Embed Size (px)
Citation preview

Aminoacizi si Proteine
Proteinele sunt macromolecule din sistemele vii care indeplinesc functii majore pentru celula: rol de catalizatori; rol de transport si depozitare a anumitor molecule (O2), in sustinerea mecanica, in imunitate, in transmiterea impulsului nervos, in coordonarea cresterii si diferentierii celulare.
Proteinele sunt polimeri constituiti din unitati monomerice numite aminoacizi.
aminoacizi
aminoacid

Aminoacizi si Proteine
Aminoacizi Structura, clasificare, proprietati fizico-chimice
Legatura peptidicaAlte tipuri de legaturi chimice din moleculele proteiceProteine- nivele de organizare
Structura primarăStructura secundarăStructura terţiarăStructura cuaternară

?
AcidAmina
Aminoacizi
OH
O
H
H2N
C
C
R
= acizi aminaţi, sunt compuşi chimici cu masă moleculară mică, ce conţin în molecula lor două funcţii caracteristice: o grupare carboxilică (-COOH) şi o grupare aminică (-NH2) si un radical (R) alifatic sau aromatic care contine diferite grupari functionale: hidroxil, sulfhidril, carboxil, amino, guanidino, fenil, indol, imidazol etc.
În natură se întâlnesc peste 150 aminoacizi, dintre care 20 sunt universal răspândiţi. Aminoacizii din constituţia proteinelor prezintă totdeauna gruparea aminică în poziţia faţă de gruparea carboxilică.

Proprietati fizice ale aminoacizilor
• Solubilitatea aminoacizilor în apă este variată (între 162 g % pentru prolină şi 0.01 % pentru cisteină). Toţi aminoacizii se dizolvă în baze şi acizi diluaţi.
• Cu excepţia glicocolului, aminoacizii sunt substanţe optic active, datorită existenţei în molecula lor a cel puţin unui atom de carbon asimetric (C).

Stereoizomerii aminoacizilor
• Se disting două serii sterice. Seria L şi seria D, în funcţie de dispoziţia grupării α-aminice: spre stânga - pentru seria L (sau configuraţia S, "sinister"), sau spre dreapta - pentru seria D (sau configuraţia R, "rectus"), dacă în formula de proiecţie plană (Fischer) carboxilul este figurat în partea de sus.
• Sensul de rotaţie a planului luminii polarizate nu depinde de apartenenţa la seria sterică. Activitatea optică se notează şi la aminoacizi cu (+) - spre dreapta ("dextrogir"), şi (-) - spre stânga ("levogir").
COOH
CH
R
H2N
COOH
C NH2
R
H
L D
Stereoizomerii aminoacizilor

L-aminoacizii
• Majoritatea aminoacizilor naturali fac parte din seria L ("levo"). Proteinele animale sunt formate numai din L-aminoacizi, legaţi între ei prin legături peptidice: -CO-NH-.
• La microorganisme, precum şi în unele tumori maligne au fost găsiţi şi aminoacizi cu configuraţia D (“dextro").

Clasificarea aminoacizilor
• Dupa polaritatea si sarcina radicalului:
1) Aminoacizi cu radical R nepolar
2) Aminoacizi cu radical R polar
3) Aminoacizi acizi (monoamino-dicarboxilici)
4) Aminoacizi bazici
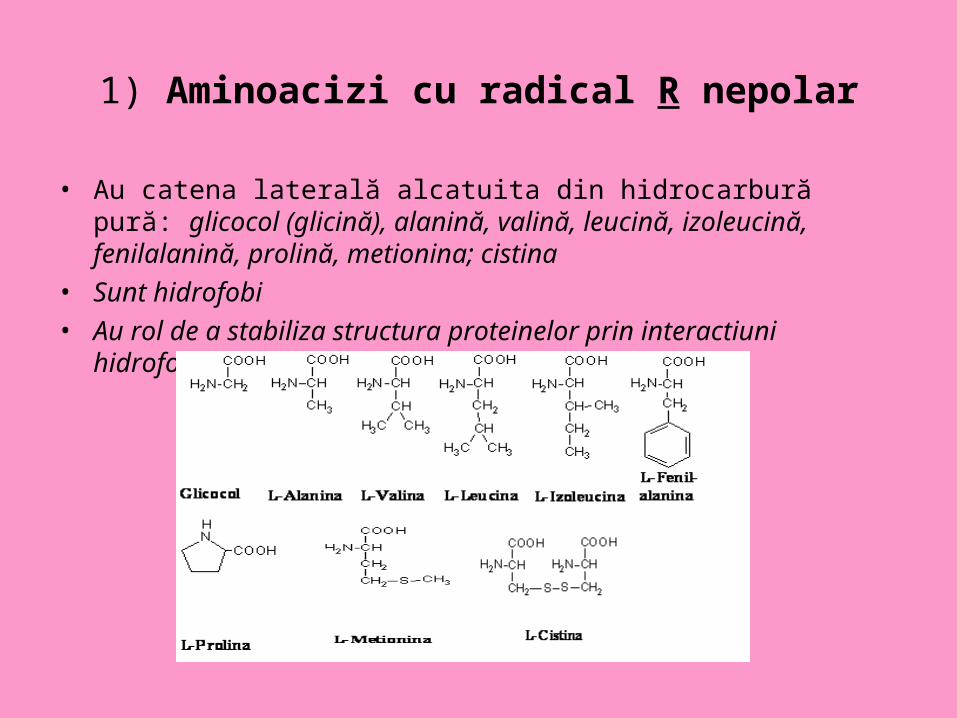
1) Aminoacizi cu radical R nepolar
• Au catena laterală alcatuita din hidrocarbură pură: glicocol (glicină), alanină, valină, leucină, izoleucină, fenilalanină, prolină, metionina; cistina
• Sunt hidrofobi • Au rol de a stabiliza structura proteinelor prin interactiuni hidrofobe.

2) Aminoacizi cu radical R polar
• Au un radical polar cu grupări neionizate în catenă: serină, treonină, cisteină, triptofan, tirozină, asparagină, glutamină.
• Sunt mai hidrofile, deoarece gruparile functionale ale radicalului polar stabilesc punti de H cu apa.
• Triptofanul si tirozina sunt partial hidrofobe si mult mai polare decat fenilalanina. Intra in constitutia centrilor activi ai unor enzime. Cei doi aminoacizi aromatici absorb lumina UV la ~280 nm.

3.Aminoacizi acizi (monoamino-dicarboxilici)
• conţin în radicalul R o a doua grupare carboxilică, ce le conferă caracter acid: acidul aspartic (asparaginic), acidul glutamic.

4. Aminoacizi bazici
• conţin în radicalul R o grupare bazică suplimentară:
lizină, arginină, histidină.
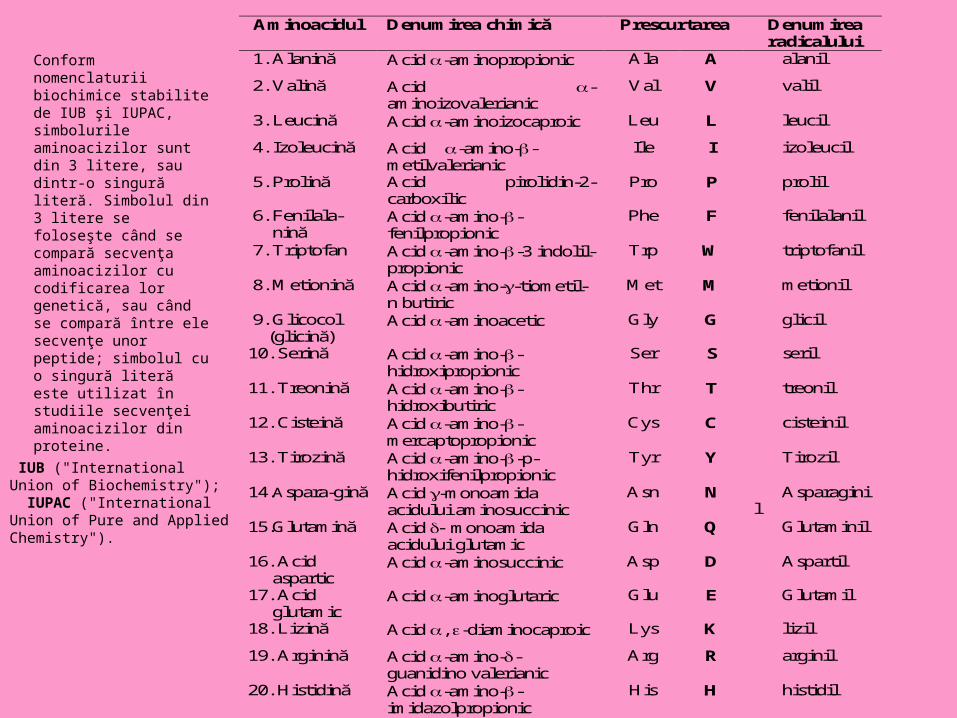
Conform nomenclaturii biochimice stabilite de IUB şi IUPAC, simbolurile aminoacizilor sunt din 3 litere, sau dintr-o singură literă. Simbolul din 3 litere se foloseşte când se compară secvenţa aminoacizilor cu codificarea lor genetică, sau când se compară între ele secvenţe unor peptide; simbolul cu o singură literă este utilizat în studiile secvenţei aminoacizilor din proteine.
IUB ("International Union of Biochemistry"); IUPAC ("International Union of Pure and Applied Chemistry").
Aminoacidul Denumirea chimică Prescurtarea Denumirea radicalului
1. Alanină Acid -aminopropionic Ala A alanil
2. Valină Acid -aminoizovalerianic
Val V valil
3. Leucină Acid -aminoizocaproic Leu L leucil
4. Izoleucină Acid -amino--metilvalerianic
Ile I izoleucil
5. Prolină Acid pirolidin-2-carboxilic
Pro P prolil
6. Fenilala- nină
Acid -amino--fenilpropionic
Phe F fenilalanil
7. Triptofan Acid -amino--3 indolil-propionic
Trp W triptofanil
8. Metionină Acid -amino--tiometil-n butiric
Met M metionil
9. Glicocol (glicină)
Acid -aminoacetic Gly G glicil
10. Serină Acid -amino--hidroxipropionic
Ser S seril
11. Treonină Acid -amino--hidroxibutiric
Thr T treonil
12. Cisteină Acid -amino--mercaptopropionic
Cys C cisteinil
13. Tirozină Acid -amino--p-hidroxifenilpropionic
Tyr Y Tirozil
14.Aspara-gină Acid -monoamida acidului aminosuccinic
Asn N Asparaginil
15.Glutamină Acid monoamida acidului glutamic
Gln Q Glutaminil
16. Acid aspartic
Acid -aminosuccinic Asp D Aspartil
17. Acid glutamic
Acid -aminoglutaric Glu E Glutamil
18. Lizină Acid , -diaminocaproic Lys K lizil
19. Arginină Acid -amino--guanidino valerianic
Arg R arginil
20. Histidină Acid -amino--imidazolpropionic
His H histidil

Aminoacizi esentiali pentru organismul uman
• = aminoacizii pe care organismul uman, ca şi cel al majoritătii animalelor nu îi poate sintetiza din lipsa unor sisteme enzimatice adecvate sau a unor precursori disponibili.
• Valina, izoleucina, leucina, treonina, metionina, fenilalanina, triptofan, lizina, arginina, histidina

Aminoacizi care intră doar în structura unui grup restrâns de proteine
• In afara celor 20 aminoacizi, există şi aminoacizi aminoacizi intră doar în structura unui grup restrâns de proteine, cum sunt 5-hidroxiprolina şi 5-hidroxilizina, care intră în structura colagenului, sau N-metillizina,care intră în structura actinei (o proteină musculară).
• Aceşti aminoacizi provin din aminoacizii naturali prolină şi lizină, cu structură modificată “postsintetic”.

Aminoacizi care nu intra in structura proteinelor
• Aminoacizii care nu intră niciodată în structura proteinelor dar care au un rol metabolic si structural important pentru organism: β-alanina (intră în structura coenzimei A), acidul γ-aminobutiric (prescurtat GABA, este un mediator chimic la nivelul sistemului nervos central), 5-hidroxitriptofanul (precursor al serotoninei - o "amină biogenă"), ornitina şi citrulina (intermediari în sinteza ureei), homocisteina şi homoserina (intermediari în metabolismul unor aminoacizi).

