Embed Size (px)
Citation preview

1
2007-2008
Régulation de l’expression du génome, L5 BH05 Travaux dirigés : Expression du génome procaryote
Exercice n°1 : Opéron Lactose Avec l’étude de l’opéron lactose, François Jacob, Jacques Monod et André Lwoff (Prix Nobel de Physiologie et Médecine, 1965) ont été les premiers scientifiques à décrire un système de régulation de la transcription des gènes ; c’est à partir de leurs travaux qu’est né le concept-même de régulation génique. L’organisation de cet opéron lactose est le suivant : I P O Z Y A
1/ Redéfinir ce que sont ces régions I, P, O, Z, Y et A
β-Galactosidase Perméase Génotype Sans lactose Avec lactose Sans lactose Avec lactose
1 I+ P+ O+ Z+ Y+ / I+ P+ O+ Z+ Y+
2 I- P+ Oc Z+ Y- / I+ P+ O+ Z- Y+
3 I+ P- Oc Z- Y+ / I- P+ Oc Z+ Y-
4 Is P+ O+ Z+ Y- / I+ P+ O+ Z- Y+
5 Is P+ O+ Z+ Y+ / I- P+ O+ Z+ Y+
6 I+ P+ Oc Z+ Y- / I- P+ O+ Z- Y+
7 I- P- O+ Z+ Y+ / I- P+ Oc Z+ Y-
8 I+ P+ O+ Z- Y+ / I- P+ O+ Z+ Y-
2/ Sous chaque génotype, schématiser chacune des situations proposées. Compléter par « + » quand l’enzyme est produite et « - » quand elle ne l’est pas.
Exercice n°2 : Opéron Lactose, d’après Monod et al. (1947, The phenomenon of enzymatic adaptation, Growth symposium XI : 223-289) E. coli pousse plus vite sur du glucose (monosaccharide) que sur du lactose (disaccharide), pour 2 raisons : - le lactose est prélevé plus lentement que le glucose, et - le lactose doit d'abord être hydrolysé, par la β-galactosidase, en glucose et galactose avant d'être métabolisé plus avant. Quand on fait pousser E. coli sur un milieu contenant un mélange de glucose et de lactose, elle présente une cinétique de croissance complexe (symbole de figure : carrés blancs). Les bactéries croissent plus vite au début qu'à la fin, et il existe une

latence entre les 2 phases de croissance, pendant laquelle elles cessent quasiment de pousser. Des mesures expérimentales des concentrations des 2 sucres dans le milieu montrent que le glucose chute à des niveaux très faibles après quelques cycles cellulaires (cercles noirs), mais le lactose reste élevé quasiment jusqu'à la fin de l'expérience. Bien que la concentration en lactose soit élevée tout au long de l'expérience, la β-galactosidase, régulée avec l'opéron lac auquel elle appartient, n'est induite qu'après plus de 100 minutes (triangles gris).
1/ Expliquer la cinétique de croissance bactérienne au cours de l'expérience. Justifier la vitesse rapide de la croissance initiale, la vitesse plus lente de la croissance finale, et l'arrêt de croissance au milieu de l'expérience. 2/ Expliquer pourquoi l'opéron lac n'est pas induit par le lactose au cours de la phase rapide de croissance bactérienne.
Croissance d'E. coli sur un mélange de glucose et de lactose.
Exercice n°3 : Opéron arabinose En absence de glucose, E. coli peut métaboliser et pousser sur un milieu contenant de l’arabinose, en utilisant un ensemble inductible de gènes disposés en 3 groupes sur le chromosome.
Localisation chromosomique des gènes impliqués dans le métabolisme de l’arabinose.
Deux groupes de gènes codent des protéines impliquées dans le transport de l’arabinose. Dans le dernier groupe, les gènes araA, araB et araD codent des enzymes métabolisant l’arabinose, tandis que le gène araC code une protéine régulatrice qui se lie à côté des promoteurs arabinose et coordonne l’expression des gènes de cet opéron. Afin de comprendre les propriétés régulatrices de la protéine araC, vous isolez une bactérie mutante possédant une délétion du gène araC. Comme le montre le tableau suivant, la souche mutante n’exprime pas le produit du gène araA quand de l’arabinose est ajouté au milieu.
