Embed Size (px)
Citation preview

Camp. Biochem. Physiol., 1977, Vol. 56A, pp. 299 to 304. Pergamon Press. Printed in Great Britain
PRODUCTION OF VOLATILE FATTY ACIDS IN TWO SPECIES OF WALLABY AND IN SHEEP
I. D. HUME
Department of Biochemistry & Nutrition, University of New England, Armidale, N.S.W. 2351, Australia
(Received 25 May 1976)
Abstract-l. The production of volatile fatty acids (VFA) was estimated in Ctro in the stomach and hind gut of two species of wallaby (Thylogale thetis and Macropus rufogriseus) and of sheep (Oh aries) fed chopped lucerne hay.
2. VFA production (kcal/day) was greater in the stomach of M. rufogriseus (42% of digestible energy (DE) intake) than in the sheep (29%) (P < 0.05) and in T thetis (21%) (P < 0.01).
3. In the hind gut, VFA production was greater in the sheep (6.9% of DE intake) than in the wallabies (mean 1.6%) (P < 0.001).
4. The difference in total VFA production between M. rufogriseus (43% of DE intake) and ‘I. thetis (22%) is probably related to the nutritional environment of their preferred habitats, and to their feeding behaviour.
INTRODUCTION
In ruminants little of the dietary carbohydrate is absorbed as hexose because of pre-gastric fermen- tation. The principal end products of this fermen- tation are the steam-volatile fatty acids (VFA) acetic, propionic, butyric and valeric acids, and methane. The VFA are the main energy source for the host animal tissues.
Macropod marsupials have been described as being “ruminant-like” in their digestive physiology (Moir et al., 1956); they have a large sacculated stomach which harbours a dense population of bacteria and protozoa prior to digestion. Significant concentrations of VFA have been found in the anterior region of the stomach, but very low concentrations in the gastric region of the stomach and in the small intestine, sug- gesting that the VFA are absorbed mainly from the stomach. This is supported by observed increases in VFA concentration in venous blood draining the stomach after feeding (Barker, 1961). In contrast to ruminants, it appears that methane is not produced in the stomach fermentation of the eastern grey kan- garoo (Macropus giganteus) (Kempton et al., 1976).
The present study was designed to estimate the apparent contribution of VFA production measured in vitro to the DE intake of two species of wallaby and sheep fed the same diet. The wallabies used were the red-necked pademelon (Thylogale thetis) and the red-necked wallaby (Macropus rufogriseus). Thylogale
thetis is a small wallaby (3.5-7.5 kg bodyweight) which has a preferred habitat of closed-forest (rainfor- est). Its range is restricted to the eastern escarpment of the Great Dividing Range in northern New South Wales and south-eastern Queensland. Macropus rufo- griseus is a larger wallaby (9-13 kg), which is found in both tall open-forest (wet sclerophyll) and open- forest (dry sclerophyll). Its range is adjacent but in- land to that of ?: thetis, and is much more extensive (Troughton, 1941).
Like ruminants, macropod marsupials have a secondary fermentation area in the caecum-proximal colon. Faichney (1969), reported that VFA production
in the ovine hind gut was about 14% of production in the reticula-rumen. In this study comparisons are made between VFA production in the hind gut and the stomach of the three species examined.
MATERIALS AND METHODS
Animals
Four adult pademelons (3.5-7.4 kg liveweight), four adult red-necked wallabies (9.1-12.7 knl and four mature merino sheep (32.8-43.0 kg) ‘were use% The pademelons were trapped near Dorrigo, 150 km east of Armidale, and main- tained in small yards on lucerne hay. The red-necked wal- labies were trapped near Jeogla, 80 km east of Armidale, and maintained indoors in metabolism cages on chopped lucerne hay until used in the experiment. The sheep were maintained on chopped lucerne hay indoors.
During the experiment all animals were housed indivi- dually in metabolism cages which allowed separate collection of urine and faeces. Feed and water containers were attached outside the cages to minimise spillage. The animals were kept in an air-conditioned room (22 + 1’C) under continuous lighting, and fed at 1 hr intervals by automatic feeder in an attempt to maintain steady state conditions of fermentation and digesta flow in the gut. The sheep and the pademelons adapted quickly to their cages. The red-necked wallabies always showed some signs of nervousness, although this decreased as the experiment progressed.