Proprietăţile acido-bazice ale aminoacizilor • Având în moleculă atât grupări acide cât şi bazice, aminoacizii au
caracter amfoter, acţionând astfel fie ca acid (donor de protoni), fie ca bază (acceptor de protoni).
• În soluţie, gruparea carboxilică poate să disocieze de H+, căpătând sarcină negativă (COO-), iar gruparea aminică, bazică, poate prelua ioni de H+ (poate fi protonată), căpătând sarcină pozitivă (-NH3
+). Dacă ambele grupări funcţionale ale unui aminoacid au sarcini, se obţin zwitterioni (din germana ioni hibrizi )=amfiioni.
(mediu bazic)(amfion)(mediu acid)
AnionIon hibridCation
OH-+H++
H2O
COO-
CH
R
H2N+
COO-
CH
R
H3N+
COOH
CH
R
H3N
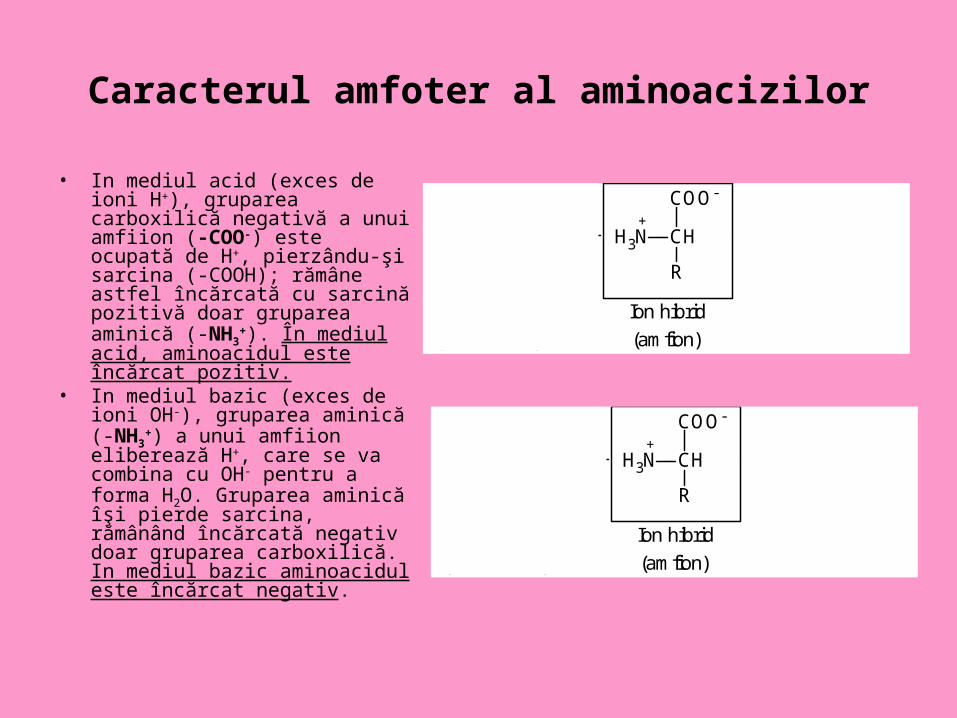
Caracterul amfoter al aminoacizilor
• In mediul acid (exces de ioni H+), gruparea carboxilică negativă a unui amfiion (-COO-) este ocupată de H+, pierzându-şi sarcina (-COOH); rămâne astfel încărcată cu sarcină pozitivă doar gruparea aminică (-NH3
+). În mediul acid, aminoacidul este încărcat pozitiv.
• In mediul bazic (exces de ioni OH-), gruparea aminică (-NH3
+) a unui amfiion eliberează H+, care se va combina cu OH- pentru a forma H2O. Gruparea aminică îşi pierde sarcina, rămânând încărcată negativ doar gruparea carboxilică. In mediul bazic aminoacidul este încărcat negativ.
(mediu bazic)(amfion)(mediu acid)
AnionIon hibridCation
OH-+H++
H2O
COO-
CH
R
H2N+
COO-
CH
R
H3N+
COOH
CH
R
H3N
(mediu bazic)(amfion)(mediu acid)
AnionIon hibridCation
OH-+H++
H2O
COO-
CH
R
H2N+
COO-
CH
R
H3N+
COOH
CH
R
H3N
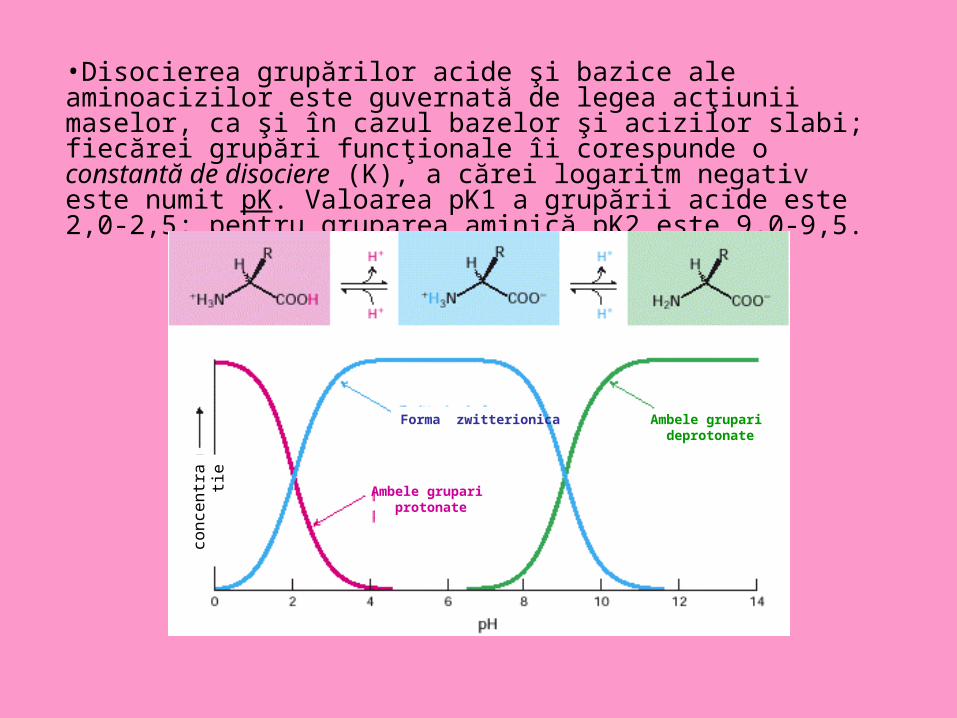
•Disocierea grupărilor acide şi bazice ale aminoacizilor este guvernată de legea acţiunii maselor, ca şi în cazul bazelor şi acizilor slabi; fiecărei grupări funcţionale îi corespunde o constantă de disociere (K), a cărei logaritm negativ este numit pK. Valoarea pK1 a grupării acide este 2,0-2,5; pentru gruparea aminică pK2 este 9,0-9,5.
Forma zwitterionica
Ambele grupari protonate
Ambele grupari deprotonate
con
cen
tra
tie

Punctul izoelectric al aminoacizilor
• Aminoacizii dizolvaţi în medii ionizate prezintă fenomenul de migrare în câmp electric, fenomen ce poartă numele de ionoforeză.
• În soluţii acide, moleculele ionizate vor migra spre catod (-), iar în soluţii alcaline spre anod (+).
• Pentru fiecare dintre aminoacizi există o valoare de pH, numit pH izoelectric sau punct izoelectric (notat pI), la care molecula este neutră, iar fenomenul de migrare electroforetică încetează. La punctul izoelectric, numărul sarcinilor pozitive şi negative sunt egale (aminoacidul este deci electroneutru), disocierea lui ca acid fiind egală cu disocierea ca bază.
• În medii cu pH mai mic decât pI, aminoacidul se va comporta ca bază, iar la pH mai mare decât pI el se va comporta ca acid. Astfel, aminoacizii pot îndeplini funcţia de tampon acido-bazic în organism, proprietate manifestată mai pregnant în jurul valorii pK1 şi pK2.
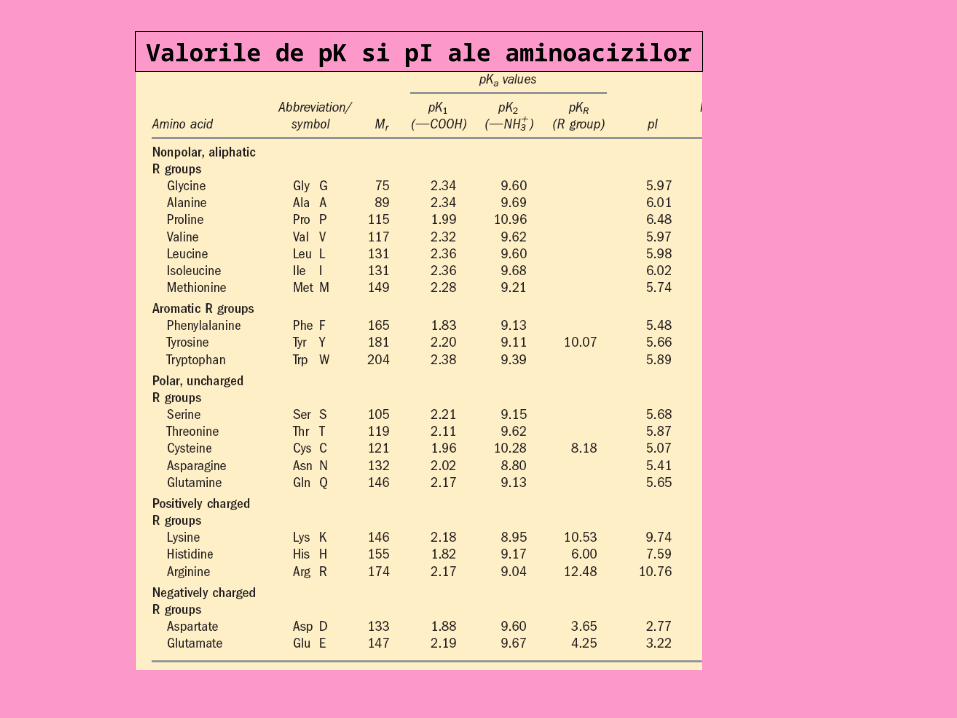
Valorile de pK si pI ale aminoacizilor
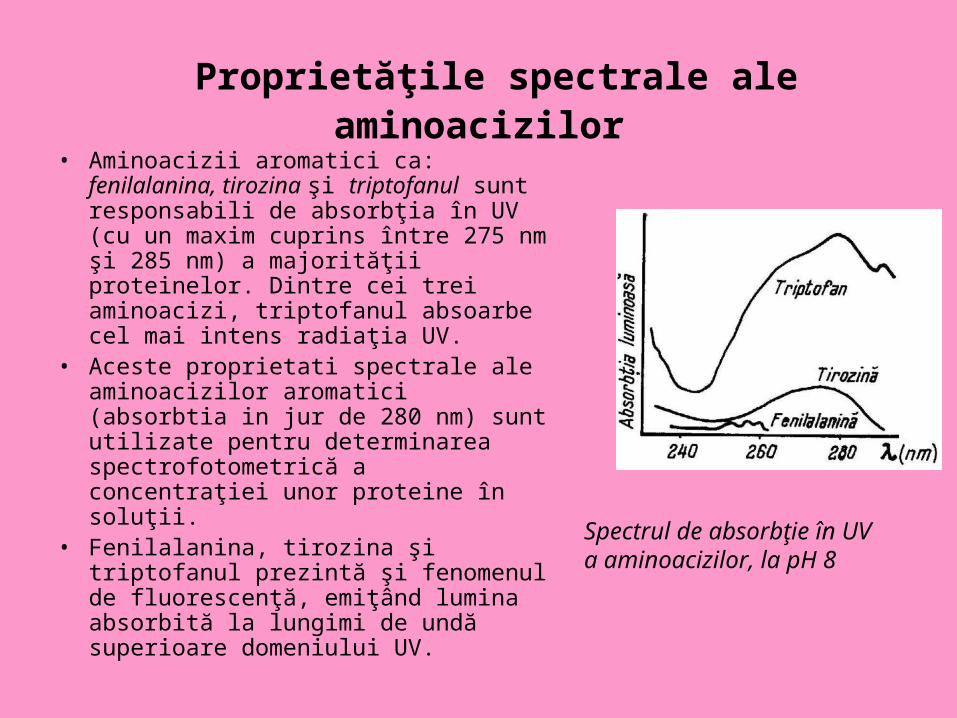
Proprietăţile spectrale ale aminoacizilor
• Aminoacizii aromatici ca: fenilalanina, tirozina şi triptofanul sunt responsabili de absorbţia în UV (cu un maxim cuprins între 275 nm şi 285 nm) a majorităţii proteinelor. Dintre cei trei aminoacizi, triptofanul absoarbe cel mai intens radiaţia UV.
• Aceste proprietati spectrale ale aminoacizilor aromatici (absorbtia in jur de 280 nm) sunt utilizate pentru determinarea spectrofotometrică a concentraţiei unor proteine în soluţii.
• Fenilalanina, tirozina şi triptofanul prezintă şi fenomenul de fluorescenţă, emiţând lumina absorbită la lungimi de undă superioare domeniului UV.
Spectrul de absorbţie în UV a aminoacizilor, la pH 8

Aminoacizi si Proteine
Aminoacizi Structura, clasificare,proprietati fizico-chimice
Legatura peptidicaAlte tipuri de legaturi chimice din moleculele proteiceProteine- nivele de organizare
Structura primarăStructura secundarăStructura terţiarăStructura cuaternară

