Embed Size (px)
Citation preview

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 268, No. 2. February 1, pp. 617-629,1989
Reduction of 7-Alkoxyresorufins by NADPH-Cytochrome P450 Reductase and Its Differential Effects on Their 0-Dealkylation by
Rat Liver Microsomal Cytochrome P450’
DAVID R. DUTTON AND ANDREW PARKINSON’
Departnwnt q~Phnrmnc:olo~~y, Toxicology, and Thfrapevctics, University of Kam-as Medical Center, Kc~mas City. h’awas 6610.3
Received August 12,1988, and in revised form September 30,1988
Antibody-inhibition experiments established that the induction of cytochrome P45Oc is largely responsible for the marked increase in liver microsomal7-ethoxyresorufin O- dealkylation in rats treated with 3-methylcholanthrene, whereas the induction of cyto- chrome P450b and/or P450e is largely responsible for the marked increase in 7-pentoxy- and 7-benzyloxyresorufin 0-dealkylation in rats treated with phenobarbital. When re- constituted with NADPH-cytochrome P450 reductase and lipid, purified cytochrome P45Oc catalyzed the 0-dealkylation of 7-ethoxyresorufin at a rate of -30 nmol/nmol P450/min, which far exceeded the rate catalyzed by either purified cytochromes P450b and P450e or microsomal cytochrome P45Oc. In contrast, purified cytochromes P450b and P450e were poor catalysts of the 0-dealkylation of 7-pentoxy- and 7-benzyloxyresorufin. However, purified cytochrome P450b is an excellent catalyst of several other reactions, such as the N-demethylation of benzphetamine, the hydroxylation of testosterone, and the 0-dealkylation of 7-ethoxycoumarin. The low rate of 7-pentoxyresorufin O-dealkyl- ation catalyzed by purified cytochrome P450b did not reflect a requirement for cyto- chrome bs, and could not be ascribed to an artifact of the method used to measure the formation of resorufin. The catalytic activity of purified cytochrome P450b toward 7- pentoxyresorufin was consistently low over a range of substrate and lipid concentra- tions, and was not stimulated by sodium deoxycholate (which stimulates the N-demeth- vlation of benzphetamine by purified cytochrome P450b). Evidence is presented which indicates that cytochrome P45Oc catalvzes the 0-dealkvlation of both the oxidized and reduced forms of 7-ethoxyresorufin, with perhaps a slight preference for the reduced form. In contrast, cytochrome P450b preferentially catalyzes the 0-dealkylation of the oxidized form of 7-pentoxyresorufin. Conditions that favored formation of the reduced form of 7-ethoxyresorufin tended to stimulate its 0-dealkylation by purified cytochrome P45Oc, whereas conditions that favored formation of the reduced form of 7-pentoxyre- sorufin decreased its rate of 0-dealkylation by purified cytochrome P450b. Such condi- tions included a molar excess of NADPH-cytochrome P450 reductase over cytochrome P450, the presence of superoxide dismutase, and the presence of DT-diaphorase (liver cytosol). Both NADPH-cytochrome P450 reductase and DT-diaphorase increased the rate of formation of the reduced form of the 7-alkosyresorufins, whereas superoxide dismutase decreased its rate of disappearance (because superoxide anion, which is formed during the redox cycling reactions described in the preceding paper, causes a
’ This research was supported by Grants ES 03765 and GM 37044 from the National Institutes of Health. from the National Institutes of Health. A prelimi- D.R.D. is supported by Training Grant ES 07079 from nary account of this work was presented in abstract the National Institutes of Health, and by Stauffer form (D. R. Dutton and A. Parkinson (1987) Fed Proc. Chemical Co., Farmington, CT. A.P. is a recipient 46,1957). of a Research Career Development Award (ES 00166) ’ To whom correspondence should be addressed.
617 0003-9861189 $3.00 Copyright Q 1989 by Academic Press. Inc. All rights of reproduction in any form reserved.

618 DUTTON AND PARKINSON
rapid reoxidation of the reduced form of the 7-alkoxyresorufinsj. Although a molar ex- cess of NADPH-cytochrome P450 reductase is required for purified cytochrome P450b to exert its full catalytic activity toward benzphetamine, testosterone, and 7-ethoxycou- marin, the 0-dealkylation of 7-pentoxyresorufin was greatest when the ratio of NADPH-cytochrome P450 reductase-to-cytochrome P450b was less than one. Inasmuch as a molar excess of NADPH-cytochrome P450 reductase compromised the catalytic activity of cytochrome P450b toward 7-pentoxy- and 7-benzyloxyresorufin, it is signifi- cant that the molar ratio of cytochrome P450 to NADPH-cytochrome P450 reductase in liver microsomes is greater than 1O:l. We speculate that if rat liver microsomes did not contain a molar excess of cytochrome P450 over NADPH-cytochrome P450 reductase, the 0-dealkylation of 7-pentoxy- and 7-benzyloxyresorufin would be of little value in monitoring the induction of cytochromes P450b and P450e by phenobarbital. ,e 1989 ~~~~ demie Press. Inc.
Rat liver microsomes contain many cy- tochrome P450 isozymes that catalyze the biotransformation of numerous xenobiot- its (such as drugs, pesticides, and chemical carcinogens) and several endogenous sub- strates (such as steroid hormones). One of the major problems in studying the micro- somal cytochrome P450 system has been a lack of specific enzymatic reactions to dis- tinguish and measure individual isozymes. This problem is related to the broad and often overlapping substrate specificity of many of the microsomal cytochrome P450 isozymes (l-3). However, some reactions are catalyzed predominantly by a single isozyme (or group of closely related iso- zymes) of rat liver microsomal cytochrome P450. Examples of such reactions include the regio- and stereoselective hydroxyla- tion of polycyclic aromatic hydrocarbons, warfarin, and steroid hormones (namely testosterone, androstenedione, and pro- gesterone) and the 0-dealkylation of 7-al- koxyresorufins (4-15).
It has been shown by immunochemical analysis that treatment of rats with phe- nobarbital causes a marked induction (-50 fold) of cytochromes P450b3 and
3 Recently, a nomenclature system for cytochrome P450 isozymes was proposed based on the genes of the cytochrome P450 superfamily whose cDNA and/or amino acid sequences have been determined (17). We have included the new committee on Standardized Nomenclature conventions in boldface in this foot- note. The nomenclature system of Ryan et nl. (1, 2, 18) is used throughout the manuscript. Cytochrome P450b (P450IIBl) is also known as P450 PB-B (3,lZj. P450 PB-4 (19), or P450 I-C (20); cytochrome P45Oc
P450e (1, 16), whereas treatment with 3- methylcholanthrene causes an equally marked induction of cytochromes P45Oc and P450d (1, 16). The induction of cyto- chrome P450b by phenobarbital is asso- ciated with a marked increase in the 16/3-hydroxylation of testosterone and the 0-dealkylation of 7-pent.oxy- and 7- benzyloxyresorufin catalyzed by rat liver microsomes (4-6, 13, 14). The induction of cytochromes P45Oc by 3-methylcholan- threne is associated with a marked in- crease in the hydroxylation of benzo[a]- pyrene and the 0-dealkylation of 7-ethoxy- resorufin (9, 14, 16). As expected, purified cytochrome P45Oc, when reconstituted with NADPH-cytochrome P450 reductase and lipid, catalyzes the 0-dealkylation of 7-ethoxyresorufin at a rate which exceeds that catalyzed by liver microsomes from 3- methylcholanthrene-induced rats (12, 15). In contrast, purified cytochrome P450b is an unexpectedly poor catalyst of the O-de- alkylation of 7-pentoxy- and 7-benzyloxy- resorufin (13, 21). Purified cytochrome P450b is, however, an effective catalyst of many other biotransformation reactions, including the lG@hydroxylation of testos- terone, the N-demethylation of benzphe- tamine, and the 0-dealkylation of ‘I-eth- oxycoumarin (4,12,22).
We have undertaken studies to deter- mine why purified cytochrome P450b, when reconstituted with NADPH-cyto-
(P450IAl) is also known as P450 PNF-B (3. 12); and cytochrome P450e (P450IIB2) as P450 PB-D (3, 12) and P450 PB-5 (19).