Diet
All animals were fed a diet of chopped lucerne hay. Both wallaby species were fed ad libitum by offering each animal 50% more than its intake on the previous day. Such a large excess was considered essential because of the highly selective feeding pattern observed in the pademelons in preliminary experiments. The amount of feed offered to sheep 1 and 2 was restricted by the size of the automatic feeder to 12OOa dailv. Sheen 1 consumed auorox 73X of this, but sheep-2 consumed*virtually all feed-offered,’ and hence was not fed ad libitum. Sheep 3 and 4 were fed by a larger automatic feeder, and were offered 25% more than their intake on the previous day, ensuring ad libitum feed- ing.
C.&P. 5613~--0 299
300 I. D. HUME
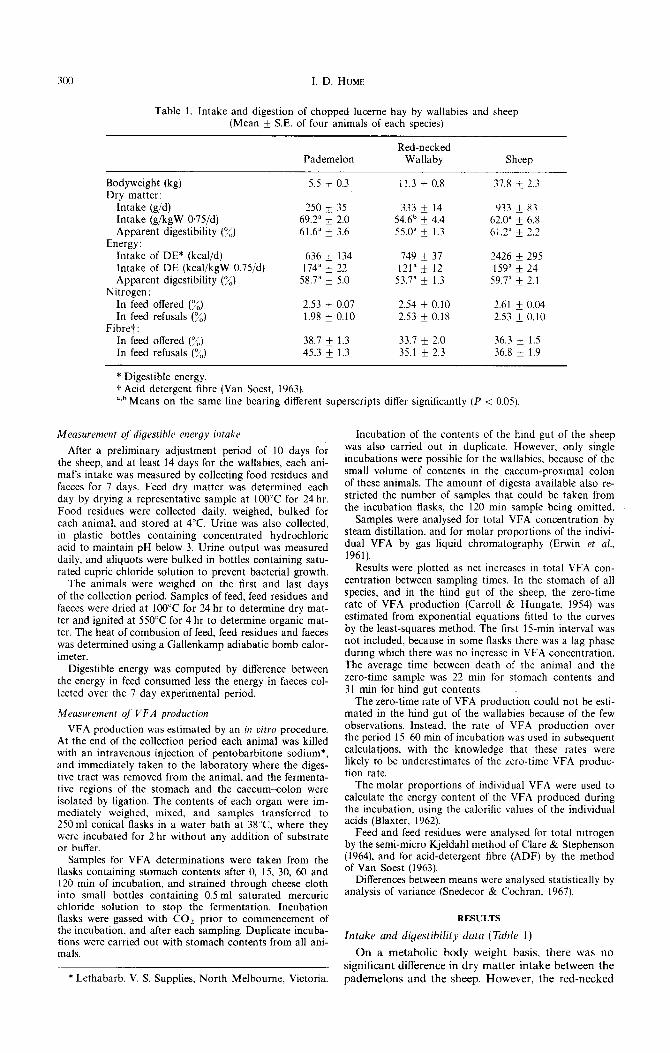
Table 1. Intake and digestion of chopped Lucerne hay by wallabies and sheep (Mean + S.E. of four animals of each species)
Pademelon Red-necked
Wallaby Sheep
Bodyweight (kg) Dry matter:
Intake (g/d) Intake (g/kgW 0,75/d) Apparent digestibility (%)
Energy: Intake of DE* (kcal/d) Intake of DE (kcal/kgW 0.75/d) Apparent digestibility (%)
Nitrogen: In feed offered (%) In feed refusals (%)
Fibret: In feed offered (%) In feed refusals (7”)
5.5 * 0.3
250 ri: 35 69.2” k 2.0 61.6” k 3.6
636 k 134 174” * 22
58.7” * 5.0
2.53 + 0.07 1.98 * 0.10
38.7 k 1.3 45.3 + 1.3
11.3 k 0.8
333 * 14 54.6b k 4.4 55.0“ + 1.3 -
749 + 37 121” * 12
53.7” & 1.3
2.54 k 0.10 2.53 f 0.18
33.7 * 2.0 35.1 & 2.3
37.8 * 2.3
933 + 83 62.0” k 6.8 61.2” + 1.2 _-
2426 f 295 159” f 24
59.7” + 7.1 _-
2.61 + 0.04 2.53 k 0.10
36.3 + 1.5 36.8 + 1.9
* Digestible energy. f Acid detergent fibre (Van Soest, 1963). “.“Means on the same line bearing different superscripts differ significantly (P < 0.05).