Legatura peptidica
• Legătura peptidică este legătura formată in urma reactiei de condensare între gruparea carboxilică (-COOH) de la C1 a unui aminoacid şi gruparea aminică (-NH2) de la C2 a altui aminoacid, cu eliminarea unei molecule de apă.
• Gruparea -CO-NH-, caracteristică legăturii peptidice, alternând cu grupări metinice formează "coloana vertebrală“ (din engl. backbone)
a catenei polipeptidice.
C COOHH2N
R
-H2OC CO NH C COOH
R R
H H H
1 22
H2N
2
H
R
H2N COOHC
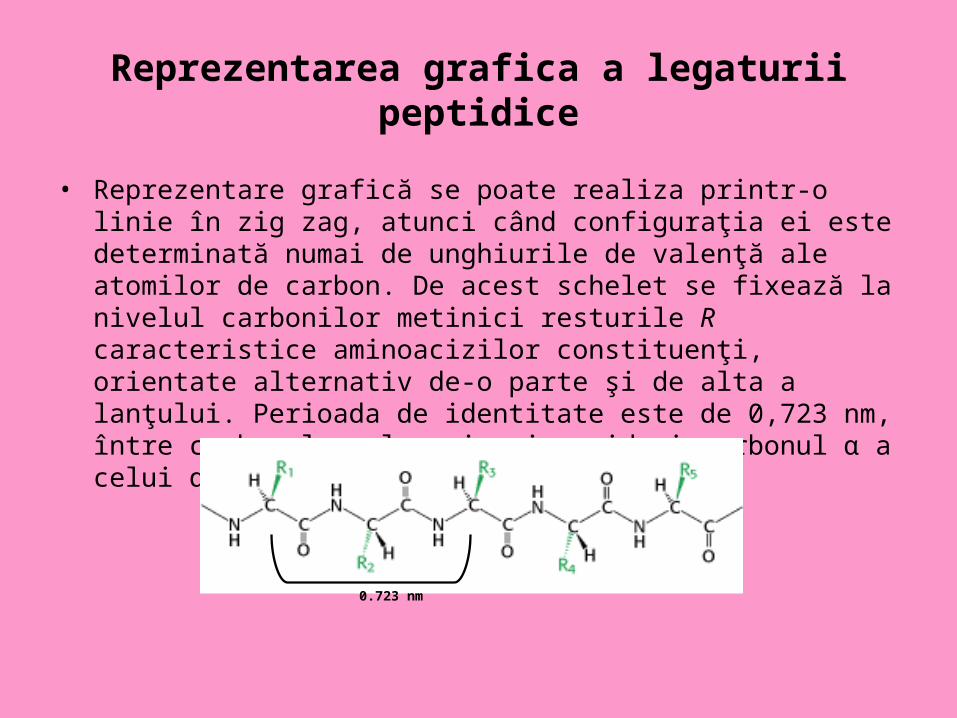
Reprezentarea grafica a legaturii peptidice
• Reprezentare grafică se poate realiza printr-o linie în zig zag, atunci când configuraţia ei este determinată numai de unghiurile de valenţă ale atomilor de carbon. De acest schelet se fixează la nivelul carbonilor metinici resturile R caracteristice aminoacizilor constituenţi, orientate alternativ de-o parte şi de alta a lanţului. Perioada de identitate este de 0,723 nm, între carbonul α al unui aminoacid şi carbonul α a celui de al treilea aminoacid
0.723 nm

Formarea şi geometria legăturii peptidice • Mezomeria lactam-lactimică se datoreaza oscilarii dublei legături
dintre atomul de carbon şi oxigen, la atomul de carbon şi azot, datorită transportului alternativ al unui electron între NH şi CO.
• Rolul mezomeriei lactam-lactimica: determina structura peptidelor şi proteinelor.
O
N
H
C
N
H
O
C+
-
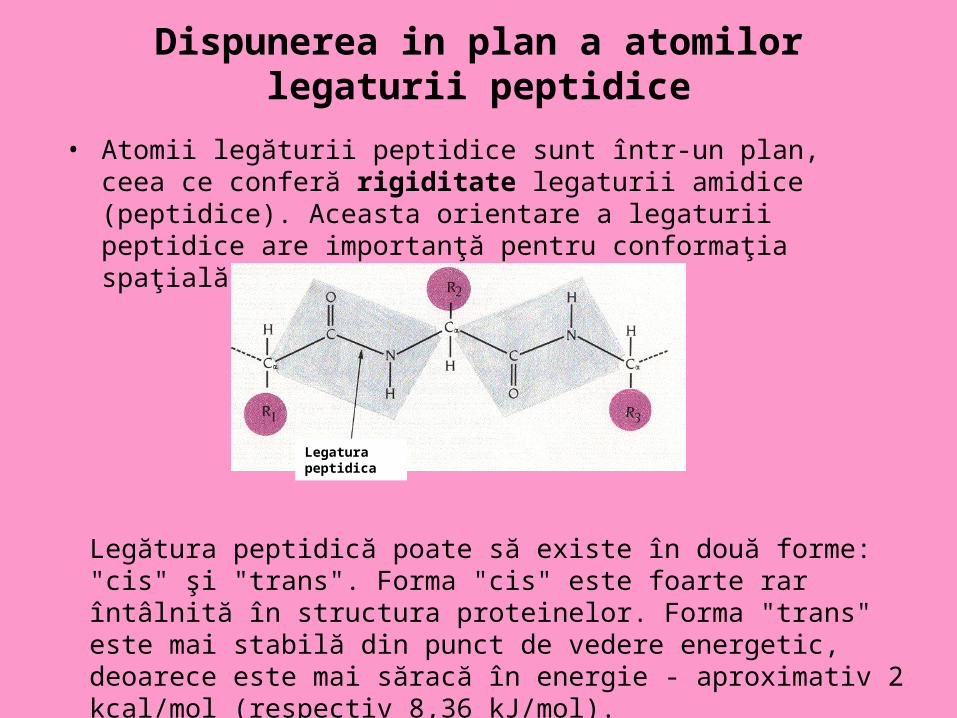
Dispunerea in plan a atomilor legaturii peptidice
• Atomii legăturii peptidice sunt într-un plan, ceea ce conferă rigiditate legaturii amidice (peptidice). Aceasta orientare a legaturii peptidice are importanţă pentru conformaţia spaţială a catenei polipeptidice.
Legatura peptidica
Legătura peptidică poate să existe în două forme: "cis" şi "trans". Forma "cis" este foarte rar întâlnită în structura proteinelor. Forma "trans" este mai stabilă din punct de vedere energetic, deoarece este mai săracă în energie - aproximativ 2 kcal/mol (respectiv 8,36 kJ/mol).
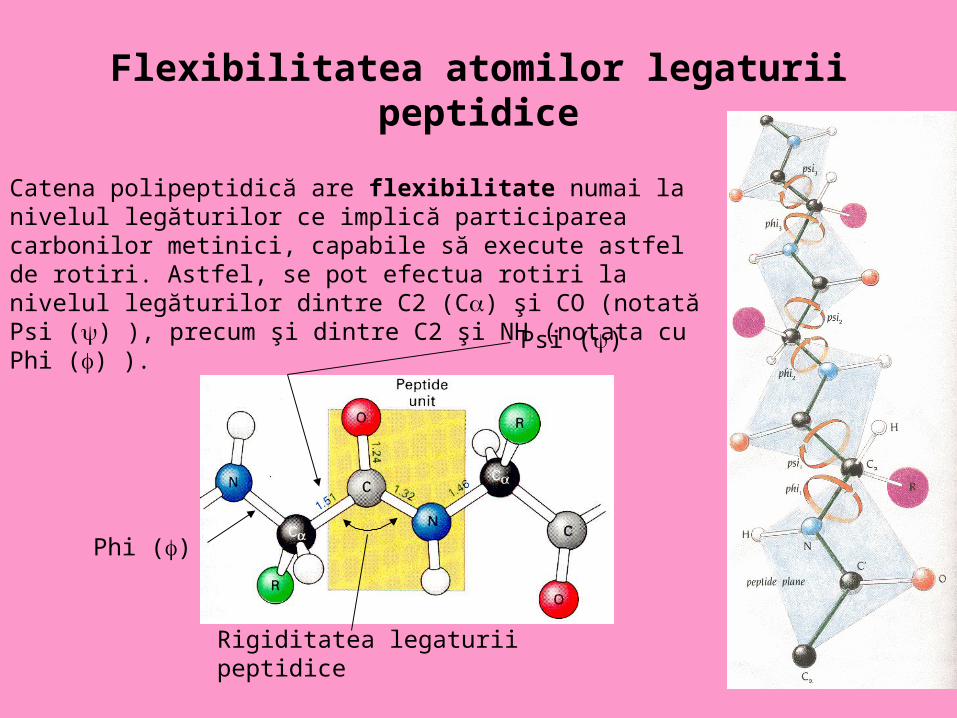
Flexibilitatea atomilor legaturii peptidice
Psi ()
Phi ()
Rigiditatea legaturii peptidice
Catena polipeptidică are flexibilitate numai la nivelul legăturilor ce implică participarea carbonilor metinici, capabile să execute astfel de rotiri. Astfel, se pot efectua rotiri la nivelul legăturilor dintre C2 (C) şi CO (notată Psi () ), precum şi dintre C2 şi NH (notata cu Phi () ).

Diagrama Ramachandran
• Conform acestei diagrame (contine 4 cadrane), atomii nelegati chimic din structura aminoacizilor adiacenti se pot apropia intre ei pana la o distanta maxima egala cu suma razelor van der Waals.
• Pentru un numar mare de proteine se calculeaza diagrama Ramachandran care arata in 3 cadrane combinatiile posibile ale celor doua unghiuri Psi () ), si Phi () ). Pentru fiecare aminoacid se obtin zone de maxima densitate notate cu α, β si L.
α
β
L

Aminoacizi si Proteine
Aminoacizi Structura, clasificare proprietati fizico-chimice
Legatura peptidicaAlte tipuri de legaturi chimice din moleculele proteiceProteine- nivele de organizare
Structura primarăStructura secundarăStructura terţiarăStructura cuaternară

Alte tipuri de legaturi chimice din moleculele proteice
- Puntea de hidrogen (numită şi legătura de hidrogen) se poate realiza în situaţia în care un atom de hidrogen este amplasat între doi atomi, ambii electronegativi. De exemplu, in catena polipeptidică, potenţialul de donare de hidrogen este reprezentat de grupările NH, iar cel de acceptare de către grupările C=O ale aceleiaşi catene, sau a unor catene învecinate. Cu alte cuvinte, puntea de hidrogen ar reprezenta o acţiune de schimb dintre atomul de azot şi cel de oxigen, a hidrogenului aparţinând grupărilor NH şi CO.
Puntea de hidrogen
•Puntea de H este mai slabă decât o legătură covalentă, având doar 1/10 din energia de legare; energia (entalpia de formare) unei legături de hidrogen este 2,39 kcal/mol (10 kJ/mol). Distanţa dintre atomul de azot şi cel de oxigen nu trebuie să depăşească 0.28 nm, pentru ca între cei doi atomi să se poată stabili o legătură de hidrogen.
Donor de Hidrogen
Acceptor de Hidrogen

Alte legaturi din moleculele proteice
• se formează între două grupări sulfhidrice aparţinând la două resturi de cisteină, prin dehidrogenare. Această legătură covalentă foarte puternică, având energia de legare de 50-100 kcal/mol (respectiv 208-418 kJ/mol), este rezistentă la hidroliza proteinei.
Puntea disulfurică
cisteina
cisteina
cistina

Alte legaturi din moleculele proteice- Legătura ionică se realizeaza între o grupare -COOH
liberă a unui aminoacid dicarboxilic dintr-o catenă polipeptidică şi o grupare -NH2 liberă a unui aminoacid diaminic din aceeaşi catenă sau dintr-o catenă învecinată. Formarea şi menţinerea legăturilor ionice depind de pH-ul mediului. Energia de atracţie dintre cele două grupări încărcate cu sarcini de semn opus (NH3
+ şi COO-), realizând o aşa-zisă punte salină, poate ajunge la cel mult -10 kcal/mol (respectiv -42 kJ/mol), la o distanţă a sarcinilor egală cu 0,3 nm. Această energie este de zece ori mai mare decât cea a forţelor van der Waals.
- Legătura eterică se realizează prin eliminarea unei molecule de apă dintre două grupări alcoolice aparţinând unor aminoacizi ca serina din catena polipeptidică.