Produit du gène araA Sans arabinose Avec arabinose
araC+ 1 1000 araC- 1 1
Réponse de bactéries normales et mutantes à la présence ou à l’absence d’arabinose
1/ Les résultats de ce tableau suggèrent-ils que la protéine araC est un régulateur positif ou négatif de l’opéron arabinose ? 2/ A quoi ressembleraient les données de ce tableau si la protéine araC avait un effet opposé sur la régulation des gènes ?
Exercice n°4 : Opéron arabinose, d’après Dunn, TM et coll., PNAS, vol.81, p5017-5020 (1984). La régulation de l’opéron arabinose chez E. coli n’est pas aussi simple que le laisse sous-entendre l’exercice n°3. Il a ensuite été montré que la protéine araC, régulant l’opéron, joue à la fois un rôle de régulateur positif et de régulateur négatif. Ainsi, dans la régulation du groupe de gènes araBAD, la liaison d’araC au site 1 en présence d’arabinose (et en absence de glucose) augmente la transcription de l’opéron d’un facteur 100 environ par rapport au taux de base mesuré en absence de protéine araC. La liaison d’araC au site 2, en absence d’arabinose, réprime la transcription des gènes araBAD 10 fois environ en dessous du taux de base, mesuré en l’absence de protéine araC. L’association des effets de régulation négative au site 2 (sans arabinose) et positive au site 1 (sans arabinose) signifie que l’addition d’arabinose provoque une multiplication par 1 000 de la transcription des gènes araBAD. La régulation positive par liaison au site 1 semble directe, étant donné que ce site est adjacent au promoteur et facilite probablement la liaison de l’ARN polymérase ou active la formation d’un complexe ouvert. Le fait que le site 2 est à 270
2

nucléotides en amont du site de transcription a laissé les auteurs de cette publication perplexes. Trois mécanismes potentiels de répression à distance peuvent être imaginés.
1/ Lesquels ?
Afin d’étudier le mécanisme de la répression au site 2, ils ont déplacé la région régulatrice dans son entier devant le gène galK dont le produit, la galactokinase, est plus simple à tester que les enzymes codés par les gènes araBAD.
2/ Comment, dans cette construction, est appelé (ou encore qualifié) le gène galK ?
Afin de déterminer l’importance de la distance entre les sites de liaison d’araC, ils ont inséré ou délété des nucléotides au point d’insertion indiqué Figure A. L’activité du promoteur en absence d’arabinose est alors testée, en faisant pousser les bactéries sur des boîtes indicatrices particulières, dans lesquelles les colonies bactériennes sont blanches si le promoteur est complètement réprimé et rouges si la galactokinase est produite. L’analyse des résultats obtenus est présentée Figure B : les couleurs des bandes correspondant aux bactéries contenant les espacements modifiés sont face à une échelle montrant le nombre de nucléotides insérés ou délétés.
3/ De ces 3 mécanismes, lequel est appuyé par les données expérimentales obtenues, et rend compte de l’organisation observée des bandes blanches et rouges ?
Fig. A : Disposition des sites de liaison d’araC dans l’opéron araBAD
Fig. B : Résultats de la modification de la distance entre les sites de liaison d’araC. Les gènes d’araBAD ont été remplacés par le gène galK, et des nombres variés de nucléotides ont été insérés ou délétés à partir du point d’insertion « 0 ». Les colonies n’exprimant pas la galactokinase sont blanches, celles qui l’expriment sont rouges.
Exercice n°5 : Gène de la Glutamine Synthase (GLN Synthase), d’après Ninfa et al. (1987, Cell 50, 1039-1046) La transcription du gène bactérien codant la glutamine synthase est régulée par la disponibilité de l'azote dans la cellule. Le régulateur-clé de cette transcription est la protéine NtrC, qui n'active la transcription que quand elle est phosphorylée. La phosphorylation de la protéine NtrC est contrôlée par la protéine NtrB, à la fois protéine kinase et phosphatase. L'équilibre entre ses activités kinasique et phosphatasique, et de là le degré de phosphorylation de NtrC et la transcription de la glutamine synthase, est déterminé par d'autres protéines répondant au rapport α-cétoglutarate / glutamine. Ce rapport est un indicateur sensible de la disponibilité en azote, car 2 azotes, sous forme d'ammonium, doivent être ajoutés à l' α-cétoglutarate pour donner de la glutamine. La transcription du gène de la glutamine synthase peut être réalisée in vitro en ajoutant à une matrice d'ADN linéaire contenant le gène et sa région régulatrice en amont du site de liaison de l'ARN polymérase, un facteur sigma particulier, et la protéine NtrC phosphorylée. Les tests de "footprinting" (empreintes sur gel) de l'ADN montrent que la protéine NtrC peut se lier à 5 sites en amont du promoteur. Bien que la liaison de la protéine NtrC ne soit que légèrement augmentée par la phosphorylation, la transcription dépend absolument de la phosphorylation. Néanmoins, l'ARN polymérase se lie facilement au promoteur, même en absence de protéine NtrC.