Measurement qf digestible energy intake
After a preliminary adjustment period of 10 days for the sheep. and at least 14 days for the wallabies, each ani- mal’s intake was measured by collecting food residues and faeces for 7 days. Feed dry matter was determined each day by drying a representative sample at 100°C for 24 hr. Food residues were collected daily, weighed, bulked for each animal, and stored at 4°C. Urine was also collected, in plastic bottles containing concentrated hydrochloric acid to maintain pH below 3. Urine output was measured daily, and aliquots were bulked in bottles containing satu- rated cupric chloride solution to prevent bacterial growth.
The animals were weighed on the first and last days of the collection period. Samples of feed, feed residues and faeces were dried at 100°C for 24 hr to determine dry mat- ter and ignited at 550°C for 4 hr to determine organic mat- ter. The heat of combusion of feed, feed residues and faeces was determined using a Gallenkamp adiabatic bomb calor- imeter.
Digestible energy was computed by difference between the energy in feed consumed less the energy in faeces col- lected over the 7 day experimental period.
Measurement of VFA production
VFA production was estimated by an in vitro procedure. At the end of the collection period each animal was killed with an intravenous injection of pentobarbitone sodium*, and immediately taken to the laboratory where the diges- tive tract was removed from the animal, and the fermenta- tive regions of the stomach and the caecum-colon were isolated by ligation, The contents of each organ were im- mediately weighed, mixed, and samples transferred to 250ml conical flasks in a water bath at 38°C where they were incubated for 2 hr without any addition of substrate or buffer.
Samples for VFA determinations were taken from the fasks containing stomach contents after 0, 15, 30, 60 and 120 min of incubation, and strained through cheese cloth into small bottles containing 0.5 ml saturated mercuric chloride solution to stop the fermentation. Incubation flasks were gassed with CO, prior to commencement of the incubation, and after each sampling. Duplicate incuba- tions were carried out with stomach contents from all ani- mals.
* Lethabarb. V. S. Supplies, North Melbourne, Victoria.
Incubation of the contents of the hind gut of the sheep was also carried out in duplicate. However, only single incubations were possible for the wallabies, because of the small volume of contents in the caecum-proximal colon of these animals. The amount of digesta available also re- stricted the number of samples that could be taken from the incubation flasks, the 120 min sample being omitted.
Samples were analysed for total VFA concentration by steam distillation, and for molar proportions of the indivi- dual VFA by gas-liquid chromatography (Erwin et al.. 1961).
Results were plotted as net increases in total VFA con- centration between sampling times. In the stomach of all species, and in the hind gut of the sheep, the zero-time rate of VFA production (Carroll & Hungate. 1954) was estimated from exponential equations fitted to the curves by the least-squares method. The first 15-min interval was not included, because in some flasks there was a lag phase during which there was no increase in VFA concentration. The average time between death of the animal and the zero-time sample was 22 min for stomach contents and 31 min for hind gut contents.
The zero-time rate of VFA production could not be esti- mated in the hind gut of the wallabies because of the few observations. Instead, the rate of VFA production over the period 1560 min of incubation was used in subsequent calculations, with the knowledge that these rates were likely to be underestimates of the zero-time VFA produc- tion rate.
The molar proportions of individual VFA were used to calculate the energy content of the VFA produced during the incubation, using the calorific values of the individual acids (Blaxter, 1962).
Feed and feed residues were anaiysed for total nitrogen by the semi-micro Kjeldahl method of Glare & Stephenson (1964). and for acid-detergent fibre (ADF) bv the method of Van Soest (1963). -
_~
Differences between means were analysed statistically by analysis of variance (Snedecor & Cochran. 1967).
