Embed Size (px)
Citation preview

Pulmonary xenotransplantation: Rapidly progressinginto the unknown
Edward Cantu1*, William Parker1, JeffreyL. Platt2 and R. Duane Davis1
1Department of Surgery, Duke University Medical Center,Durham, NC;2Departments of Surgery, Immunology, and Pediatrics,Mayo Clinic, Rochester, MN, USA*Corresponding author: Edward Cantu, [email protected]
As one approach to circumventing the dire shortageof human lungs for transplantation, a handful ofinvestigators have begun to probe the possibility ofpulmonary xenotransplantation. The immunologicand perhaps physiologic barriers encountered bythese investigators are considerable and progress inpulmonary xenotransplantation has lagged behindprogress in cardiac and kidney xenotransplantation.However, during the last few years there have beensubstantial advances in the field of pulmonary xeno-transplantation including, most noticeably, significantprogress in attenuating hyperacute dysfunction. Pro-gress has been made in understanding the barriersimposed by xenoreactive antibodies, complement,coagulation incompatibility and porcine pulmonaryintravascular macrophages. Although our under-standing of the barriers to pulmonary xenotransplant-ation is far from complete and the clinical applicationof pulmonary xenotransplantation is not yet in sight,current progress is fast paced. This progress providesa basis for future work and for a hope that the short-age of human lungs for transplantation will notalways be a matter of life and death.
Received July 17 2003, revised and accepted for pub-lication September 10 2003.
Introduction
In the last four decades following the first clinical lung
transplant (1), advances in immunosuppression and refine-
ments in surgical technique have transformed clinical lung
transplantation into an effective treatment for many
patients with end stage lung disease. Reported cumula-
tive world experience exceeds 14 000 lung transplants
with 73% 1-year and 45% 5-year overall survival (2). The
moderate long-term success achieved, growth of the can-
didate pool and the relatively fixed number of available
donors has resulted in a health crisis where the median
time to transplant has doubled and for every two candidates
who receive an allograft, one dies while still on the
waiting list (3). It has been this trend and the relatively
primitive long-term mechanical replacements available
that have encouraged investigators to evaluate pulmon-
ary xenotransplantation. Such an approach offers a
potential unlimited source of donor organs.
For reasons that have been reviewed elsewhere (4,5),
the pig is currently considered to be the most promising
potential donor for xenotransplantation. In light of this
view, those experiments which model pig-to-human
pulmonary xenotransplantation are considered to be
the most clinically relevant, and are the topic of this
review. Models utilizing pig to primate pulmonary xeno-
transplantation and lung perfusion models using human or
occasionally baboon blood products are discussed. In some
cases, data from other models and from other solid organ
xenotransplants provide information that may provide
insight into pulmonary xenotransplantation, and the implica-
tions of this research for pulmonary xenotransplantation are
discussed.
Hyperactute Pulmonary XenograftDysfunction
The initial experience with pulmonary xenotransplantation
using porcine organs was reported in 1968, prior to the
development of extracorporeal mechanical oxygenators.
Using an ex vivo perfusion apparatus, Bryant and col-
leagues tested pig lungs as potential biologic oxygenators
for humans undergoing cardiac surgical procedures (6).
Unlike the use of lungs from monkeys, the use of swine
lungs was associated with rapid failure of the lung. This
failure was characterized by (a) elevated pulmonary vas-
cular resistance and (b) the rapid development of massive
pulmonary edema evident by gross examination (6). The
presence of these two ‘components’, thrombosis and
edema, has been confirmed by a number of studies
since the initial description in 1968. Histologic examination
of porcine lungs following exposure to human or primate
blood has confirmed the severe pulmonary edema and
demonstrated that the elevated pulmonary vascular resist-
ance is associated with micro- and macrovascular throm-
bosis. Because little evidence existed that immunologic
rejection by the immune system of the recipient was
responsible for pulmonary xenograft failure, the milieu of
thrombosis and edema associated with pulmonary xeno-
transplantation has been termed ‘hyperacute xenograft
dysfunction’. Subsequent studies, described below, have
supported the idea that factors other than the recipient
American Journal of Transplantation 2004; 4 (Suppl. 6): 25–35 Copyright # Blackwell Munksgaard 2004Blackwell Munksgaard
25
immune system play a key role in hyperacute xenograft
dysfunction. Further, the milieu associated with pulmon-
ary xenograft dysfunction is distinct from that associated
with the hyperacute xenograft rejection and the acute
vascular rejection of porcine hearts or kidneys. For these
reasons, the term ‘hyperacute xenograft dysfunction’
rather than the term ‘hyperacute rejection’ or other related
terms is still used to describe the milieu associate with
pulmonary xenograft failure.
In addition to thrombosis and edema, pulmonary xeno-
transplantation has been associated with profound sys-
temic problems. Most noticeably, profound hypotension
occurs in the organ recipient, which requires inotropic
support to maintain the recipient’s hemodynamics in
many experiments. It is likely that this adverse response
is due to a variety of molecules released from the organ,
including C5a and perhaps a number of cytokines, includ-
ing TNFa. In addition, activation of the coagulation cascade
results in a disseminated intravascular coagulopathy (DIC)
that is reversible upon explant of the xenogeneic lung (7).
The coagulopathy is characterized by increased PT, deple-
tion of platelets, fibrinogen, and a dramatic increase in
circulating thrombin-antithrombin complex. Yet another
problem associated with pulmonary xenotransplantation
is a profound depletion of leukocytes from the recipient’s
blood.
A variety of therapies have been successful in eliminating
or delaying one or more of the pathologic process asso-
ciated with pulmonary xenotransplantation (Figures 1–3;
Tables 1 and 2). The results of these experiments and their
implications for the pathogenesis of pulmonary xenograft
dysfunction are described below.
