Embed Size (px)
Citation preview

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 329, No. 1, May 1, pp. 9–16, 1996Article No. 0185
Purification and Characterization of an NADPH-CytochromeP450 (Cytochrome c) Reductase from Spearmint(Mentha spicata) Glandular Trichomes1
Krishan Ponnamperuma and Rodney Croteau2
Institute of Biological Chemistry and Department of Biochemistry and Biophysics,Washington State University, Pullman, Washington 99164-6340
Received November 21, 1995, and in revised form February 13, 1996
portant plant natural products such as phenylpropa-Solubilized NADPH-cytochrome c (P450) reductase noids, alkaloids, and terpenoids. In addition to their
was purified to homogeneity from an extract of spear- role in the biosynthesis of natural products, plant P450mint (Mentha spicata) glandular trichomes by dye- cytochromes also participate in the catabolism of herbi-ligand interaction chromatography on Matrex-Gel cides and other xenobiotic compounds (1). Our interestRed A and affinity chromatography on 2 *,5*-adenosine in this family of catalysts stems from their involvementdiphosphate agarose. SDS–PAGE of the purified en- in terpenoid metabolism, particularly the role of regio-zyme preparation revealed the presence of two simi- specific cytochrome P450 (0)-limonene hydroxylases inlar proteins with masses of 82 kDa (major) and 77 kDa essential oil formation in the glandular trichomes (oil(minor) that crossreacted on immunoblot analysis glands) of peppermint (Mentha piperita), native spear-with polyclonal antibodies directed against NADPH- mint (M. spicata), and Scotch spearmint (M. gracilis)cytochrome P450 reductase from Jerusalem arti-
and their various mutants (Fig. 1) (2–6).choke and from mung bean. Complete immunoinhibi-The cytochrome P450 hydroxylase system consiststion of reductase activity was observed with both
of an integral membrane multienzyme complex of thetypes of polyclonal antibodies, while only partial inhi-substrate binding hemoprotein cytochrome P450 andbition of activity resulted using a family of mono-the FAD/FMN containing flavoprotein NADPH-cyto-clonal antibodies directed against the Jerusalem arti-chrome P450 reductase. Several NADPH-P450 reduc-choke cytochrome P450 reductase. Inhibition of thetases have been purified to homogeneity from mamma-spearmint oil gland cytochrome c reductase was alsolian (7–11), microbial (12–14), and insect (15, 16)observed with the diphenyliodonium ion. The Km val-
ues for the cosubstrates NADPH and cytochrome c sources that are capable of reconstituting functionalwere 6.2 and 3.7 mM, respectively, and the pH optimum P450-dependent monooxygenase activities. cDNA clon-for activity was at 8.5. The NADPH-cytochrome c re- ing has been carried out and the deduced primary se-ductase reconstituted NADPH-dependent (0)-4S-lim- quences have been reported for rat (17), rabbit (18), pigonene-6-hydroxylase activity in the presence of cyto- (19), house fly (20), yeast (21, 22), and bacterial (23)chrome P450, purified from the microsomal fraction reductases. Cytochrome P450 reductases have alsoof spearmint oil gland cells and dilauroyl phosphati- been isolated from several higher plant sources; how-dyl choline. These characteristics establish the iden- ever, deduced primary sequences have been describedtity of the purified enzyme as a NADPH-cytochrome
only for the enzymes from mung bean (24) and Catha-P450 reductase. q 1996 Academic Press, Inc.ranthus roseus (25).
The first reported purification of a plant cytochromeCytochrome P450-dependent hydroxylases play a P450 reductase was from etiolated Catharanthus ro-
pivotal role in the biosynthesis of a wide range of im- seus seedlings, and this enzyme reconstituted C10 hy-droxylation of geraniol as an intermediate step in iri-
1 This investigation was supported in part by National Science doid monoterpene biosynthesis (26). NADPH-cyto-Foundation Grant IBN-9104983 and Hatch Project 0268 from the chrome c reductases have been isolated from sweetAgricultural Research Center, Washington State University.
potato (27), cell suspension cultures of Catharanthus2 To whom correspondence should be addressed. Fax: (509) 335-7643. roseus (28), and petunia flowers (29); the former was
90003-9861/96 $18.00Copyright q 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
/ 6b17$$9407 03-28-96 23:24:44 arca AP: Archives

10 PONNAMPERUMA AND CROTEAU
P450 reductase was a generous gift from Dr. Ronald Estabrook (Uni-versity of Texas Southwestern Medical Center, Dallas, TX). Rabbitpolyclonal antibodies (immune serum and IgG) and rat monoclonalantibodies (AD(3)3 and C5A5) directed against Jerusalem artichokeNADPH-cytochrome P450 reductase were generous gifts from Dr.Irene Benveniste (Centre National De La Recherche Scientifique,Strasbourg, France).
