Embed Size (px)
Citation preview

Biochimica et Biophysica Acta, 320 (1973) 635-647 Elsevier Scientific Publishing Company, Amsterdam - Printed in The Netherlands
BBA 27252
PREPARATION AND PROPERTIES OF 2'(or Y)-O-(2,4,6-TRINITROPHE-
NYL) ADENOSINE 5 ' -TRIPHOSPHATE, AN A N A L O G OF ADENOSINE
TRIPHOSPHATE
T. HIRATSUKA and K. UCHIDA
Department of Chemistry, Faculty of Science, Hokkaido University, Sapporo (Japan)
(Received May 21st, 1973)
SUMMARY
An analog of adenosine triphosphate, 2'(or Y)-O-(2,4,6-trinitrophenyl)ade- nosine 5'-triphosphate (TNP-ATP), was synthesized as a reporter-labeled substrate of heavy meromyosin ATPase. TNP-ATP was hydrolyzed by heavy meromyosin in the presence of CaC12 MgC12 or EDTA.
TNP-ATP had absorption maxima at 259 nm, 408 nm and 470 nm at neutral pH. When bound to heavy meromyosin, TNP-ATP underwent the characteristic spectral shift. The difference spectrum resulting from the binding of TNP-ATP to heavy meromyosin at pH 8.0 had positive peaks at 415 nm and 518 nm, and a nega- tive trough at 458 nm.
The difference spectrum due to the binding of 2'(or 3')-O-(2,4,6-trinitrophe- nyl)adenosine (TNP-adenosine) to heavy meromyosin had small positive peaks at 420 nm and 495 nm. This difference spectrum was similar to that of TNP-ATP or TNP-adenosine produced by 20 % (v/v) ethyleneglycol perturbation. The positive peak at 495 nm in the difference spectrum due to the binding of TNP-adenosine to heavy meromyosin shifted toward 505 nm, when pyrophosphate or ATP was added to the reaction mixture.
These results suggest that the difference spectrum of TNP-ATP due to the interaction with heavy meromyosin arises not only from the binding of the chromo- phoric portion of the TNP-ATP molecule but also from that of the phosphate portion.
INTRODUCTION
A number of analogs of ATP have been synthesized to elucidate the roles of adenine base, ribose and triphosphate 1 and the role of adenosine residue 2'3 in the
Abbreviations: TNBS, 2,4,6-trinitrobenzene-l-sulfonate; TNP-ATP, 2'(or 3")-O-(2,4,6-trini- trophenyl)adenosine 5'-triphosphate including 2',3'-O-(2,4,6-trinitrocyclohexadienylidene)adenosine 5"-triphosphate at neutral and basic pH6; TNP-ADP, 2' (or 3')-O-(2,4,6-trinitrophenyl) adenosine 5'-diphosphate including 2',3'-O-(2,4,6-trinitrocyclohexadienylidene)adenosine 5"-diphosphate; TNP- adenosine, 2'(or 3')-O-(2,4,6-trinitrophenyl)adenosine including 2',3'-O-(2,4,6-trinitrocyclohexadi- enylidene)adenosine; TNP-, 2,4,6-trinitrophenyl.

636 T. H I R A T S U K A , K. U C H I D A
enzymic hydrolysis of ATP by myosin ATPase and other ATP-relating enzymes. Recently, the analog of ATP, which has a chromophore 4 or fluorophore 5, has also been synthesized to clarify the structure around the active site of myosin ATPase. To provide information about the microenvironment within the binding site, a rea- gent with an environmentally sensitive group for a particular binding site is designed by the use of the specific binding characteristics of heavy meromyosin ATPase.
N
H H H Adenine
- o - o - o - I
' ~-----4 ' -I-H* ~. I I
-14" q~" 'gH
Fig. I. Structure proposed for TNP-ATP at acidic and basic pH.
Azegami and Iwai 6 have reported that the ribose residue of adenosine is tri- nitrophenylated by 2,4,6-trinitrobenzene-l-sulfonate (TNBS). In the present paper, the ribose residue of ATP was also trinitrophenylated under the mild condition. 2'(or Y)-O-(2,4,6-Trinitrophenyl)adenosine 5'-triphosphate (TNP-ATP) resulting from the reaction of ATP with TNBS has visible absorption characterisitic of the typical Meisenheimer compound linking 7'8, among 2' and Y-OH groups of ribose and trinitrobenzene at neutral pH (Fig. 1). The TNP-ATP obtained is useful as a reporter-labeled substrate for heavy meromyosin ATPase. This compound is not a native substrate, but it interacts with heavy meromyosin and is then hydrolyzed.
MATERIALS A N D METHODS
Chemicals Reagent grade chemicals were used throughout. ATP was obtained from
Sigma Chemical Co. or Kyowa Hakko Co. Adenosine was obtained from Sanko Junyaku Co. Sodium 2,4,6-trinitrobenzene-l-sulfonate was purchased from Tokyo Kasei Kogyo Co. and was used without further purification. Deuterium oxide (99.8 ~ ) was purchased from Merck (Darmstadt).
Enzyme Heavy meromyosin was prepared from myosin by digestion with trypsin for
10 min at 23 °C as described previously I o, 11. For spectral measurements, heavy me- romyosin was purified by eluting from DEAE-cellulose according to a similar method to that of Mueller and Perry 12. Protein concentration was determined from the mea-

TRINITROPHENYL-ADENOSINE TRIPHOSPHATE 637
surement of the absorbance at 280 nm, assuming that a solution containing 1.0 mg of protein per ml has an absorbance of 0.63. The molecular weight of heavy meromy- osin was taken as 3.5 • 105 (ref. 13).
Preparation of TNP-adenosine Direct trinitrophenylation of adenosine with TNBS was accomplished by the
method of Azegami and Iwai 6.