Xenoreactive Antibody and PulmonaryXenotransplantation
There is evidence that xenoreactive antibodies play a role
in hyperacute pulmonary xenograft dysfunction. Pierson
and colleagues, using an ex vivo swine heart-lung model
perfused with human blood, have demonstrated that
increases in pulmonary vascular resistance and pulmonary
edema are associated with immunohistochemical demon-
stration of prominent deposition of IgM and the comple-
ment fragment C4 in the alveolar interstitium, somewhat
less deposition of IgG and patchy deposition of properdin,
C3, and C9. These preliminary studies indicate that xeno-
reactive natural antibody, predominately IgM, binds to the
xenograft and activates complement. Other investigators,
using ex vivo isolated lung perfusion models have repro-
duced these results, finding an association between the
rapid failure of the pulmonary xenografts with deposition
of IgM, IgG, Clq, C3, C4, and C9 (8,9). However, whether
xenoreactive antibody actually remains bound to porcine
endothelium remains in question. Studies performed by
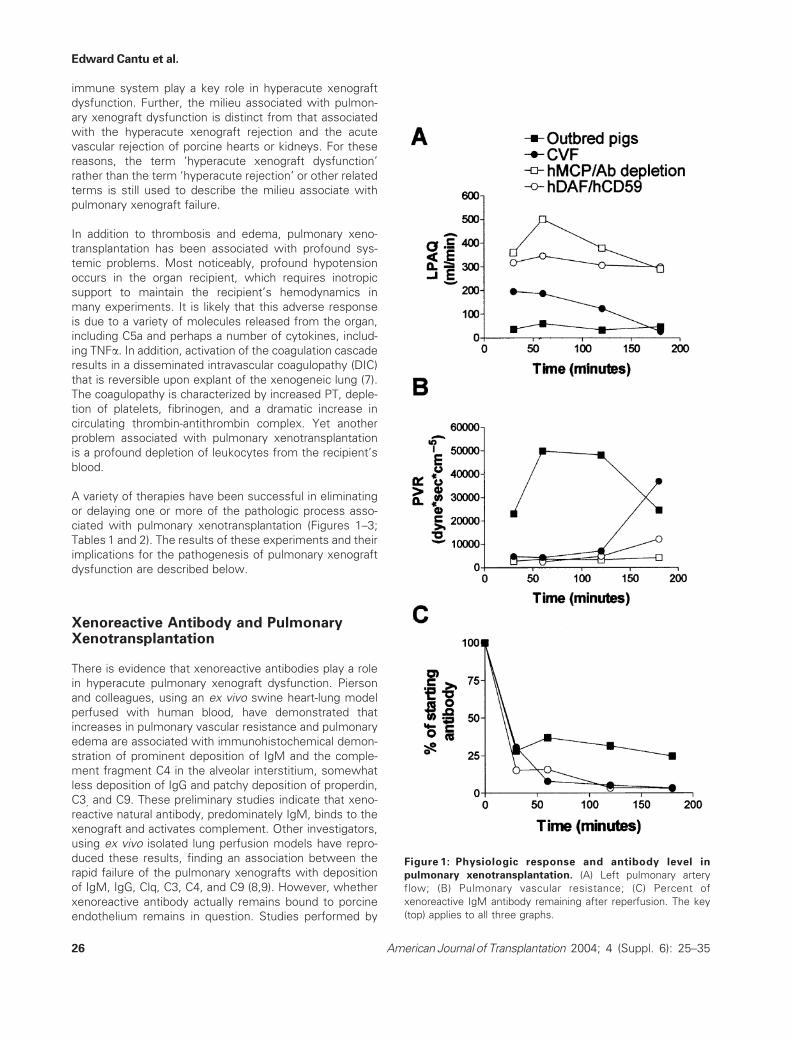
Figure 1: Physiologic response and antibody level in
pulmonary xenotransplantation. (A) Left pulmonary artery
flow; (B) Pulmonary vascular resistance; (C) Percent of
xenoreactive IgM antibody remaining after reperfusion. The key
(top) applies to all three graphs.
Edward Cantu et al.
26 American Journal of Transplantation 2004; 4 (Suppl. 6): 25–35
Parker, Platt and Davis (10) have demonstrated that,
although xenoreactive antibodies can bind to pulmonary
xenografts, many of these antibodies are shed as anti-
body-antigen complexes. A prominent role of these
immune complexes in the activation of complement dur-
ing pulmonary xenotransplantation seems likely, but has
yet to be proven.
Some experiments in which xenoreactive antibody were
depleted from recipients demonstrated improved pulmon-
ary xenograft function in the absence of xenoreactive
antibodies. For example, Pierson and associates demon-
strated that elevated pulmonary vascular resistance could
be prevented with depletion of xenoreactive antibody
utilizing incubation of human plasma with pig splenocytes
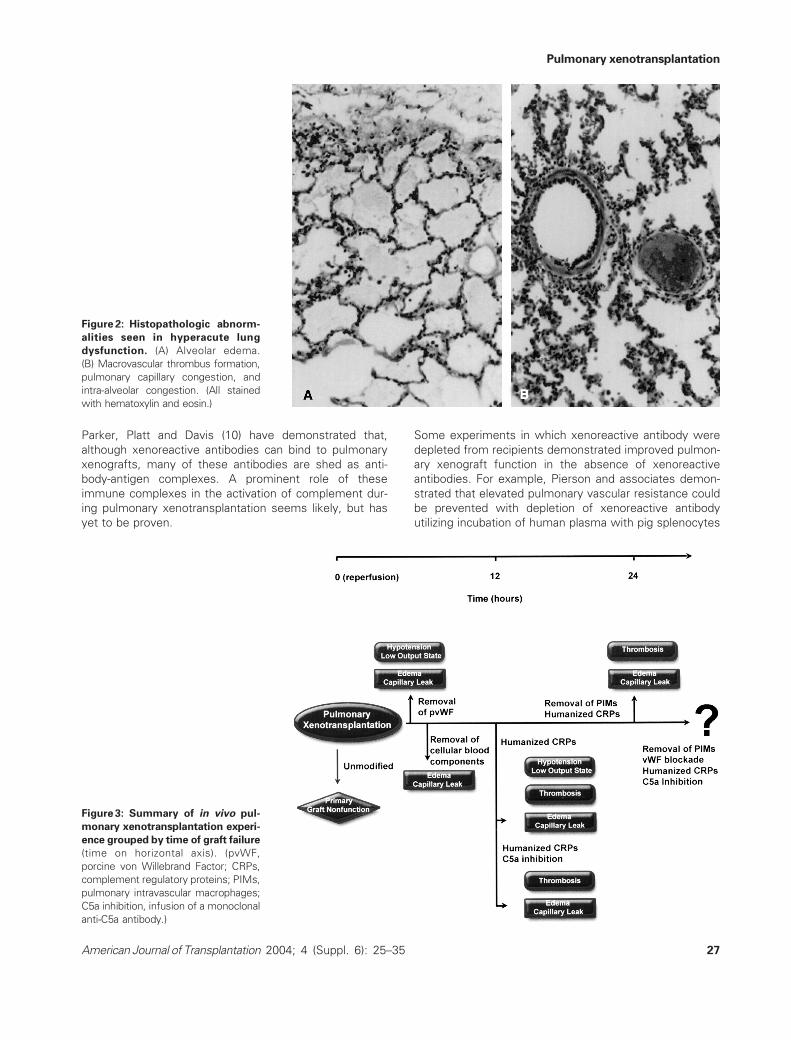
Figure 2: Histopathologic abnorm-
alities seen in hyperacute lung
dysfunction. (A) Alveolar edema.
(B) Macrovascular thrombus formation,
pulmonary capillary congestion, and
intra-alveolar congestion. (All stained
with hematoxylin and eosin.)
Figure 3: Summary of in vivo pul-
monary xenotransplantation experi-
ence grouped by time of graft failure
(time on horizontal axis). (pvWF,
porcine von Willebrand Factor; CRPs,
complement regulatory proteins; PIMs,
pulmonary intravascular macrophages;
C5a inhibition, infusion of a monoclonal
anti-C5a antibody.)
Pulmonary xenotransplantation
American Journal of Transplantation 2004; 4 (Suppl. 6): 25–35 27
and peripheral blood cells (11). Macchiarini (12) demons-
trated that prolonged porcine lung survival could be achieved
by first depleting human blood of xenoreactive antibodies
using a porcine lung. Similarly, Lau et al. (13) demonstrated
that depletion of xenoreactive antibodies using an extracor-
poreal lung perfusion resulted in prolonged xenograft survi-
val. However, perfusion of blood through a porcine lung and
perhaps incubation with porcine splenocytes depletes other
components of human and primate blood, including com-
plement, fibrinogen, platelets and leukocytes. Thus, experi-
ments more specifically targeted at xenoreactive antibodies
are required to address the importance of the antibodies in
pulmonary xenotransplantation.