1/ Si l'ARN polymérase peut se lier au promoteur du gène de la glutamine synthase en absence de protéine NtrC, pourquoi cette protéine NtrC est-elle nécessaire pour activer la trancription?
L'activation de la transcription par NtrC a ensuite été étudiée en utilisant 3 matrices différentes : A- le gène normal et ses séquences de régulations intactes B- le gène après délétion des sites de liaison de NtrC, C- le gène après déplacement, à son extémité 3', de 3 sites de liaisons de NtrC.
En absence de protéine NtrC phosphorylée, aucune transcription ne se produit quelle que soit la matrice. En présence de NtrC phosphorylée à 100 nM, les 3 matrices subissent une transcription maximale. Cependant, les 3 matrices diffèrent significativement quant à la concentration en protéine NtrC phosphorylée nécessaire pour atteindre la moitié du taux maximum de transcription.
A- le gène normal requiert de la protéine NtrC phosphorylée à 5 nM, B- le gène sans site de liaison, 50 nM,
C- le gène avec les sites en 3', 10 nM.
2/ Si la protéine NtrC peut se lier à ses sites de liaison quel que soit son état de phosphorylation, pourquoi (comment est-il pos sible) la phosphorylation est-elle nécessaire à la transcription? 3/ Si la protéine NtrC phosphorylée peut activer la transcription même quand ses sites de liaison sont absents, quel rôle jouent ces sites de liaison?
3

Trois matrices pour l'étude du rôle des sites de liaison de ntrC dans la transcription du gène de la glutamine synthase.
Exercice n°6 : Opéron Histidine, d’après Delorme et al. (1999, J. Bact. 181, 2026-2037) et partie du sujet d’examen 2ème session 2000-2001 de Mr. Kahlem : Régulation par atténuation (cf Opéron TRP) L'opéron histidine de Lactococcus lactis contient tous les gènes nécessaires à la biosynthèse de l'histidine. Le premier cistron, hisC, est situé à environ 300 pb de l'origine de transcription. Cette séquence, appelée région L (leader), comprend :
1) 6 séquences répétées inversées (nommées de A à F) qui permettent la formation de différentes structures en tige-boucle de l'ARN,
2) un terminateur transcriptionnel potentiel rho-dépendant, 3) une « boîte T », entre D et E, connue pour son affinité pour les ARNt non chargés ainsi qu'une séquence CAC, entre B
et C, importante pour la régulation. Aucun cadre de lecture ouvert n'est identifié dans cette région. Afin d'étudier le mécanisme moléculaire de cette régulation, C. Delorme et coll. ont analysé les transcrits produits par les bactéries placées en présence ou non d'histidine. Après extraction et électrophorèse puis transfert sur membrane, les ARN sont hybridés avec une sonde radioactive correspondant à la région L. L'autoradiographie obtenue est présentée sur la figure 1. D’autres études ont montré que les deux transcrits possèdent la même origine et que leur proportion varie en fonction de la quantité d'histidine disponible.
Figure n°1 : Transcription analysis of the histidine operon by Northern blot. Total RNA was extracted from L. lactis NCDO2118 cells grown with (+) and without (-) histidine and hybridized with oligonucleotide 963R. The size of the band is indicated. No transcript was detected in cells grown with histidine.
1) Représentez schématiquement l'opéron his et les transcrits obtenus dans les différentes conditions de culture. Indiquez sur votre schéma les régions essentielles pour la transcription et la traduction et précisez leur fonction respective.