Enzyme isolation. NADPH-cytochrome c reductase was isolatedfrom developing glandular trichomes of immature spearmint leavesby modification of a procedure previously used for the isolation oflimonene-6-hydroxylase (5). Buffers used were: methyl cellulose ex-traction buffer (pH 6.6) containing 25 mM Mopso,3 200 mM sorbitol,10 mM sucrose, 0.5 mM KH2PO4, 10 mM Na2S2O5, 10 mM sodiumascorbate, 1 mM EDTA, 1% PVP-40, 0.6% methyl cellulose, and 1FIG. 1. Pathways for monoterpene biosynthesis in peppermintmM DTT; wash buffer (pH 6.6) containing 25 mM KH2PO4, 1 mM(Mentha piperita), native spearmint (M. spicata), and Scotch spear-EDTA, 10% glycerol, and 1 mM DTT; sonication buffer (pH 7.4) con-mint (M. gracilis) and mutant 643 illustrating the roles of regiospe-taining 100 mM KH2PO4, 250 mM sucrose, 1 mM EDTA, 1 mM PMSF,cific cytochrome P450 monooxygenases in establishing the oxygen-1 mM DTT, 5 mM FAD, and 5 mM FMN.ation patterns of the monoterpene products of the different mint
For the isolation of leaf oil glands (6) and the extraction of theiressential oils.contents, 45–60 g of spearmint apices were harvested and placed inglass-distilled water for 1 h at 47C to maximize turgor. The tissuewas then drained and placed in a 500-ml polycarbonate Bead Beaterchamber (Biospec Products, Bartlesville, OK) with 35 g XAD-4 (poly-only tentatively identified as a P450 reductase,styrene resin) beads, 140 ml of 500-mm glass beads and Ç300 mlwhereas the latter two showed sequence homology withmethyl cellulose extraction buffer. The rotor was attached and theFMN-NADPH binding domains of other P450 reductases. contents of the chamber were stirred for 3 min at a drive-base setting
NADPH-cytochrome P450 reductases have also been iso- of 85 to 95 V, followed by filtration through a 350-mm, then 105-mm,lated from elicitor-challenged Pueraria lobata (30) and nylon mesh. The 105-mm filtrate was next filtered through a 20-mm
nylon mesh which retained the yellow oil glands (Ç60 mm diam.). Thesoybean cell suspension cultures (31); these enzymes re-isolated oil glands were washed with wash buffer and then washed offconstitute activities for the conversion of liquiritigenin tothe mesh with sonication buffer into a 50-ml steel sonication cup and2,7,4*-trihydroxyisoflavanone, and 3,9-dihydroxypterocar- made up to 50 ml with sonication buffer. After the addition of 5 g of
pan-6a-hydroxylase, respectively. The cytochrome P450 XAD-4 resin and 0.5 g PVPP, the gland suspension was sonicatedfive times for 15 s each with 45-s cooling intervals between bursts.reductase isolated from Jerusalem artichoke is capableThe sonicated suspension was filtered through a 20-mm nylon meshof reconstituting cinnamate-4-hydroxylase activity (32).to remove XAD beads, PVPP, and gland cell debris, and the filtrateMultiple forms of cytochrome P450 reductase from thiswas homogenized in a tight-fitting, chilled, 40-ml glass-on-glass (Ten-
source have also been characterized (33), suggesting the broeck) homogenizer for 10 strokes with each 10-ml aliquot. Theexistence of isoforms of this enzyme in higher plants. No pooled homogenate, made up to 100 ml with sonication buffer, was
centrifuged at 18,000g for 30 min at 47C and the supernatant wasisoforms of cytochrome P450 reductase have been de-then centrifuged at 195,000g for 90 min at 47C. The microsomaltected to date in mammalian, microbial, or insect systems.pellet was used as a source of cytochrome P450 (0)-4S-limonene-6-Failure to reliably reconstitute purified mint hydrox-hydroxylase and NADPH-cytochrome c reductase, while the 195,000g
ylases for the conversion of limonene to trans-carveol or supernatant (S195), containing appreciable amounts of the apparentlytrans-isopiperitenol, with cytochrome P450 reductases solubilized reductase, was used as the principal source for enzyme
purification. If not used immediately, these preparations were frozenthat are more readily available from other plant (mungat 0807C.bean (24)) or animal (rat liver (34)) sources, prompted
Detergent-solubilization of microsomal NADPH-cytochrome c re-us to examine the reductase from spearmint glandularductase. For a standard preparation, four microsomal pellets (rep-trichomes which does reconstitute the monoterpene resentingÇ60 g of starting tissue each) were resuspended by homog-
monooxygenase, limonene-6-hydroxylase (Fig. 1), from enization in 7 ml of 25 mM Hepes buffer, pH 7.7, containing 25 mM
the same source. This P450 reductase from mint oil NaCl, 1 mM DTT, 10 mM MgCl2, 20% glycerol, 5 mM PMSF, and 5mM leupeptin, using a tight-fitting chilled Tenbroeck homogenizer.glands is similar in gross properties to other reductasesThe resuspended microsomal membranes were diluted with an equalof plant origin.volume of the same buffer containing 2% Emulgen 911, and thesuspension was stirred for 1 h at 47C and then centrifuged at
EXPERIMENTAL PROCEDURES 195,000g for 90 min. The pellet was discarded and the supernatantwas used as a source of detergent-solubilized, microsomal reductase.Plant materials, substrates, and reagents. Spearmint (Mentha
spicata) plants were grown under controlled greenhouse conditionsas previously described (35). Young shoots of actively growing plantswere used as the enzyme source. NADPH and cytochrome c (oxidized 3 Abbreviations used: DTT, dithiothreitol; Hepes, N-2-hydroxyethyl-
piperazine-N*-2-ethanesulfonic acid; Mes, 2-(N-morpholino)ethanesul-cytochrome c, bovine heart, 98% pure) were purchased from SigmaChemical Co. Emulgen 911 was from the Kao Soap Co. (Japan), and fonic acid; Mopso, [3-(N-morpholino)-2-hydroxypropanesulfonic acid;
Tes, N-tris-(hydroxymethyl)-methyl-2-aminoethanesulfonic acid; PVP,diphenyliodonium chloride was purchased from Aldrich ChemicalCo. Other biochemicals were obtained from Sigma or Research Or- polyvinylpyrrolidone; PVPP, polyvinylpolypyrrolidone; SDS, sodium do-
decyl sulfate; S195, 195,000g supernatant; PAGE, polyacrylamide gelganics Inc., and the remaining reagents were purchased from Baker.Purified rat IgG directed against mung bean NADPH-cytochrome electrophoresis; PMSF, phenylmethylsulfonyl fluoride.