Experiments designed specifically to test the importance of
xenoreactive antibodies in pulmonary xenotransplantation
have not led to the conclusion that xenoreactive antibodies
are critical for pulmonary xenograft dysfunction. For
example, depletion of xenoreactive antibodies by extra cor-
poreal perfusion through a porcine kidney, unlike depletion
using a porcine lung, did not result in prolonged xenograft
survival (13). Similarly, extra corporeal perfusion through
porcine liver or spleen proved unsuccessful in prolong-
ing pulmonary xenograft survival (12). Further, depletion
of anti-Gala1-3Gal antibodies using a column containing
synthetically coupled aGal did not result in prolonged
pulmonary xenograft function. The lack of improved xeno-
graft function in these experiments may be associated
with the release of anaphylatoxins such as C5a, dis-
cussed below. However, hyperacute pulmonary xenograft
dysfunction also occurred in a model using neonatal
primate recipients, which lack anti-Gala1-3Gal antibodies,
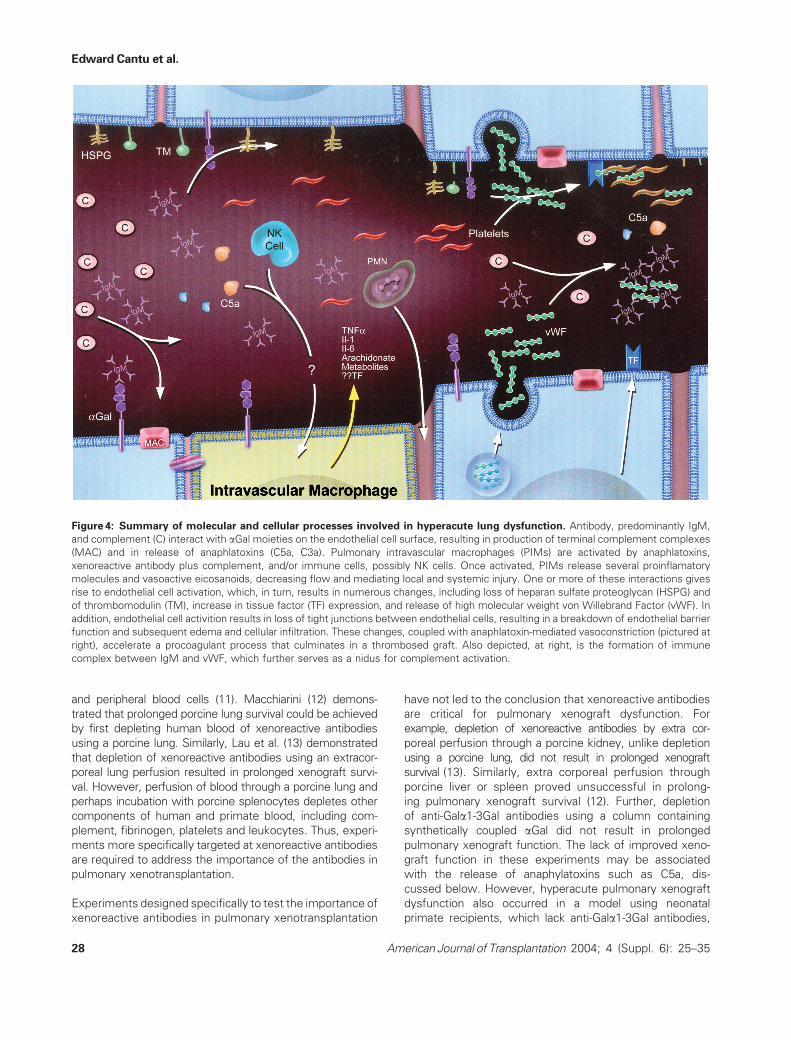
Figure 4: Summary of molecular and cellular processes involved in hyperacute lung dysfunction. Antibody, predominantly IgM,
and complement (C) interact with aGal moieties on the endothelial cell surface, resulting in production of terminal complement complexes
(MAC) and in release of anaphlatoxins (C5a, C3a). Pulmonary intravascular macrophages (PIMs) are activated by anaphlatoxins,
xenoreactive antibody plus complement, and/or immune cells, possibly NK cells. Once activated, PIMs release several proinflamatory
molecules and vasoactive eicosanoids, decreasing flow and mediating local and systemic injury. One or more of these interactions gives
rise to endothelial cell activation, which, in turn, results in numerous changes, including loss of heparan sulfate proteoglycan (HSPG) and
of thrombomodulin (TM), increase in tissue factor (TF) expression, and release of high molecular weight von Willebrand Factor (vWF). In
addition, endothelial cell activition results in loss of tight junctions between endothelial cells, resulting in a breakdown of endothelial barrier
function and subsequent edema and cellular infiltration. These changes, coupled with anaphlatoxin-mediated vasoconstriction (pictured at
right), accelerate a procoagulant process that culminates in a thrombosed graft. Also depicted, at right, is the formation of immune
complex between IgM and vWF, which further serves as a nidus for complement activation.
Edward Cantu et al.
28 American Journal of Transplantation 2004; 4 (Suppl. 6): 25–35

indicating that anti-Gala1-3Gal antibodies are not absolutely
required to initiate pulmonary xenograft dysfunction. The
neonatal recipients do, however, contain polyreactive,
xenoreactive antibodies (14,15), suggesting the idea that
antibodies other than anti-Gala1-3Gal antibodies might be
involved in hyperacute pulmonary xenograft dysfunction. On
the other hand, the results may reflect the importance of
factors other than xenoreactive antibodies in pulmonary
xenograft failure. Experiments outlined below utilizing spe-
cific depletion of porcine macrophages from the donor lung
are consistent with this latter interpretation.
Complement in PulmonaryXenotransplantation
Experiments utilizing porcine lungs expressing human
complement regulatory proteins (hCRP) have provided a
wealth of information regarding the role of complement in
pulmonary xenotransplantation. Porcine organs expres-
sing hCRPs, unlike lungs lacking such expression, sub-
stantially attenuate human or primate complement
activation, although perhaps not as efficiently as human
or primate organs (16,17). A number of experiments have
consistently shown improved function of xenografted
porcine lungs expressing hCRP’s vs. those lacking such
expression. For example, Dagget et al. in an ex vivo
perfusion study with human plasma in lungs expressing
DAF and CD59 (Nextran, Princeton, NJ), demonstrated
better function and decreased pulmonary vascular resist-
ance (18) than in non transgenic organs. Similarly, in
Pierson’s ex vivo model, there was a mild prolongation
in xenograft function using swine lungs transgenic for the
hDAF (19). In another series of orthotopic pulmonary
xenotransplants, using swine transgenic for another
CRP, human membrane cofactor protein (MCP, Nextran,
Princeton, NJ), were conducted. As with lungs from
hDAF/CD59 donors, lungs expressing human MCP exhibi-
ted greater blood flow through the xenotransplant, lower
pulmonary vascular resistances, and delayed evidence of
histologic injury as compared to controls expressing no
hCRPs (20).