Preparation of TNP-ATP ATP (1.0 g, disodium salt, 1.65 mmoles) was dissolved in 10 ml of water and
the pH was adjusted to 9.5 with 4 M LiOH. To this solution with continuous stirring TNBS (1.7 g, 5.4 mmoles) in 10 ml of water was added dropwise over 3 h at 30 °C. The pH was immediately adjusted to and maintained at 9.5 by titration with 0.4 M LiOH. The stirring was further continued for 4 days to ensure the reaction at room temperature in the dark. The reaction mixture was then evaporated to dryness at 30 °C. The residues obtained were suspended in 40 ml of acetone-methanol (3 : 1, v/v), and then filtered. The residues on a sintered glass were repeatedly washed with acetone.
The reddish orange residues were dissolved in 10 ml of water and purified on a Sephadex LH-20 column packed in water.
Paper chromatography Ascending paper chromatography on Whatman No. 2 paper was carried out
in the following solvent systems; (A) isobutyric acid-1 M ammonium hydroxide- 0.I M ethylenediamine tetraacetic acid (100 : 60 : 1.6, v/v/v), (B) n-butanol-acetic acid-water ( 4 0 : 1 0 : 2 5 , v/v/v) and (C) e thanol-I M ammonium acetate ( 7 : 3 , pH 7.5, v/v). The ultraviolet-absorbing materials were located on paper by obser- vation under an ultraviolet lamp (2536 A), extracted with water and measured for ultraviolet absorption.
Enzyme assay ATPase activity of heavy meromyosin was measured in a solution containing,
if not otherwise stated, 0.1 mg of heavy meromyosin per ml, 0.27 mM ATP, 0.5 M KC1, 50 mM Tris-maleate buffer (pH 7.0) at 25 °C and, alternatively, 5 mM CaCI 2, 5 mM EDTA or 10 mM MgC1 z. Only when the reaction was performed in the pre- sence of MgClz, 2.6 mg of heavy meromyosin per ml was used.
Measurements using TNP-ATP as a substrate were also carried out under the same conditions as those of ATPase assay. Incubation was terminated by addition of 1 ml of 10 ~o trichloroacetic acid. Pi liberated was determined by the method of Fiske-SubbaRow 14 or Mar t in-Doty 15.
Acid hydrolysis For complete hydrolysis, TNP-adenosine and TNP-ATP were incubated in
1 M HC1 at 100 °C for 1.5 h. Picric acid in the hydrolyzates was directly determined f rom the absorbance at 356 nm in 0.5 M N a O H using the extinction coefficient of 14200 M -1 • cm -1 (ref. 16).
For determination of acid-labile phosphate, TNP ATP and ATP were hydrol- yzed in I M HC1 at 100 °C for 7 min 17. The acid-labile phosphate liberated was

638 T. HIRATSUKA, K. UCHIDA
determined by the method of Fiske-SubbaRow ~4. Aliquots of hydrolyzates obtained in these experiments were also subjected
to paper chromatography for further characterization. Adenine fractionated by paper chromatography and extracted with water was determined from the absorbance at 269 nm in 0.01 M NaOH using the extinction coefficient of 12300M - 1 . cm -~ (ref. 18).
Spectral measurements Proton magnetic resonance experiments were carried out in 2H20 with a
JEOL PS-100 high resolution N M R spectrometer operating at 100 MHz. Chemical shifts were measured with respect to an external reference of tetramethylsilane. As- signments were made by references for ATP 19 and trinitrocyclohexadienate anion % 20.
Infrared spectra of TNP-ATP and ATP were obtained with a JASCO I R - G infrared spectrophotometer. A KBr pellet was prepared from a mixture obtained by grinding 700 mg of oven-dried spectrograde KBr with 3 mg of the compound. The instrument was calibrated against polystyrene film.
Absorption spectra, difference spectra and solvent perturbation spectra were obtained at room temperature by means of a Hitachi model 124 spectrophotometer. The measurements of difference spectra were made as described previously 21. Spectral scans were always started after incubation of the reaction mixture at room tempera- ture for 10 rain. In all the spectral measurements, the pH of the reaction mixture was checked by a glass electrode after each spectrophotometric measurement. No sig- nificant change of the desired pH was found.
Concentrations of TNP-adenosine and TNP-ATP were determined by measu- rements of the absorbance at 408 nm at pH 8.0 with use of the extinction coef- ficient of 26400 M-1 • cm-~ in the following experiments.
RESULTS
Purification and characterization of TNP-adenosine The products were applied on a Sephadex LH-20 column (2.2 cm × 25 cm)
packed in water and TNP-adenosine was eluted with ethanol. The preparation was reproducible and the yield of TNP-adenosine after purification was 48 ~o. Upon paper chromatography, TNP-adenosine purified gave a single orange spot (Table I).
Aliquots of the effluent containing TNP-adenosine were pooled and evaporat- ed to dryness under the reduced pressure.
Analysis calculated for C16H 13NSNaO10 • 1.9 H20 (534.544): C, 35.95; H, 3.17; N, 20.96. Found: C, 36.52; H, 3.38; N, 20.50.
The infrared spectrum of TNP-adenosine agreed well with those reported by Azegami and Iwai 6 and the spectra reported by Dyal111 and Foster et al. 7 for Meisen- heimer complexes.
Azegami and Iwai 6 reported that O-TNP-nucleosides are hydrolyzed by treat- ment with 1 M HCI at 100 °C to give an equimolar amount of picric acid and base. The TNP-adenosine prepared was also completely hydrolyzed for 1.5 h under the same conditions. The stoichiometric relation between adenine and picric acid in the hydrolyzate of TNP-adenosine agrees well with that reported by Azegami and Iwai 6 (Table 1).