7-ALKOXYRESORUFIN 0-DEALKYLATION BY PURIFIED CYTOCHROME P450 619
chrome P450 reductase and lipid, is an un- expectedly poor catalyst of the O-dealkyl- ation of 7-pentoxy- and 7-benzyloxyreso- rufin. In the preceding paper (23), we showed that NADPH-cytochrome P450 re- ductase catalyzes the reduction of resoru- fin and the 7-alkoxyresorufins which, in the presence of 02, initiates a redox cycling re- action. The studies described in this paper show that reduction of the 7-alkoxyresoru- fins by NADPH-cytochrome P450 reduc- tase impedes their 0-dealkglation by cyto- chrome P&Ob, but actually enhances their 0-dealkylation by cytochrome P45Oc.
EXPERIMENTAL PROCEDURES
Chenticuls. 3-Methylcholanthrene and cumene hy- droperoxide were purchased from Sigma Chemical Co. (St. Louis, MO); potassium superoxide was from Pfaltz and Bauer Inc. (Waterbury, CT), and sodium phenobarbital was obtained from the Department of Pharmacy, Kansas University Medical Center. The sources of all other reagents are given in the preced- ing paper (23).
Animal treatnmlt a td prepumtinn (d liver wz i- crosomes a& cytosot. Male Sprague-Dawley rats (Charles River Labs, Wilmington, MA) were killed at 9 weeks of age after 4 consecutive daily injections (ip) of corn oil (10 ml/kg), phenobarbital (80 mg/kg), or 3-methylcholanthrene (27 mg/kg). Alternatively, 56- day-old-male Sprague-Dawley (Charles River Labs) or 28-day-old-male Long-Evans rats (Blue Spruce Farms, Altamont, NY) were killed 4 days after a sin- gle injection of Aroclor 1254 (500 mg/kg) or corn oil. Liver microsomes were prepared by the method of Lu and Lerin (24), and stored as a suspension in 0.25 M
sucrose at -80°C. Cytosol was prepared as the post- microsomal supernatant fraction by the same proce- dure, except that livers were first perfused with ho- mogenization buffer (50 mM Tris-HCI, pH ‘7.4, at 4”C, containing 150 rnM KCI and 2 rnbc EDTA) to remove blood.
Protein purQiecrtim. All proteins were purified from liver microsomes from Aroclor 1254-induced Long-Evans rats. Cytochromes P450b and P45Oc were purified to electrophoretic homogeneity as described by Ryan et a/. (22), with modifications described pre- viously (15). One of t.hese modifications was an exten- sion of the NaCl gradient applied to the initial What- man DE 52 anion-exchange column to elute cyto- chrome bS, which was further purified essentially as described by Waxman and Walsh (25). Purified cyto- chrome bS had a specific content of -42-44 nmol/mg protein, was free of contamination with NADPH-cy- tochrome P450 reductase. and migrated as single
polppeptide when subjected to SDS-PAGE’ in lo- 15% acrglamide gels. Cytochrome P450e was purified as described by Ryan ef ul. (26i, except the purified protein was not subjected to immunoaffinity chroma- tography with antibody against cytochrome P45Oc. Despite this omission, cytochrome P450e was con- taminated with less than 27 cytochrome P45Oc, based on its catalytic activity toward ‘7-ethoxyresoru- fin and the results of SDS-PAGE. NADPH-cyto- chrome P450 reductase was purified as described by Yasukochi and Masters (27), with modifications re- ported previously (15 ). One nanomole of purified en- zyme reduced -3 pmol cytochrome c per minute at 22”C, in the presence of 330 rnM potassium phosphate buffer (pH 7.4), 1 mM EDTA. 3 mM MgC&, 100 PM
KCN, 50 pM cytochrome c, and 100 pM NADPH. All purified proteins were stored at ~80°C.
rl~tibody protlwtiorr u)rd ioc rn1,~tonbscllrptiotc. Anti- bodies against cytochromes P450b and P-450~ were raised in male New Zealand White rabbits, essen- tially as described by Thomas et ~1. (16, 28). The IgG fraction was isolated from high-titer antiserum (de- termined by ELI%) as described by McKinney and Parkinson (29). Each IgG preparation was subjected to immunoabsorption chromatography to remove an- tibody that unavoidably cross-reacted with other cy- tochrome P-150 isozymes with shared epitopes (anti- genie determinantsj (30). The procedure used was a modification of that described by Thomas ef trl. (16, 31). Three columns (2.5 X 15 cm) of A’-octylamino- Sepharose were prepared as described by Guengerich (32), and were equilibrated with 350 ml of 100 rnM po- tassium phosphate butfer, pH 7.4, containing 100 GM EDTA. Liver microsomes from control and phenobar- bital- or 3-methylcholanthrene-treated rats (500 nmol cytochrome P450) were diluted to 10 mg pro- tein/ml in 100 rnhf potassium phosphate buffer, pH 7.4, containing 204 glycerol and 100 PM EDTA, and were solubilized with sodium cholate (final concen- tration 2? ). Each sample was diluted with 9 vol of equilibration buffer (to give 1 mg protein/ml and 0.2’7 cholatei and loaded onto an ?J-octylamino- Sepharose column. Columns I, II, and III contained liver microsomes from control and phenobarbital- and 3-methylcholanthrene-treated rats, respectively. Partially purified cytochrome P450d. present in the void volume from the Whatman DE-52 anion-es- change column used to purify cytochromes P450h, P45Oc, and P45Oe (26.31). was also loaded onto column II (300 nmol total cytochrome P450). Each column was washed with 350 ml equilibration buffer to re- move sodium cholate before antibody (diluted to 5 mg/ml in equilibration buffer) was passed through the column. Rabbit anti-rat cytochrome P450b (1.7 g IgG protein) was passed through columns I and III.
’ Abbreviations used: SDS-PAGE, sodium dodecgl sulfate-polyacrylamide gel electrophoresis; ELI%, enzyme-linked immunosorbent assay.

620 DUTTON AND PARKINSON
Rabbit anti-rat cytochrome P45Oc (1.5 g IgG protein) was passed through column II. Columns were washed with 350 ml equilibration buffer, followed by 150 ml of 2 M KSCN (to elute specifically bound antibody). Column eluant was monitored at 280 nm, collected in 20-ml fractions, and screened for specificity by ELISA and Western immunoblot (33-35). Those frac- tions from column II that recognized only cytochrome P45Oc were pooled, as were fractions from columns I and III that recognized only cytochromes P450b and P450e (NB: cytochromes P450b and P450e share 97% amino acid sequence homology (36,37) and are indis- tinguishable with polyclonal antibody against cyto- chrome P450b (1, 30). The pooled fractions were treated with ammonium sulfate (277 mg/ml) to pre- cipitate IgG, which was resuspended in and dialyzed against phosphate-buffered saline. The dialyzed sam- ples were treated with XAD-2 beads (100 mg/ml [wet weight] for 1 hr), diluted or concentrated to 20 mg/ml (based on Em of 1.3 for 1 mg IgG/ml), and stored at -20°C.
Enzyme assays. 7-Alkoxyresorufin 0-dealkylase activity was measured by the fluorometrie method of Burke and Mayer (38). with minor modifications (39). Reactions were carried out at 37°C in l- or 3-ml incu- bation mixtures containing potassium phosphate buffer (100 ITIM, pH 7.4), MgC& (3 IKIM), EDTA (1 mM), NADP (1 mM), glucose 6-phosphate (5 mM), glucose- 6-phosphate dehydrogenase (1 unit/ml), and either 7- pentoxyresorufin or 7-ethoxyresorufin (l-10 PM). Liver microsomes were incubated at 0.05-1.0 mg pro- tein/ml. Purified cytochromes P450 were incubated at 20-200 pmol/ml in the presence of NADPH-cyto- chrome P450 reductase (0.05-1.0 nmol/ml) and di- lauroylphoshatidylcholine (usually 15pMj. at the final concentrations indicated. Reactions were started by
addition of the NADPH-generating system. Forma- tion of resorufin in the 3-ml incubation mixtures was monitored continuously as the increase in fluores- cence intensity (hexeitatiun - 535 nm, Xpmi39i,,n - 585 nm) with a Shimadzu RF 540 fluorometer. Formation of resorufin in the l-ml incubation mixtures was mea- sured after the reactions were stopped with 2 ml ice- cold acetone following a l- to lo-min incubation, as described (39). The latter method allows any reduced resorufin to reoxidize, which restores the fluorescent properties of resorufin (23).