Alte legaturi din moleculele proteice
- Forţele van der Waals. Între resturile aminoacizilor din catena polipeptidică se pot stabili legături apolare prin forţe van der Waals. Aceste forţe sunt de natura unor atracţii electrostatice şi sunt invers proporţionale cu puterea a 6-a a distanţei dintre atomi. Ele acţionează numai la distanţe foarte mici.
- Efect al forţelor van der Waals sunt aşa numitele "forţe hidrofobe" cu care catenele laterale ale aminoacizilor din lanţurile peptidice se atrag reciproc, orientându-se adesea spre interiorul macromoleculei proteice şi împiedică pătrunderea apei în anumite spaţii ale edificiului proteic. Forţele hidrofobe influenţează puternic structura spaţială a moleculelor proteice în soluţii apoase.
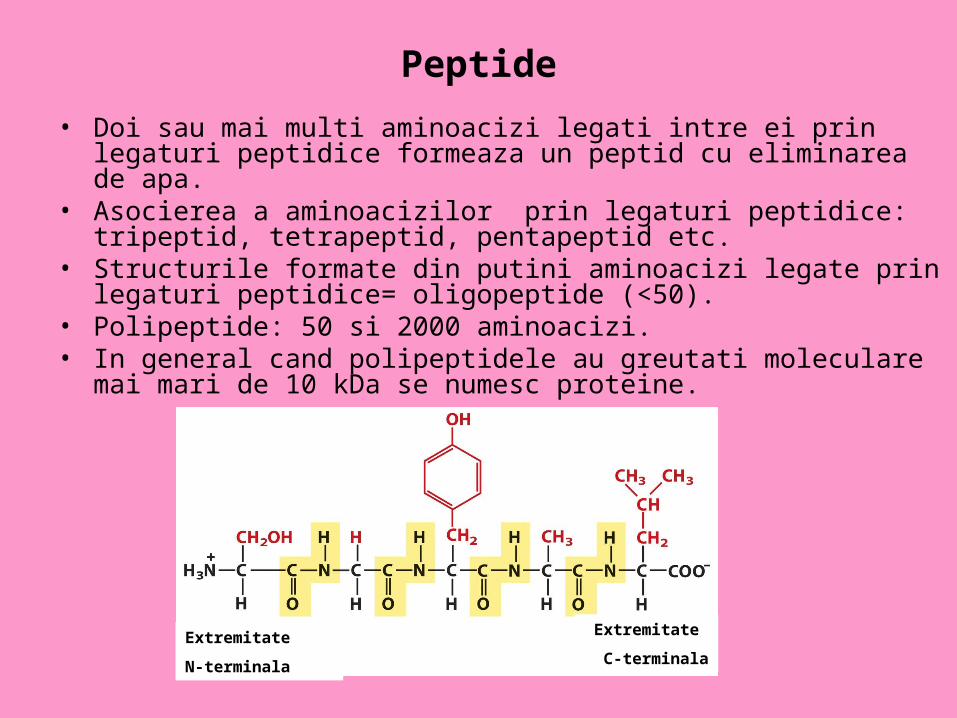
Peptide
• Doi sau mai multi aminoacizi legati intre ei prin legaturi peptidice formeaza un peptid cu eliminarea de apa.
• Asocierea a aminoacizilor prin legaturi peptidice: tripeptid, tetrapeptid, pentapeptid etc.
• Structurile formate din putini aminoacizi legate prin legaturi peptidice= oligopeptide (<50).
• Polipeptide: 50 si 2000 aminoacizi.• In general cand polipeptidele au greutati moleculare mai mari de 10 kDa
se numesc proteine.
Extremitate
N-terminala
Extremitate
C-terminala

Daltonul
• Daltonul (Da) este o unitate de masă, apropiată cu cea a atomului de hidrogen (mai precis, este egală cu 1,0 pe scala maselor atomice). A fost numită după John Dalton (1766-1844), care a dezvoltat teoria atomică a materiei. Un kilodalton (1 kDa) este egal cu 1.000 daltoni. Noţiunea de dalton este utilizată în biochimie pentru a exprima masa moleculară a macromoleculelor (proteine, acizi nucleici).
• Masa moleculara medie a unui aminoacid este de aprox. 110 Da.

Aminoacizi si Proteine
Aminoacizi Structura, clasificare, proprietati fizico-chimice
Legatura peptidicaAlte tipuri de legaturi chimice din moleculele proteiceProteine- nivele de organizare
Structura primarăStructura secundarăStructura terţiarăStructura cuaternară
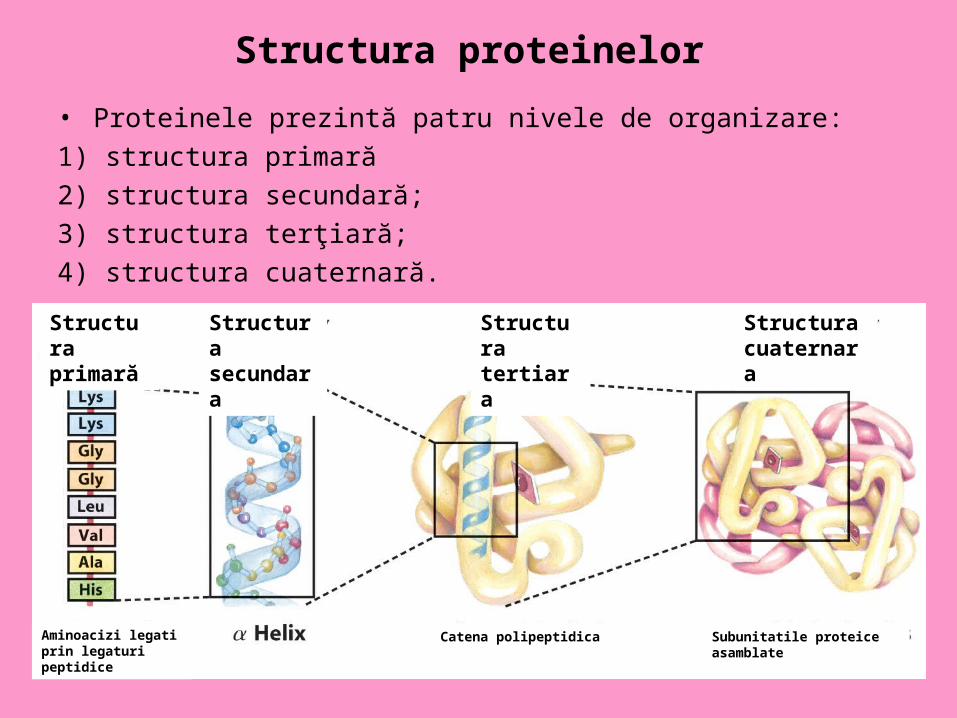
Structura proteinelor • Proteinele prezintă patru nivele de organizare:
1) structura primară
2) structura secundară;
3) structura terţiară;
4) structura cuaternară.
Structura primară
Aminoacizi legati prin legaturi peptidice
Structura secundara
Structura tertiara
Structura cuaternara
Catena polipeptidica Subunitatile proteice asamblate

Nivele de organizare structurala ale
proteinelor
• Elementele structurii secundare se combina si formeaza structura tertiara
• Structura cuaternara este caracteristica complexelor enzimatice.– Multe proteine sunt
active in stare de homodimeri, homo/ heterotetrameri, etc
Structura proteica primara= Succesiunea aminoacizilor din structura proteinei
aminoacizi
Structuri β α helix
Structura proteica secundara determinata de legaturi de H intre aminoacizi ai lantului polipeptidic
Structuri β
α helix
Structura proteica tertiara cand au loc diferite interactiuni (legaturi) intre elementele structurii secundare
Structura cuaternara care este prezenta cand proteina contine mai multe catene polipeptidice

Aminoacizi si Proteine
Aminoacizi Structura, clasificare proprietati fizico-chimice
Legatura peptidicaAlte tipuri de legaturi chimice din moleculele proteiceProteine- nivele de organizare
Structura primarăStructura secundarăStructura terţiarăStructura cuaternară

Structura primară a proteinelor • Structura primară este definită ca fiind
succesiunea (secvenţa) aminoacizilor în lanţul polipeptidic. Această structură va determina în mod esenţial celelalte planuri de organizare a macromoleculei proteice.
• Prima secvenţă cunoscută a fost cea a insulinei, un polipeptid alcătuit din 51 aminoacizi; a fost realizată de Sanger în 1953.
• Lanţul polipeptidic, înşiruire de aminoacizi legaţi între ei prin legătură peptidică, are la o extremitate o grupare -NH2 liberă (extremitatea amino-terminală sau N-terminală), iar la extremitatea cealaltă o grupare -COOH liberă (extremitatea carboxi-terminală sau C-terminală).
• Resturile R libere ale aminoacizilor din lanţul polipeptidic pot detine grupari functionale care contribuie în ansamblu atât la stabilirea structurii proteice, cât şi la realizarea interacţiunilor specifice dintre proteine sau dintre proteine şi alţi compuşi organici (acizi nucleici, substratul enzimatic, etc).

Denumirea unui lant oligo/polipeptidic
• Prin convenţie, ordinea de prezentare a secvenţei începe întotdeauna cu aminoacidul care are gruparea aminică liberă (care devine astfel aminoacidul nr.1) şi se termină cu acela care are gruparea carboxilică liberă (care devine aminoacidul n). Altfel spus, prezentarea unei secvenţe începe cu extremitatea N-terminală.
• Denumirea unui polipeptid se face enumerând radicalii aminoacizilor în ordinea secvenţei, începând cu cel de la extremitatea N-terminală; în această enumerare vor avea sufixul "-il" toţi aminoacizii a căror carboxil este legat peptidic, cu excepţia aminoacidului C-terminal care îşi va păstra terminaţia neschimbată.

Siclemia sau anemia falciforma (anemia cu hematii in forma de secera)
• Activitatea biologică a proteinelor depinde de particularităţile lor structurale. Modificarea secvenţei, înlocuirea, pierderea (deleţia) sau introducerea (inserţia) unor aminoacizi poate duce la modificarea activităţii biologice. Bettelheim & March (1990) Introduction to Organic & Biochemistry
(International Edition) Philadelphia: Saunders College Publishing, p301

Aminoacizi si Proteine
Aminoacizi Structura, clasificare proprietati fizico-chimice
Legatura peptidicaAlte tipuri de legaturi chimice din moleculele proteiceProteine- nivele de organizare
Structura primarăStructura secundarăStructura terţiarăStructura cuaternară

Structura secundară a proteinelor
• Structura secundară este definită de poziţiile spaţiale pe care le au resturile R ale aminoacizilor, unul faţă de altul, de-a lungul aceleiaşi catene polipeptidice.
• Poate un lant polipeptidic sa se plieze in mod regulat intr-o structura repetitiva?
• In 1951, Linus Pauling si Robert Corey- 2 structuri repetitive: α helix si structura in foaie β pliata.
• Ulterior, s-au evidentiat alte structuri nerepetitive: structuri in “agrafa de par” (engl. "β-turn" sau "hairpin turn"), bucle (engl. loop), regiuni de intoarcere (engl. turn).
• Toate aceste structuri repetitive si nerepetitive contribuie la formarea nivelului de organizare secundar al proteinei.
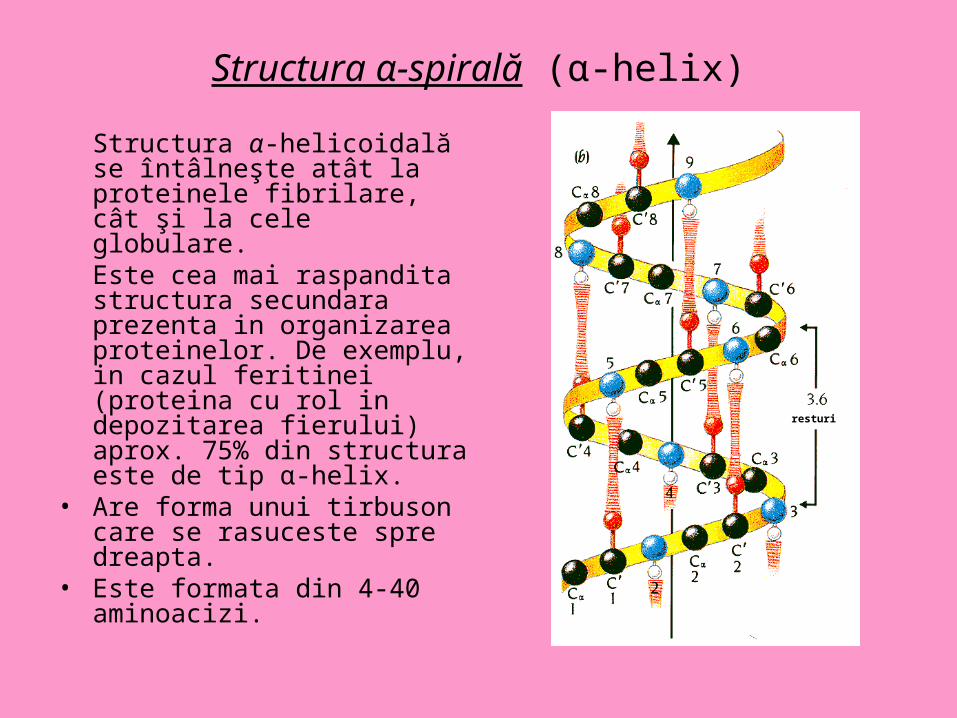
Structura α-spirală (α-helix)
Structura α-helicoidală se întâlneşte atât la proteinele fibrilare, cât şi la cele globulare. Este cea mai raspandita structura secundara prezenta in organizarea proteinelor. De exemplu, in cazul feritinei (proteina cu rol in depozitarea fierului) aprox. 75% din structura este de tip α-helix.
• Are forma unui tirbuson care se rasuceste spre dreapta.
• Este formata din 4-40 aminoacizi.
resturi

Proprietatile α-helixului
Catena polipeptidică este înfăşurată pe o spirală. Fiecare spiră conţine 3,6 aminoacizi, iar distanţa dintre spire (perioada mică de identitate) este de 0.54 nm, unui aminoacid revenindu-i 0.15 nm în pasul helixului. Gruparea CO a fiecarui aminoacid formeaza punti de hidrogen cu hidrogenul grupării NH a celui de al 4-lea aminoacid, care urmeaza in structura lantului dupa aminoacidul cu gruparea CO. Gruparea CO a unui aminoacid va forma punti de H la o depărtare de 0,28 nm, distanţă ce asigură realizarea unei punţi de hidrogen. Energia legăturilor de hidrogen intracatenare asigură o deosebită stabilitate structurii helicoidale a catenei polipeptidice.