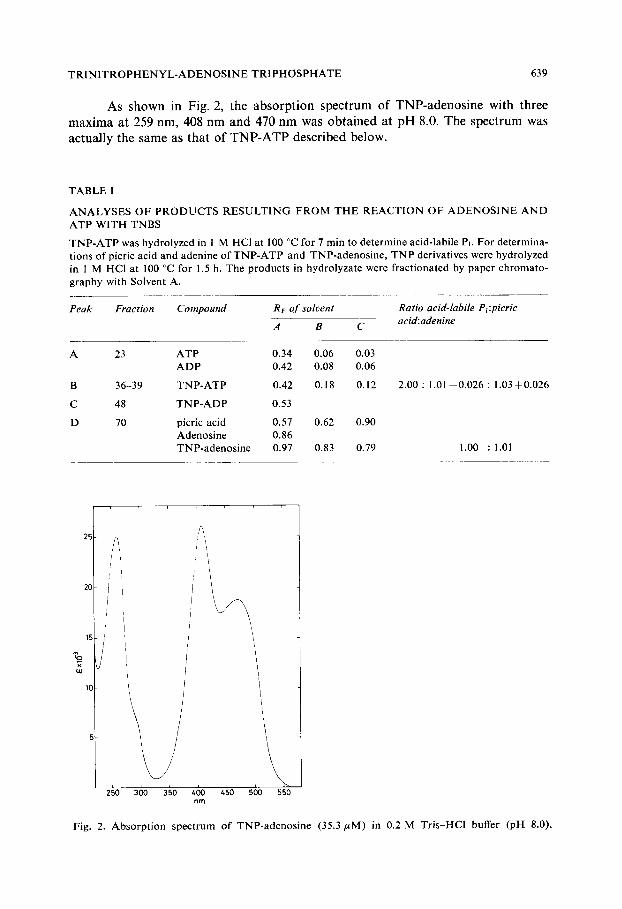
T R I N I T R O P H E N Y L - A D E N O S I N E T R I P H O S P H A T E 639
As shown in Fig. 2, the absorption spectrum of TNP-adenosine with three maxima at 259 nm, 408 nm and 470 nm was obtained at pH 8.0. The spectrum was actually the same as that of TNP-ATP described below.
T A B L E I
A N A L Y S E S O F P R O D U C T S R E S U L T I N G F R O M T H E R E A C T I O N OF A D E N O S I N E A N D A T P W I T H TNBS
T N P - A T P was hydrolyzed in 1 M HC1 at 100 °C for 7 min to determine acid-labile PI. For determina- t ions o f picric acid and adenine o f T N P - A T P and TNP-adenos ine , T N P derivatives were hydrolyzed in 1 M HCI at 100 °C for 1.5 h. The p roduc t s in hydrolyzate were f ract ionated by paper ch romato - g raphy with Solvent A.
Peak Fraction Compound Rv of solvent Ratio acid-labile P+:picric acid:adenine A B C
A 23 A T P 0.34 0.06 0.03 A D P 0.42 0.08 0.06
B 36-39 T N P - A T P 0.42 0.18 0.12
C 48 T N P - A D P 0.53
D 70 picric acid 0.57 0.62 0.90 Adenos ine 0.86 TNP-adenos ine 0.97 0.83 0.79
2.00 : 1.01 ±0 .026 : 1 .0350.026
1.00 : 1.01
w
25 /,~
,/ ,I i ,!
20 /
2~o
! '
J //]
i
36o 3~o 46o 4;o 50o ~so nm
Fig. 2. Absorp t ion spec t rum o f TNP-adenos ine (35.3/~M) in 0.2 M Tris-HC1 buffer (pH 8.0).
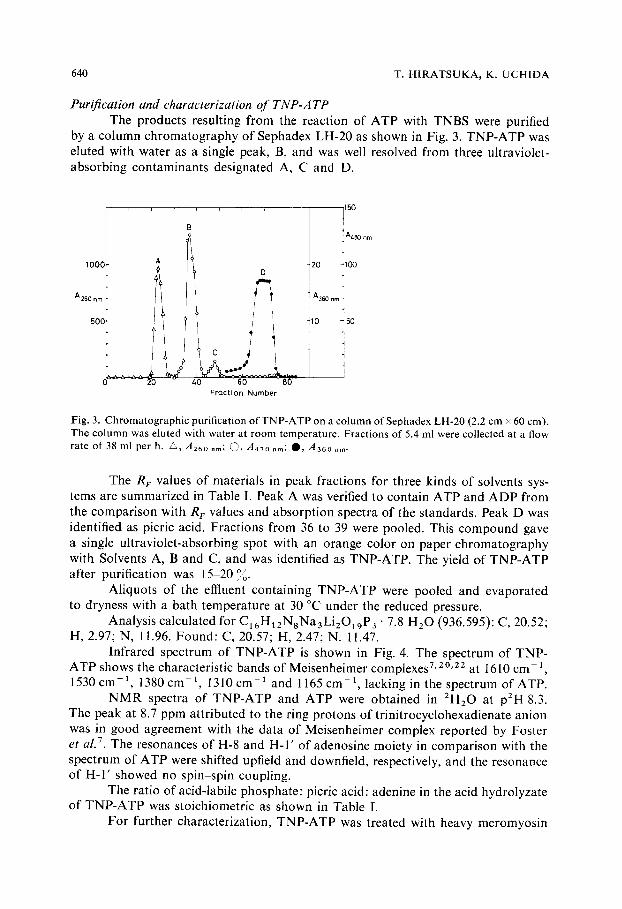
640 T. HIRATSUKA, K. UCHIDA
Purification and characterization o f T N P - A T P The products resulting from the reaction of ATP with TNBS were purified
by a column chromatography of Sephadex LH-20 as shown in Fig. 3. TNP-ATP was eluted with water as a single peak, B, and was well resolved from three ultraviolet- absorbing contaminants designated A, C and D.
1000
A26O nrn
500
A
I i
C ' i
2O 4 0 ~ Fraction Number
150
I A480 nm
2 0 1100
A360 r'm t
10 -~50
Fig. 3. Chromatographic purification of TNP-ATP on a column ofSephadex LH-20 (2.2 cm × 60 cm). The column was eluted with water at room temperature. Fractions of 5.4 ml were collected at a flow rate of 38 ml per h. •, A260 nm; C~, A470 rim; O , A36o nm.