RESULTS
Intake and digestibility data (Table 1)
On a metabolic body weight basis, there was no
significant difference in dry matter intake between the
pademelons and the sheep. However, the red-necked

VFA production in wallabies 301
Table 2. Dry matter content and weights of stomach’ and hind gut’ digesta of wallabies and sheep
(Mean 5 SE. of four animals of each species)
Pademelon Red-necked
Wallaby Sheep
Stomach : Weight of contents (g) Weight of contents
r/, of body wt) Dry matter of contents (%)
Hind gut: Weight of contents (g) Weight of contents
(% of body wt) Dry matter of contents (%f
530 + 82 1035 + 118 5240 2 422 9.5” * 0.2 9.4” * 1.3 14.2’ & 1.9
18.5k t_ 0.3 16.3’ _t 0.5 14.7” 2 0.8
73 + 5 58 + 14 1802 _t 207 1.3” * 0.2 0.5” If: 0.1 4.9’ * 0.9
12.6” * 0.4 13.Q * 2.7 13.8” & 0.1
’ The fermentation region of the stomach (i.e. the rumen-reticulum of the sheep, and the regions anterior to the fundic pouch of the wallabies).
’ The caecum plus the proximal colon. “.bMeans on the same line bearing different superscripts differ significantly (P -c 0.05). k,‘*m Means on the same line bearing different superscripts differ significantly (P < 0.01). ‘+Means on the same line bearing different superscripts differ significantly (P < 0.001).
wallabies ate less dry matter (54,6 g/kg 0.75/day) than the pedemelons (69.2g/kg 0.75/day) (P < 0.05). There were no significant differences between any species in dry matter digestibility, nor in the intake of digest- ible energy.
Compuriso~ of capacities of digestive tracts (Tub& 2)
The contents of the rumen-reticulum of the sheep constituted a larger proportion of body weight (14.2%) than did the contents of the fermentative regions of the stomach of the pademelons (9.5%) and red-necked wallabies (9.4yi) (P < 0.001). Similarly, the contents of the sheep hind gut (i.e. caecum and proxi- mal colon) constituted a larger proportion of body- weight (4.9%) than did the hind gut contents of the two macropod species (1.3% and 0.5%) (P < 0.001). There was no significant difference between the two macropod species in the relative size of the stomach nor of the caecum-proximal colon.
The dry matter content of the red-necked wallaby stomach digesta (16,3oj,) was higher than that of the sheep (14.7%) (P < @OS), but lower than that of the pademelon (18.5%) (P -C 0.01). In the hind gut the dry matter content of the digesta from the red-necked wallaby (19.6%) was higher than both the sheep (13.8%) and the pademelon (12.6%) (P < 0.05). There was no consistent difference in dry matter content between the stomach and the hind gut.
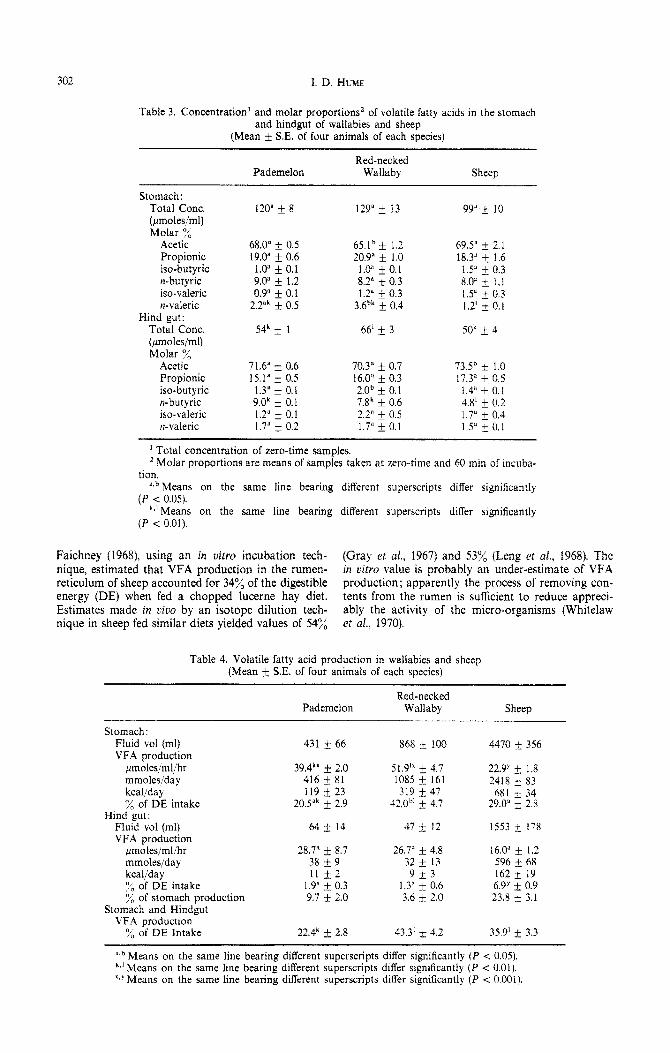
Concentrations and molar proportions of VFA (Table 3)
TotaI VFA concentrations were higher in the stomach than in the hind gut of all species (P =C 0.001). Among species there were no significant differences in VFA levels in the stomach, but jhe hind gut of the red-necked wallaby contained a higher VFA concentration (66 ,umoles/ml) than that of the other two species (54 and 50 pmoles/ml) (P < 0.01).