Stabilitatea α-helixului
• Stabilitatea helixului poate fi influentata de compozitia structurii primare a proteinei.
• Anumiti aminoacizi favorizeaza structura spiralata: Ala, Glu, Leu, Met. Alti aminoacizi intrerup structura helicala: Pro, Gly, Tyr, Ser.
• Stabilitatea helixului este mult micşorată în stare dizolvată a proteinei. Acidul poliglutamic, de natură sintetică, are înfăţişare α-helix în mediul acid, în timp ce în mediul bazic, datorită ionizării grupărilor carboxilice laterale, respingerea lor electrostatică rupe legăturile de hidrogen şi molecula capătă o înfăşurare întâmplătoare (din engl. "random coil").

Structura în “foaie pliată” (structura β)
• Structura în “foaie pliată” (structura β) este asigurată tot prin punţi de hidrogen realizate între mai multe regiuni dintr-o catena polipeptidica avand orientare paralelă sau antiparalelă.
• Distanţa între 2 aminoacizi adiacenti este de 0.35 nm.
• În esenţă, modelul poate fi reprezentat ca o foaie de hârtie pe care s-au desenat catene polipeptidice paralele şi apoi pliată alternativ în sus şi în jos, în dreptul carbonilor alfa. Catenele laterale R ale aminoacizilor sunt aşezate alternativ, de-o parte şi de alta a foii pliate, perpendicular pe crestele cutelor.
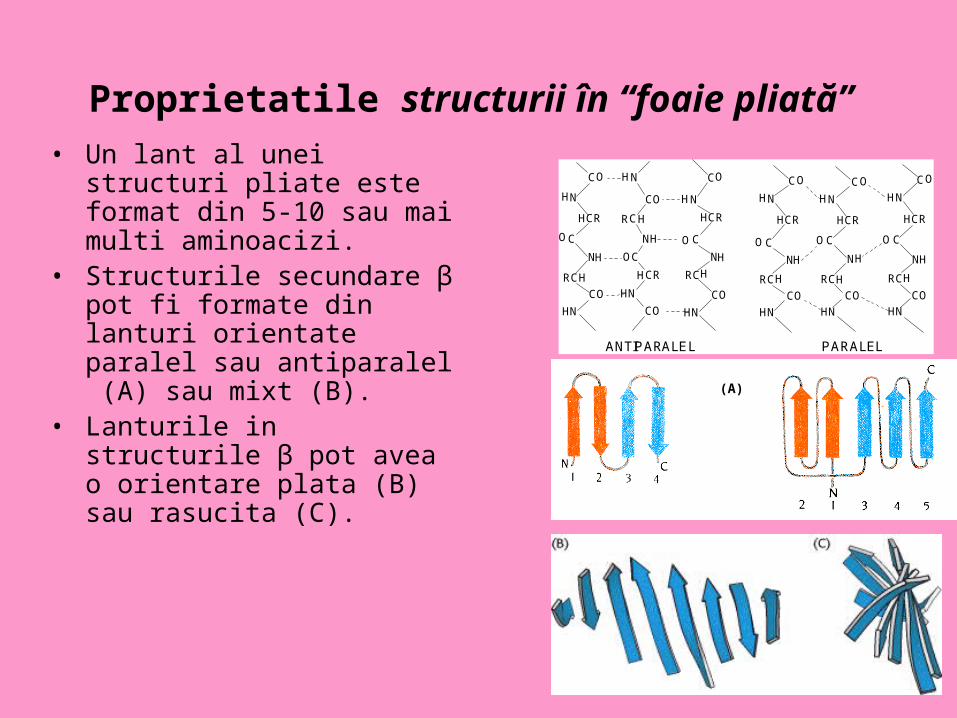
Proprietatile structurii în “foaie pliată” • Un lant al unei structuri pliate
este format din 5-10 sau mai multi aminoacizi.
• Structurile secundare β pot fi formate din lanturi orientate paralel sau antiparalel (A) sau mixt (B).
• Lanturile in structurile β pot avea o orientare plata (B) sau rasucita (C).
N
C
RC
HN
C
HCR
N
CO
O
H
O
H
NH
CO
C
NH
OC
CR
HN
CO
R
H
H
H
H
O
H
O
CO
N
HCR
C
HN
RC
C
N
H
H
O
H
O
CO
N
HCR
C
HN
RC
C
N
H
H
O
H
O
CO
N
HCR
C
HN
RC
C
N N
C
RC
HN
C
HCR
N
CO
O
H
H
HH
O
ANTIPARALEL PARALEL
(A)
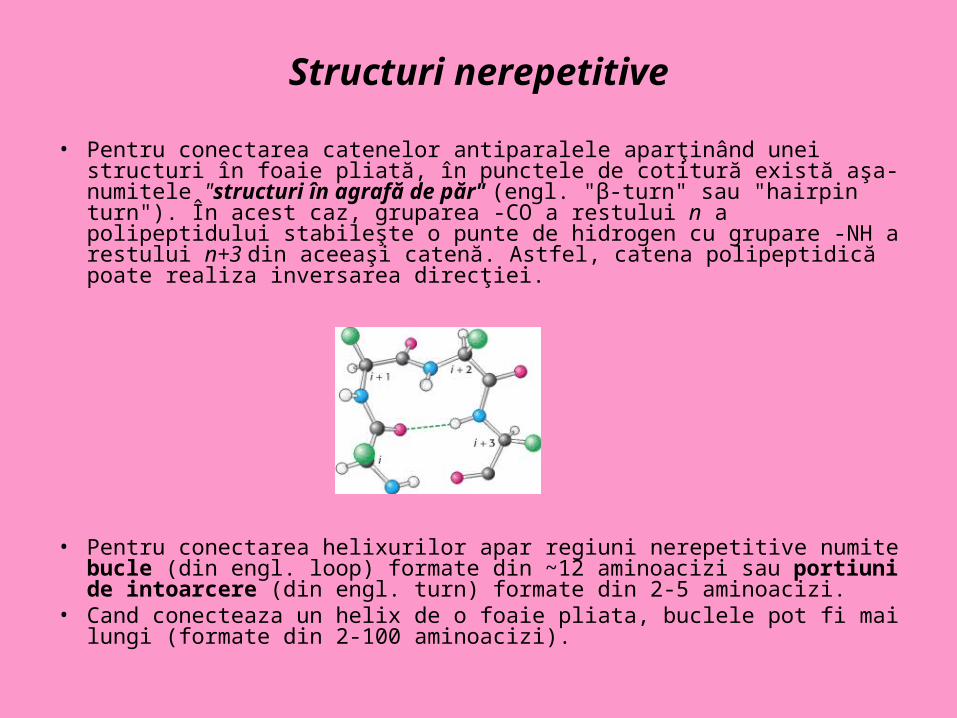
Structuri nerepetitive
• Pentru conectarea catenelor antiparalele aparţinând unei structuri în foaie pliată, în punctele de cotitură există aşa-numitele "structuri în agrafă de păr" (engl. "β-turn" sau "hairpin turn"). În acest caz, gruparea -CO a restului n a polipeptidului stabileşte o punte de hidrogen cu grupare -NH a restului n+3 din aceeaşi catenă. Astfel, catena polipeptidică poate realiza inversarea direcţiei.
• Pentru conectarea helixurilor apar regiuni nerepetitive numite bucle (din engl. loop) formate din ~12 aminoacizi sau portiuni de intoarcere (din engl. turn) formate din 2-5 aminoacizi.
• Cand conecteaza un helix de o foaie pliata, buclele pot fi mai lungi (formate din 2-100 aminoacizi).

Aminoacizi si Proteine
Aminoacizi Structura, clasificare proprietati fizico-chimice
Legatura peptidicaAlte tipuri de legaturi chimice din moleculele proteiceProteine- nivele de organizare
Structura primarăStructura secundarăStructura terţiarăStructura cuaternară

Structura terţiară a proteinelor
• Structura terţiară se referă la atitudinea spaţială pe care o au, una faţă de alta, diferitele regiuni ale aceleiaşi catene polipeptidice. Această structură variază foarte mult de la o proteină la alta.
• Cristalografia cu raze X sau rezonanta magnetica nucleara au pus in evidenta structura tridimensionala a mii de proteine.

Mioglobina- exemplu de organizare tridimensionala
Mioglobina constă în mare măsură din structuri α-helix. Grupul hem specific este reprezentat în negru iar atomul de fier printr-o sferă.
Resturi de aminoacizi hidrofili la exterior si resturi de aminoacizi hidrofobi la interior
1958, John Kendrew
1962 -Premiul Nobel in chimie- John Kendrew si Max Perutz – pentru caracterizarea structurii 3D a unor globuline (hemoglobina).

Porina - exemplu de organizare tridimensionala
Resturi de aminoacizi hidrofili la interior si resturi de aminoacizi hidrofobi la exterior
Exterior preponderent hidrofob
Canal hidrofilic la interior

Factorii de care depinde structura terţiară
• 1) structura primară a proteinei; • 2) prezenţa grupărilor -SH ale cisteinei;
• 3) natura resturilor aminoacizilor din catena polipeptidică.

1) Structura primară a proteinei
- Prezenţa prolinei, aminoacid care nu conţine o grupare aminică liberă, va altera structura unui α -helix, prin întreruperea structurii regulate şi modificarea orientării spaţiale a restului catenei.

2) Prezenţa grupărilor -SH ale cisteinei
- Între grupările -SH aparţinând la două resturi de cisteină situate în zone diferite ale catenei se pot stabili punţi disulfurice (-S-S-), ce obligă catena polipeptidică să se plieze şi să adopte o anumită conformaţie spaţială. Rolul determinant al structurii terţiare a fost prima dată demonstrat pe ribonucleaza pancreatică bovină, enzimă ce are în stare nativă 4 punţi disulfurice intracatenare.

3) Natura resturilor aminoacizilor din catena polipeptidică
- Resturile R hidrofile ale unor aminoacizi ca: Ser, Thr, Tyr, Asn, Gln, Asp, Glu, Lys, Arg şi His, se orientează spre exterior, pentru a lega moleculele de apă, în timp ce resturile R hidrofobe ale aminoacizilor Gly, Ala, Val Leu, Ile, Phe, Cys, Met şi Trp, se vor orienta spre interior, obligând catena polipeptidică să adopte o conformaţie de ghem.
• Structura terţiară a proteinei tinde spre aranjamentul spaţial cel mai favorabil din punct de vedere energetic (conformaţia de minimă energie)=aranjamentul spatial cel mai stabil.
Bettelheim & March (1990) Introduction to Organic & Biochemistry (International Edition) Philadelphia: Saunders College Publishing, p302
Regiune hidrofoba

Structuri suprasecundare si domenii
• Asigura edificarea structurii tertiare• Structuri suprasecundare =motiv• Motive= O substructură regulată, cu proprietăţi distincte, specifice,
ce apare în diferite domenii; o combinaţie simplă, specifică, de structuri secundare.
• Domeniile= motive cu identitate structurala si functionala din structura proteinelor. Domeniile pot avea o functionalitate distincta in cadrul catenei polipeptidice (pot functiona ca o unitate proteica distincta). Un domeniu poate conţine între 30 şi 400 de aminoacizi.
• In unele cazuri, domeniile si motivele se suprapun (nu exista inca in literatura de specialitate o distinctie clara intre domenii si motive).