The R F values of materials in peak fractions for three kinds of solvents sys- tems are summarized in Table I. Peak A was verified to contain ATP and ADP from the comparison with R r values and absorption spectra of the standards. Peak D was identified as picric acid. Fractions from 36 to 39 were pooled. This compound gave a single ultraviolet-absorbing spot with an orange color on paper chromatography with Solvents A, B and C, and was identified as TNP-ATP. The yield of TNP-ATP after purification was 15-20')o.
Aliquots of the effluent containing TNP-ATP were pooled and evaporated to dryness with a bath temperature at 30 °C under the reduced pressure.
Analysis calculated for C16HI2NsNa3Li2019P 3 • 7.8 H20 (936.595): C, 20.52; H, 2.97; N, 11.96. Found: C, 20.57; H, 2.47; N. 11.47.
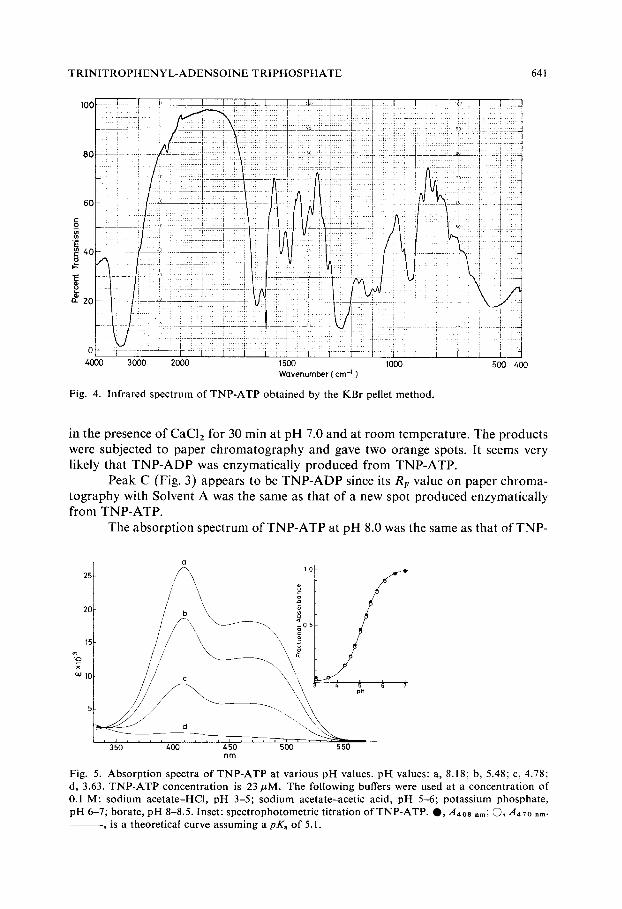
Infrared spectrum of TNP-ATP is shown in Fig. 4. The spectrum of TNP- ATP shows the characteristic bands of Meisenheimer complexes 7,2°'2z at 1610 cm-1, 1530 cm- 1, 1380 cm- 1, 1310 cm- ~ and 1165 cm- l, lacking in the spectrum of ATP.
N M R spectra of TNP-ATP and ATP were obtained in 2HzO at p2H 8.3. The peak at 8.7 ppm attributed to the ring protons of trinitrocyclohexadienate anion was in good agreement with the data of Meisenheimer complex reported by Foster et al. 7. The resonances of H-8 and H-I ' of adenosine moiety in comparison with the spectrum of ATP were shifted upfield and downfield, respectively, and the resonance of H- I ' showed no spin-spin coupling.
The ratio of acid-labile phosphate: picric acid: adenine in the acid hydrolyzate of TNP-ATP was stoichiometric as shown in Table I.
For further characterization, TNP-ATP was treated with heavy meromyosin
T R I N I T R O P H E N Y L - A D E N S O I N E TRIPHOSPHATE
100
641
80
6O
c ~ 40
2o
0 4000 3000 2000 1500 1000
Wavenumber ( cm -~ )
Fig. 4. Infrared spectrum of TNP-ATP obtained by the KBr pellet method.
500 400
in the p resence o f C a C l z fo r 30 min at p H 7.0 a n d at r o o m t e m p e r a t u r e . The p r o d u c t s
were sub jec ted to pape r c h r o m a t o g r a p h y a n d gave two o r a n g e spots. I t seems very
l ikely tha t T N P - A D P was e n z y m a t i c a l l y p r o d u c e d f r o m T N P - A T P .
P e a k C (Fig. 3) appea r s to be T N P - A D P since its R F va lue on pape r c h r o m a -
t o g r a p h y wi th So lven t A was the s a m e as tha t o f a new spot p r o d u c e d e n z y m a t i c a l l y f r o m T N P - A T P .
