growth of the smaller adjacent buds. Direct cytokinin applications have been shown to release lateral buds from correlative inhibition in intact pea seedlings (Sachs and Thimann 1964). One likely explanation of our results is that the largest bud inhibits the growth of smaller adjacent buds. Inclusion of 0.75 mg L-I BAP in our nutrient medium, which did not release any of the smaller buds of a leaf axil from inhibition, resulted in uniform growth of the largest bud derived from all four nodes examined. With 5.0 and 10.0 mg L-' BAP the dominance exerted by larger buds from the cotyledonary node (and to a smaller extent from node 1) was broken; all buds present in these nodal segments grew out and themselves proliferated (Table 2). Dominance of the largest bud in explants derived from higher nodes was not broken, and nor&ally a single leader shoot grew out. At the high cytokinin level (50.0 mg L-' BAP) dominance of the largest bud over adjacent buds was broken in explants derived from all nodes, and all resultant shoots proliferated.
Sorokin and Thimann (1964) observed up to two vascular traces leading to each of three inhibited buds in the axil of the scale leaf at node 2 in cv. Alaska, with a branch trace extend- ing from the bud into the primary tissues of the stele. Their results imply that the multiple buds occumng within a leaf axil in Pisum are derived independently. Dominance of one inde- pendent bud over another at the same node, although not in the same leaf axil, has been demonstrated in Hygrophila poly- sperma and in Alternanthera philoxeroides (Cutter 1967, 1972), both of which are decussate species showing aniso- clady. Grafting experiments on cultured nodes of Hygrophila led to the conclusion that the larger of each pair of buds exerted dominance over the smaller by hormonal, rather than nutri- tional, means (Cutter and Chiu 1972). We propose that in Pisum, the largest axillary bud of a group exerts dominance over adjacent buds by constituting a preferential sink for available cytokinins. Only when levels of cytokinins exceed the requirement of the largest bud are smaller buds released from correlative inhibition.
It might well be advantageous for the commercial grower interested in producing maximal numbers of shoots from isolated nodes in culture to perform an initial screening for positional differences in the response to in vitro conditions. Efforts could then be concentrated on those nodes with maxi- mal proliferation.
Acknowledgements We thank Dr. G. Hussey for demonstrating the in vitro
technique and Mr. I. W. Miller A.B.I.P.P., for assistance with the photography. Work was supported by a Science and Engineering Research Council Cooperative Award in Science and Engineering to K. S. G.
BIENIEK, M. E., and W. F. MILLINGTON. 1967. Differentiation of lateral shoots as thorns in Ulex europaeus. Am. J. Bot. 54: 61 -70.
BOOKER, C. E., and R. S. DWIVEDI. 1972. Ultrastructure of meri- stematic cells of dormant and released buds of Tradescantia palu- dosa. Exp. Cell Res. 82: 255-261.
CUTTER, E. G. 1967. Morphogenesis and developmental potential- ities of unequal buds. Phytomorphology , 17: 437 -445.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F W
YO
MIN
G o
n 09
/20/
13Fo
r pe
rson
al u
se o
nly.

GOULD ET AL. 411
1972. Regulation of branching in decussate species with unequal lateral buds. Ann. Bot. (London), 36: 207-220.
CUTTER, E. G., and H. CHIU. 1972. Grafting experiments on correla- tive effects between lateral buds. Ann. Bot. (London), 36: 221 -228.
1975. Differential responses of buds along the shoot to factors involved in apical dominance. J. Exp. Bot. 26: 828-839.
FEDER, N., and T. P. O'BRIEN. 1968. Plant microtechnique: some principles and new methods. Am. J . Bot. 55: 123 - 142.
GLAUERT, A. M. 1975. Fixation, dehydration and embedding of biological specimens. Practical methods in electron microscopy. North Holland Publishing Co., Amsterdam.
GOEBEL, K. 1905. Organography of plants. Part I . Translated by I . B. Balfour. Clarendon Press, Oxford.
GOLDENBERG, J. B. 1965. "Afila" a new mutation in pea (Pisurn sativum L). Bol. Genet. 1: 27-3 1.
COULD, K. S., E. G. CUTTER, and J. P. W. YOUNG. 1986. Morphogenesis of the compound leaf in three genotypes of the pea, Pisum sativum. Can. J. Bot. 64: 1268 - 1276.
HUSAIN, S. M., and A. J . LINCK. 1966. Relationship of apical dominance to the nutrient accumulation pattern in Pisum sativum var. Alaska. Physiol. Plant. 19: 992 - 1010.
HUSSEY, G. 1978. The application of tissue culture to the vegetative propagation of plants. Sci. Prog. (London), 65: 185 -208.
HUSSEY, G., and H. V. GUNN. 1984. Plant production in pea (Pisurn sativum L. cvs. Puget and Upton) from long-term callus with superficial meristems. Plant Sci. Lett. 37: 143 - 148.
KUJALA, V. 1953. Felderbse, bei welcher die ganze Blattspreite in Ranken umgewandelt ist. Arch. Soc. Zool. Bot. Fenn. Vanamo, 8: 44-45.
MCDANIEL, C. 1978. Determination for growth pattern in axillary buds of Nicotiana tabacum L. Dev. Biol. 66: 250-255.
1984. Competence, determination, and induction in plant
development. 111 Pattern formation. Edited by G. M. Malacinski and S. V. Bryant. Macmillan, New York. pp. 393-412.
MCDANIEL, C., and F. C. HSU. 1976. Position-dependent develop- ment of tobacco meristems. Science (N.Y.), 259: 564-565.
MURASHIGE, T., and F. SKOOG. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15: 473-497.
NAYLOR, J . M. 1958. Control of nuclear processes by auxin in axil- lary buds of Tradescantia paludosa. Can. J. Bot. 36: 221 -232.
PHILLIPS, I. D. J . 1975. Apical dominance. Annu. Rev. Plant Physiol. 26: 341-367.
SACHS, T., and K. V. THIMANN. 1964. Release of lateral buds from apical dominance. Nature (London), 201: 939-940.
SOROKIN, H. P., and K. V. THIMANN. 1964. The histological basis for inhibition of axillary buds in Pisurn sativurn and the effects of auxin and kinetin on xylem development. Protoplasma, 59: 326 - 349.
VILMORIN, P. DE, and W. BATESON. 1912. A case of gametic coupling in peas. Proc. R. Soc. London Ser. B, 84: 9- 11.
WARDLAW, C. W. 1943a. Experimental and analytical studies of pteridophytes. I . Preliminary observations on the development of buds on the rhizome of the ostrich fern (Matteucia strrtthiopteris Tod.). Ann. Bot. (London), 7: 171 - 184.
1943b. Experimental and analytical studies of pteridophytes. 11. Experimental observations on the development of buds in Onoclea sensibilis and in species of Dryopteris. Ann. Bot. (London), 7: 357 -377.
WHITE, 0 . 1917. Studies of inheritance in Pisurn. 11. The present state of knowledge of heredity and variation in peas. Proc. Am. Philos. Soc. 56: 487-588.
ZIV, M., A. H. HALVEY, and R. SHILO. 1970. Organs and plantlets regeneration of Gladiolus through tissue culture. Ann. Bot. (London), 34: 67 1 - 676.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F W
YO
MIN
G o
n 09
/20/
13Fo
r pe
rson
al u
se o
nly.





