In all cases, experiments were performed to verify that product formation was proportional to incuba- tion time and cytochrome P450 concentration. For antibody-inhibition studies, microsomes were prein- cubated for 5 min at 37°C with rabbit anti-rat cyto- lauroylphoshatidylcholine (usually 15 PM), at the final concentration of antibody was adjusted to 1 mg/ml with an IgG fraction purified from preimmune rabbit
serum. Other assays. Protein concentration was measured
by the method of Lowry et al. (401, with bovine serum albumin as standard. The concentration of cyto-
chrome P450 was determined by the method of Omura and Sato (41), from the carbon monoxide difference spectrum of dithionite-reduced micro- somes, based on an extinction coefficient of 91 mMm' cm-l. The concentration of cytochrome b5 was deter- mined from the difference spectrum between oxidized and reduced cytochrome b5, based on an extinction co- efficient of 185 rtIM-' cm-l (41). Microsomal cyto- chrome b5 was reduced by addition of 100 pM NADH, whereas purified cytochrome b5 was reduced with so- dium dithionite. The activity of DT-diaphorase, NAD(P)H-quinone oxidoreductase (EC 1.6.99.2), was measured with resorufin as substrate, as described by Nims et al. (42).
RESULTS AND DISCUSSION
7’-Eth.oxyresorufin 0-Dea.lkylation
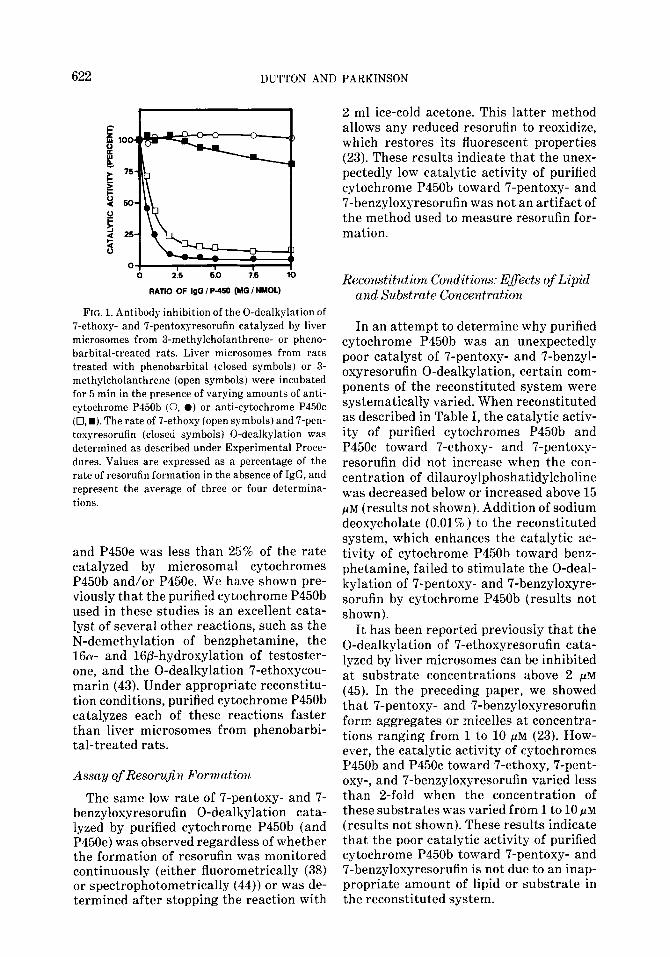
Treatment of rats with the cyto- chrome P45Oc inducer, S-methylcholan- threne, caused a 33-fold increase in the rate of 7-ethoxyresorufin 0-dealkylation catalyzed by liver microsomes (calculated on a per milligram of protein basis), whereas treatment of rats with phenobar- bital caused less than a 2-fold increase (Table I). Antibody against cytochrome P45Oc inhibited greater than 85% of the 7-ethoxyresorufin 0-dealkylation cata- lyzed by liver microsomes from 3-methyl- cholanthrene-treated rats, as shown in Fig. 1. When reconstituted with saturating amounts of NADPH-cytochrome P450 re- ductase and optimal amounts of lipid, puri- fied cytochrome P45Oc catalyzed the O-de- alkylation of 7-ethoxyresorufin at a rate of 30 nmol/nmol P450/min (Table I). The cat- alytic activity of purified cytochrome P45Oc toward 7-ethoxyresorufin exceeded by at least an order of magnitude the cata- lytic activity of liver microsomes (regard- less of the source) and of purified cyto- chromes P450b and P450e. Even when one takes into account the fact that cyto- chrome P45Oc comprises only -50% of the total cytochrome P450 in liver microsomes from 3-methylcholanthrene-treated rats (16), purified cytochrome P45Oc was still at least 5 times more active than microsomal cytochrome P45Oc at catalyzing ?-ethoxy- resorufin 0-dealkylation.
T-Pentoxyresomjin 0-Dea.bkylatim
Treatment of rats with the cyto- chrome P450b/P450e inducer, phenobarbi-

7-ALKOXYRESORLIFIN 0-DEALKYLATION BY PURIFIED CYTOCHROME P450 621
TABLE I
O-DEALKYLATIONOF'~-ETHOXY-AND~-PENTOXYRESORUFINBY RATLIVERMICROSOMES ANDBYPURIFIEDCYTOCHROMEP~~O
Enzymesystem i’-Ethoxpresorufm” 7-Pentoxyresorufin”
Microsomesb Control
Phenobarbital
3-Methylcholanthrene
Purified isozyme Qtochrome P450b Cytochrome P45Oc Cytochrome P450e
97 + 6 12 + 1 (109+7) (13kl)
79 f 5 453 + 60 (183 f 11) (1050 + 140 1 2370 _t 150 30 ‘- 3
(3630 k 230) (46 f 5)
230 200 30,000 150
600 50
Arote. Liver microsomes from control rats were incubated at a final concentration of 1 mg protein/ml with both substrates, as described under Experimental Procedures. Microsomes from phenobarbital-treated rats were incubated at 1.0 and 0.05 mg/ml with 7-ethoxy- and 7-pentoxyresorufin, respectively, whereas micro- somes from 3-methglcholanthrene-treated rats were incubated with these substrates at 0.05 and 1.0 mg/ml, respectively. Purified cytochrome P450b and P450e were incubated at a final concentration of 0.1 to 0.2 nmol/ ml with both substrates. Purified cgtochrome P45Oc was incubated at 0.02 and 0.2 nmol/ml for 7-ethoxp- and 7-pentoxyresorufin, respectively. Incubation mixtures with purified cytochrome P450 also contained 1.0 nmol/ ml NADPH-cytochrome P450 reductase and 15 P&I dilauroylphosphatidylcholine.
’ Values are pmol resort& formed/nmol cytochrome P-450/min. Values in parentheses are pmol resorufin formed/mg microsomal proteinjmin. Values obtained with microsomes represent the means + SE of four determinations. Values for the purified cytochromes P450 isozymes represent the average of at least two sepa- rate determinations carried out in triplicate (which deviated less than 5q, 1.
‘Liver microsomes from control and phenobarbital- or 3-methylcholanthrene-treated rats contained 1.12, 2.32, and 1.53 nmol cytochrome P450/mg protein, respectively.
t.al, caused an M-fold increase in the rate of 7-pentoxyresorufin 0-dealkylation cata- lyzed by liver microsomes, whereas treat- ment with 3-methylcholanthrene caused less than a 4-fold increase (Table I). Anti- body against cytochrome P450b (and P450e) inhibited greater than 90% of the 0-dealkylation of 7-pentoxyresorufin cata- lyzed by liver microsomes from phenobar- bital-treated rats (Fig. 1). The results in Fig. 1 clearly established that the induc- tion of cytochromes P450b and/or P450e was responsible for the marked increase in 7-pentoxyresorufin 0-dealkylation caused by treatment of rats with phenobarbital. However, when reconstituted with satu- rating amounts of NADPH-cytochrome P450 reductase and optimal amounts of lipid, cytochromes P450b and P450e were unexpectedly poor catalysts of the O-deal- kylation of 7-pentoxyresorufin (Table I). Results identical to those shown in Table
I for 7-pentoxyresorufin were obtained when 7-benzyloxyresorufin was the sub- strate (results not shown).