2) Proposez une méthode permettant d'analyser l'origine des deux transcrits. 3) Proposez, sous forme de 2 schémas, un modèle moléculaire de la régulation quantitative de cet opéron
sachant que l'anticodon de l'ARNt-His est GUG. Exercice n°7 : Réponse SOS, d’après Little et al. (1979, PNAS, 76, 6147-6151) Le régulon SOS est réprimé par le produit du gène lexA, un répresseur protéique. A la différence du répresseur lac, les propriétés de liaison de la protéine LexA à l’ADN ne sont pas régulées par une petite molécule effectrice. Une autre différence importante par rapport à l’opéron lactose est à noter : la répression de l’opéron SOS ne peut être ré-établie que dans des bactéries en phase active de synthèse protéique. L’inactivation de la protéine LexA se fait grâce au produit du gène recA (RecA). Le gène recA peut être réprimé par LexA. Little et collaborateurs ont voulu préciser le mécanisme par lequel le produit du gène recA inactive LexA en incubant in vitro LexA ou LexA3 (un mutant protéique de LexA) avec RecA, et ont ensuite analysé ces mélanges par électrophorèse sur gel. Les résultats sont présentés figure suivante.
4

Résultats du Western-blot Lex.
1- Quel est probablement le mécanisme par lequel RecA inactive LexA ? Comment ce mécanisme peut
expliquer que le chloramphénicol, un inhibiteur de synthèse des protéines bactériennes, bloque la re-répression des gènes SOS ?
2- La mutation lexA3 vous paraît-elle récessive ou dominante ? Pourquoi ? 3- Proposez un mécanisme par lequel la répression SOS peut être rétablie dans les bactéries en phase
active de synthèse de RecA.
Exercice n°8 : ermC, d’après Bechhofer et al. (1987, PNAS 84, 498-502) Le gène ermC code pour une méthyl-transférase, enzyme qui réalise la modification d’un ARN ribosomal. Cette réaction rend les ribosomes insensibles à certains antibiotiques de la famille MLS (macrolides, lincosamide, streptogramine B). La régulation de l’expression du gène ermC a fait l’objet de nombreuses études : la traduction de l’ARNm d’ermC est dépendante de la présence d’une faible quantité d’érythromycine (antibiotique appartenant à la famille MLS). La figure n°1 présente le début de la séquence transcrite du gène ermC. Afin de préciser le mécanisme de la régulation de la synthèse de la protéine ErmC, D.H. Bechhofer et D. Dubnau ont réalisé les expériences qui seront analysées dans cet exercice.
FIG. 1 : Uninduced structure of the ermC mRNA regulatory region. The six complementary segments are numbered. The TAA and CCC substitutions are described in the text. Shine-Dalgarno (SD) sequences are marked with hatched bars, followed by the respective ATG initiation codons (thin lines). The stop codon for the leader peptide is indicated by a solid bar.
Les bactéries Bacillus subtilis, contenant le plasmide pE194 renfermant le gène ermC, sont mises en présence d’érythromycine pendant 15 min. Un traitement à la rifampicine est alors réalisé, ce qui permet d’inhiber la transcription. A des temps variables (de 0 à 120 min) après l’addition de rifampicine, les ARN des bactéries sont extraits puis soumis à électrophorèse. Les ARNm ermC sont révélés à l’aide d’une sonde correspondant à un fragment du gène ermC. Une analyse équivalente est réalisée en parallèle à partir de bactéries non traitées à l’érythromycine. L’autoradiogramme obtenu est présenté figure n°2.
FIG. 2 : Blot-hybridization analysis of induced stability of ermC mRNA. Parallel cultures of a pBD144-containing wild-type strain were grown in minimal medium in the absence of Em to the mid-logarithmic stage of growth. Em (0.02 µg/ml) was added to one of the cultures. Induction was allowed to proceed for 15 min, at which time rifampicin was added to 150 µg/ml. One-milliliter samples were removed at various times after addition of rifampicin and were added to 0.4 ml of ice-cold 50 mM NaN3. Time (in minutes) after rifampicin addition is given above each lane.