/ 6b17$$9407 03-28-96 23:24:44 arca AP: Archives

11CYTOCHROME P450 REDUCTASE FROM SPEARMINT
Enzyme assay. The assay mixture for NADPH-cytochrome c re- Trans-Blot Electrophoretic Transfer Cell. Blocking with 3% nonfatductase contained 5–100 ml enzyme extract, 100 mM Tris–HCl (pH milk was followed by incubation with primary antibody or the appro-7.7), 5 mM FAD, 5 mM FMN, 10 mM MgCl2, and 60 mM cytochrome c priate preimmune controls (1/1,000–1/10,000 dilution). Visualizationin a total volume of 1 ml in a quartz cuvette. The assay was initiated of antibody bound proteins was carried out by binding to secondaryby the addition of NADPH to a final concentration of 0.12 mM. The antibody (Bio-Rad, goat anti-rabbit (or anti-rat) IgG, 1/1,000 dilution)contents were mixed and the changes with time in the absorbance conjugated to horse radish peroxidase and chemiluminescence detec-at 550 nm (cytochrome c reduction) and at 340 nm (NADPH oxida- tion (DuPont Western Blot Chemiluminescence Reagent) on photo-tion) were monitored at 257C using a Hewlett–Packard HP8452 di- graphic film (Kodak). Rabbit polyclonal antibodies (IgG fraction) di-ode-array spectrophotometer. An extinction coefficient of 21 rected against Jerusalem artichoke NADPH-cytochrome P450 reduc-mM01cm01 (cytochrome c reduction) was used to calculate enzyme tase did not store well in our hands; stored preparations recordedactivity. Data were collected every 3 s over a period of 1 min and low molecular weight (õ77 kDa) species on immunoblots that wereinitial rates were calculated from the linear portion of the progress not observed using fresh preparations or preimmune serum.curve. NADPH-cytochrome c reductase activity is expressed in nkatal
Inhibition experiments. The purified NADPH-cytochrome c re-(1 katal corresponding to the conversion of 1 mol substrate/s). Proteinductase preparation (400 ml) was dialyzed into N2-purged, pH 7.7,concentrations were determined using the Bio-Rad microassay or thebuffer containing 50 mM Tris–HCl, 1 mM EDTA, and 20% glycerol.Micro BCA (Pierce) protein assay.To examine the inhibitory effect of diphenyliodonium chloride (39),Best-fit Km values for cytochrome c and NADPH were obtained50 ml of dialyzed enzyme preparation was mixed with 1 ml of a 6 mMusing the direct linear plot method after confirming that Eadie-NADPH solution and 1 ml of a 100 mM stock solution of diphenylio-Hofstee plots were linear. In determining the Km value for NADPH,donium chloride in 10% DMSO in an Eppendorf tube to final concen-the assays were performed in the presence of an NADPH-regenerat-trations of 0.12 and 2 mM, respectively, and the contents were prein-ing system consisting of 2 mM glucose-6-phosphate and 10 units/cubated for 10 min at 257C followed by 20-fold dilution in Tris–ml glucose-6-phosphate dehydrogenase in order to maintain fixedHCl buffer to below inhibitory concentration. NADPH-cytochrome cconcentrations of reductant.reductase activity was then measured by adding cytochrome c, fla-Enzyme purification. The pooled 195,000g supernatant fractionvins, and NADPH to the final assay concentrations previously notedfrom four preparations of oil gland extract (Ç400 ml) was concen-in a 1-ml volume and monitoring the reduction of cytochrome c in thetrated under N2 pressure by ultrafiltration (Amicon cell with YM-usual way. Control experiments performed were: (1) preincubation30 membrane) to Ç20 ml (without significant loss of activity), andwithout NADPH and diphenyliodonium chloride; (2) preincubationthe concentrate was dialyzed into buffer A (25 mM Mes–NaOH,in the presence of NADPH without inhibitor; and (3) preincubationpH 6.1, containing 10 mM MgCl2 , 20% glycerol, and 0.1 mM DTT)in the presence of inhibitor without NADPH.at 47C. The dialyzed preparation was centrifuged at 3,000g for
The inhibitory effects of NADP/, NAD/, and 2*-AMP were deter-30 min to remove precipitated protein and then mixed, using amined by varying the concentration of the nucleotide added to theLabquake Shaker (LabIndustries, Inc.), with 35 ml of Matrex Gelreaction mixture immediately prior to the addition of NADPH toRed A (Amicon) that had been preequilibrated with buffer A for 1initiate the assay. The range of concentrations examined for eachh at 47C. The slurry was then poured into a 2.6 1 13.5-cm columnnucleotide were: 0.1–0.5 mM for NADP/; 5–20 mM for NAD/; andand washed with 200 ml buffer A. NADPH-cytochrome c reductase0.25–7.5 mM for 2 *-AMP.activity was eluted with 150 ml buffer B (100 mM Tris–HCl, pH
7.7, containing 1 mM EDTA, 20% glycerol, and 0.1 mM DTT) while Immunoinhibition of NADPH-cytochrome c (P450) reductase. Ly-40-ml fractions were collected. The active fraction was concen- ophilized polyclonal rabbit antibodies (serum and purified IgG) andtrated without significant loss to 3–5 ml by centrifugation at rat monoclonal antibodies directed against Jerusalem artichoke2,500g in an Amicon Centriprep-30 concentrator and the concen-
NADPH-cytochrome P450 reductase (40, 41) were resuspended in antrate was then applied to a 0.9 1 6.5-cm column of 2 *,5*-ADPequal amount of phosphate-buffered saline (pH 7.2). The rat poly-agarose (Sigma; 2-ml bed volume) that had been preequilibratedclonal antibody preparation directed against mung bean NADPH-with buffer C (25 mM Tris–HCl, pH 7.7, containing 1 mM EDTA,cytochrome P450 reductase (24) was available as purified IgG in20% glycerol, and 0.1 mM DTT). The column was washed with 6solution. For immunoinhibition experiments, the affinity chromatog-ml of buffer B followed by 9 ml of buffer C, and the pure NADPH-raphy purified, glandular NADPH-cytochrome c reductase was di-cytochrome c reductase was then eluted with 6 ml of buffer Cluted with resuspended antibody preparation to a 1:1 protein:anti-containing 1 mM 2 *-AMP; 3-ml fractions were collected. Activebody ratio and incubated at 257C for 10 min prior to assay. Controlsfractions were used immediately or were stored frozen at 0807Cwere performed with the appropriate rat or rabbit preimmune serum.under N2 without loss of activity for several months.The percentage of remaining reductase activity was calculated with
SDS–PAGE. Denaturing polyacrylamide gel electrophoresis was respect to parallel control incubations in which buffered saline wascarried out using the discontinuous system of Laemmli (36) with used in place of the antibody preparation.0.1% SDS (10% resolving gel and 5% stacking gel, 1-mm thickness).