The catalyt.ic activity of purified cyto- chrome P450b and P450e toward 7-pen- toxyresorufin was actually less than their catalytic activity toward 7-ethoxyresoru- fin (Table I). Furthermore, the rate of 7- pentoxyresorufin 0-dealkylation catalyzed by purified cytochromes P450b and P450e was comparable to that catalyzed by puri- fied cytochrome P45Oc, and was less than half the rate of 7-pentoxyresorufin O-deal- kylation catalyzed by liver microsomes from phenobarbital-treated rats (based on per nanomole of P450 basis). If one takes into account the fact that cytochromes P450b and P450e comprise -50%’ of the to- tal cytochrome P450 in liver microsomes from phenobarbital-treated rats (16), the rate of 7-pentoxyresorufin 0-dealkylation catalyzed by purified cytochromes P450b
622 DUTTON AND
RATIO OF IgG / P-450 (MG I NMOL)
FIG. 1. Antibody inhibition of the 0-dealkylation of 7-ethoxy- and 7-pentoxyresorufin catalyzed by liver microsomes from 3-methylcholanthrene- or pheno- barbital-treated rats. Liver microsomes from rats treated with phenobarbital (closed symbols) or 3- methylcholanthrene (open symbols) were incubated for 5 min in the presence of varying amounts of anti- cytochrome P450b (0, 0) or anti-cytochrome P45Oc (0, w). The rate of 7-ethoxy (open symbols) and 7-pen- toxyresorufin (closed symbols) 0-dealkylation was determined as described under Experimental Proce- dures. Values are expressed as a percentage of the rate of resorufin formation in the absence of IgG, and represent the average of three or four determina- tions.
and P450e was less than 25% of the rate catalyzed by microsomal cytochromes P450b and/or P450e. We have shown pre- viously that the purified cytochrome P450b used in these studies is an excellent cata- lyst of several other reactions, such as the N-demethylation of benzphetamine, the 16~~ and 16P-hydroxylation of testoster- one, and the 0-dealkylation 7-ethoxycou- marin (43). Under appropriate reconstitu- tion conditions, purified cytochrome P450b catalyzes each of these reactions faster than liver microsomes from phenobarbi- tal-treated rats.
Assay of Resorujh Forma.tio?l.
The same low rate of ‘I-pentoxy- and 7- benzyloxyresorufin 0-dealkylation cata- lyzed by purified cytochrome P450b (and P450e) was observed regardless of whether the formation of resorufin was monitored continuously (either fluorometrically (38) or spectrophotometrically (44)) or was de- termined after stopping the reaction with
PARKINSON
2 ml ice-cold acetone. This latter method allows any reduced resorufin to reoxidize, which restores its fluorescent properties (23). These results indicate that the unex- pectedly low catalytic activity of purified cytochrome P450b toward 7-pentoxy- and 7-benzyloxyresorufin was not an artifact of the method used to measure resorufin for- mation.
Recorrstitution Conditions: Eflects of Lipid and Substrate Concentration
In an attempt to determine why purified cytochrome P450b was an unexpectedly poor catalyst of 7-pentoxy- and 7-benzyl- oxyresorufin 0-dealkylation, certain com- ponents of the reconstituted system were systematically varied. When reconstituted as described in Table I, the catalytic activ- ity of purified cytochromes P450b and P45Oc toward 7-ethoxy- and 7-pentoxy- resorufin did not increase when the con- centration of dilauroylphoshatidylcholine was decreased below or increased above 15 PM (results not shownj. Addition of sodium deoxycholate (0.01 lo ) to the reconstituted system, which enhances the catalytic ac- tivity of cytochrome P450b toward benz- phetamine, failed to stimulate the O-deal- kylation of 7-pentoxy- and 7-benzyloxyre- sorufin by cytochrome P450b (results not shown).
It has been reported previously that the 0-dealkylation of 7-ethoxyresorufin cata- lyzed by liver microsomes can be inhibited at substrate concentrations above 2 PM
(45). In the preceding paper, we showed that 7-pentoxy- and 7-benzyloxyresorufin form aggregates or micelles at concentra- tions ranging from 1 to 10 PM (23). How- ever, the catalytic activity of cytochromes P450b and P45Oc toward 7-ethoxy, 7-pent- oxy-, and 7-benzyloxyresorufin varied less than 2-fold when the concentration of these substrates was varied from 1 to 10 FM
(results not shown). These results indicate that the poor catalytic activity of purified cytochrome P450b toward 7-pentoxy- and 7-benzyloxyresorufin is not due to an inap- propriate amount of lipid or substrate in the reconstituted system.

7-ALKOXYRESORUFIN 0-DEALKYLATION BY PURIFIED CYTOCHROME P450 623
Reconstitu,tion Gmditions: Erects of Cytoch rome b5
Previous studies have shown that the catalytic activity of certain forms of cyto- chrome P450 toward certain substrates is markedly influenced by addition of cyto- chrome b5 to the reconstituted system. For example, the K,n for the N-demethylation of dimethylnitrosamine by purified cyto- chrome P45Oj decreases in the presence of cytochrome b5, and thereby resembles the high affinity enzyme that catalyzes this re- action in liver microsomes (46). Cyto- chrome b5 caused a concentration-depen- dent decrease in the rate of 7-ethoxy- and 7-pentoxyresorufin 0-dealkylation cata- lyzed by purified cytochromes P450b and P45Oc, respectively (results not shown). Cytochrome b5 has been shown previously to inhibit the catalytic activity of purified isozymes of cytochrome P450 (47). These results indicate that the poor catalytic ac- tivity of purified cytochrome P450b toward 7-pentoxyresorufin does not reflect a re- quirement for cytochrome bs.
Recovstit,zrtiov Conditions: Effects qf NADPH-Cytochrome PAiO Reductase
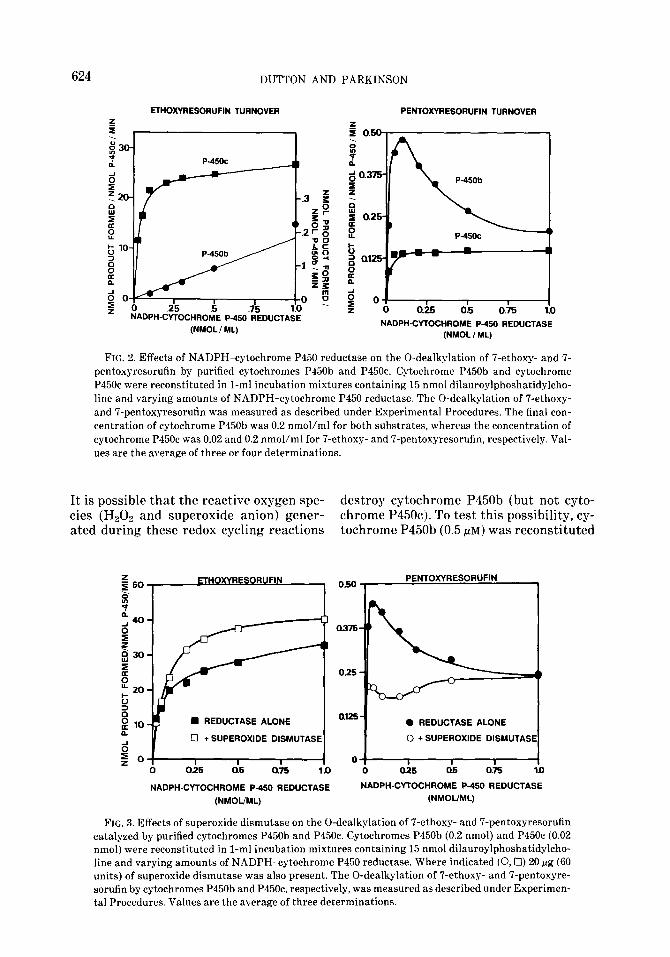
The effects of varying the concentration of NADPH-cytochrome P450 reductase on the 0-dealkylation of ‘7-ethoxy- and 7-pent- oxyresorufin by cytochromes P450b and P45Oc are shown in Fig. 2. The rate of 7- ethoxyresorufin 0-dealkylation catalyzed by cytochrome P45Oc (0.02 nmol/ml) in- creased rapidly as the concentration of NADPH-cytochrome P450 reductase was increased from 0.025 to 0.10 nmol/ml, and increased gradually as the concentration of reductase was increased further to 1.0 nmol/ml. The rate of 7-pentoxyresorufin 0-dealkylation catalyzed by cytochrome P45Oc (0.2 nmol/ml) was less than l%, of that with 7-ethoxyresorufin, and the rate did not increase further as the concentra- tion of NADPH-cytochrome P450 reduc- tase was increased above 0.1 nmol/ml (Fig. 2). An import.ant difference between the O- dealkylation of 7-ethoxyresorufin and 7- pentoxyresorufin by purified cytochrome P45Oc was the molar ratio of reductase to
P45Oc required for maximal rates of O-de- alkylation. Whereas equimolar amounts of NADPH-cytochrome P450 reductase and cytochrome P45Oc supported maximal rates of 7-pentoxyresorufin O-dealkyl- ation, the rate of7-ethoxyresorufin O-deal- kylation could not be fully saturated even with a 50-fold molar excess of reductase over cytochrome P45Oc.