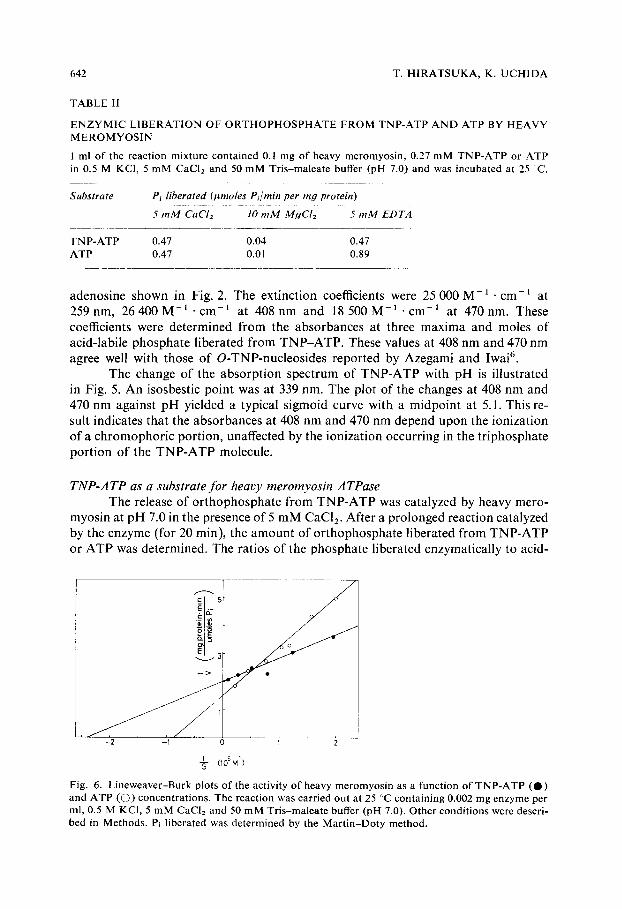
T h e a b s o r p t i o n s p e c t r u m o f T N P - A T P at p H 8.0 was the s a m e as tha t o f T N P -
(1 1 2 5 / ~ S
jl . . . . \ /
~ d " ' ~ 35o 4oo 4so 5oo s~o nm
Fig. 5. Absorpt ion spectra o f TNP-ATP at various p[-[ values, p]-[ values: a, 8.18; b, 5.48; c, 4.78; d, 3.63. TNP-ATP concentration is 23 #M. The following buffers were used at a concentration of 0.1 M: sodium acetate-HCl, pH 3-5; sodium acetate-acetic acid, pH 5-6; potassium phosphate, pH 6-7; borate, pH 8-8.5. Inset: spectrophotometric titration of TNP-ATP. O, A4oa nm; (3, , 4470 nm.
, is a theoretical curve assuming a pKa of 5.1.
642 T. H I R A T S U K A , K. U C H I D A
TABLE II
E N Z Y M I C L I B E R A T I O N OF O R T H O P H O S P H A T E FROM TNP-ATP A N D ATP BY HEAVY M E R O M Y O S I N
1 ml of the reaction mixture contained 0.1 mg of heavy meromyosin, 0.27 mM TNP-ATP or ATP in 0.5 M KCI, 5 mM CaC12 and 50 mM Tris-maleate buffer (pH 7.0) and was incubated at 25 C .
Substrate Pt liberated (#moles P~/min per my protein)
5 mM CaClz 10 mM MgCI2 5 mM EDTA
TNP-ATP 0.47 0.04 0.47 ATP 0.47 0.01 0.89
adenosine shown in Fig. 2. The extinction coefficients were 25 000 M - 1 . cm-~ at 259 nm, 26 400 M - I . cm-1 at 408 nm and 18 500 M - J . c m - I at 470 nm. These coefficients were determined from the absorbances at three maxima and moles of acid-labile phosphate liberated from TNP-ATP. These values at 408 nm and 470 nm agree well with those of O-TNP-nucleosides reported by Azegami and Iwai 6.
The change of the absorption spectrum of TNP-ATP with pH is illustrated in Fig. 5. An isosbestic point was at 339 nm. The plot of the changes at 408 nm and 470 nm against pH yielded a typical sigmoid curve with a midpoint at 5.1. This re- sult indicates that the absorbances at 408 nm and 470 nm depend upon the ionization of a chromophoric portion, unaffected by the ionization occurring in the triphosphate portion of the TNP-ATP molecule.
TNP-ATP as a substrate for heavy meromyosin ATPase The release of orthophosphate from TNP-ATP was catalyzed by heavy mero-
myosin at pH 7.0 in the presence of 5 mM CaC12. After a prolonged reaction catalyzed by the enzyme (for 20 rain), the amount of orthophosphate liberated from TNP-ATP or ATP was determined. The ratios of the phosphate liberated enzymatically to acid-
-2 -I
1 (10 5b~I)
1 2
Fig. 6. Lineweaver-Burk plots of the activity of heavy meromyosin as a function of TNP-ATP (@) and ATP (©) concentrations. The reaction was carried out at 25 °C containing 0.002 mg enzyme per ml, 0.5 M KCI, 5 mM CaCI2 and 50 mM Tris-maleate buffer (pH 7.0). Other conditions were descri- bed in Methods. P~ liberated was determined by the Mar t in-Doty method.

TR[NITROPHEN YL-ADENOSINE TRIPHOSPHATE 643
labile phosphate were 0.94 : 2.00 for TNP-ATP and 0.92 : 2.00 for ATP, indicating that an equimolar phosphate is liberated by heavy meromyosin from TNP-ATP as well as from ATP.
The release of orthophosphate from TNP-ATP was also catalyzed by heavy meromyosin even when CaCl/ was replaced by MgCI 2 or EDTA and the specific activities were compared with those of ATP (Table II). The rate of TNP-ATP hydro- lysis decreased as well as that of ATP hydrolysis by addition of MgC12 in the place of CaCI 2. It should be noted that the rate of TNP-ATP hydrolysis in the presence of EDTA was the same as that in the presence of CaCI2, while the distinct activation of ATP hydrolysis was observed in the presence of EDTA.
A Lineweaver-Burk plot of the hydrolysis of TNP-ATP by heavy meromyo- sin ATPase is shown in Fig. 6. That of ATP is also shown for comparative purpose. The K m and V values for TNP-ATP were 4.2/~M and 0.50 ~tmole Pl per rain per rag, respectively. Those for ATP were l 1.3 pM and 0.66/~mole P~ per rain per rag, res- pectively. These results indicate that TNP-ATP can replace ATP as a substrate for heavy meromyosin.
Glycerinated psoas fibers without a load, when placed in a solution contain- ing TNP-ATP, shortened to the same extent as did ATP.
Difference absorption spectra In visible absorption spectra of TNP-ATP and TNP-adenosine, specific shifts
toward higher wavelengths were produced when heavy meromyosin was added. The difference spectrum due to the interaction of TNP-ATP with heavy meromyosin showed two positive peaks at 414 nm and 518 nm, and a negative trough at 455 nm (Curve A in Fig. 7). The difference spectrum resulting from the interaction of heavy meromyosin with TNP-ADP (Fraction 48 from Peak C in Fig. 3) also had the same positive peaks.