Reconstitution of (0)-4S-limonene-6-hydroxylase activity. Puri-Electrophoresis was performed in a Bio-Rad Mini-PROTEAN II verti-fied NADPH-cytochrome c reductase (1.8–2.5 nkatal) and detergent-cal slab gel apparatus at 20 mA (150 V) for 1 h. Protein samples
were diluted with 31 loading buffer (186 mM Tris–HCl, pH 6.8, free cytochrome P450 limonene-6-hydroxylase (200 pmol) were com-containing 30% glycerol, 6% SDS, 90 mM DTT, and 0.006% bromo- bined in a minimum volume of buffer (100–200 ml) on ice. Flavins,phenol blue) and denatured by steam heating for 5 min. Aliquots MgCl2, and dilauroyl phosphatidyl choline were next added, again(10–20 ml) were loaded into each lane, and molecular weight markers in a small volume, and the temperature was raised to 327C. The(Diversified Biotech, 12.4–95.5 kDa) were run in parallel with the mixture was then diluted to 0.5 ml with 25 mM Tris–HCl buffer, pHdenatured protein samples. Protein bands were visualized by the 7.7, containing 1 mM EDTA, 1 mM DTT, and 20% glycerol such thatthiosulfate-silver staining method (37). the final component concentrations of the reconstituted system were
5 mM FAD, 5 mM FMN, 30.4 mM dilauroyl phosphatidyl choline, 10Electroblotting and immunodetection. Electrophoretic transfer ofmM MgCl2, 0.4 mM cytochrome P450, and 3.6–5.0 nkatals/ml purifiedproteins, separated by SDS–PAGE, onto nitrocellulose membranesNADPH-cytochrome c reductase. Saturating concentrations ofwas performed according to a published method (38). NitrocelluloseNADPH (2 mM) and (0)-4S-limonene (200 mM) were finally added,membranes and SDS–PAGE gels were soaked in pH 8.3 transblotterand the assay for hydroxylation of the olefinic substrate to trans-buffer (20 mM Tris, 150 mM glycine, 20% methanol, 0.04% SDS) and
electroblotting then carried out at 100 V (250 mA) in a Bio-Rad Mini carveol was carried out as described previously (3).
/ 6b17$$9407 03-28-96 23:24:44 arca AP: Archives
12 PONNAMPERUMA AND CROTEAU
TABLE I
Purification of NADPH-Cytochrome c Reductase from Spearmint Oil Glands
Total protein Total activity Specific activity Purification factor YieldPurification step (mg) (nkatal) (nkatal/mg) (fold) (%)
195,000g supernatant 32.0 120.3 3.7 1.0 100Matrex Gel Red A 3.6 78.0 21.7 5.9 652*,5*-ADP agarose 0.05 36.0 720 195 30
Note. The numbers represent the means of four preparations, each consisting of roughly 60 g of leaf tissue.
RESULTS least in part, to the loss of flavins, and to residual, moretightly bound activity (at least 10%) that eluted at 300Isolation and Purification of Spearmint Oil GlandmM Tris–HCl; the latter was not used for further puri-NADPH-Cytochrome c Reductasefication.
NADPH-cytochrome c reductase activity was found Affinity chromatography on NADP-agarose was in-to be associated with both the microsomal (40% of total) vestigated as a prospective fractionation step, but noand 195,000g supernatant (60% of total) fractions ob- binding of the reductase could be obtained under a verytained from oil gland extracts. To enable enzyme puri- wide range of conditions. However, the reductase didfication with sufficient amounts of protein, the S195 frac- bind to 2 *,5*-ADP-agarose and could be selectivelytions from multiple oil gland preparations were com- eluted from this matrix with 1 mM of 2 *-AMP, whichbined and concentrated by ultrafiltration to a small provided an effective step in terms of reproducibility,volume atÇ1 mg protein/ml in preparation for fraction- recovery and purification. A considerable number ofation. The resulting concentrate could be stored frozen contaminating protein species were removed in the var-at 0207C for several months without loss of activity. ious wash fractions, whereas the material eluted withThe microsomal NADPH-cytochrome c reductase was 1 mM of 2 *-AMP showed only a major band at 82 kDaquantitatively solubilized using Emulgen 911 or Triton and a minor band at 77 kDa on SDS–PAGE (Fig. 2).N-101. The detergent-solubilized reductase behaved The purified material could be stored frozen at 0807Csimilarly to the S195 reductase that was solubilized dur-
for several months without loss of activity, as coulding gland extraction, and both species ostensibly repre-the crude material, microsomal pellets, and detergentsent the same protein (see below).solubilized reductase.The scheme for purification of the NADPH-cyto-