The rate of 7-ethoxyresorufin O-dealkyl- ation cat.algzed by purified cytochrome P450b (0.2 nmol/ml) also increased as the concentration of NADPH-cytochrome P450 reductase was increased up to 1.0 nmol/ml, although the highest rate ob- tained was less than 1% of that catalyzed by purified cytochrome P45Oc (Fig. 2). In contrast, the rate of 7-pentoxyresorufin O- dealkylation catalyzed by cytochrome P450b (0.2 nmol/ml) increased as the con- centration of NADPH-cytochrome P450 reductase increased up to 0.1 nmol/ml, at which point the molar ratio of cytochrome P450b to NADPH-cytochrome P450 reduc- tase was 2:l. Increasing the concentration of NADPH-cytochrome P450 reductase further caused a progressive decline in the cytochrome P450b-dependent O-dealkyl- ation of 7-pentoxyresorufin. It should be emphasized, however, that even when the molar ratio of P450b to reductase was 2:1, cytochrome P450b was still a relatively poor catalyst of 7-pentoxyresorufin O-de- alkylation. It should also be emphasized that a molar excess of NADPH-cyto- chrome P450 reductase does not inhibit the catalytic activity of cytochrome P450b to- ward other substrates, such as benzphe- tamine, testosterone, and 7-ethoxycouma- rin (6, 22, 43). Results identical to those shown in Fig. 2 for 7-pentoxyresorufin were obtained when 7-benzyloxyresorufin was the substrate (results not shown ).
Destrwtkw qfcytochrome PJX
The observation that a molar excess of NADPH-cytochrome P450 reductase in- hibited the 0-dealkylation of 7-pentoxy- and 7-benzyloxyresorufin by cytochrome P450b (and P450e) led to the discovery that NADPH-cytochrome P450 reductase initi- ates the redox cycling of resorufin and the 7-alkoxyresorufins (see preceding paper).
624 DUTTON AND PARKINSON
PENTOXVRESORlJFlN TURNOVER ETHOXVRESORUFIN TURNOVER
z I ,
.i5 .6 I-CYTOCHROYE P
(NMOL / ML)
‘ASE 0.25 05 0.15 1.0
NADPH-CYTOCHROME P-450 REDUCTASE (NMOL / ML)
FIG. 2. Effects of NADPH-cytochrome P450 reductase on the 0-dealkylation of i’-ethoxy- and 7- pentoxyresorufin by purified cytochromes P450b and P45Oc. Cytochrome P450b and cytochrome P45Oc were reconstituted in l-ml incubation mixtures containing 15 nmol dilauroylphoshatidylcho- line and varying amounts of NADPH-cytochrome P450 reductase. The 0-dealkylation of 7-ethoxy- and 7-pentoxyresorufin was measured as described under Experimental Procedures. The final con- centration of cytochrome P450b was 0.2 nmol/ml for both substrates, whereas the concentration of cytochrome P45Oc was 0.02 and 0.2 nmol/ml for 7-ethoxy- and 7-pentoxyresorufin, respectively. Val- ues are the average of three or four determinations.
It is possible that the reactive oxygen spe- destroy cytochrome P450b (but not cyto- ties (H202 and superoxide anion) gener- chrome P45Ocj. To test this possibility, cy- ated during these redox cycling reactions tochrome P450b (0.5 PM) was reconstituted
ETHOXVRESORUFIN , 0.50
025
n REDUCTASE ALONE a12!?
0 + SUPEROXIDE DISMUTASE
Cl25 a5 0.75 1.0 i
0
PENTOXYRESORUFIN
0 REDUCTASE ALONE
0 + SUPEROXIDE DISMUTASI
o& ci5 0.k
NADPH-CVTOCHROME P-450 REDUCTASE NADPH-CVTOCHROME P-450 REDUCTASE
(NMOUML) (NMOUML)
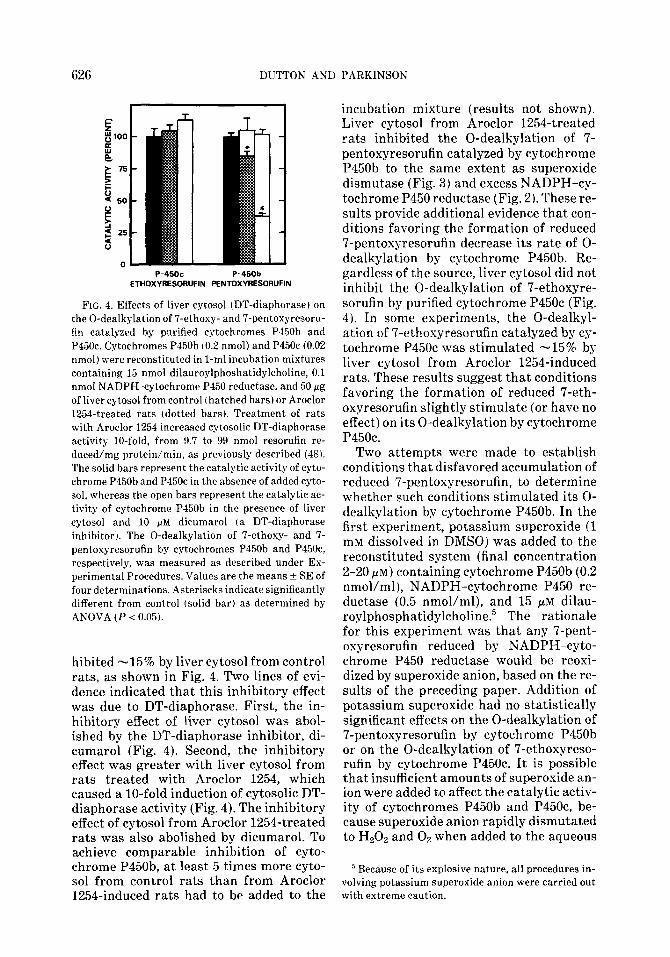
FIG. 3. Effects of superoxide dismutase on the 0-dealkylation of 7-ethoxy- and 7-pentoxyresorufin catalyzed by purified cytochromes P450b and P45Oc. Cytochromes P450b (0.2 nmol) and P45Oc (0.02 nmol) were reconstituted in l-ml incubation mixtures containing 15 nmol dilauroylphoshatidylcho- line and varying amounts of NADPH-cytochrome P450 reductase. Where indicated (0,O) 20 fig (60 units) of superoxide dismutase was also present. The 0-dealkylation of ‘I-ethoxy- and IT-pentoxyre- sorufin by cytochromes P450b and P45Oc, respectively, was measured as described under Experimen- tal Procedures. Values are the average of three determinations.

7-ALKOXYRESORUFIN 0-DEALKYLATION BY PURIFIED CYTOCHROME P-150 625
with 15 pM dilauroylphoshatidylcholine, 0.5 pM NADPH-cytochrome P450 reduc- tase, and 5 pM pentoxyresorufin. The con- centration of cytochrome P450 was deter- mined by the method of Omura and Sato (41j before and after a 5-min incubation with 100 pM NADPH. Less than 5% of cy- tochrome P450b was converted to the cata- lytically inactive form, cytochrome P420, during the 5-min incubation (results not shown). This result established that the redox cycling reactions supported by NADPH-cytochrome P450 reductase do not inactivate cytochrome P450b to a sig- nificant extent.