Difference spectra induced by addition of heavy meromyosin to TNP-adeno- sine in the presence and absence of pyrophosphate or ATP are shown in Curves B, C and D in Fig. 7. Interaction of heavy meromyosin with TNP-adenosine resulted in a smaller difference spectrum with positive peaks at 420 nm and 495 nm, and negative troughs at 450 nm and 520 nm (curve B in Fig. 7). When pyrophosphate or ATP was added to the reaction mixture, the positive peak at 495 nm in Curve B shifted toward higher wavelength, 505 nm, and the negative trough at 520 nm dis- appeared (Curves C and D in Fig. 7). The hypochromicity at 450 nm became larger with such polyphosphate compounds.
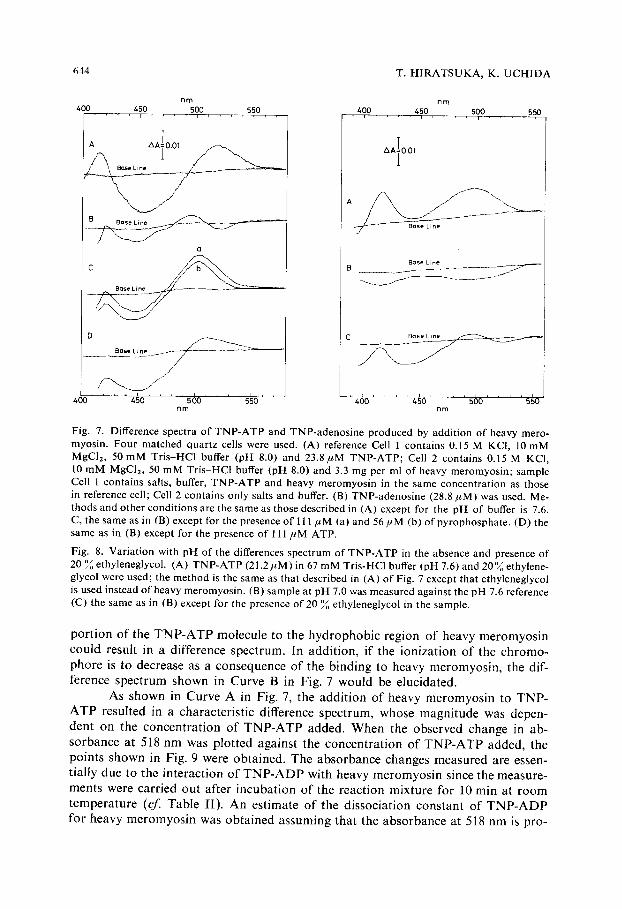
As shown in Curve A in Fig. 8, the difference spectrum with positive peaks at 417 nm and 496 nm was obtained for TNP-Aq'P at pH 7.6 in 20 ~ ethyleneglycol (v/v) and also in 20 0/o dioxane. A similar spectral shift in the absorption spectrum of TNP-adenosine was also obtained by the solvent perturbation.
A fall of visible absorption of TNP-ATP in the range from 400 to 550 nm was made by lowering the pH of the reaction mixture and only a negative phase was observed in the difference spectrum (Curve B in Fig. 8). The difference spectrum due to the pH change was altered when the polarity of the solvent was decreased by the addition of ethyleneglycol (Curve C in Fig. 8). The resulting difference spectrum re- sembled more closely that due to the interaction of TNP-adenosine with heavy me- romyosin (Curve B in Fig. 7). It appears, then, that the binding of the chromophoric
644 T. HIRATSUKA, K. UCHIDA
n m
4O0 450 5OO 550 . . . . [ . . . . i . . . . ~ , ,
A AAIO.O]
400
nm 400 450 500 550
' , . . . . i . . . . ~ . . . . r
AAIO.O1
U Base L l n e ~ - ~ ~ C Base L ne
. . . . 4~o . . . . 56o . . . . s ; o ' ' 4 6 o . . . . ~ o . . . . 5~o . . . . s~o nm nm
Fig. 7. Difference spectra of TNP-ATP and TNP-adenosine produced by addition of heavy mero- myosin. Four matched quartz cells were used. (A) reference Cell 1 contains 0.15 M KCI, 10mM MgClz, 50mM Tris-HCl buffer (pH 8.0) and 23.8#M TNP-ATP; Cell 2 contains 0.15 M KCI, 10 mM MgCI2, 50 mM Tris-HCl buffer (pH 8.0) and 3.3 mg per ml of heavy meromyosin; sample Cell 1 contains salts, buffer, TNP-ATP and heavy meromyosin in the same concentration as those in reference cell; Cell 2 contains only salts and buffer. (B) TNP-adenosine (28.8/zM) was used. Me- thods and other conditions are the same as those described in (A) except for the pH of buffer is 7.6. C, the same as in (B) except for the presence of 111 #M (a) and 56/~M (b) of pyrophosphate. (D) the same as in (B) except for the presence of 111 ,uM ATP.
Fig. 8. Variation with pH of the differences spectrum of TNP-ATP in the absence and presence of 20 % ethyleneglycol. (A) TNP-ATP (21.2/tM) in 67 mM Tris-HCl buffer (pH 7.6) and 20~o ethylene- glycol were used; the method is the same as that described in (A) of Fig. 7 except that ethyleneglycol is used instead of heavy meromyosin. (B) sample at pH 7.0 was measured against the pH 7.6 reference (C) the same as in (B) except for the presence of 20 ~ ethyleneglycol in the sample.
p o r t i o n o f the T N P - A T P m o l e c u l e to the h y d r o p h o b i c r eg ion o f heavy m e r o m y o s i n c o u l d resul t in a d i f fe rence spec t rum. In add i t ion , i f the i on i za t i on o f the c h r o m o -
p h o r e is to decrease as a c o n s e q u e n c e o f the b ind ing to heavy m e r o m y o s i n , the dif- f e rence s p e c t r u m shown in C u r v e B in Fig. 7 w o u l d be e luc ida ted .