chrome c reductase from the S195 concentrate is pre-sented in Table I. Several fractionation steps, in addi-tion to red dye–ligand interaction chromatography,were explored initially, including anion-exchange(DEAE-Sepharose), gel-filtration (on Sephacryl S-200,the reductase sized at about 80 kDa), and hydrophobicinteraction (methyl-agarose) chromatography. Al-though the purification factors and recoveries achievedwere comparable to those obtained by Matrex Gel RedA chromatography, only the combination of dye-ligandchromatography and chromatography on 2 *,5*-ADP-agarose yielded a purified reductase that was free of acontaminating 30- to 35-kDa protein species. A rela-tively low pH (6.1) was required for binding of the re-ductase to the Matrex Gel Red A dye-ligand column;
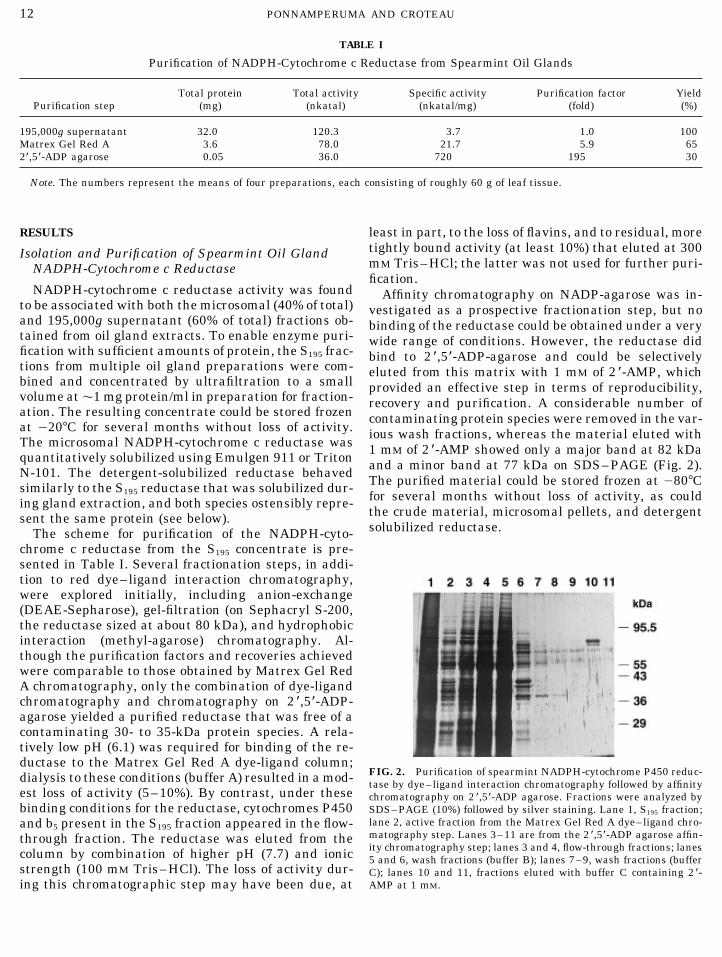
FIG. 2. Purification of spearmint NADPH-cytochrome P450 reduc-dialysis to these conditions (buffer A) resulted in a mod- tase by dye–ligand interaction chromatography followed by affinityest loss of activity (5–10%). By contrast, under these chromatography on 2*,5*-ADP agarose. Fractions were analyzed bybinding conditions for the reductase, cytochromes P450 SDS–PAGE (10%) followed by silver staining. Lane 1, S195 fraction;
lane 2, active fraction from the Matrex Gel Red A dye–ligand chro-and b5 present in the S195 fraction appeared in the flow-matography step. Lanes 3–11 are from the 2 *,5*-ADP agarose affin-through fraction. The reductase was eluted from theity chromatography step; lanes 3 and 4, flow-through fractions; lanescolumn by combination of higher pH (7.7) and ionic 5 and 6, wash fractions (buffer B); lanes 7–9, wash fractions (buffer
strength (100 mM Tris–HCl). The loss of activity dur- C); lanes 10 and 11, fractions eluted with buffer C containing 2*-AMP at 1 mM.ing this chromatographic step may have been due, at
/ 6b17$$9407 03-28-96 23:24:44 arca AP: Archives
13CYTOCHROME P450 REDUCTASE FROM SPEARMINT
TABLE II 8% of the reaction rate supported by NADPH as thereductant at saturation.Inhibition of NADPH-Reduced NADPH-Cytochrome c
Reductase by the Diphenyliodonium Ion The purified enzyme was reversibly inhibited byNADP/ (I50 Ç0.15 mM), NAD/ (I50 Ç25 mM) and 2 *-
Cytochrome c AMP (I50 Ç1.25 mM). The relatively high I50 value forreductase activity 2 *-AMP allowed the affinity-purified enzyme to be as-
Preincubation conditions (pkatal) sayed directly without prior dialysis or gel filtration toremove the AMP. The diphenyliodonium ion at 2 mMReductase alone 320completely and irreversibly inhibited the NADPH-re-Reductase / 0.12 mM NADPH / 2 mM
diphenyliodonium ion 6 duced form of the enzyme (Table II). Attempts to re-Reductase / 0.12 mM NADPH 300 store activity by removal of excess inhibitor by gel-fil-Reductase / 2 mM diphenyliodonium ion 240 tration or dialysis were unsuccessful.
Note. The affinity purified enzyme was used for these studies and,Immunodetection and Immunoinhibitionfollowing 20-fold dilution of the preincubation mixture, the standard
NADPH-cytochrome c reductase assay was employed.Electroblotted SDS–PAGE gels of affinity-purified
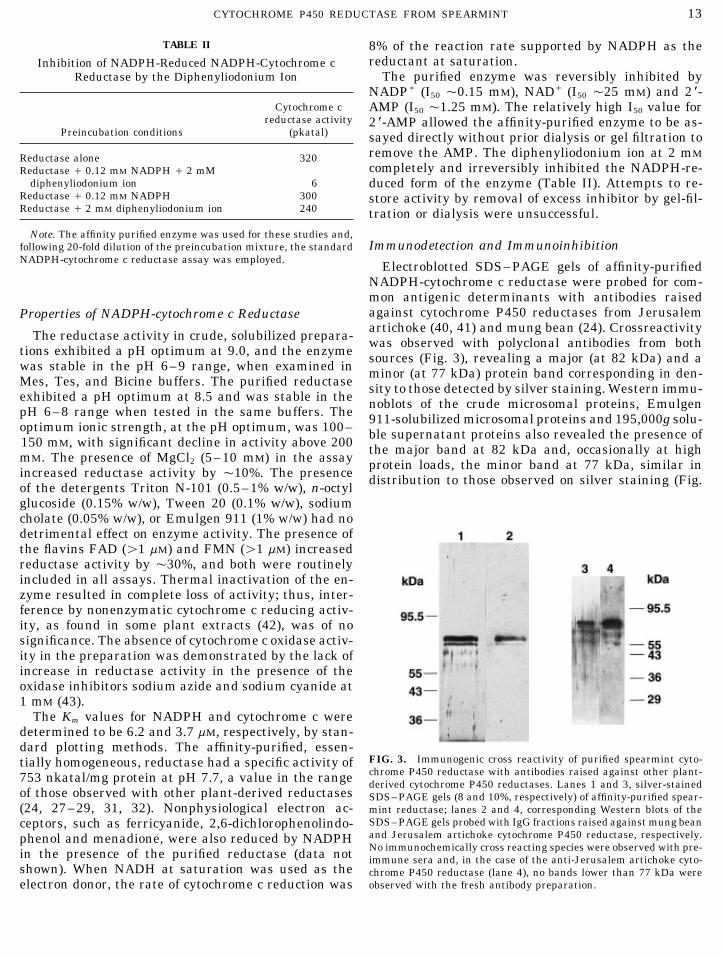
NADPH-cytochrome c reductase were probed for com-mon antigenic determinants with antibodies raisedagainst cytochrome P450 reductases from JerusalemProperties of NADPH-cytochrome c Reductaseartichoke (40, 41) and mung bean (24). Crossreactivity
The reductase activity in crude, solubilized prepara- was observed with polyclonal antibodies from bothtions exhibited a pH optimum at 9.0, and the enzyme sources (Fig. 3), revealing a major (at 82 kDa) and awas stable in the pH 6–9 range, when examined in minor (at 77 kDa) protein band corresponding in den-Mes, Tes, and Bicine buffers. The purified reductase sity to those detected by silver staining. Western immu-exhibited a pH optimum at 8.5 and was stable in the noblots of the crude microsomal proteins, EmulgenpH 6–8 range when tested in the same buffers. The 911-solubilized microsomal proteins and 195,000g solu-optimum ionic strength, at the pH optimum, was 100– ble supernatant proteins also revealed the presence of150 mM, with significant decline in activity above 200 the major band at 82 kDa and, occasionally at highmM. The presence of MgCl2 (5–10 mM) in the assay protein loads, the minor band at 77 kDa, similar inincreased reductase activity by Ç10%. The presence distribution to those observed on silver staining (Fig.of the detergents Triton N-101 (0.5–1% w/w), n-octylglucoside (0.15% w/w), Tween 20 (0.1% w/w), sodiumcholate (0.05% w/w), or Emulgen 911 (1% w/w) had nodetrimental effect on enzyme activity. The presence ofthe flavins FAD (ú1 mM) and FMN (ú1 mM) increasedreductase activity by Ç30%, and both were routinelyincluded in all assays. Thermal inactivation of the en-zyme resulted in complete loss of activity; thus, inter-ference by nonenzymatic cytochrome c reducing activ-ity, as found in some plant extracts (42), was of nosignificance. The absence of cytochrome c oxidase activ-ity in the preparation was demonstrated by the lack ofincrease in reductase activity in the presence of theoxidase inhibitors sodium azide and sodium cyanide at1 mM (43).