The molar proportion of acetic acid was lower in the stomach than in the hind gut (P < O.OOl), and prapionic acid higher (P -C 0.001). The proportion of acetic acid was lower in the macropod species than
in the sheep (P < O.Ol), and lower in the red-necked wallaby than in the pademelon (P < 0.05). There were no significant species differences in propionic acid proportions.
The molar proportion of n-butyric acid was similar in stomach and hind gut digesta, and lower in the sheep than in the macropods (P -C 0.01).
The proportion of n-valeric acid was higher in stomach than in hind gut digesta (P < O.Ot), and lower in the sheep than in the macropods (P < 0.01).
The molar proportions of VFA did not change sig- nificantly over the 12%min incubation period.
Production of VFA (Table 4)
The zero-time rate of VFA production &moles/ m&r) was faster in the stomach than the hind gut of all species (P < 0.01). In the stomach, VFA produc- tion was faster in the macropods than in the sheep (P -=z O.#l), and faster in the red-neck~.~vallaby than in the pademelon (P < 0.01). In the hind gut, the rate of VFA production was more variable but similar in both macropod species, and not significantly different from the sheep, even though the sheep mean was only 58% of the macropod mean.
As a proportion of the animal’s intake of DE, VFA production was greater in the stomach of the red- necked wallaby (42.0%) than in the sheep (29.00& (P < 0.05) and the pademelon (20.5”/,) (P < 0.01). There was no significant difference in the contribution made by VFA production in the hind gut of either macropod (mean I.@?& and both were less than in the sheep (6.9%) (P -C 0.001).
Total VFA production (i.e. stomach and hind gut together) was not significantly different between the red-necked wallaby (43.3%) and the sheep (35.9%), and both were greater than the pademelon (22.4%) (P C 0.01).
DISCUSSJON
The production of VFA has been measured in ruminants such as sheep both in vitro and in viva.
302 I. D. HUME
Table 3. Concentration’ and molar proportions’ of volatile fatty acids in the stomach and hindgut of wallabies and sheep
(Mean k SE. of four animals of each species)
Pademelon Red-necked
Wallaby Sheep
Stomach: Total Cont. 120” * 8 129” + 17 - . 99”+ 10 (pmoles/ml) Molar y/,
Acetic 68.0” k 0.5 65.1b i. 1.2 69.5” k 2.1 Propionic !9.01 & 0.6 20.9” + 1.0 18.3” + 1.6 iso-butyric 1.0” f 0.1 1.0” + 0.1 1.5” + 0.3 n-butyric 9.0” f 1.2 8.2’ f 0.3 8.0” & 1.1 iso-valeric 0.9” * 0.1 1.2” * 0.3 1.5” + 0.3 n-valeric 2.2’k + 0.5 - 3.6bk + 04 - 1 1.2’ * 0.1
Hind gut: Total Cont. 54k + I 66’ & 3 50k + 4 (pmoles/ml) Molar sj’,
Acetic 71.6” 2 0.6 70.3” f 0.7 73.5h + 1.0 Propionic 15.1” f 0.5 16.0” f 0.3 17.3” f 0.5 iso-butyric 1.3” * 0.1 2.0b + 0.I 1.4;’ -t 0 I n-butyric 9.0k * 0.1 7.8’ + 0.6 4.8’ + 0:2 iso-valeric 1.2” * 0.1 2.2” * 0.5 1.7” + 04 n-valeric 1.7” + 0.2 1.7” * 0.1 1.5” IO:1
’ Total concentration of zero-time samples. ’ Molar proportions are means of samples taken at zero-time and 60 min of incuba-
tion. ‘JI Means on the same line bearing different superscripts differ significantly
(P < 0.05). k+‘Means on the same line bearing different superscripts differ significantly
(P < 0.01).