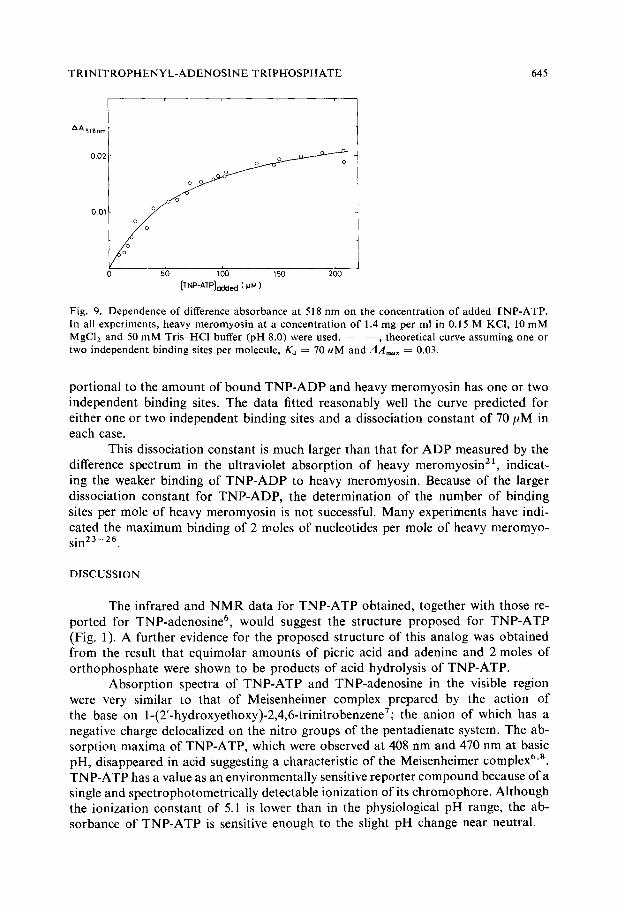
As s h o w n in C u r v e A in Fig. 7, the a d d i t i o n o f heavy m e r o m y o s i n to T N P - A T P resu l ted in a cha rac te r i s t i c d i f ference spec t rum, whose m a g n i t u d e was depen- den t on the c o n c e n t r a t i o n o f T N P - A T P added . W h e n the obse rved change in ab-
s o r b a n c e at 518 n m was p lo t t ed aga ins t the c o n c e n t r a t i o n o f T N P - A T P added , the po in t s s h o w n in Fig. 9 were ob t a ined . T h e a b s o r b a n c e changes m e a s u r e d are essen- t ia l ly due to the i n t e r ac t i on o f T N P - A D P wi th heavy m e r o m y o s i n since the measu re - men t s were ca r r i ed ou t a f te r i n c u b a t i o n o f the r eac t i on mix tu r e fo r l 0 rain at r o o m t e m p e r a t u r e (cf. T a b l e II) . A n e s t ima te o f the d i s soc ia t ion c o n s t a n t o f T N P - A D P fo r heavy m e r o m y o s i n was o b t a i n e d a s s u m i n g tha t the a b s o r b a n c e at 518 n m is p ro-
TRINITROPHENYL-ADENOS1NE TRIPHOSPHATE 645
AA518n m
0.02
O01
o o o [
i
l 0 50 100 150 200
[TNP-AI'P]oo~Iecl (.uM)
Fig. 9. Dependence of difference absorbance at 518 nrn on the concentration of added TNP-ATP. In all experiments, heavy meromyosin at a concentration of 1.4 mg per ml in 0.15 M KCI, 10 mM MgC12 and 50 mM Tris-HCl buffer (pH 8.0) were used. - - , theoretical curve assuming one or two independent binding sites per molecule, Ka = 70 uM and A A , , a x - - 0.03.
portional to the amount of bound TNP-ADP and heavy meromyosin has one or two independent binding sites. The data fitted reasonably well the curve predicted for either one or two independent binding sites and a dissociation constant of 70 ttM in each case.
This dissociation constant is much larger than that for ADP measured by the difference spectrum in the ultraviolet absorption of heavy meromyosin 21, indicat- ing the weaker binding of TNP-ADP to heavy meromyosin. Because of the larger dissociation constant for TNP-ADP, the determination of the number of binding sites per mole of heavy meromyosin is not successful. Many experiments have indi- cated the maximum binding of 2 moles of nucleotides per mole of heavy meromyo- sin 23-26.
DISCUSSION
The infrared and N M R data for TNP-ATP obtained, together with those re- ported for TNP-adenosine 6, would suggest the structure proposed for TNP-ATP (Fig. 1). A further evidence for the proposed structure of this analog was obtained from the result that equimolar amounts of picric acid and adenine and 2 moles of orthophosphate were shown to be products of acid hydrolysis of TNP-ATP.
Absorption spectra of TNP-ATP and TNP-adenosine in the visible region were very similar to that of Meisenheimer complex prepared by the action of the base on 1-(2'-hydroxyethoxy)-2,4,6-trinitrobenzene7; the anion of which has a negative charge deiocalized on the nitro groups of the pentadienate system. The ab- sorption maxima of TNP-ATP, which were observed at 408 nm and 470 nm at basic pH, disappeared in acid suggesting a characteristic of the Meisenheimer complex 6'8. TNP-ATP has a value as an environmentally sensitive reporter compound because of a single and spectrophotometrically detectable ionization of its chromophore. Although the ionization constant of 5.1 is lower than in the physiological pH range, the ab- sorbance of TNP-ATP is sensitive enough to the slight pH change near neutral.

646 T. HIRATSUKA, K. UCHIDA
By placing a sensitive reporter group at one specific position in a protein mole- cule, the changes in its local environment can be examined. In this note an analog of the substrate with a reporter group enabled the placement of the environ- mentally-sensitive chromophore in a specific position on heavy meromyosin, namely the ATP binding site. In addition, TNP-ATP was found to be practically nonfluorescent in aqueous solution, but fluorescent when dissolved in ethanol-water solutions (unpublished results).