The Km values for NADPH and cytochrome c weredetermined to be 6.2 and 3.7 mM, respectively, by stan-dard plotting methods. The affinity-purified, essen-
FIG. 3. Immunogenic cross reactivity of purified spearmint cyto-tially homogeneous, reductase had a specific activity ofchrome P450 reductase with antibodies raised against other plant-753 nkatal/mg protein at pH 7.7, a value in the range derived cytochrome P450 reductases. Lanes 1 and 3, silver-stained
of those observed with other plant-derived reductases SDS–PAGE gels (8 and 10%, respectively) of affinity-purified spear-(24, 27–29, 31, 32). Nonphysiological electron ac- mint reductase; lanes 2 and 4, corresponding Western blots of the
SDS–PAGE gels probed with IgG fractions raised against mung beanceptors, such as ferricyanide, 2,6-dichlorophenolindo-and Jerusalem artichoke cytochrome P450 reductase, respectively.phenol and menadione, were also reduced by NADPHNo immunochemically cross reacting species were observed with pre-in the presence of the purified reductase (data not immune sera and, in the case of the anti-Jerusalem artichoke cyto-
shown). When NADH at saturation was used as the chrome P450 reductase (lane 4), no bands lower than 77 kDa wereobserved with the fresh antibody preparation.electron donor, the rate of cytochrome c reduction was
/ 6b17$$9407 03-28-96 23:24:44 arca AP: Archives

14 PONNAMPERUMA AND CROTEAU
Reconstitution
(0)-4S-Limonene-6-hydroxylase activity was recon-stituted with affinity-purified NADPH-cytochrome c re-ductase and cytochrome P450 purified from microsomalpreparations from spearmint oil gland cells, in the pres-ence of dilauroyl phosphatidyl choline. In several ex-periments, trans-carveol formation corresponding toturnover numbers in the range 0.1–1.0/s were observedin the presence of saturating concentrations of NADPHand (0)-4S-limonene.
DISCUSSION
The purification and characterization of NADPH-cy-FIG. 4. Immunogenic cross-reactivity of crude spearmint cyto-chrome P450 reductase with anti-mung bean cytochrome P450 reduc- tochrome c reductase from the glandular trichomes oftase IgG. Lanes 1, 3, and 5 are silver-stained SDS–PAGE gels (10%) spearmint, the site of monoterpene biosynthesis (2, 6),of crude reductase fractions: lane 1, 195,000g supernatant fraction represents part of a larger effort to define the micro-(4 mg); lane 3, resuspended microsomes (3 mg); lane 5, Emulgen 911-
somal cytochrome P450-dependent monooxygenasesolubilized microsomal fraction (1.5 mg). Lanes 2, 4, and 6 are thecorresponding Western blots of the SDS–PAGE gels probed with the systems of commercial mint species responsible for theanti-mung bean cytochrome P450 reductase IgG (the faint band at oxidative transformation of (0)-4S-limonene to severalÇ65 kDa seen in lanes 4 and 6 is most likely a proteolytic product). families of essential oil monoterpenes (44).Results obtained with the anti-Jerusalem artichoke cytochrome P450
The 200-fold purification of the reductase to near ho-reductase IgG were similar, and no immunochemically cross reactingmogeneity (Table I) contrasts to the several thousand-proteins were observed with the preimmune serum controls.fold purifications required for other plant-derived cyto-chrome P450 reductases isolated from other tissuesand organs, such as tubers (27), flowers (29), and seed-
4). No immunochemically crossreacting proteins were lings (24), indicating that the oil glands represent aobserved on Western blots probed with preimmune highly enriched source of this enzyme. The propertiessera. of the purified reductase resemble those of other plant-
The anti-mung bean and anti-Jerusalem artichoke cy- derived NADPH cytochrome c reductases with respecttochrome P450 reductase IgGs were found to inhibit to molecular weight (Ç80,000 by SDS–PAGE and gel-strongly the solubilized spearmint reductase activity filtration chromatography), kinetic constants for cyto-when employed in a 1:1 antibody:enzyme ratio, whereas chrome c and NADPH, inhibition by NADP/, NAD/,preimmune sera were without effect (Table III). Of the and 2 *-AMP, ability to reduce nonphysiological elec-two families of anti-Jerusalem artichoke P450 reductase tron acceptors, and strong preference for NADPH overmonoclonal antibodies, C5A5 and AD(3)3 (41), only the for- NADH as the electron donor.