Although pulmonary xenografts expressing hCRPs fare
better than those lacking hCRPs, they are still subject to
hyperacute pulmonary xenograft dysfunction within hours
(Figure 3). This observation may reflect the idea that
Table 1: Ex vivo pulmonary xenotransplantation experience. Antibody depletion was performed using various strategies prior to
transplant: (1) baboon-to-pig organ perfusion (lung, liver, or spleen); (2) column plasmapheresis (column); or (3) plasma incubation with
aGal antigen (Ag incubation). Transgenic donors were of several types: (1) human decay accelerating factor (hDAF); (2) human CD59; or
(3) combinations thereof. Complement modulation was accomplished either through: (1) heat inactivation; (2) soluble complement
receptor 1 (sCR1); (3) K76-COOH (classical complement activation blockade); (4) FUT-175 (classical and alternative complement activation
blockade). Coagulant modulation was imposed with either heparin or recombinant hirudin (rHirudin)
Donor Recipient n Treatment Survival (h) Reference
Swine Human 7 None 0.2 Azimzadeh et al./2003
10 None 0.5 Azimzadeh et al./2003
4 None 1.9 Wiebe et al./2000
6 None 0.6 Kulick et al./2000
10 None 2.0 Yeatman et al./1999
6 None 2.0 Dagget et al./1997
6 None 1.9 Macchiarini et al./1997
11 None 0.3 Pierson et al./1997
7 None 0.2 Blum et al./1998
6 Ab Depletion(lung perfusion) 5.0 Macchiarini et al./1998
6 Ab Depletion(liver perfusion) 2.5 Macchiarini et al./1998
6 Ab Depletion(spleen perfusion) 1.0 Macchiarini et al./1998
6 Ab Depletion(column) 1.8 Macchiarini et al./1998
3 Ab Depletion(column) 1.5 Pierson et al./1997
12 Transgenic Donor(hDAF) 1.5 Azimzadeh et al./2003
4 Transgenic Donor(hDAF) 4.0 Wiebe et al./2000
5 Transgenic Donor(hCD59) 4.0 Kulick et al./2000
10 Transgenic Donor(hDAF/hCD59) 2.0 Yeatman et al./1999
6 Transgenic Donor(hDAF/hCD59) 2.0 Dagget et al./1997
4 Complement Modulation(sCR1) 0.6 Azimzadeh et al./2003
5 Complement Modulation(heat inactivated) 0.5 Pierson et al./1997
6 Complement Modulation(K76–COOH) 0.6 Blum et al./1998
5 Complement Modulation(FUT�175) 0.8 Blum et al./1998
6 Coagulation Modulation(heparin) <1.0 Schelzig et al./2002
6 Coagulation Modulation(rHirudin) 3.0 Schelzig et al./2002
6 Transgenic Donor(hDAF)/Complement Modulation(heat inactivated) 5.5 Azimzadeh et al./2003
3 Ab Depletion(Ag incubation)/Complement Modulation(heat inactivated) 3.5 Pierson et al./1997
Pulmonary xenotransplantation
American Journal of Transplantation 2004; 4 (Suppl. 6): 25–35 29

complement regulation by hCRPs is insufficient during
pulmonary xenotransplantation.
Some evidence suggests that pulmonary xenografts may
be uniquely sensitive to complement activation, particu-
larly the anaphylatoxin C5a. Of particular interest are
experiments using cobra venom factor (CVF), a molecule
which depletes complement but produces substantial
amounts of anaphylatoxins, including C5a. Yeatman and
associates, in a pig to baboon single orthotopic lung trans-
plantation model, were only able to show partial protection
from pulmonary xenograft injury in CVF (Quidel, San
Diego, CA) pretreated baboons (21). Using an isolated
working heart–lung perfusion system, Pierson and col-
leagues perfused swine lungs with human blood in order
to determine the mechanisms of pulmonary xenograft dys-
function. Pretreatment of human blood with CVF was
unsuccessful in preventing the development of acute
lung injury (19), consistent with the idea that anaphylatoxin
may injure pulmonary xenografts. Other evidence that
anaphylatoxins may be important in pulmonary xenograft
dysfunction comes from the efficacy of anti-C5a mono-
clonal antibodies (rC5a, Sigma, St.Louis, MO) in prolong-
ing pulmonary xenograft function, although the effect is
short lived (<12 h survival; unpublished data). Although
the lack of efficacy of anticomplement therapy could be
due to an inadequate complement control, other lines
of evidence described below indicate that factors other
than complement play a key role in pulmonary xenograft
dysfunction.
Coagulopathy and Pulmonary XenograftDysfunction
Disseminated intravascular coagulation (DIC) has recently
been proposed as the ‘third’ barrier to xenotransplantation
(22), with xenoreactive antibodies and complement
incompatibility being the first two barriers. DIC is asso-
ciated with the chronic rejection of kidney xenografts
(23,24), developing over a period of weeks in these xeno-
grafts. DIC is also associated with acute processes follow-
ing bone marrow xenotransplantation (25), developing
over a period of days following bone marrow xenotrans-
plantation. In contrast, DIC is observed within minutes to
hours of pulmonary xenograft reperfusion (7).
Several incompatibilities between the human and porcine
coagulation systems have been identified as potential
underlying causes of the coagulopathy associated with
pulmonary xenotransplantation. One potential incom-
patibility is that porcine von Willebrand Factor (vWF) inter-
acts in an aberrant fashion with human platelets. Von
Willebrand Factor is a protein stored by platelets and
endothelial cells that is released upon activation of those
cells (26–28). In normal individuals, platelet activation and
adhesion occurs when vWF binds to GPIb on platelets
only if the platelets are subjected to shear stress (29–
34). In contrast, porcine von Willebrand Factor (pvWF)
binds human (or primate) GPIb on quiescent platelets,
leading to platelet aggregation even in the absence of
shear stress (35,36). Thus, aberrant interactions between
Table 2: In vivo pulmonary xenotransplantation experience. Antibody depletion was performed using various strategies prior to
transplant: (1) baboon-to-pig lung perfusion (lung perfusion); (2) column plasmapheresis (column); or (3) aGal conjugated to polyethelene
glycol (aGal-PEG). Transgenic donors were of several types: (1) human membrane cofactor protein (hMCP); (2) human decay accelerating
factor (hDAF); (3) human CD59 (hCD59); or (4) combinations thereof. Macrophage depletion strategies utilized an encapsulated heavy
metal chelator (clodronate), which selectively destroys macrophages by induction of apoptosis
Donor Recipient n Treatment Survival (h) Reference
Swine Baboon 3 None <3 Yeatman et al./1999
3 None <3 Dagget et al./1998
2 None <1 Norin et al./1996
6 None <72 Kaplon et al./1995
3 None <3 Lau et al./2003
3 CVF <1 Yeatman et al./1999
4 Ab Depletion(lung perfusion) 11 Dagget et al./1998
4 Transgenic Donor (hDAF/CD59) 3 Yeatman et al./1999
4 Transgenic Donor(hDAF/CD59) 3 Lau et al./2000
3 Transgenic Donor(hMCP) <2 Gaca et al./2002
4 Transgenic Donor(hMCP) 12 Gonzalez-Stawinski et al./2002
3 Transgenic Donor(hCD59) <12 Norin et al./1996
2 vWFd <3 Lau et al./2003
2 Transgenic Donor(hDAF/CD59)/Ab Depletion(column) <1 Lau et al./2000
1 Transgenic Donor(hMCP)/Ab Depletion(lung perfusion) <1 Gaca et al./2002
4 Transgenic Donor(hMCP)/Ab Depletion(column) 4 Gonzalez-Stawinski et al./2002
5 Transgenic Donor(hMCP)/Ab Depletion(aGAL–PEG) 4 Gonzalez-Stawinski et al./2002
3 Transgenic Donor(hMCP)/antiGP1b <3 Gaca et al./2002
1 Transgenic Donor(hMCP)/Ab Depletion(column)/antiC5a 9 Gaca et al./2002
5 Transgenic Donor(hMCP)/Ab Depletion(aGAL–PEG)/PIM depletion(Clodronate) 22.6 Unpublished
3 vWFd Donor/Ab Depletion(aGAL–PEG)/PIM depletion(Clodronate) 67 Unpublished
Edward Cantu et al.