Faichney (1968). using an in vitro incubation tech- (Gray et al., 1967) and 53% (Leng et at., 1968). The nique, estimated that VFA production in the rumen- in t&o value is probably an under-estimate of VFA reticulum of sheep accounted for 34% of the digestible production; apparently the process of removing con- energy (DE) when fed a chopped lucerne hay diet. tents from the rumen is sufficient to reduce appreci- Estimates made in ~iuo by an isotope dilution tech- ably the activity of the micro-organisms (Whitelaw nique in sheep fed similar diets yielded values of 54% et al., 1970).
Table 4. Volatile fatty acid production in wallabies and sheep (Mean + SE. of four animals of each species)
Pademelon Red-necked
Wallaby Sheep
Stomach: Fluid vol (ml) 431 i: 66 868 100 jI 4470 -f 356 VFA production
pmoles/ml/hr 39.4k” * 2.0 51.9’” ir 4.7 22.9y + 1.8 mmoles/day 416rf:81 1085 rt 161 2418 + 83 kcal/day 119 rf: 23 319 + 47 681 34 “/d of DE intake 20.5”k + 2.9 42 ( Oh’ - + 4.7
I_ 29.0” ri: 2.8
Hind gut: Fluid vol (ml) 64 i: 14 47 & 12 1553 t 178 VFA production
~moles/ml/hr 28.7” k 8.7 26.7” rt 4.8 16.0” ri; 1.2 mmoles/day 38 i9 32;t 13 596 + 68 kcal/day 11 +2 9+3 162 rt 19 y0 of DE intake 1.9” 2 0.3 1.3” I 0.6 6.9’ + 0.9 Y0 of stomach production 9.1 rfr 2.0 3.6 i_ 2.0 23.8 i 3.1
Stomach and Hindgut VFA production
‘>* of DE Intake 22.4” 2 2.8 43.3’ i: 4.2 35.9’ & 3.3
“.h Means on the same line bearing different superscripts differ significantiy (P < 0.05). k~’ Means on the same line bearing different superscripts differ significantly (P < 0.01). ‘ay Means on the same line bearing different superscripts differ significantly (P < 0.001).

VFA production in wallabies 303
In the present study the choice of method for esti- mating VFA production rates in the stomach was re- stricted by the dependence of the in uiuo approach on rapid mixing and uniform distribution of infused “C-labelled acid throughout the stomach contents. This is known to be achieved in the rumen-reticulum of sheep (Leng & Leonard, 1965), but the tubular structure of the macropod stomach (Moir et al., 1956; Griffiths & Barton, 1966) makes complete mixing of contents unlikely. This is supported by the relatively slow mixing of coloured dye in the stomach of the quokka (Setonix brachyurus) observed by Ward (un- published). Hence the comparisons between sheep and macropods in this study were necessarily made in dtro. Albeit that, in sheep, published estimates of VFA production made in vitro differ from those made irl vice, there is little reason to doubt that the com- parisons made between species in vitro are valid. All the animals were kept under constant illumination, and were offered feed hourly to encourage uniform rates of eating throughout the day, so as to ensure that the fermentation in the stomach and in the hind gut approached a steady state. This, together with thorough mixing of the contents of the regions of fer- mentation before sampling for the in uitro incuba- tions, should have minimised any differences in VFA production rate between species caused by diurnal variations in substrate availability.
The contribution made by ruminal VFA produc- tion to the DE intake of the sheep in this experiment (29%) is close to that estimated by Faichney (1968) by a similar technique in sheep fed a similar diet
(34%). Apparent VFA production in the stomach of the
pademelon made a relatively minor contribution to DE intake (21%). The diet selected by this small wal- laby in its preferred habitat of sub-tropical rain forest is likely to be relatively low in fibre content at all times of the year. This would be consistent with a comparatively limited fermentation in the stomach.
In contrast, VFA production in the stomach of the red-necked wallaby was unexpectedly high (42% of DE intake), at least as high as in the sheep. The differ- ence in VFA production between the pademelon and the red-necked wallaby may be related to the habitat of the latter species of drier eucalypt forest, in which the fibre content of the diet selected by the macropods probably varies through the year. Unfortunately no detailed studies have been made of the dietary habits of either of the above macropod species.