Reconstitutiovl. Conditions: Efects of Ca talase a.nd Superoxide Dismutase
To test further the possibility that reac- tive oxygen species generated during the redox cycling of resorufin and the 7-alk- oxyresorufins destroy cytochrome P450b (but not cytochrome P45Oc j, superoxide dismutase and catalase were added to the reconstitution system. Catalase (480 units/ml) had little or no effect on the cyto- chrome P450b-dependent 0-dealkylation of 7-pentoxyresorufin or on the cyto- chrome P450c-dependent 0-dealkylation of 7-ethoxyresorufin, which further indi- cated that these isozymes were not inacti- vated by any Hz02 formed (results not shownj. The effects of superoxide dismu- tase on the catalytic activity of cyto- chromes P450b and P45Oc are shown in Fig. 3. Unexpectedly, superoxide dismu- tase inhibited, rather than stimulated, the 0-dealkylation of 7-pentoxyresorufin (and 7-benzyloxyresorufin) by cytochrome P450b. This result confirmed that cyto- chrome P450b was not inactivated by any superoxide anion formed, but raised the question: Why does superoxide dismutase inhibit the catalytic activity of cytochrome P450b toward 7-pentoxyresorufin? The degree to which superoxide dismutase in- hibited cytochrome P450b decreased as the concentration of NADPH-cytochrome P450 reductase increased. Superoxide dis- mutase caused a -50% inhibition of cyto- chrome P450b reconstituted with 0.1 nmol/ml NADPH-cytochrome P450 re- ductase, but caused no inhibition with 1.0
nmol/ml reductase. In contrast to its effects on cytochrome P450b, superoxide dismutase stimulated the 0-dealkylation of 7-ethoxyresorufin by cytochrome P45Oc (Fig. 3). When cytochrome P45Oc was re- constituted with 1.0 nmol NADPH-cyto- chrome P450 reductase, superoxide dismu- tase increased the rate of 7-ethoxyresoru- fin 0-dealkylation to 40 nmol/nmol P45Oc/ min, which is the highest turnover rate re- ported for this reaction.
Kewv&tution Cbnditiow Effects of Liwv Cytosol (DT-d,iaphorase), Su,peroxide Anion, and Ckmene Hydroperoxide
In terms of their catalytic activity to- ward 7-ethoxy- and 7-pentoxyresorufin, the results in Figs. 3 and 4 indicate that cytochromes P45Oc and P450b respond op- positely to the presence of a molar excess of NADPH-cytochrome P450 reductase and to the presence of superoxide dismu- tase. Both these conditions favor the ac- cumulation of the reduced form of the 7-alkoxyresorufins, as demonstrated in the preceding paper (23). NADPH-cyto- chrome P450 reductase increases the rate of formation of reduced 7-alkoxyresoru- fins, whereas superoxide dismutase de- creases their rate of disappearance (due to the ability of superoxide anion to reoxidize the reduced form of the 7-alkoxyresorufins (23)). The following experiments were de- signed to test the hypothesis that condi- tions favoring the accumulation of reduced 7-pentosyresorufin inhibit its O-dealkyl- ation by qtochrome P450b, whereas condi- tions favoring the accumulation of reduced 7-ethoxyresorufin slight.ly stimulate its O- dealkylation by cytochrome P45Oc.
The 0-dealkylation of 7-ethoxy- and 7- pentoxyresorufin by cgtochromes P45Oc and P450b, respectively, was determined in the presence and absence of liver cytosol. Due to the presence of DT-diaphorase, ad- dition of liver cytosol favors formation of the reduced form of 7-ethoxgresorufin and 7-pentoxyresorufin. Cytochromes P450b and P45Oc were reconstituted under those conditions that responded maximally to the effects of superoxide dismutase (Fig. 3). The 0-dealkylation of 7-pentoxyresoru- fin by purified cyt,ochrome P450b was in-
626 DUTTON AND PARKINSON
P-45oc P-450b ETHOXYRESORUFIN PENTOXYRESORUFIN
FIG. 4. Effects of liver cytosol (DT-diaphorase) on the 0-dealkylation of ‘i-ethoxy- and 7-pentoxyresoru- fin catalyzed by purified cytochromes P450b and P45Oc. Cytochromes P150b (0.2 nmol) and P45Oc (0.02 nmol) were reconstituted in l-ml incubation mixtures containing 15 nmol dilauroylphoshatidylcholine, 0.1 nmol NADPH-cytochrome Pi50 reductase, and 50 Fg of liver cytosol from control (hatched bars) or Aroclor 12Wtreated rats (dotted bars). Treatment of rats with Aroclor 1254 increased cytosolic DT-diaphorase activity lo-fold, from 9.7 to 99 nmol resorufin re- duced/mg protein/min, as previously described (48). The solid bars represent the catalytic activity of cyto- chrome P450b and P45Oc in the absence of added cyto- sol, whereas the open bars represent the catalytic ac- tivity of cytochrome PS5Ob in the presence of liver cytosol and 10 FM dicumarol (a DT-diaphorase inhibitor’). The 0-dealkylation of ‘I-ethoxg- and 7- pentoxyresorufin by cptochromes P450b and P45Oc, respectively, was measured as described under Ex- perimental Procedures. Values are the means + SE of four determinations. Asteriscks indicate significantly different from control (solid bar) as determined by ANOVA (P < 0.05).
hibited -15% by liver cytosol from control rats, as shown in Fig. 4. Two lines of evi- dence indicated that this inhibitory effect was due to DT-diaphorase. First, the in- hibitory effect of liver cytosol was abol- ished by the DT-diaphorase inhibitor, di- cumarol (Fig. 4). Second, the inhibitory effect was greater with liver cytosol from rats treated with Aroclor 1254, which caused a lo-fold induction of cytosolic DT- diaphorase activity (Fig. 4). The inhibitory effect of cytosol from Aroclor 1254-treated rats was also abolished by dicumarol. To achieve comparable inhibition of cyto- chrome P450b, at least 5 times more cyto- sol from control rats than from Aroclor 1254-induced rats had to be added to the
incubation mixture (results not shown). Liver cytosol from Aroclor 1254-treated rats inhibited the 0-dealkylation of 7- pentoxyresorufin catalyzed by cytochrome P450b to the same extent as superoxide dismutase (Fig. 3) and excess NADPH-cy- tochrome P450 reductase (Fig. 2 ). These re- sults provide additional evidence that con- ditions favoring the formation of reduced 7-pentoxyresorufin decrease its rate of O- dealkylation by cytochrome P450b. Re- gardless of the source, liver cytosol did not inhibit the 0-dealkylation of 7-ethoxyre- sorufin by purified cytochrome P45Oc (Fig. 4). In some experiments, the O-dealkyl- ation of ?-ethoxyresorufin catalyzed by cy- tochrome P45Oc was stimulated -15% by liver cytosol from Aroclor 1254-induced rats. These results suggest that conditions favoring the formation of reduced 7-eth- oxyresorufin slightly stimulate (or have no effect) on its 0-dealkylation by cytochrome P45Oc.
Two attempts were made to establish conditions that disfavored accumulation of reduced 7-pentoxyresorufin, to determine whether such conditions stimulated its O- dealkylation by cytochrome P450b. In the first experiment, potassium superoxide (1 mM dissolved in DMSO) was added to the reconstituted system (final concentration 2-20 PM) containing cytochrome P450b (0.2 nmol/ml), NADPH-cytochrome P450 re- ductase (0.5 nmol/ml), and 15 PM dilau- roylphosphatidylcholine.5 The rationale for this experiment was that any 7-pent- oxyresorufin reduced by NADPH-cyto- chrome P450 reductase would be reoxi- dized by superoxide anion, based on the re- sults of the preceding paper. Addition of potassium superoxide had no statistically significant effects on the 0-dealkylation of 7-pentoxyresorufin by cytochrome P450b or on the 0-dealkylation of 7-ethoxyreso- rufin by cytochrome P45Oc. It is possible that insufficient amounts of superoxide an- ion were added to affect the catalytic activ- ity of cytochromes P450b and P45Oc, be- cause superoxide anion rapidly dismutated to H202 and OZ when added to the aqueous
’ Because of its explosive nature, all procedures in- volving potassium superoxide anion were carried out with extreme caution.