30 American Journal of Transplantation 2004; 4 (Suppl. 6): 25–35

pvWF and human or primate GPIb could lead to wide-
spread activation of the coagulation system, resulting in
DIC.
The interaction between pvWF and human GPIb may be
particularly important in pulmonary xenotransplantation, as
evidence suggests that pulmonary xenografts shed more
pvWF than either heart or kidney xenografts (10). As a
direct test of this idea, the interaction between pvWF
and primate GPIb in the DIC associated with pulmonary
xenotransplantation was blocked in a series of swine-
to-baboon pulmonary xenotransplants using a monoclonal
antibody to GPIb (monoclonal Lp-J3) (37). Treatment with
the anti-GPIb antibody resulted in a loss of platelets,
through splenic sequestration. Treatment with anti-GPIb
antibody in splenectomized baboons (normal number of
platelets) and in baboons with intact spleens (elimination
of platelets) prevented the decreases in fibrinogen and
increases in D-dimers observed in control xenotransplants,
indicating that activation of platelets by porcine vWF plays
a significant role in the DIC associated with pulmonary
xenograft dysfunction.
On the other hand, elevations in the PT and generation of
thrombin were not prevented by anti-GPIb therapy (37),
suggesting that activation of the coagulation system
occurs by means other than or in addition to the interac-
tion of pvWF with platelets. Indeed, there are at least two
other molecular incompatibilities between porcine and
human proteins that might cause or at least contribute to
coagulopathy. Porcine TFPI, which is present in lung tis-
sues (38), and is a modulator of thrombosis in the lung
(39), does not inhibit human factor Xa (40,41). Further,
porcine TM does not activate the anticoagulant human
(or primate) protein C (42). Thrombomodulin may be pre-
sent in lung tissues to a greater extent than in other
tissues (43), suggesting that problems associated with
the ineffectiveness of porcine TM may be especially pro-
nounced in pulmonary xenotransplantation compared to
heart and kidney xenotransplantation.
Role of Pulmonary IntravascularMacrophages in Pulmonary XenograftDysfunction
Swine lungs differ from primate lungs because, in addition
to the resident pulmonary alveolar macrophages, there are
pulmonary intravascular macrophages (PIMs) that com-
prise more than 16% of microvascular surface area (44).
Importantly, these pulmonary intravascular macrophages
(PIMs) are largely absent in humans and primates. PIMs
are a resident population of macrophages, morphologically
resembling Kupffer cells, which are tightly adherent to the
endothelium by cell junctions and are not readily dis-
placed. Whether they can be induced in humans or
primates remains controversial.
The physiologic function of these cells is to filter blood of
any foreign material or bacteria passing through the lung, a
process, which normally takes place in the liver or spleen
in species without large populations of PIMs (45–48). In
addition to their scavenging function, PIMs, produce
arachidonate metabolites including thromboxane, cytokines
including IL-1, IL-2, and TNF-a, and procoagulant factors,
including tissue factor and PAI-1 (48–50). In acute lung
injury and some xenotransplants models evidence sug-
gests PIMs may contribute significantly to the rapid devel-
opment of pulmonary hypertension and edema (45,47,51).
Using a technique to deplete macrophages using lipo-
somal clodronate (Roche, Mannheim, Germany) developed
by Van Rooijen (52,53), Staub was able to reliably deplete
>90% of the PIM population in sheep (54). In the PIM
depleted sheep, there was no pulmonary vasoconstrictive
response to particle infusion. In PIM depleted vs. non-
depleted sheep, LPS infusion resulted in complete abroga-
tion of increases in PVR and >90% attenuation of the
capillary leak (54,55).
Pierson and colleagues using lungs from swine treated with
liposomal clodronate for their heterologous perfusion experi-
ments, found decreased production of thromboxane and
preservation of a PVR comparable to the homologous con-
trol perfusion. In addition, PIM depletion decreased platelet
sequestration, C3a levels, TNF-a release, and prolonged
pulmonary function (50). More recently, Gaca and associ-
ates using an in vivo pig to primate orthotopic lung transplant
model with PIM depletion have been able to abrogate the
consumptive coagulopathy seen in controls and extend graft
function to 24 h (Figure 3, data unpublished).
Hypothetical Barriers
There are a number of hypothetical barriers to pulmonary
xenotransplantation. Perhaps most concerning is a
hypothetical problem common to all xenografts – xoono-
sis. Considerable attention has been placed on the pos-
sibility that infectivity of transplanted pig organs may place
the community as a whole at risk (5). Although cross-
species transmission of diseases has and continues to
be a health problem, the transmission of disease in pig-
to-human transplantation is a hypothetical problem. Many
consider that following strict housing and breeding guide-
lines would make the risk of transferring known swine
infections less likely (56). Further, selective breeding or
gene targeting strategies to further reduce the risk of
known infections are likely possible (57). Thus, given our
current level of understanding, community risk of known
infections is minimal (5); however, it is also recognized
that a cavalier attitude is inappropriate, and that we should
proceed with caution (58–62).
Another hypothetical problem is that physiologic incompat-
ibility may pose a barrier to pulmonary xenotransplantation.
Pulmonary xenotransplantation
American Journal of Transplantation 2004; 4 (Suppl. 6): 25–35 31

Although porcine organs have been able to support
primates without the assistance of a native lung or other
means of oxygenation (63), there are some known physio-
logic differences between porcine and human lungs. For
example, porcine lungs contain less substantial collateral
channels and more musculature in the arterial walls than
do human lungs (64). This should make pulmonary hyper-
tension more of a concern in xenografts than in allografts.
On the other hand, because collateral resistance is much
greater than airway resistance, the collateral pathways do
not apparently play a ‘significant role’ under normal physio-
logic circumstances, even in humans (64). Thus, it remains
unknown what effect a decrease in collateral pathways
might have on a xenograft recipient.
Another hypothetical concern regarding pulmonary xeno-
transplantation is that, being an immune organ, the lung
may itself need to be substantially immunosuppressed to
facilitate engraftment, and that such immunosuppression
may lead to infection of the organ. The fact that elimination
of PIMs from the organ dramatically improves xenograft
function substantiates the idea that immunosuppression
of the organ may be required. This hypothetical problem
will likely materialize or be dismissed as the duration of
xenograft survival in clinically relevant models is extended.
Yet another hypothetical barrier to pulmonary xenotrans-
plantation is one that has already been encountered in
heart and kidney xenotransplantation. Specifically, the
recipient may mount a T-cell dependent response to the
graft, resulting in the production of high affinity IgG
specific for the porcine tissue. It is hoped that, if indeed
this barrier is encountered, therapies currently showing
promise in heart and kidney xenotransplantation will prove
equally successful in pulmonary xenotransplantation.
Future Directions
The hurdles to the application of clinical pulmonary xeno-
transplantation are substantial, and even though signifi-
cant advances have been made, it seems apparent that
work toward that application is only beginning. Preliminary
results from our group indicate that depletion of PIMs
combined with attenuation of xenoreactive antibodies
and elimination of porcine vWF (vWF-deficient swine
bred at the University of North Carolina, Chapel Hill, NC)
can prolong pulmonary xenograft survival up to five days,
even in the absence of complement inhibition. Although
the etiology of graft failure in the absence of PIMs appears
similar to the more rapid graft failure observed in the
presence of the PIMs, the mechanisms underlying this
failure remain unknown. The failure may be due, at least
in part, to PIMs which were either not depleted or which
are regenerated. In addition, factors such as complement
activation and problems with coagulation regulation prob-
ably play a role in the failure of PIM-depleted lungs just as
in PIM-sufficient lungs. However, the presence of other
barriers has not been ruled out and future studies will
necessarily address this issue. In addition, the ability of
lungs depleted of PIMs to withstand infection will need to
be addressed.