The contribution to the sheep’s intake of DE made by VFA production in the hind gut in this experiment (6.9%) is similar to an estimate of 5.3% in sheep fed a dried grass diet made in ciuo by Faichney (1969). VFA production in the hind gut of both wallaby spe- cies was small (mean 1.6”: of DE intake) compared with the sheep (Table 4). This can be explained by the comparatively small size of the wallaby hind gut (Table 2) rather than by a slower rate of VFA pro- duction. In fact, the rates of VFA production in the macropod hind guts (Table 4) are probably underesti- mates of the zero-time production rate because of the restrictions imposed by the small size of the caecum- proximal colon on the number of samples that could be used for the production estimates.
Rapid fermentation rates in ruminants appear to
be associated with high rumen turnover rates (Hungate et al., 1960) and with lower molar propor- tions of acetate in the VFA produced (Hume, 1970). The relationship between rapid fermentation and high turnover rate is consistent with observations of Del- low & Hume (unpublished) that the retention times of both solid and liquid digesta are significantly shorter in the stomach of iY thetis than in the ovine rumen. A relationship between rapid fermentation and lower acetate molar proportions is evident in this experiment; acetate molar proportions were lower in both wallaby species than in the sheep, and the rate of VFA production higher. Similarly, within the mac- ropods, the molar proportion of acetate in the stomach of the red-necked wallaby was lower than in the pademelon stomach, and the rate of VFA pro- duction higher.
In all three species studied, the rate of VFA produc- tion was higher in the stomach than in the hind gut (Table 4). This difference in fermentation rate was again reflected in the VFA produced; the molar pro- portion of acetate was lower in the stomach than the hind gut, and that of propionate higher. Further, the total concentration of VFA in the stomach was double the concentration in the hind gut. These find- ings are all consistent with results reported by Hun- gate et al. (1960) and can be explained in terms of the composition of the substrate entering the two fer- mentation regions. The digesta entering the caecum- proximal colon of sheep contains relatively more structural carbohydrate and less soluble carbohydrate and protein than ingested food (Oh et al., 1972). A similar difference would be expected in macropods. Fermentation of high-cellulose substrates yields high molar proportions of acetate, and low proportions of propionate (Annison, 1954) consistent with the VFA patterns found in the hind gut of all three spe- cies studied in this experiment.
Hungate et al. (1960) concluded that a fast fermen- taion rate, and hence a lower retention time, was necessary in small ruminants because of an apparent limitation to the size the rumen-reticulum could reach as a proportion of body size; in ruminants ranging in weight from 3.7 to 522 kg, the contents of the rumen-reticulum averaged 12% of the body- weight. They argued that this limitation should be reflected in selection by the smaller ruminants for a more readily fermentable diet, in a higher food intake per unit metabolic bodyweight, or both. In this exper- iment, the small macropod, T thrtis, consumed signi- ficantly more dry matter per day, on a metabolic bodyweight basis, than the red-necked wallaby. How- ever, the higher food intake was associated with a significantly slower rate of VFA production in 7: thetis, contrary to the hypothesis advanced by Hun- gate et al. (1960). An explanation for this apparent anomaly may be forthcoming from present studies on the relationship between stomach structure and digesta flow rate in a number of macropod species.
The pademelons also selected a diet of higher nutri- tive value than did the red-necked wallabies or the sheep. The feed consumed by the pademelons was 22% higher in nitrogen and 17% lower in fibre con- tent than the feed offered (Table 1). In contrast, the red-necked wallabies consumed a diet no different in nitrogen or fibre content to the feed offered. The diet

304 I. D. HUME
consumed by the sheep was 3% higher in nitrogen and 774 lower in fibre content than the feed offered. Thus, although it might be expected that the greater VFA production in the red-necked wallabies would be associated with a more complete dry matter diges- tion, this was not the case (Table l), due, in part at least, to the highly selective feeding behaviour of the pademelons.
On the basis of the relatively minor contribution of VFA production to DE intake, 7: thetis can be described as being less “ruminant-like” than M. rufo- griseus. It would seem that the importance of mi- crobial fermentation is minimised in ?: thetis by a range restricted to sub-tropical rain forest, and further, by its highly selective feeding behaviour.