Clasificarea structurală a proteinelor
Baza de date: The Structural Classification of Proteins (SCOP) (http://scop.mrc-lmb.cam.ac.uk/scop) - pe baza structurilor suprasecundare si a motivelor -4 clase de proteine
Clasa Caracteristici Exemple
α- helix
“all α”
Structuri secundare exclusiv sau aproape numai α- helix
Albumina serica (om), feritina, mioglobina
Foi β pliate “all β” Structuri secundare exclusiv sau aproape numai Foi β pliate
Colagenaza-3 (om)
UDP N-acetilglucosamin aciltransferaza (Escherichia coli)
α+ β α- helixuri si foi β pliate separat distribuite in molecula
Triozfosfat izomeraza (muschi- puiul de gaina),
Proteine fluorescente: green fluorescent protein GFP meduza (Aequorea victoria)
α/ β Helixuri si foi asamblate in unitati βαβ
Ribonucleaza A (pancreasul bovin)

Motive
• Clasificare: simple: formate din mai multe structuri sunt de tipul: αα, ββ, βαβ. complexe: formate din succesiuni numai de structuri α (all α), din
succesiuni numai de structuri β (all β), din structuri α+β, α/β

Motive simple
Motivul αα sau helix-bucla-helix
Este alcatuit din doua helixuri unite printr-o portiune nehelicala.
Exemple: in structura proteinelor care interactioneaza cu ADN-ul, a proteinelor care leaga ionii de calciu: parvalbumina din muschi, calmodulina, troponina C.
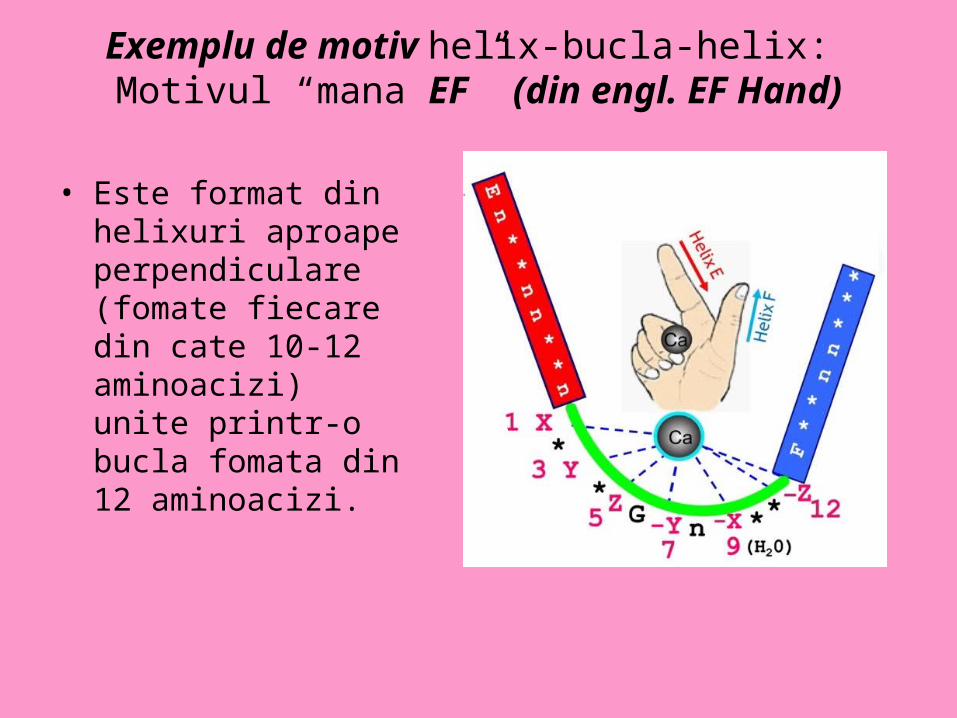
Exemplu de motiv helix-bucla-helix: Motivul “mana EF” (din engl. EF Hand)
• Este format din helixuri aproape perpendiculare (fomate fiecare din cate 10-12 aminoacizi) unite printr-o bucla fomata din 12 aminoacizi.

Exemplu de motiv helix-bucla-helix: Motivul “mana EF” (din engl. EF Hand)
Calmodulina cu rol in reglarea Ca-ului celular, modularea unor enzime si a unor proteine de transport
• Este prezent la proteinele care leaga Ca2+
• La nivelul unui astfel de motiv se gaseste un singur situs de legare a Ca2+ . Ionii de Ca se leaga de bucla de obicei prin intermediul a doua grupari carboxil provenite de la un rest de acid aspartic si respectiv acid glutamic, de o grupare carbonil din regiunea de bucla si de o molecula de apa.
• Dupa anumiti autori “motivul mana EF” este considerat un domeniu functional.

•Este alcatuit din 2 a helixuri si o portiune de intoarcere (din engl. turn) formata dintr-un lant scurt de aminoacizi • Extremitatile C-terminale ale helixurilor interactioneaza cu ADN-ul la nivelul adanciturii mari a ADN-ului. Este specific proteinelor de legare a ADN (proteine reglatoare): represorul bacteriofagului λ, represorul triptofanului, proteina activata de catabolit (din engl. catabolite activator protein (CAP))
Motivul helix-intoarcere-helix (din engl. Helix Turn Helix)

Motivul fermoarului de leucina (din engl. Leucine zipper)
Acest motiv este format din 2 helixuri cu portiuni amfipatice care prezinta aminoacizii hidrofobi dispusi la nivelul suprafetei de contact dintre helixuri. In aceasta zona, cele doua regiuni polipeptidice formeaza o regiune de dimer. La nivelul suprafetei hidrofobe fiecare al 7-lea rest de aminoacid este leucina. Initial s-a crezut ca resturile de leucina din fiecare helix se intercaleaza cu resturile de leucina din cel de-al doilea helix formand un fermoar de leucina la nivelul suprafetei hidrofobe. Ulterior s-a constatat ca resturile de leucina se dispun unul dupa altul in urma rasucirii celor doua helixuri unul in jurul celuilalt. Sunt caracteristice proteinelor reglatoare ale ADN-ului (factori de transcriere). Motivul “leucine zipper” este prezent atat la proteine eucariote cat si cele procariote.

Motivul ββ sau agrafa sau cotul β
• Este constituit din doua structuri de tip β legate printr-o intoarcere formata din 2-5 aminoacizi.
• O varianta a acestui motiv este motivul “cheie greceasca”. Este intalnit la nucleaza de la genul Staphylococus, in structura plastocianinei (o proteina din lantul transportor de electroni de la organismele fotosintetizante), in structura anticorpilor.

Motivul “cheie greceasca”
• Denumirea provine de la motivele decorative folosite in Grecia antica in constructii si imbracaminte.

Motivul “butoiul β”
• Motivul “butoiul β” (din engl. β barrel) este format din foi β torsionate ca si doagele unui butoi formand o structura cilindrica. Sunt caracteristice proteinelor membranare in care 20 sau mai multe regiuni transmembranare formeaza foi β organizate intr-un cilindru. Regiunile de aminoacizi hidrofobe ale foilor β interactioneaza cu regiunile hidrofobe ale lipidelor membranare.
• Exemplu: porinele prezente in membrana externa a bacteriilor Gram negative si care permit trecerea substantelor polare prin porul creat in membrana externa, α hemolizinele de la Staphylococus aureus care penetreaza celulele animale.
Vedere de sus a motivului” β barrel”

Motivele
Macrostructura de stangaprezenta de exemplu la enzima proteolitica subtilizina de la Bacilus subtilis
Realizeaza jonctiunea intre doua foi paralele prin intermediul unui α helix. Cel mai raspandit motiv βαβ este “macrostructura”de dreapta fiind prezent in toate proteinele care poseda foi β paralele. Helixul este paralel cu foile β si mascheaza regiunile cu aminoacizi hidrofobi ale acestora. Regiunile de bucla au lungimi diferite (2-100 aminoacizi). Bucla care leaga capatul C-terminal al foii β de capatul N-terminal al helixului are de obicei functie de recunoastere.
Macrostructura de dreapta
N C
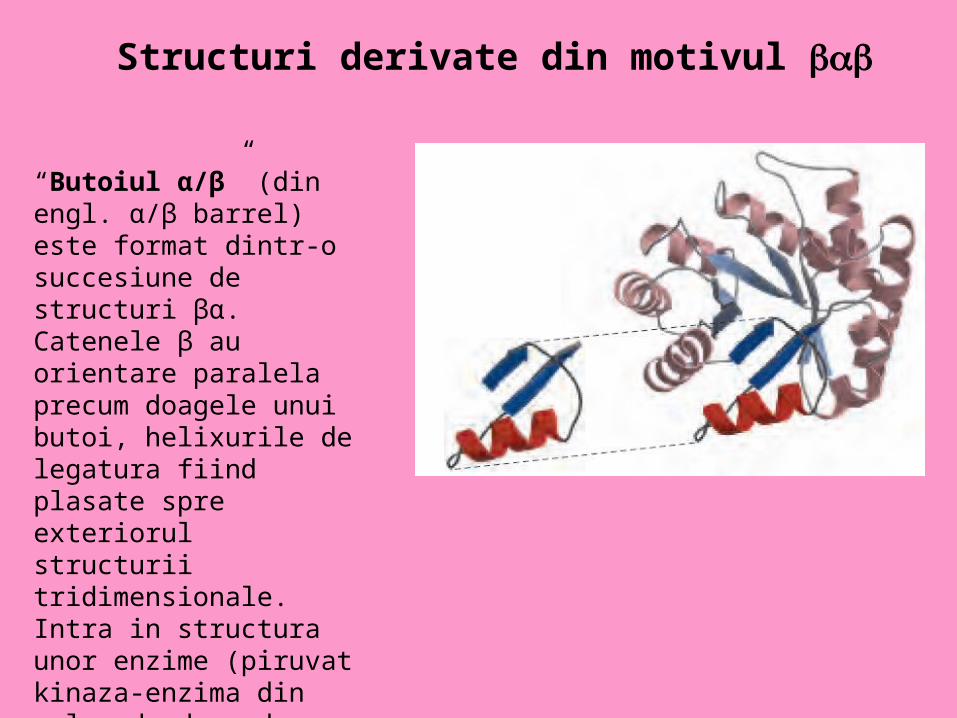
Structuri derivate din motivul
“Butoiul α/β” (din engl. α/β barrel) este format dintr-o succesiune de structuri βα. Catenele β au orientare paralela precum doagele unui butoi, helixurile de legatura fiind plasate spre exteriorul structurii tridimensionale. Intra in structura unor enzime (piruvat kinaza-enzima din calea de degradare a glucozei)

Domenii
• Domeniile pot fi structurale si functionale.
Domenii structurale: domeniile constante si variabile din structura anticorpilor si care diferentiaza diferite clase de anticorpi.
Domenii functionale: domeniile proteinelor de legare a ionilor de Ca, domeniile proteinelor de legare a ADN-ului.
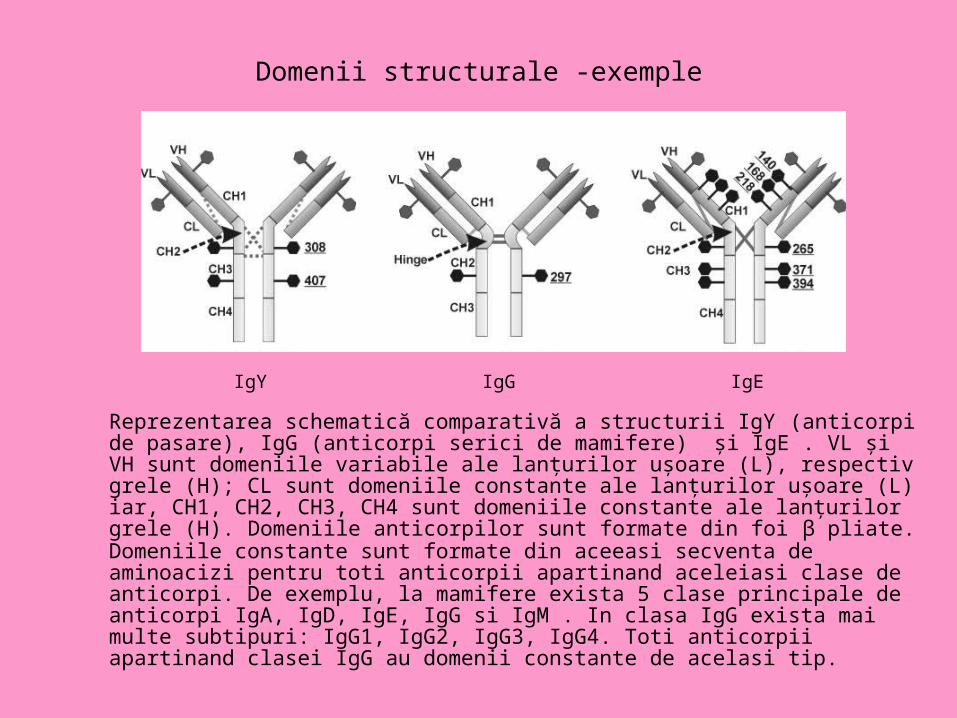
Domenii structurale -exemple
Reprezentarea schematică comparativă a structurii IgY (anticorpi de pasare), IgG (anticorpi serici de mamifere) şi IgE . VL şi VH sunt domeniile variabile ale lanţurilor uşoare (L), respectiv grele (H); CL sunt domeniile constante ale lanţurilor uşoare (L) iar, CH1, CH2, CH3, CH4 sunt domeniile constante ale lanţurilor grele (H). Domeniile anticorpilor sunt formate din foi β pliate.Domeniile constante sunt formate din aceeasi secventa de aminoacizi pentru toti anticorpii apartinand aceleiasi clase de anticorpi. De exemplu, la mamifere exista 5 clase principale de anticorpi IgA, IgD, IgE, IgG si IgM . In clasa IgG exista mai multe subtipuri: IgG1, IgG2, IgG3, IgG4. Toti anticorpii apartinand clasei IgG au domenii constante de acelasi tip.
IgY IgG IgE

Domenii functionale-exemple
Domeniile functionale: domeniile proteinelor de legare a ionilor de Ca, domeniile proteinelor de legare a ADN-ului.
Domeniul “degetului de zinc”(din engl. zinc finger).
• Proteinele pot contine unul sau mai multe degete de Zn. Fiecare motiv este format din 2 foi β antiparalele urmate de un α helix Un ion de Zn este legat prin 4 legaturi coordinative de doua resturi de histidina si doua de cisteina.
• Aceste domenii sunt prezente in structura factorilor de transcriere (Zif268) asigurand legarea de ADN.
• Aceste proteine interactioneaza cu ADN-ul la nivelul marii adancituri (depresiuni) astfel incat fiecare α-helix din structura unui “zinc finger” sa se intinda in jurul moleculei de ADN.