Inevitably, experiments aimed at achieving tolerance in pul-
monary xenotransplantation will take place. However, suc-
cess in this area is not a short-term goal. Work toward
tolerance to porcine kidney xenografts is progressing but
is facing serious hurdles (65–67). It is anticipated that
work in this area with porcine lungs may prove even
more challenging, given that the lung is, like the gut, an
immune organ.
Future studies will necessarily address therapies to over-
come the problems with coagulation regulation in pulmon-
ary xenotransplantation. Although vWF deficiency has
proven effective in our model, the ultimate goal is to
block aberrant interactions between porcine vWF and
gp1b without eliminating the normal function of vWF.
Achieving this goal may provide us with insight into other
problems with coagulation regulation that need to be
addressed. Importantly, if early events unrelated to coagu-
lation initiate the associated DIC, the complex nature of
the DIC may be less important from a clinical perspective
if the early events can be blocked. Thus, future studies in
pulmonary xenotransplantation should be aimed not only
at understanding the factors that contribute directly to
DIC, but also to factors that potentially initiate the process.
The future of xenotransplantation is inexorably tied to the
development of genetic engineering in the pig. Cloning of
pigs has recently been achieved and provides new and
promising avenues of investigation. Healthy galactosyl a1–3galactosyl transferase deficient swine have been born
earlier this year (68), and this technology may now be
applied to other molecules (69). Gene transfer technolo-
gies in pigs have recently advanced, allowing for the ability
to either knock out or knock in genes of interest quicker,
with greater efficiency, and with less expense (70–76).
Our increasing ability to create genetically modified pigs,
coupled with our rapidly improving understanding of the
immunology surrounding pulmonary xenograft dysfunc-
tion, fuel the hope that pulmonary xenotransplantation
will one day be a viable option for patients with end
stage pulmonary disease.
Conclusion
Substantial progress has been made in the last 10 years
regarding the role of xenoreactive antibodies, comple-
ment, coagulopathy and, more recently, pulmonary intra-
vascular macrophages in the pathologic milieu associated
with pulmonary xenotransplantation. Although this review
has treated each of these factors separately, the interplay
between them is certainly an integral part of the patholo-
gical process (Figure 4). For example, porcine vWF shed
Edward Cantu et al.
32 American Journal of Transplantation 2004; 4 (Suppl. 6): 25–35

from pulmonary xenografts may play a key role not only in
the coagulopathy and DIC associated with pulmonary
xenotransplantation, but, when complexed with xenoreac-
tive antibodies, may be a significant factor in the produc-
tion of C5a. This anaphylatoxin, in turn, may interact with
C5a receptors present on macrophages (77) thus leading
to macrophage stimulation and subsequent increases in
the proinflammatory and procoagulant milieu (78).
Despite this progress, the limited survival of pulmonary
xenografts that is currently observed in large animal
models means that the clinical application of pulmonary
xenotransplantation is not yet in sight, and, although none
can foresee what lies around the next corner, much addi-
tional work is required.
References
1. Hardy JDWW, Dalton ML. Lung homotransplantation in man.
JAMA 1963; 186: 1065–1069.
2. Hertz MI, Taylor DO, Trulock EP et al. The Registry of the Inter-
national Society for Heart and Lung Transplantation: Nineteenth
Official Report 2002. J Heart & Lung Transplant 2002; 21:
950–970.
3. UNOS. Annual Report. Richmond, VA; US Department of Health
and Human Services, 2001.
4. Lambrigts D, Sachs DH, Cooper DK. Discordant organ xeno-
transplantation in primates: world experience and current status.
Transplantation 1998; 66: 547–561.
5. Cooper DK, Keogh AM, Brink J et al. Report of the
Xenotransplantation Advisory Committee of the International
Society for Heart and Lung Transplantation: the present status
of xenotransplantation and its potential role in the treatment of
end-stage cardiac and pulmonary diseases. J Heart & Lung
Transplant 2000; 19: 1125–1165.
6. Bryant LR, Eiseman B, Avery M. Studies of the porcine lung as
an oxygenator for human blood. J Thoracic & Cardiovascular
Surgery 1968; 55: 255–263.
7. Gaca JG, Lesher A, Aksoy O et al. Disseminated intravascular
coagulation in association with pig-to-primate pulmonary
xenotransplantation. Transplantation 2002; 73: 1717–1723.
8. Macchiarini P, Mazmanian GM, Oriol R et al. Ex vivo lung model
of pig-to-human hyperacute xenograft rejection. J Thorac &
Cardiovasc Surg 1997; 114: 315–325.
9. Kamholz SL, Brewer RJ, Grijalva G et al. Laboratory studies in
cross-species lung transplantation. World J Surgery 1997; 21:
951–955.
10. Holzknecht ZE, Coombes S, Blocher BA et al. Immune complex
formation after xenotransplantation: evidence of type III as well
as type II immune reactions provide clues to pathophysiology.
Am J Pathol 2001; 158: 627–637.
11. Pierson RN, 3rd, Kasper-Konig W, Tew DN et al. Hyperacute lung
rejection in a pig-to-human transplant model: the role of anti-pig
antibody and complement. Transplantation 1997; 63: 594–603.
12. Macchiarini P, Oriol R, Azimzadeh A et al. Evidence of human
non-alpha-galactosyl antibodies involved in the hyperacute
rejection of pig lungs and their removal by pig organ perfusion.
J Thoracic & Cardiovascular Surgery 1998; 116: 831–843.
13. Lau CL, Daggett WC, Yeatman MF et al. The role of antibodies in
dysfunction of pig-to-baboon pulmonary transplants. J Thoracic
& Cardiovascular Surgery 2000; 120: 29–38.
14. Gaca JG, Lee W, Aksoy O et al. Evidence of polyreactive xenor-
eactive antibodies in the repertoire of human anti-swine lung
antibodies: The ‘next’ humoral barrier to xenotransplantation?
Transplant Immunol 2001; 9: 19–27.
15. Lee W, Gaca JG, Braedehoeft SJ, Parker W, Davis RD. Binding
of polyreactive antibodies to self vs. foreign antigens. Immuno-
biology 2001.
16. Dalmasso AP, Vercellotti GM, Platt JL, Bach FH. Inhibition of
complement-mediated endothelial cell cytotoxicity by decay-
accelerating factor. Potential for prevention of xenograft
hyperacute rejection. Transplantation 1991; 52: 530–533.
17. Hourcade D, Holers VM, Atkinson JP. The regulators of comple-
ment activation (RCA) gene cluster. Advances in Immunology
1989; 45: 381–416.
18. Daggett CW, Yeatman M, Lodge AJ et al. Swine lungs expres-
sing human complement-regulatory proteins are protected
against acute pulmonary dysfunction in a human plasma perfusion
model. J Thorac & Cardiovasc Surg 1997; 113: 390–398.
19. Pierson RN, 3rd, Dunning JJ, Konig WK et al. Mechanisms
governing the pace and character of pig heart-lung rejection by
human blood. Transplantation Proceedings 1994; 26: 2337.