T-ALKOXYRESORUFIN 0-DEALKYLATION BY PURIFIED CYTOCHROME P450 627
incubation mixture (inferred from the effervescent reaction). Attempts to stimu- late the catalytic activity of cytochrome P450b with superoxide anion generated en- zymatically with xanthine and xanthine oxidase were also unsuccessful, mainly be- cause xanthine itself inhibited the cata- lytic activity of cytochrome P450b (and P45Oc) (results not shown).
The second attempt to establish condi- tions that disfavored formation of reduced 7-pentoxyresorufin involved substituting NADPH and NADPH-cytochrome P450 reductase with cumene hydroperoxide. Previous studies have shown that cyto- chrome P450 can function as a peroxidase, such that oxygen from various organic hy- droperoxides can be used to generate the tertiary complex between cytochrome P-150, substrate, and activated oxygen that leads to hydroxylation of the substrate (49, 50). However, cumene hydroperoxide (final concentration 500 PM) did not effectively support the 0-dealkylation of 7-ethoxy- and 7-pentoxyresorufin by cytochrome P45Oc and P450b (results not shown), and led to a progressive conversion of these iso- zymes to cytochrome P420, as previously reported (51). As a result of these problems with superoxide anion and cumene hydro- peroxide, we were unable to establish ap- propriate conditions to determine whether conditions favoring formation of the oxi- dized form of 7-pentoxyresorufin would stimulate its 0-dealkglation by cyto- chrome P450b.
CONCLUSIONS
The antibody-inhibition experiments described in this study confirm that the in- duction of cytochrome P45Oc is largely re- sponsible for the marked increase in liver microsomal 7-ethoxyresorufin O-dealkyl- ation in rats treated with 3-methylcholan- threne, whereas the induction of cyto- chrome P450b and/or P450e in rats treated with phenobarbital is largely responsible for the marked increase in 7-pentoxy- and 7-benzyloxyresorufin 0-dealkylation (13, 14). In contrast to purified cytochrome P45Oc, which catalyzed the 0-dealkylation of 7-ethoxyresorufin with high turnover
when reconstituted with NADPH-cyto- chrome P450 reductase and lipid, purified cytochromes P450b and P450e were unex- pectedly poor catalysts of the O-dealkyl- ation of 7-pentoxy- and 7-benzyloxyreso- rufin. We have previously shown that the cytochrome P450b used in these experi- ments is an excellent catalyst of several other reactions, such as the N-demethyl- ation of benzphetamine, the 160~ and IS@- hydroxylation of testosterone, and the O- dealkylation of 7-ethoxgcoumarin (43). The low rate of 7-pentoxyresorufin O-deal- kylation catalyzed by purified cytochrome P450b did not reflect a requirement for cy- tochrome bsr and could not be ascribed to an artifact of the method used to measure the formation of resorufin. The catalytic activity of purified cytochrome P450b to- ward 7-pentoxyresorufin was consistently low over a range of substrate and lipid con- centrations, and was not stimulated by so- dium deoxycholate (which stimulates the N-demethylation of benzphetamine by pu- rified cytochrome P450b ).
As shown in Fig. 5, we propose that cyto- chrome P45Oc catalyzes the 0-dealkylation of both the oxidized and reduced forms of 7-ethoxyresorufin, with perhaps a slight preference for the reduced form. In con- trast, we propose that cytochrome P450b preferentially catalyzes the O-dealkyl- ation of the oxidized form of 7-pentoxy- and 7-benzyloxgresorufin. Conditions that favor formation of the reduced form of 7- pentoxyresorufin decrease its rate of O-de- alkylation by cytochrome P450b, whereas conditions that favor formation of the re- duced form of 7-ethoxyresorufin tend to stimulate its 0-dealkylation by cyto- chrome P45Oc. Such conditions include a molar excess of NADPH-cytochrome P450 reductase over cytochrome P450 (Fig. 2), the presence of superoxide dismutase (Fig. 3), and the presence of DT-diaphorase (Fig. 4). Both NADPH-cytochrome P450 reduc- tase and DT-diaphorase increase the rate of formation of the reduced form of the 7- alkoxyresorufins. In contrast, superoxide dismutase decreases their rate of disap- pearance, because superoxide anion, which is formed during the redox cycling reac- tions described in the preceding paper (23j, causes a rapid reoxidation of the reduced

628 DUTTON AND PARKINSON
0
ETHOXYRESORUFIN 0xbJ,z.3d ------& -r------ EniOxmEsoR”FIN md”Ced
I
0
P45OC P45OC
0 I RESORWIN Olldlzad --------------& RESOR”F,N reduced ~--------------
t
@
P450b P45Ob
0 T PEHTOxmEXWIUFlN oxkl!zed -----% PENlOnaEsoR"FlN red"cfd .z----- 8 FIG. 5. Proposed scheme for the 0-dealkylation of
the oxidized and reduced forms of 7-ethoxy- and 7- pentoxyresorufin by cytochromes P45Oc and P450b, respectively. As indicated by the thickness of the ar- rows, cytochrome P45Oc is proposed to catalyze the O- dealkylation of both the oxidized and reduced forms of 7-ethoxyresorufin (perhaps with a slight prefer- ence for the reduced form). In contrast, cgtochrome P450b is proposed to catalyze preferentially the O-de- alkylation of the oxidized form of 7-pentoxyresorufin (as well as ‘7-benzyloxyresorufin). The interconver- sion of the oxidized and reduced forms of resorufin and the 7-alkoxyresorufins (---) is based on the re- sults of the preceding paper (23). Reaction 1 is cata- lyzed by NADPH-cytochrome P450 reductase and DT-diaphorase, while reaction 2 is driven by Oe and superoxide anion radical.
form of the 7-alkoxyresorufins. For techni- cal reasons, we were unable to establish conditions that disfavored formation of the reduced form of the 7-alkoxyresorufins to test whether cytochrome P450b cata- lyzed the 0-dealkylation of oxidized 7- pentoxyresorufin with high catalytic turn- over. It should be emphasized that, in con- trast to its effects on the 0-dealkylation of 7-pentoxyresorufin, a molar excess of NADPH-cytochrome P450 reductase does not inhibit the catalytic activity of cyto- chrome P450b toward benzphetamine, tes- tosterone, or 7-ethoxycoumarin (43).
The reason reduction of 7-pentoxyreso- rufin and 7-benzyloxyresorufin decreases their rate of 0-dealkylation by cytochrome P450b, whereas reduction of 7-ethoxyre- sorufin slightly stimulates its O-dealkyl- ation by cytochrome P45Oc remains un- known. It is interesting that the oxidized forms of the 7-alkoxyresorufins are co-pla- nar molecules, whereas the reduced forms proposed in the proceeding paper are non co-planar molecules. It is possible that the active site of P450b accommodates only the
co-planar form of 7-pentoxyresorufin and 7-benzyloxyresorufin.
If cytochrome P450b preferentially cata- lyzes the 0-dealkylation of the oxidized form of 7-pentoxy- and 7-benzyloxyresoru- fin, it is significant that the molar ratio of cytochrome P450 to NADPH-cytochrome P450 reductase in liver microsomes is greater than 1O:l (52). We speculate that if rat liver microsomes did not contain a molar excess of cytochrome P450, the O- dealkylation of 7-pentoxy- and 7-benzyl- oxyresorufin would be of little value in monitoring the induction of cytochrome P450b and P450e by phenobarbital. Fi- nally, it has been proposed previously that the relatively low levels of NADPH-cyto- chrome P450 reductase in liver micro- somes may be a safeguard to protect hepa- tocytes from t.he potentially deleterious one-electron reduction reactions catalyzed by this flavoprotein. The results of this and the preceding study provide additional support for this proposal.
REFERENCES
1. RYAN, D. E., THOMAS, P. E., REIK, L. M., AND
LEVIN, W. (1982) Xe)zobioticu 12,727-744.
2. CONNEY, A. H. (1986) Life Sci. 39,2493-2518.
3. GUENGERICH, F. P., DANNAN, G. A., WRIGHT,
S. T., MARTIN, M. W., AND KAMINSKY, L. S.
(1982) Xexobiotico 12.701-716.