Aminoacizi si Proteine
Aminoacizi Structura, clasificare proprietati fizico-chimice
Legatura peptidicaAlte tipuri de legaturi chimice din moleculele proteiceProteine- nivele de organizare
Structura primarăStructura secundarăStructura terţiarăStructura cuaternară

Structura cuaternara
• Multe proteine, ca de exemplu enzimele, sunt alcătuite din două sau mai multe lanţuri polipeptidice numite subunităţi, legate între ele prin diferite legături necovalente ca: punţi de hidrogen, legături ionice şi forţe van der Waals.
• Structura cuaternară descrie relaţiile spaţiale stabilite între diferitele catene polipeptidice ce intră în alcătuirea unei proteine (are în vedere existenţa a cel puţin două catene polipeptidice independente ce intră în asamblarea unei proteine funcţionale).
• Structura cuaternară prezintă distribuţia spaţială a subunităţilor şi stabileşte ansamblul interacţiunilor şi contactelor dintre subunităţile unei proteine.

Rolul structurii cuaternare
• Structura cuaternară are consecinţe asupra activităţii biologice a proteinelor active. Subunităţile ce alcătuiesc proteina enzimatică se intercondiţionează reciproc. Modificările în conformaţia spaţială a uneia din subunităţi, ca urmare a legării unei molecule de substrat, coenzimă, activator sau inhibitor, pot induce modificări conformaţionale celorlalte subunităţi ale proteinei. Asemenea interacţiuni stau la baza fenomenului de alosterism.

Numarul de subunitati ale unei proteine
În general, numărul de subunităţi ale unei proteine este număr par:
• dimer• tetramer• hexamer• octamer• decamer• dodecamer etc.

Exemplu de proteine cu numar impar de subunitati: Proteinele G
• Există însă şi proteine cu număr impar de subunităţi, ca de exemplu proteinele G membranare care sunt alcătuite din trei subunităţi (trimer).Proteinele G sunt heterotrimeri αβγ alcătuiţi din trei subunităţi diferite: α, şi . Subunitatea α prezintă un situs de legare pentru GDP/GTP.
• Proteina G inactivă este sub formă de trimer: Gα -GDP-GβGγ, în timp ce proteina G activă se găseşte sub formă de monomer legat de GTP: Gα-GTP. Procesul de activare/inactivare este reversibil.

Exemplu de proteina formata din catene polipeptidice diferite structural şi funcţional: Protein-kinaza A (PKA)
• Protein-kinaza A (PKA) este formată din patru subunităţi: două cu rol catalitic şi două cu rol reglator. Când acestea sunt asociate, proteina nu manifestă activitate enzimatică, în timp ce disocierea lor, cu îndepărtarea subunităţilor reglatoare, conduce la câştigarea activităţii enzimatice a subunităţilor catalitice.

Exemplu de complex enzimatic: complexul acid gras sintetazic
- Pot fi asociate catene polipeptidice având mai multe tipuri de structuri şi funcţii. De exemplu: La procariote (Escherichia coli) si la plante, sintetaza acizilor graşi este un complex enzimatic multifuncţional format din şase enzime diferite şi o proteină de transport ,ACP (din engl. “Acyl Carrier Protein”).

Proprietăţile proteinelor
• Proteinele sunt substanţe solide, incolore, stabile la temperatura camerei.
• La temperaturi mai mari de 50-60°C, proteinele se denaturează.
• Unele proteine pot fi obţinute în stare cristalină, altele numai în stare amorfă.

Solubilitatea proteinelor
• Proteinele se deosebesc unele de altele prin gradul lor de solubilitate. Unele proteine sunt solubile în apă distilată, altele în soluţii saline diluate, iar altele în soluţii diluate de acizi sau baze; toate proteinele sunt însă insolubile în solvenţi organici.
• Există şi proteine insolubile în mod normal, cum sunt cele fibrilare, care intră în structura ţesuturilor de susţinere sau de protecţie a organismului (unghii, copite, cioc, coarne etc.).
• Moleculele proteice pot fixa mari cantităţi de apă (până la 0,3 g/g) prezentând caracter de coloizi hidrofili reversibili.
• Unele proteine cu structură fibrilară, cum este gelatina, se îmbibă puternic cu apa formând geluri. Imbibiţia este influenţată de ionii anorganici absorbiţi. Gradul de îmbibare şi de stabilitate a proteinei este minim la punctul izoelectric, unde proteinele precipită foarte uşor.

Punctul izoelectric al proteinelor
• Majoritatea proteinelor sunt electroliţi amfoteri, având atât proprietăţi ale bazelor, cât şi ale acizilor. În fiecare proteină există numeroase grupări aminice (-NH2) şi carboxilice (-COOH) libere, aparţinând aminoacizilor diaminomono-carboxilici, respectiv monoaminodicarboxilici.
• În cursul titrării unei proteine apare o valoare de pH la care încărcarea aparentă este zero; acest pH se numeşte punct izoelectric (pI) şi se evidenţiază experimental prin lipsa migrării în câmpul electric. La punctul izoelectric, sarcina netă a moleculei proteice este nulă, iar solubilitatea proteinei este minimă.
• Din cauza numărului mare de sarcini purtate, proteinele sunt polielectroliţi. La trecerea unui curent electric, ionii se vor deplasa spre polul de semn contrar, fenomen numit electroforeză. Combinarea electroforezei cu precipitarea la punctul izoelectric ("isoelectric focussing") constituie o metodă modernă şi extrem de sensibilă pentru separarea şi izolarea diverselor proteine din amestecuri.

Denaturarea proteinelor• =pierderea proprietăţilor caracteristice stării native a unei proteine, sub acţiunea unor
agenţi chimici, fizici şi mecanici. Denaturarea presupune dezorganizarea structurii spaţiale (cuaternare, terţiare şi secundare) a moleculei proteice, datorită ruperii legăturilor necovalente inter- şi intracatenare, a deplierii lanţului polipeptidic şi, în unele cazuri, a scindării legăturilor disulfurice.
• Denaturarea se poate manifesta prin: pierderea activităţii biologice, creşterea reactivităţii unor grupări chimice din moleculă, micşorarea solubilităţii, precipitare sau coagulare, schimbarea formei sau a dimensiunii moleculei, schimbarea activităţii optice specifice.
• Dintre agenţii denaturanţi menţionăm: ureea, clorura de guanidină, detergenţii, alcoolii, modificările drastice de pH, de tărie ionică şi de temperatură.
• Denaturare poate fi reversibilă, proteina recăpătându-şi proprietăţile ei native când agentul denaturant a fost îndepărtat. Renaturarea este operaţia inversă denaturării. Când însă denaturarea persistă şi după îndepărtarea agentului denaturant, procesul este considerat ireversibil. În cazul denaturării ireversibile are loc ruperea legăturilor chimice propriu-zise (în special a legăturilor disulfurice inter- şi intracatenare).

Precipitarea proteinelor
• Reprezintă trecerea din stare dizolvată în stare solidă (precipitat). Proteinele precipită la punctul izoelectric. Precipitările pot fi reversibile sau ireversibile.
• Ionii anorganici în concentraţie mare precipită proteinele prin spargerea sferei de hidratare care înconjoară şi stabilizează molecula proteică dizolvată, pentru a se dizolva ei înşişi în apa extrasă. Fenomenul acesta se numeşte salifiere.
Sare (sulfat de amoniu)

Precipitarea proteinelor
• Concentraţia salină de precipitare variază de la o proteină la alta, ceea ce permite fracţionarea proteinelor dintr-un amestec neomogen. Sărurile de sulfat de amoniu, de sulfat de magneziu şi sulfat de sodiu, precipită în mod reversibil proteinele, cu păstrarea activitaţii biologice. Unele săruri pot produce precipitarea ireversibilă a proteinelor, ca în cazul sărurilor metalelor grele: acetatul de plumb, sulfatul de zinc, azotatul de argint, clorura mercurică etc.
• O clasă de precipitanţi proteici des utilizaţi în laborator include acizi ( acidul tricloracetic, acidul percloric, acidul sulfosalicilic, acidul fosfotungstic (fosfowolframic), acidul picric) sau baze care pot desface puntile saline din structura lanturilor polipeptidice.

Funcţia tampon a proteinelor
• Proteinele, ca şi aminoacizii, au capacitate tampon datorită existenţei grupărilor aminice (slab acide) protonate şi a grupărilor carboxilice (slab acide) disociate. Sistemele tampon realizate de proteine sunt foarte sensibile şi eficiente; circa 75% din capacitatea de tamponare a plasmei sanguine este asigurată de hemoglobină.
• Proteinele au putere de tamponare la două valori de pH, corespunzătoare valorilor pK1 şi pK2 corespunzătoare grupărilor -COOH şi -NH2 libere, măsurate pe scara de pH. Deoarece moleculele proteice pot cuprinde numeroase grupări carboxilice şi aminice libere, având pK diferite, se poate uşor deduce că proteinele au capacitate de tamponare în mai multe regiuni de pH, astfel încât, în orce domeniu de pH, adăugarea unor cantităţi apreciabile de acid sau bază determină variaţii minime de pH ale soluţiei proteice.

Clasificarea proteinelor
După structura chimică:
1. Holoproteine ("Proteine simple")
1.1. Scleroproteine ("Proteine fibrilare“)
1.2. Sferoproteine ("Proteine globulare")
2. Heteroproteine ("Proteine conjugate")

Proteine simple (holoproteine)
• Holoproteinele dau prin hidroliză numai α-aminoacizi.
În funcţie de aspectul exterior, se împart în:
proteine fibrilare ("scleroproteine")
proteine globulare ("sferoproteine").

Proteinele fibrilare
• Sunt de origine animală, sunt insolubile în apă şi au rezistenţă mare la acţiunea enzimelor proteolitice. Datorită insolubilităţii lor, nu li se poate determina cu precizie masa moleculară. Au molecula alungită, filiformă: două sau trei catene polipeptidice (helix triplu) se răsucesc una în jurul celeilalte formând o fibră lungă.
• Se găsesc în firul de mătase, în pânza de paianjen, în firul de lână şi cel de păr, în piele, coarne, ţesut conjunctiv şi oase.
• Se subdivid în: colageni, elastine şi cheratine.

Proteinele globulare• Lanţurile polipeptidice au structura terţiară ce le conferă aspectul globular (de ghem).
Cuprind proteine cu funcţie biologică dinamică cum sunt enzimele, hormonii proteici, proteinele de transport. Sunt solubile în apă sau în soluţii de săruri minerale, acizi, baze. Sunt de origine animală şi vegetală.
a) Albuminele • Albuminele de origine animală : serumalbumina (în plasma sanguină), lactalbumina
(în lapte), ovalbumina (în albuşul de ou), miogenul (în ţesutul muscular). • Albuminele vegetale : faseolina (în fasole), legumelina (în mazăre), leucozina (în
orez, grâu, secară). b) Globulinele: Globulinele de origine animală: miozina (în muşchi), globulinele serice,
fibrinogenul (în plasma sanguină), ovoglobulina (din ou), lactoglobulina (din lapte). Globulinele vegetale menţionăm: legumina (în mazăre şi linte) şi edestina (în seminţele de cânepă).
c) Protaminele conţin mari cantităţi de aminoacizi bazici (până la 80 %), aşa cum sunt lizina, histidina şi, în special, arginina. Din compoziţia lor lipsesc complet aminoacizii cu sulf cum este cisteina. Au caracter bazic, sunt solubili în apă şi în soluţii diluate de acizi sau baze. Protaminele intră în compoziţia celulelor germinale mature la animale.
d) Histonele sunt proteine mai complexe decât protaminele. Sunt alcătuite în cea mai mare parte din aminoacizi bazici, în special din arginină. Nu conţin triptofan şi nici aminoacizi cu sulf. Au punctul izoelectric în jurul valorii de pH 10. În celulă, histonele sunt asociate cu ADN.