Few specific, mechanism-based inhibitors of cyto-mer partially inhibited cytochrome c reductase activity.
TABLE III
Immunoinhibition of Spearmint NADPH-Cytochrome c Reductase by Antibodies Raisedagainst NADPH-Cytochrome P450 Reductase from Jerusalem Artichoke and Mung Bean
NADPH-cytochrome creductase activity
Antibody (pkatal) % of control
Control (no antibody) 359 100Rabbit preimmune serum 328 91Rat preimmune serum 340 95Rat anti-mung bean reductase (IgG) 14.3 4Rabbit anti-Jerusalem artichoke reductase
(1) immune serum 11.9 3.3(2) IgG 9.9 3.0
Rat anti-Jerusalem artichoke monoclonal C5A5 211 59Rat anti-Jerusalem artichoke monoclonal AD(3)3 348 97
/ 6b17$$9407 03-28-96 23:24:44 arca AP: Archives

15CYTOCHROME P450 REDUCTASE FROM SPEARMINT
chrome P450 reductases have been reported. Recently, chrome c reductase purified from spearmint oilglands is an NADPH-cytochrome P450 reductasethe diphenyliodonium ion was shown to be an irrevers-
ible inhibitor of liver microsomal P450 reductase, and and, thus, an essential component of the glandularcytochrome P450-dependent limonene hydroxylaseinactivation was demonstrated to result, in part, from
covalent modification of the reduced FMN prosthetic system.group (39). Complete, irreversible inhibition of thespearmint gland NADPH-reduced cytochrome P450 re- ACKNOWLEDGMENTSductase by the diphenyliodonium cation thus supports We thank Ronald Estabrook and Irene Benveniste for generouslythe flavoprotein nature of this enzyme. Immunogenic providing antibodies, Greg Wichelns for raising the plants, and Joyce
Tamura-Brown for typing the manuscript.cross-reactivity on immunoblotting with polyclonalIgGs from mung bean and Jerusalem artichoke indi-cate the presence of epitopes common to these P450 REFERENCESreductases, a finding confirmed by immunoinhibition 1. Bolwell, G. P., Bozak, K., and Zimmerlin, A. (1994) Phytochemis-of the spearmint reductase activity by antibodies from try 37, 1491–1506.both sources. 2. Gershenzon, J., Maffei, M., and Croteau, R. (1989) Plant Physiol.
The presence of NADPH-cytochrome c reductase in 89, 1351–1357.the 195,000g supernatant of the glandular trichome 3. Karp, F., Mihaliak, C. A., Harris, J. L., and Croteau, R. (1990)
Arch. Biochem. Biophys. 276, 219–226.extract suggested the possibility of proteolytic solubili-4. Croteau, R., Karp, F., Wagschal, K. C., Satterwhite, D. M., Hyatt,zation of the microsomal enzyme. However, immunode-
D. C., and Skotland, C. B. (1991) Plant Physiol. 96, 744–752.tection of two proteins of sizes 82 kDa (major) and 775. Croteau, R., and Gershenzon, J. (1994) in Genetic EngineeringkDa (minor) on Western blots of Emulgen 911-solubi-
of Plant Secondary Metabolism: Recent Advances in Phytochem-lized microsomes, which were identical to those ob- istry, (Ellis, B. E., Kuroki, G., and Stafford, H. A., Eds.), Vol. 28,served with the NADPH-cytochrome c reductase puri- pp. 193–229, Plenum Press, New York.fied from the 195,000g supernatant, indicated that pro- 6. Gershenzon, J., McCaskill, D., Rajaonarivony, J. I. M., Mihaliak,
C., Karp, F., and Croteau, R. (1992) Anal. Biochem. 200, 130–teolysis was not likely responsible for the solubilization138.of the reductase or the presence of the two copurifying
7. Bergh, A. F., and Strobel, H. W. (1992) J. Neurochem. 59, 575–reductase species. Rather, the detection of two proteins581.on the immunoblots, and the possibility of a minor,
8. Yasukochi, Y., and Masters, B. S. S. (1976) J. Biol. Chem. 251,tighter binding species on dye-ligand chromatography,5337–5344.suggest the occurrence of reductase isoforms from this
9. Dignam, J. D., and Strobel, H. W. (1975) Biochem. Biophys. Res.source. Further evidence against proteolytic solubiliza- Commun. 63, 845–852.tion was provided by the ability of the purified reduc- 10. Vermilion, J. L., and Coon, M. J. (1974) Biochem. Biophys. Res.tase to reconstitute limonene-6-hydroxylase activity Commun. 60, 1315–1322.with cytochrome P450 purified from the same source. 11. Iscan, M. Y., and Arinc, E. (1986) J. Biochem. 18, 731–741.In studies with mammalian P450 reductases, the pro- 12. Fukuda, H., Fujii, T., Sukita, E., Tazaki, M., Nagahama, S., and
Ogawa, T. (1994) Biochem. Biophys. Res. Commun. 201, 516–teolytically solubilized form of the enzyme, although522.functional in the reduction of cytochrome c, was unable
13. Black, S. D., Linger, M. H., Freck, L. C., Kazemi, S., and Gal-to reduce cytochrome P450 and, therefore, was incapa-braith, J. A. (1994) Arch. Biochem. Biophys. 310, 126–133.ble of reconstituting P450-dependent hydroxylase ac-
14. Peterson, J. A., and Boddupalli, S. S. (1992) Arch. Biochem. Bio-tivity (45, 46).phys. 294, 654–661.The appearance of reductase activity in the high
15. Mayer, R. T., and Durrant, J. L. (1979) J. Biol. Chem. 254, 756–speed supernatant fraction of the gland extract may 761.result from protein solubilization by endogenous 16. Vincent, D. R., and Terriere, L. C. (1985) Insect Biochem. 15,small molecules from these oil storage structures. 299–307.Gas chromatographic analyses of ether extracts of 17. Porter, T. D., and Kasper, C. B. (1985) Proc. Natl. Acad. Sci.