20. Daggett CW, Platt JL, Davis RD. The expression of human
membrane cofactor protein is protective in swine-to-primate
pulmonary xenotransplantation. 4th Annual International
Congress of the Society of Xenotransplantation 1997. (Abstract)
21. Yeatman M, Daggett CW, Parker W et al. Complement-
mediated pulmonary xenograft injury: studies in swine-to-
primate orthotopic single lung transplant models. Transplantation
1998; 65: 1084–1093.
22. D’Apice AJ, Cowan PJ. Profound coagulopathy associated with
pig-to-primate xenotransplants: How many transgenes will be
required to overcome this new barrier? Transplantation 2000;
70: 1273–1274.
23. Ierino FL, Kozlowski T, Siegel JB et al. Disseminated intravas-
cular coagulation in association with the delayed rejection of
pig-to-baboon renal xenografts. Transplantation 1998; 66:
1439–1450.
24. Cowan PJ, Aminian A, Barlow H, et al. Renal xenografts from
triple-transgenic pigs are not hyperacutely rejected but cause
coagulopathy in non-immunosuppressed baboons. Transplant-
ation 2000; 69: 2504–2515.
25. Buhler L, Basker, M., Alwayn, I.P et al. Coagulation and
thrombotic disorders associated with pig organ and hematopoietic
cell transplantation in nonhuman primates. Transplantation 2000;
70: 1323–1331.
26. Coller BS, Hirschman RJ, Gralnick HR. Studies on the Factor VIII/
von Willebrand factor antigen on human platelets. Thrombosis
Research 1975; 6: 469–480.
27. Wagner DD, Marder VJ. Biosynthesis of von Willebrand protein
by human endothelial cells: processing steps and their intra-
cellular localization. J Cell Biol 1984; 99: 2123–2130.
28. Pareti FI, Fujimora Y, Dent JA, Holland LZ, Zimmerman TS,
Ruggeri ZM. Isolation and characterization of a collagen binding
domain in human von Willebrand factor. J Biol Chem 1986; 261:
15310–15315.
29. O’Brien JR. Shear induced platelet aggregation. Lancet 1990;
335: 711–713.
30. Howard MA, Firkin BG. Ristocetin: a new tool in the investigation
of platelet aggregation. Thrombosis et diathesis haemorrhagica
1971; 26: 362–369.
31. Inbal A, Loscalo J.Glycocalicin binding to von Willebrand Factor
adsorbed onto collagen-coated or polystyrene surfaces. Throm-
bosis Research 1989; 56: 347–357.
Pulmonary xenotransplantation
American Journal of Transplantation 2004; 4 (Suppl. 6): 25–35 33

32. Kroll MH, Harris TS, Moake JL, Handin RI, Schafer AI. Von
Willebrand factor binding to platelet GPIb initiates signals for
platelet activation. J Clin Investigation 1991; 88: 1568–1573.
33. Goto S, Salomon DR, Ikeda Y, Ruggeri ZM. Characterization of
the unique mechanism mediating the shear-dependent binding
of soluble von Willebrand factor to platelets. J Biol Chem 1995;
270: 23352–23361.
34. Ruggeri ZM. Structure and function of von Willebrand factor.
Thrombosis & Haemostasis 1999; 82: 576–584.
35. Mazzucato M, De Marco L, Pradella P, Masotti A, Pareti FI.
Porcine von Willebrand factor binding to human platelet GPIb
induces transmembrane calcium influx. Thrombosis &
Haemostasis 1996; 75: 655–660.
36. Pareti FI, Mazzucato M, Bottini E, Mannucci PM. Interaction of
porcine von Willebrand factor with the platelet glycoproteins Ib
and IIb/IIIa complex. Br J Haematol 1992; 82: 81–86.
37. Gaca JG, Lesher A, Aksoy O et al. The role of porcine von
Willebrand factor – baboon platelet interactions in disseminated
intravascular coagulation associated with pulmonary xenotrans-
plantation. 2002; 74: 1596–1603.
38. Bajaj MS, Kuppuswamy MN, Manepalli AN, Bajaj SP. Transcrip-
tional expression of tissue factor pathway inhibitor, thrombo-
modulin and von Willebrand factor in normal human tissues.
Thrombosis & Haemostasis 1999; 82: 1047–1052.
39. Fujii M, Hayakawa H, Urano T et al. Relevance of tissue factor
and tissue factor pathway inhibitor for hypercoagulable state
in the lungs of patients with idiopathic pulmonary fibrosis.
Thrombosis Research 2000; 99: 111–117.
40. Nagayasu T, Saadi S, Holzknecht RA, Plummer TB, Platt JL.
Expression of tissue factor mRNA in cardiac xenografts: clues
to the pathogenesis of acute vascular rejection. Transplantation
2000; 69: 475–482.
41. Kopp CW, Siegel JB, Hancock WW et al. Effect of porcine
endothelial tissue factor pathway inhibitor on human coagulation
factors. Transplantation 1997; 63: 749–758.
42. Siegel JB,GreyST,LesnikoskoBAet al. Xenogeneicendothelial cells
activate human prothrombin. Transplantation 1997; 64: 888–896.
43. DeBault LE, Esmon NL, Olson JR, Esmon CT. Distribution of the
thrombomodulin antigen in the rabbit vasculature. Lab Invest
1986; 54: 172–178.
44. Winkler GC, Cheville NF. Postnatal colonization of porcine lung
capillaries by intravascular macrophages: an ultrastructural, mor-
phometric analysis. Microvascular Research 1987; 33: 224–232.
45. Staub NC. Pulmonary intravascular macrophages. Ann Rev
Physiol 1994; 56: 47–67.
46. Brain JD, Molina RM, DeCamp MM, Warner AE. Pulmonary intra-
vascularmacrophages: their contribution to themononuclear phago-
cyte system in 13 species. Am J Physiol 1999; 276: L146–L154.
47. Miyamoto K, Schultz E, Heath T et al. Pulmonary intravascular
macrophages and hemodynamic effects of liposomes in sheep.
J Applied Physiol 1988; 64: 1143–1152.
48. Chitko-McKown CG, Chapes SK, Brown RE, et al. Porcine
alveolar and pulmonary intravascular macrophages: comparison
of immune functions. J Leukocyte Biol 1991; 50: 364–372.
49. Bertram TA, Overby LH, Danilowicz R et al. Pulmonary intravas-
cular macrophages produce prostaglandins and leukotrienes in
vitro. Chest 1988; 93: 82S–84S.
50. Collins BJ, Blum MG, Parker RE et al. Thromboxane mediates
pulmonary hypertension and lung inflammation during hyperacute
lung rejection. J Applied Physiol 2001; 90: 2257–2268.
51. Tector AJ, Fridell JA, Watanabe TJ et al. Pulmonary injury in
recipients of discordant hepatic and renal xenografts in the
dog-to-pig model. Xenotransplantation 1998; 5: 44–49.
52. van Rooijen N. The liposome-mediated macrophage ‘suicide’
technique. J Immunol Methods 1989; 124: 1–6.
53. van Rooijen N. Liposome-mediated elimination of macrophages.
Research in Immunol 1992; 143: 215–219.
54. Sone Y, Nicolaysen A, Staub NC. Effect of particles on sheep
lung hemodynamics parallels depletion and recovery of intravas-
cular macrophages. J Applied Physiol 1997; 83: 1499–1507.
55. Sone Y, Serikov KB, Staub NC. Intravascular macrophage
depletion attenuates endotoxin in lung injury in anesthetized
sheep. J Applied Physiol 1999; 87: 1354–1359.
56. Onions D, Cooper DK, Alexander TJ et al. An approach to the
control of disease transmission in pig-to-human xenotransplant-
ation. Xenotransplantation 2000; 7: 143–155.