In view of the facts that 2',3'-O-diacetyl ATP 27 and 2',3'-O-isopropylidene ATP z8 were hydrolyzed by myosin and that 2-(dansylamino)ethyl triphosphate 5 was hydrolyzed by heavy meromyosin, it is readily expected that TNP-ATP is hydro- lyzed as a substrate for heavy meromyosin. The similarity of the K m and V values for TNP-ATP and ATP indicates the suitability of TNP-ATP as an analog to ATP. The result that TNP-ATP shortened glycerinated psoas fibers is consistent with those for 2',3'-O-diacetyl ATP 27 and 2',3'-O-isopropylidene ATP zS.
The present studies show that heavy meromyosin produces a specific red shift in the visible absorption spectrum of TNP-ATP. Morita 21 has demonstrated that a red shift in the absorption spectrum around 290 nm of heavy meromyosin is due to the formation of a Michaelis complex between ATP and heavy meromyosin. The difference spectrum resulting from the interaction of ATP with heavy mero- myosin showed positive peaks quite typical for the spectral red shift in the ultraviolet absorption. Although the results were not presented, it was observed in the present experiments that TNP-ATP produces a spectral red shift in the ultraviolet absorption of heavy meromyosin. This shift appears to result from the specific interaction of TNP -ATP with the active site of heavy meromyosin. The finding that T N P - A T P is hydrolyzed as a substrate for heavy meromyosin strongly supports a conclusion that the specific red shift in the visible absorption spectrum of TNP-ATP is due to the interaction of a chromophore of the T N P - A T P molecule with or near the binding site of heavy meromyosin.
By addition of pyrophosphate to the reaction mixture, the positive peak at 495 nm in the difference spectrum resulting from the interaction of TNP-adenosine with heavy meromyosin shifted toward higher wavelength. Pyrophosphate can bind to heavy meromyosin and produces the difference spectrum in the ultraviolet absorption of heavy meromyosin 2 ~. The pyrophosphate-binding process may involve a localized conformational change in heavy meromyosin, a consequence of which may be the relocation of certain tyrosine and tryptophan residues 2~. These results suggest that the difference spectrum of TNP-ATP resulting from the interaction with heavy meromyosin arises not only from the binding of the chromophoric portion of the TNP-ATP molecule but also from that of the phosphate portion.
ACKNOWLEDGEMENT
We are indebted Dr K. Yagi for his discussions and supports. This work was supported by a grant from Muscular Dystrophy Association of America Inc.

T RINITROPHENYL-ADENOSINE TRIPHOSPHATE 647
REFERENCES
1 lkehara, M., Ohtsuka, E., Kitagawa, S., Yagi, K. and Tonomura, Y. (1961) J. Am. Chem. Soc. 83, 2679-2686
2 Miller, D. L. and Westheimer, F. H. (1966) J. Am. Chem. Soc. 88, 1511-1513 3 Lecocq, J. (1968)J. Med. Chem. 11, 1096-1097 4 Murphy, A. J. and Morales, M. F. (1970) Biochemistry 9, 1528-1532 5 0 n o d e r a , M. and Yagi, K. (1971) Biochim. Biophys. Acta 253, 254-265 6 Azegami, M. and lwai, K. (1964) J. Biochem. (Tokyo) 55, 346-348 7 Foster, R., Fyfe, C. A. and Morris, J. W. (1965) Recl. Tra~,. Chim. Pays-Bus 84, 516-520 8 Murto, J. (1965) Suom. Kern. (B), 38,255-257 9 Rao, C. N. R. (1969) in The Chemistry of the Nitro and Nitroso Group (Feuer, H., ed.), Part I,
pp. 79-135, lnterscience Publishers, New York 10 Yagi, K. and Yazawa, Y. (1966) J. Biochem. (Tokyo) 60, 450-458 11 Uchida, K. and Hiratsuka, T. (1971) Biochim. Biophys. Acta 229, 310-321 12 Mueller, H. and Perry, S. V. (1961) Biochem. J. 80, 217-223 13 Mueller, H. (1964) J. Biol. Chem. 239, 797-804 14 Fiske, C. H. and SubbaRow, Y. (1925) ,I. Biol. Chem. 66, 375-400 15 Martin, J. B. and Doty, D. M. (1949) Anal. Chem. 21,965-967 16 Mohler, H. (1943) Helv. Chim. Acta 26, 121-129 17 LePage, G. A. (1951) in Manometric Techniques and Related Methods for the Stud>' o[" Tissue
Metabolism (Umbreit, W. W., ed.), pp. 185-197, Burgess Publishing Co., Minneapolis 18 Beaven, G. H., Holiday, E. R. and Johnson, E. A. (1955) in The Nucleic Acids (Chargaff, E. and
Davidson, J. N., eds), Vol. I, pp. 493-553, Academic Press, New York 19 Jardetzky, C. D. and Jardetzky, O. (1960) J. Am. Chem. Soc. 82, 222-229 20 Strauss, M. J. (1970) Chem. Rev. 70, 667-712 21 Morita, F. (1967) J. Biol. Chem. 242, 4501-4506 22 Dyall, L. K. (1960) J. Chem. Soc. 5160-5167 23 Nauss, K. M., Kitagawa, S. and Gergely, J. (1969) J. Biol. Chem. 244, 755-765 24 Lowey, S. and Luck, S. M. (1969) Biochemistry 8, 3195-3199 25 Morita, F. (1969) Biochim. Biophys. Acta 172, 319-327 26 Morita, F. (1971) J. Biochem. (Tokyo) 69, 517-524 27 Hasselbach, W. (1956) Biochim. Biophys. Acta 20, 355-368 28 Ikehara, M., Ohtsuka, E., Uno, H., lmamura, K. and Tonomura, Y. (1965) Biochim. Biophys.
Acta 100, 471-478





