4. WOOD, W. A., RYAN, D. E., THOMAS, P. E., AND
LEVIN, W. (1983) J. Biol Chem. 258,8839-8847. 5. WAXMAN, D. J. (1988) Biochem. PharmacoL 37,
71-84. 6. SONDERFAN, A. J., ARLOTTO, M. P., DUTTON, D. R.,
MCMILLEN, S. K., AND PARKINSON, A. (1987)
Arch.. Biochem. Bioph ys. 255,27-41. 7. SWINNEY, D. C., RYAN, D. E., THOMAS, P. E., AND
LEVIN, W. (1987) Biochemistry 26,7073-7083. 8. CHENG, K. C., AND SCHENKMAN, J. B. (1984) Drug
hfetab. Dispos. 12.222-234.
9. LEVIN, W., WOOD, W. A., CHANG. R., RYAN, D. E.
THOMAS, P. E., YAGI, H., THAKKER, D., VYAS,
K., BOYD, C., CHU, S., CONNEY, A., AND JERINA,
D. (1982) Dwg hfetab. Rev. 13,555-580.
10. CONNEY, A. H. (1982) Cancer Res. 42,4875-4917. 11. KAMINSKY, L. S., FASCO. M. J., AND GUENGERICH,
F. P. (1980) J. Biol. Chem. 255,85-91. 12. GUENGERICH, F. P., DANNAN, G. A., WRIGHT,
S. T., MARTIN, M. W., AND KAMINSKY, L. S.
(1982) Biochemistry 21.6019-6030.
13. LUBET, R. A., MAYER, R. T., CAMERON, J. W., NIMS,
R. W.. BURKE, M. D., WOLFF, T., AND GUENGE-

7-ALKOXYRESORUFIN 0-DEALKYLATION BY PURIFIED CYTOCHROME P450 629
RICH, F. P. (1985) Arch.. Biochem. Biophys. 238, 43-48.
l-1. BURKE, M. D., THOMPSON, S. T., ELCOMBE, C. R., HALPERT, J., HAAPARANTA, T., AND MAYER, R. T. (1985) Biochem. Pharmacol. 34, 3337- 3345.
15. DUTTON, D. R., MCMILLEN, S. K., SONDERFAN, A. J., THOMAS, P. E., AND PARKINSON, A. (1987) Arch. Biochem. Biophys. 255.316-328.
16. THOMAS, P. E., REIK, L. M., RYAN, D. E., AND LEVIN, W. (1981) J. Biol. Chem. 256,1044-1052.
17. NEBERT, D. W.. ADESNIK, M., COON, M. J., ESTA- BROOK, R. W., GONZALEZ, F. J., GUENGERICH, F. P., GUNSALUS, I. C., JOHNSON, E. F., KEMPER, B., LEVIN, W., PHILLIPS, I. R., SATO, R., AND WATERMAN, M. R. (1987) DNA 6,1-11.
18. RYAN, D. E., IIDA, S., WOOD, A. W., THOMAS, P. E., LIEBER, C. S., AND LEVIN, W. (1984) J. Biol. Chem. 259, 1239-1250.
19. WAXMAN, D. J., Ko, A., AND WALSH, C. (1983) J. Biol. Chem. 258, 11937-11947.
20. KAMATAKI, T.. MAEDA, K., YAMAZOE. Y., NAGAI, T., AND KATO, R. (1982) Li$ Sci. 31,2603-2610.
21. DUTTON, D. R., AND PARKINSON, A. (1987) Fed. Proc. 46,1957.
22. RYAN, D. E., THOMAS, P. E., KORZENIOWSKI, D., AND LEVIN, W. ( 1979 j J. Biol. Chem. 254,1365- 1374.
23. DUTTON, D. R., REED, G. R.. AND PARKINSON, A. (1989) Arch. Biochem. Biophys. 268,605-616.
24. LLI, il. Y. H., AND LEVIN, W. (1972) Biochem. Birr ph y/s. Res. Corn w un. 46, 1334-1339.
25. WAXMAN, D. J., AND WALSH, C. (1982) J. Biol. C’ht~m. 257, 10416-10457.
26. RYAN, D. E., THOMAS, P. E., AND LEVIN, W. (1982) Arch. Biochem. Bioph ys. 216,272-288.
27. YASUKOCHI, Y., AND MASTERS, B. S. S. (1976) J. BioL Chem 251,5337-5344.
28. THOMAS, P. E., KORZENIO~SKI, D., RYAN, D. E., AND LEVIN, W. I 1979) Arch. Biochem. Bioph ys. 192.524-532.
29. MCKINNEY, M. M.. AND PARKINSON, A. (1987) J. Im to 71 nol. Methods 96, 271-278.
30. THOMAS, P. E., BANDIERA, S., REIK, L. M., MAINES, S. L., RYAN, D. E., AND LEVIN, W. I 1987) Fed. Proc. 46,2563-2566.
31. REIK, L. M., LEVIN. W., RYAN. D. E.. AND THOMAS, P. E. ( 1982) J. Biol. Chew. 257,3950-3957.
32. GUENGERICH, F. P. (1982) in Principals and Meth- ods of Toxicology (Hayes, W., Ed.) pp. 609-634, Raven Press, New York.
33. ENGVALL, E. (1980) in Methods in Enzymology (Van Vunakis, H., and Langone, J. J., Eds.), Vol. 70, Part A, pp. 419-439, Academic Press, San Diego.
34. TOBIN, H., ST?\EHELIN, T., AND GORDON, J. (1979) Proc. Natl. Acad. Sci. US.4 76.4350-4354.
35. THOMAS, P. E., REIK, L. M., RYAN, D. E., AND LENIN, W. (1984) J BioL Chem. 259,3890-3899.
36. YALIN, P. M., RYAN. D. E.. LEVIN, W., AND SHIVELY, J. E. (1983) Proe. XatL Acad. Sci. USA 80,1169-1173.
37. FUJI-KIJRIYAM-4, Y., MIZUKAMI, Y., KAWAJIRI, K., SOGARA, K., .~ND MURAMATSU, M. (1982) Proc. Ncrtl. Acad. Sci. USA 79,2793-2797.
38. BURKE, M. D., AND MAYER, R. T. (1974) Drug Metab. Dispos. 2,583-588.
39. GORSKI. J. R., ARLOTTO, M. P., KLAASSEN, C. D., AND PARKINSON, A. (1985) Carcinoge?u&s 6, 617-624.
40. LOWRY, 0. H., ROSEBROUGH, N. J., FARR, A. L., .~ND RANDALL, R. J. (1951) J. BioL Chem. 193, 265-275.
41. OMUR.~, T., AND SATO. R. (1964) J. Biol. Chem. 239, 2379-2385.
42. NIMS, R. W., PROUGH, R. A., END LUBET, R. A. (1984 J Arch. Biochem. Bioph ys. 299,459-465.
43. DLITTON, D. R., MCMILLEN, S. Ii., AND PARKINSON, rl. (1988) J. Biochem. Toxicol. 3, 131-145.
44. KLOTZ, A. V., STEGEMAN, J. J., AND WALSH, C. (1984) At/al. Biochem. 140,138-145.
45. BURKE, M. D., PROUGH, R. A., AND MAYER, R. T. (1977) Drug Metab. Dispos. 5, l-8.
46. LEVIN, W., THOMAS, P. E., OLDFIELD. N., AND RY.~N, D. E. (1986, Arch. Biochem. Biuphys. 248,158-165.
47. MORGAN, E. T., AND COON, M. J. (1984) Drug Metab. Dispos. 12,358-364.
48. NIMS, R. W., PROUGH, R. A., AND LUBET, R. -4. (1984) A&. Biochem. Biophys. 229,459-465.
49. WHITE, R. E., AND COON, M. J. (1980) .411nz(. Rev. Biochem. 49,315-356.
50. DAWSON, J. H. (1988) Sciexc 240,433-439. 51. PARKINSON, A., THOMAS, P. E., RYAN, D. E., GOR-
SKY, L. D., SHIVELY, J. E., SAYER, J. M., JERINA, D. M., AND LENIN, W. (1986) J. BioL Chem. 261. 11487-11495.
52. ESTABROOK, R. W., FRANKLIN, M. R., COHEN, B., SHIGAMATZU, A., AND HILDEBRANDT, A. G. (1971) Mefabolism 20, 187-199.





