USA 82, 973–977.resuspended microsomes from the oil gland cells re-vealed the presence of lipophilic terpenoids, includ- 18. Katagiri, M., Murakami, H., Yabusaki, Y., Sugiyama, T., Oka-
moto, M., Yamano, T., and Ohkawa, H. (1986) J. Biochem. 100,ing carvone and germacrene D, despite treatment of945–954.the gland homogenate with polystyrene beads and
19. Haniu, M., Iyanagi, T., Miller, P., Lee, T. D., and Shively, J. E.vinylpyrrolidone polymers that was designed to re-(1986) Biochemistry 25, 7906–7911.move these materials. The terpenoids, together with
20. Koener, J. F., Carino, F. A., and Feyereisen, R. (1993) Insectflavonoids which are also known to be present in high Biochem. Molec. Biol. 23, 439–447.concentration in mint glands (47), could readily par- 21. Yabusaki, Y., Murakami, H., and Ohkawa, H. (1988) J. Biochem.tition into the microsomal membranes during extrac- 103, 1004–1010.tion and promote protein solubilization. In summary, 22. Sutter, T. R., Sangard, D., and Loper, J. C. (1990) J. Biol. Chem.
265, 16428–16436.there is compelling evidence that the NADPH-cyto-
/ 6b17$$9407 03-28-96 23:24:44 arca AP: Archives

16 PONNAMPERUMA AND CROTEAU
23. Ruettinger, R. T., Wen, L. P., and Fulco, A. J. (1989) J. Biol. 34. Dignam, J. D., and Strobel, H. W. (1977) Biochemistry 16, 1116–1123.Chem. 264, 10987–10995.
35. Alonso, W. R., Rajaonarivony, J. I. M., Gershenzon, J., and Cro-24. Shet, M. S., Sathasivan, K., Arlotto, M. A., Mehdy, M. C., andteau, R. (1992) J. Biol. Chem. 267, 7582–7587.Estabrook, R. W. (1993) Proc. Natl. Acad. Sci. USA 90, 2890–
36. Laemmli, U. K. (1990) Nature 227, 680–685.2894.37. Blum, H., Beier, H., and Gross, H. J. (1987) Electrophoresis 8,25. Meijer, A. H., Cardoso, M. I. L., Voskuilen, J. T., de Waal, A.,
93–99.Verpoorte, R., and Hoge, J. H. C. (1993) Plant J. 4(1), 47–60.38. Towbin, H., Staehelin, T., and Gordon, J. (1979) Proc. Natl. Acad.
26. Madyastha, K. M., and Coscia, C. J. (1979) J. Biol. Chem. 254, Sci. USA 76, 4350–4354.2419–2427.
39. Tew, D. G. (1993) Biochemistry 32, 10209–10215.27. Fujita, M., and Asahi, T. (1985) Plant Cell Physiol. 26(3), 397– 40. Benveniste, I., Lesot, A., Hasenfratz, M.-P., and Durst, F. (1989)
405. Biochem. J. 259, 847–853.28. Meijer, A. H., Pennings, E. J. M., de Waal, A., and Verpoorte, 41. Lesot, A., Benveniste, I., Hasenfratz, M.-P., and Durst, F. (1992)
R. (1990) in Progress in Plant Cellular and Molecular Biology Plant Physiol. 100, 1406–1410.(Nijkamp, H. J., Van der Plas, L. H. W., and Van Aertrijk, J., 42. Pohl, U., and Wiermann, R. (1981) Anal. Biochem. 116, 425–Eds.), pp. 769–774, Kluwer Academic Publishers, Dordrecht. 432.
43. Tolbert, N. E. (1974) in Methods in Enzymology (Fleischer, S.,29. Menting, J. G. T., Cornish, E., and Scopes, R. K. (1994) Plantand Packer, L., Eds.), Vol. 31, pp. 734–746, Academic Press,Physiol. 106, 643–650.New York.30. Hakamatsuka, T., Hashim, M. F., Ebizuka, Y., and Sankawa,
44. Lupien, S., Karp, F., Ponnamperuma, K., Wildung, M., and Cro-U. (1991) Tetrahedron 47, 5969–5978.teau, R. (1995) Drug Metab. Drug Interact. 12, 245–260.
31. Kochs, G., and Grisebach, H. (1989) Arch. Biochem. Biophys. 45. Gum, J. R., and Strobel, H. W. (1979) J. Biol. Chem. 254, 4177–273, 543–553. 4185.32. Benveniste, I., Gabriac, B., and Durst, F. (1986) Biochem. J. 235, 46. Black, S. D., and Coon, M. J. (1982) J. Biol. Chem. 257, 5929–
365–373. 5937.47. Voirin, B., Bayet, C., and Colson, M. (1993) Phytochemistry 34,33. Benveniste, I., Lesot, A., Hasenfratz, M. P., Kochs, G., and Durst,
F. (1991) Biochem. Biophys. Res. Commun. 177, 105–112. 85–87.
/ 6b17$$9407 03-28-96 23:24:44 arca AP: Archives




















![Fertilizer Guide: Peppermint and Spearmint - [email protected] Home](https://img.pdfslide.net/doc/110x75/613d71dd736caf36b75d68f6/fertilizer-guide-peppermint-and-spearmint-emailprotected-home.jpg)