Acknowledyrrnerlts-The author is indebted to Mr. & Mrs. C. Rodgers, on whose property the pademelons were trapped, and to Mr. G. Gray, on whose property the red- necked wallabies were trapped, for their willing cooper- ation at all times.
The work was supported by the Australian Research Grants Committee.
REFERENCES
ANNISON E. F. (1954) Some observations on volatile fatty acids in the sheep’s rumen. Biochem. J. 57, 400-405.
BARKER J. M. (1961) The metabolism of carbohydrate and volatile fatty acids in the marsupial Setonix hrachyurus. Quart. J. Exp. Physiol. 46, 54-68.
BLAXTER K. L. (1962) The Energy Metabolism of Ruminants. Hutchinson & Co., London.
CARROI.L E. J. & HUNGATE R. E. (1954) The magnitude of the microbial fermentation in tde boiine rumen. Appl. Microhiol. 2, 205-214.
CLARE N. T. & STEPHENSON A. E. (1964) Measurement of feed intake by grazing cattle and sheep. X. Determina- tion of nitrogen in faeces and feeds using an Auto Ana- lyser. N.Z. J. Agric. Res. 7, 198.
EKWIN E. S.. MARCO G. J. & EMERY E. M. (1961) Volatile fatty acid analyses of blood and rumen fluid by gas chro- matography. J. Dairy Sci. 44, 1768-1771.
FAICHNEY G. J. (1968) The production and absorption of volatile fatty acids from the rumen of the sheep. Aust. J. Agric. Res. 19, 791-802.
FAICHNEY G. J. (1969) Production of volatile fatty acids in the sheep caecum. Aust. J. Agric. Res. 20, 491498.
GRAY F. V., WELLER R. A., PILGRIM A. F. & JONES G. B. (1967) Rates of production of volatile fatty acids in the rumen V. Evaluation of fodders in terms of volatile fatty acid produced in the rumen of the sheep. Aust. J. Agric. Res. 18, 625-634.
GRIFFITHS M. & BARTON A. A. (1966) The ontogeny of the stomach in the pouch young of the red kangaroo. C.S.I.R.O., Wildl. Res. 11, 169-183.
HUME I. D. (1970) Synthesis of microbial protein in the rumen. II. A response to higher volatile fatty acids. Aust. J. Agric. Res. 21. 297-304.
HUNGATE R. E., PHILLIPS G. D., MCGREGOR A., HUNGATE D. P. & BUECHNER H. K. (1960) Microbial fermentation in certain mammals. Science, N.Y 130, 1192-1194.
KEMPTON T. J.. MURRAY R. M. & LENG R. A. (1976) Rates of production of methane in the grey kangaroo and sheep. Aust. J. Biol. Sci. 29, 209-214.
LENG R. A., CORE& J. L. & BRETT D. J. (1968) Rates of production of volatile fatty acids in the rumen of graz- ing sheep and their relation to ruminal concentrations. Br. J. Nutr. 22, 5748.
LENG R. A. & LEONARD G. J. (1965) Measurement of the rates of production of acetic, propionic and butyric acids in the rumen of sheep. Br. J. Nutr. 19, 469484.
MOIR R. J., SOMERS M. & WARING H. (1956) Studies on marsupial nutrition. I. Ruminant-like digestion in the herbivorous marsupial Setonix brachyurus (Quoy & Gai- mard). Aust. J. Biol. Sci. 9, 293-304.
OH J. H., HUME I. D. & TORELL D. T. (1972) Development of microbial activity in the alimentary tract of lambs. J. Anim. Sci. 35, 450-459.
SNEDECOR G. W. & C~CHRAN W. G. (1967) Statistical Methods. 6th Edn Iowa State University Press, Ames.
TROUGHTON E. (1941) Furred Animals of Australia. Angus & Robertson, Sydney.
VAN SOEX P. J. (1963) Use of detergents in the analysis of fibrous feeds. II. A rapid method for the determina- tion of fiber and lignin. J. Ass. Ojf Agric. Chem. 46, 829-835.
WHITELAW F. G., HYLDGAARD-JENSEN J., REID R. S. & KAY M. G. (1970) Volatile fatty acid production in the rumen of cattle given an all-concentrate diet. Br. J. Nutr. 24. 179-195.





