Proteine conjugate (heteroproteine)• Dau prin hidroliză, pe lângă proteină şi o componentă neproteică, numită
grupare prostetică. Au proprietăţi asemănătoare proteinelor simple. În funcţie de natura grupării prostetice, heteroproteinele se pot clasifica în: metalo-, fosfo-, lipo-, nucleo-, glico-, cromoproteine.
- a) Metaloproteinele au ca grupare prostetică un metal fixat prin legături complexe de molecula proteică simplă. Conţin fier, cupru, zinc, mangan, cobalt, molibden, magneziu etc. Spre deosebire de unele cromoproteine ce conţin metale în grupările prostetice, în metaloproteine metalul este legat de resturile de aminoacizi din proteină. Dintre metaloproteinele de natură animală, un interes deosebit îl prezintă cele care au rol de depozitare cum este feritina - pentru fier, de transport de metale cum sunt transferina (“siderofilina”) - pentru fier şi ceruloplasmina - pentru cupru, precum şi unele enzime ca ascorbatoxidaza (ce conţine cupru) şi anhidraza carbonică (ce conţine zinc).
- b) Fosfoproteinele au ca grupare prostetică acidul fosforic ce esterifică permanent grupările alcoolice ale serinei şi treoninei unor proteine de constituţie. Fosfoproteinele au caracter acid, sunt insolubile în apă şi în acizi, dar sunt solubile în soluţii bazice, în special în bicarbonat. Cele mai importante fosfoproteine de origine animală sunt: cazeina(din lapte), vitelina şi fosfovitina din gălbenuşul de ou.

Proteine conjugate (heteroproteine)
- c) Lipoproteinele pot fi clasificate pe baza densităţii lor sau pe baza mobilităţii lor electroforetice. Densitatea particulelor lipoproteice este determinată prin centrifugare în soluţie de NaCl cu densitatea 1,063. Lipoproteinele au densitate mică şi o constantă de flotaţie proprie (constanta de flotaţie definindu-se ca fiind inversul constantei de sedimentare).
- Se pot distinge cinci fracţiuni: 1) lipoproteine cu densitate ridicată (HDL - ‘high-density lipoproteins”); 2) lipoproteine cu densitate scăzută (LDL - “low-density lipoproteins”); 3) lipoproteine cu densitate intermediară (IDL - “intermediate-density lipoproteins”); 4) lipoproteine cu densitate foarte scăzută (VLDH - “very low-density lipoproteins”); 5) Chilomicroni.

Exemplu de lipoproteine-chilomicroni
• Trigiceridele, colesterolul impreuna cu lipoproteine (ApoA, B, C, E) sunt impachetate in chilomicroni care le transporta in sistemul limfatic si apoi in sange.
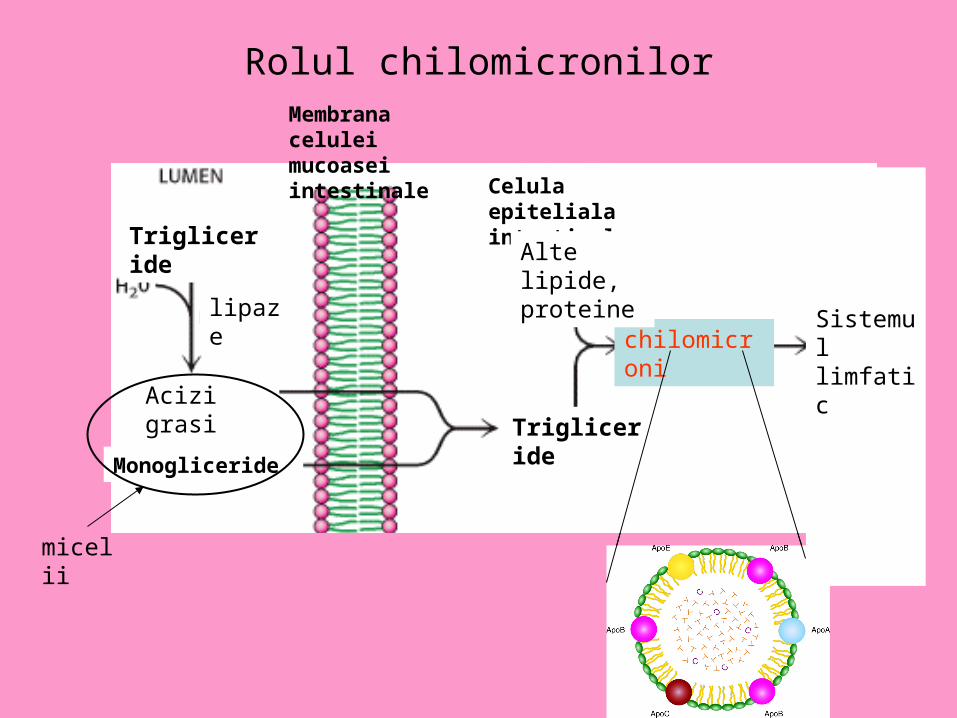
Rolul chilomicronilor
Celula epiteliala intestinala
Trigliceride
lipaze
Acizi grasi
Monogliceride
micelii
Trigliceride
chilomicroni
Alte lipide, proteine
Sistemul limfatic
Membrana celulei mucoasei intestinale

Proteine conjugate (heteroproteine)
- d)Glicoproteinele au ca grupare prostetică glucide. Au fost tratate in capitolul de Glucide (glicocalix, antigenele ABO).

Proteine conjugate (heteroproteine)
- f) Cromoproteinele au ca grupare prostetică substanţe colorate, uneori şi un metal. După natura grupării prostetice se clasifică în: cromoproteine porfirinice şi neporfirinice. După rolul lor biologic, se subdivid în: pigmenţi cu funcţie respiratorie şi pigmenţi fără funcţie respiratorie.
• Cromoproteinele porfirinice sunt: hemoglobina, citocromii, citocromoxidaza, catalaza, peroxidaza, cloroplastinele (clorofile), ficocianinele (la algele albastre) şi ficoeritrinele (la algele roşii).
• Cromoproteinele neporfirinice sunt: hemeritrina (colorant sanguin la unele nevertebrate), hemocianinele (coloranţi sanguini având cupru din hemolimfa moluştelor şi crustaceelor), rodopsina (pigment vizual – purpura retiniană din celulele cu bastonaşe ale retinei), flavoproteinele (enzime oxidoreducătoare), carotenproteinele.
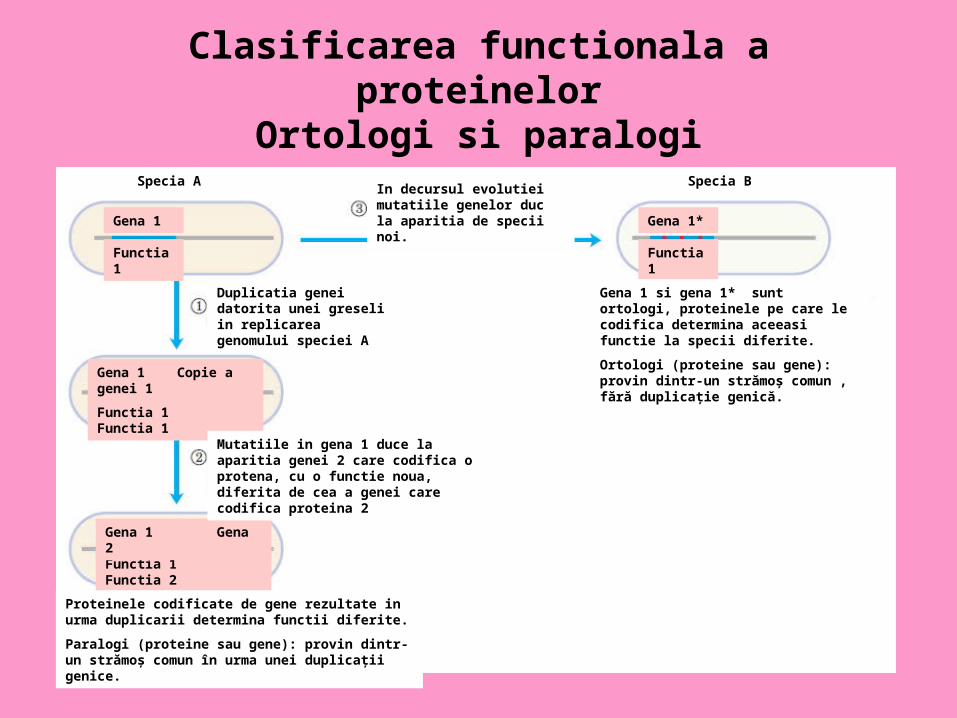
Clasificarea functionala a proteinelorOrtologi si paralogi
Specia A Specia B
Duplicatia genei datorita unei greseli in replicarea genomului speciei A
Gena 1 Copie a genei 1
Functia 1 Functia 1
Gena 1
Functia 1 Functia 2
Functia 1
Gena 1 Gena 2
Gena 1*
Functia 1
Proteinele codificate de gene rezultate in urma duplicarii determina functii diferite.
Paralogi (proteine sau gene): provin dintr-un strămoş comun în urma unei duplicaţii genice.
Mutatiile in gena 1 duce la aparitia genei 2 care codifica o protena, cu o functie noua, diferita de cea a genei care codifica proteina 2
Gena 1 si gena 1* sunt ortologi, proteinele pe care le codifica determina aceeasi functie la specii diferite.
Ortologi (proteine sau gene): provin dintr-un strămoş comun , fără duplicaţie genică.
In decursul evolutiei mutatiile genelor duc la aparitia de specii noi.
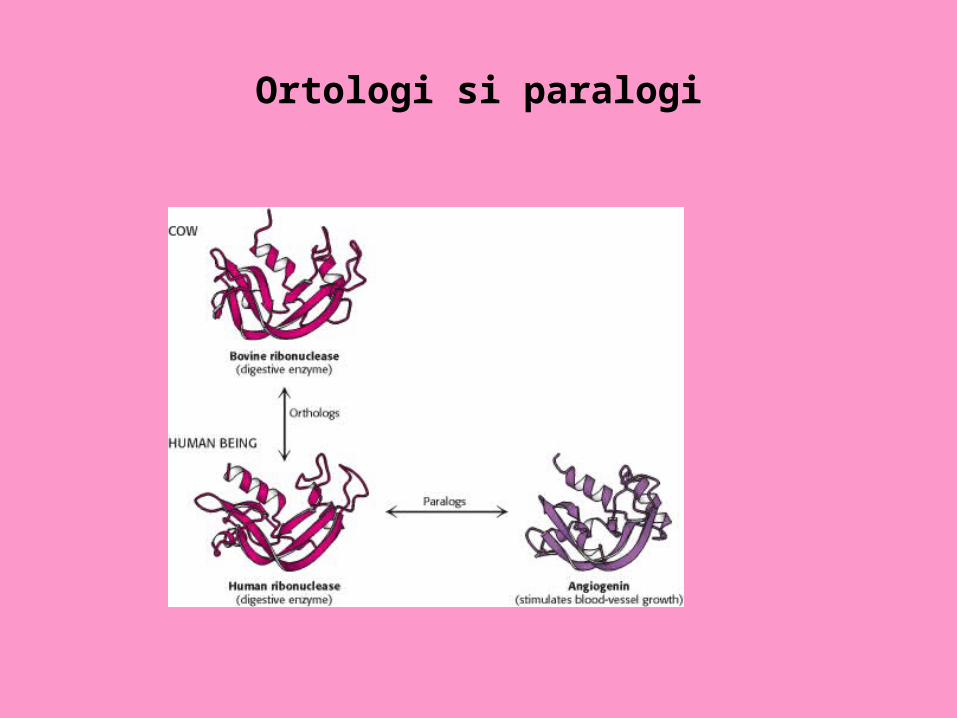
Ortologi si paralogi

Exemplu de clasificare funcţională
COGs – Cluster Orthologous Groups (http://www.ncbi.nlm.nih.gov/COG/old/palox.cgi?fun=all)
Baza de date COGs clasifică proteinele codificate de genomuri în 18 clase funcţionale. Fiecare clasă cuprinde un numar variabil de proteine, de la câteva zeci până la câteva sute





