57. Blusch JH, Patience C, Martin U. Pig endogenous retroviruses
and xenotransplantation. Xenotransplantation 2002; 9: 242–251.
58. Bach FH, Fineberg HV. Call for moratorium on xenotransplants.
Nature 1998; 391: 326.
59. Bach FH, Fishman JA, Daniels N et al. Uncertainty in xenotrans-
plantation: individual benefit versus collective risk. Nat Medical
1998; 4: 141–144.
60. Chapman LE, Folks TM, Salomon DR, Patterson AP,
Eggerman TE, Noguchi PD. Xenotransplantation and xenogeneic
infections. N Engl J Med 1995; 333: 1498–1501.
61. Fishman JA. Xenosis and xenotransplantation: addressing the
infectious risks posed by an emerging technology. Kidney Intern
1997; 58: S41–S45.
62. Stoye JP, Coffin JM. The dangers of xenotransplantation. Nat
Medical 1995; 1: 1100.
63. Daggett CW, Yeatman M, Lodge AJ et al. Total respiratory
support from swine lungs in primate recipients. J Thorac &
Cardiovasc Surg 1998; 115: 19–27.
64. Delaunois L. Anatomy and physiology of collateral respiratory
pathways. Eur Respir J 1989; 2: 893–904.
65. Sachs DH. Mixed chimerism as an approach to transplantation
tolerance. Clin Immunol 2000; 95: S63–S68.
66. Sablinski T, Gianelloi PR, BailinM et al. Pig tomonkey bonemarrow
and kidney xenotransplantation. Surgery 1997; 121: 381–391.
67. Tanaka M, Latinne D, Gianello P et al. Xenotransplantation from
pig to cynomolgus monkey: the potential for overcoming xeno-
graft rejection through induction of chimerism. Transplantation
Proceedings 1994; 26: 1326–1327.
68. Phelps CJ, Koike C, Vaught TD et al. Production of alpha 1,3-
galactosyltransferase-deficient pigs. Science 2003; 299: 411–414.
69. Cooper DK. Sixth Congress of the International Xenotransplant-
ation Association. Xenotransplantation 2003; 10: 7–9.
70. Polejaeva IA CS, Vaught TD, Page RL. Cloned pigs produced by
nuclear transfer from adult somatic cells. Nature 2000; 407: 86–89.
71. Wilmut I, Beaujean N, de Sousa PA et al. Somatic cell nuclear
transfer. Nature 2002; 419: 583–586.
72. Dinnyes A, De Sousa P, King T et al. Somatic cell nuclear transfer.
recent progress and challenges. Cloning StemCells 2002; 4: 81–90.
73. Hawley RJ. Genetic modification of pigs by nuclear transfer.
Xenotransplantation 2002; 9: 159–160.
74. Dai Y, Vaught TD, Boone J et al. Targeted disruption of the
alpha1,3-galactosyltransferase gene in cloned pigs. Nat Biotech-
nol 2002; 20: 251–255.
75. De Sousa PA, Dobrinsky JR, Zhu J et al. Somatic cell nuclear
transfer in the pig: control of pronuclear formation and integra-
tion with improved methods for activation and maintenance of
pregnancy. Biol Reprod 2002; 66: 642–650.
76. Kaiser J. Xenotransplantation. Cloned pigs may help overcome
rejection. Science 2002; 295: 25–27.
Edward Cantu et al.
34 American Journal of Transplantation 2004; 4 (Suppl. 6): 25–35

77. Haynes DR, Harkin DG, Bignold LP, Hutchens MJ, Taylor SM,
Fairlie DP. Inhibition of C5a-induced neutrophil chemotaxis and
macrophage cytokine production in vitro by a new C5a receptor
antagonist. Biochemical Pharmacology 2000; 60: 729–733.
78. Fine R, Shaw JO, Rogers WR. Effects of C5a on baboon
alveolar macrophage migration. Am Rev Resp Dis 1981; 123:
110–114.
79. Azimzadeh A, Zorn GL, 3rd, Blair KS et al. Hyperacute lung
rejection in the pig-to-human model. 2. Synergy between soluble
and membrane complement inhibition. Xenotransplantation
2003; 10: 120–131.
80. Wiebe K, Steinhoff G, Poeling J et al. Ex vivo perfusion of
swine lungs: lung function in a pig-to-human model of
xenotransplantation. Transplantation Proceedings 2000; 32:
1149–1150.
81. Kulick DM, Salerno CT, Dalmasso AP et al. Transgenic swine
lungs expressing human CD59 are protected from injury in a pig-
to-human model of xenotransplantation. J Thorac & Cardiovasc
Surg 2000; 119: 690–699.
82. Yeatman M, Daggett CW, Parker WW et al. Complement-
mediated pulmonary injury following heterologous perfusion of
swine lungs with human plasma. 1999.
83. Macchiarini P, Mazmanian GM, Oriol R et al. Ex vivo lung model
of pig-to-human hyperacute xenograft rejection. J Thorac &
Cardiovasc Surg 1997; 114: 315–325.
84. Blum MG, Collins BJ, Chang AC, Zhang JP, Knaus SA, Pierson
RN, 3rd. Complement inhibition by FUT-175 and K76-COOH in a
pig-to-human lung xenotransplant model. Xenotransplantation
1998; 5: 35–43.
85. Macchiarini P, Oriol R, Azimzadeh A et al. Evidence of human
non-alpha-galactosyl antibodies involved in the hyperacute rejec-
tion of pig lungs and their removal by pig organ perfusion. J
Thorac & Cardiovasc Surg 1998; 116: 831–843.
86. Schelzig H, Vogel A, Krischer C, Simon F, Abendroth D. Role of
recombinant hirudin in a pig-to-human lung transplantation
model. Transplantation Proceedings 2002; 34: 2384–2386.
87. Yeatman M, Daggett CW, Lau CL et al. Human complement
regulatory proteins protect swine lungs from xenogeneic injury.
Annals of Thoracic Surgery 1999; 67: 769–775.
88. Daggett CW, Yeatman M, Lodge AJ, et al. Total respiratory
support from swine lungs in primate recipients. J Thorac &
Cardiovasc Surg 1998; 115: 19–27.
89. Norin AJ, Brewer RJ, Lawson N et al. Enhanced survival of
porcine endothelial cells and lung xenografts expressing human
CD59. Transplantation Proceedings 1996; 28: 797–798.
90. Kaplon RJ, Platt JL, Kwiatkowski PA et al. Absence of
hyperacute rejection in pig-to-primate orthotopic pulmonary
xenografts. Transplantation. 1995; 59: 410–416.
91. Lau CL, Cantu III E, Gonzalez-Stawinski GV et al. The Role of
Antibodies and Von Willebrand Factor in Discordant Pulmonary
Xenotransplantation. Am J Trans 2003; 3: 1065–1075.
92. Lau CL, Daggett WC, Yeatman MF et al. The role of antibodies in
dysfunction of pig-to-baboon pulmonary transplants. J Thorac &
Cardiovasc Surg, 120: 29–38.
93. Gonzalez-Stawinski GV, Daggett CW, Lau CL et al. Non-anti-Gal
alpha 1-3 Gal antibody mechanisms are sufficient to cause
hyperacute lung dysfunction in pulmonary xenotransplantation.
J Am Coll Surg 2002; 194: 765–773.
Pulmonary xenotransplantation
American Journal of Transplantation 2004; 4 (Suppl. 6): 25–35 35