5
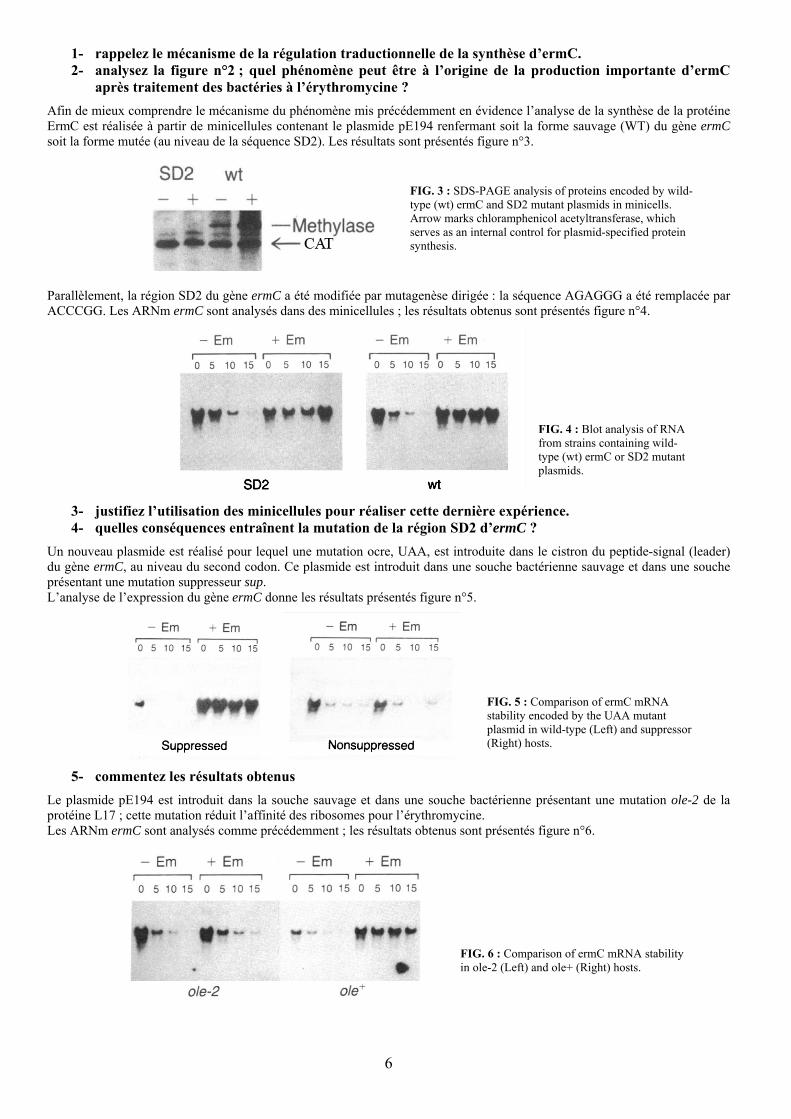
1- rappelez le mécanisme de la régulation traductionnelle de la synthèse d’ermC. 2- analysez la figure n°2 ; quel phénomène peut être à l’origine de la production importante d’ermC
après traitement des bactéries à l’érythromycine ?
Afin de mieux comprendre le mécanisme du phénomène mis précédemment en évidence l’analyse de la synthèse de la protéine ErmC est réalisée à partir de minicellules contenant le plasmide pE194 renfermant soit la forme sauvage (WT) du gène ermC soit la forme mutée (au niveau de la séquence SD2). Les résultats sont présentés figure n°3.
FIG. 3 : SDS-PAGE analysis of proteins encoded by wild-type (wt) ermC and SD2 mutant plasmids in minicells. Arrow marks chloramphenicol acetyltransferase, which serves as an internal control for plasmid-specified protein synthesis.
Parallèlement, la région SD2 du gène ermC a été modifiée par mutagenèse dirigée : la séquence AGAGGG a été remplacée par ACCCGG. Les ARNm ermC sont analysés dans des minicellules ; les résultats obtenus sont présentés figure n°4.
FIG. 4 : Blot analysis of RNA from strains containing wild-type (wt) ermC or SD2 mutant plasmids.
3- justifiez l’utilisation des minicellules pour réaliser cette dernière expérience. 4- quelles conséquences entraînent la mutation de la région SD2 d’ermC ?
Un nouveau plasmide est réalisé pour lequel une mutation ocre, UAA, est introduite dans le cistron du peptide-signal (leader) du gène ermC, au niveau du second codon. Ce plasmide est introduit dans une souche bactérienne sauvage et dans une souche présentant une mutation suppresseur sup. L’analyse de l’expression du gène ermC donne les résultats présentés figure n°5.
FIG. 5 : Comparison of ermC mRNA stability encoded by the UAA mutant plasmid in wild-type (Left) and suppressor (Right) hosts.
5- commentez les résultats obtenus
Le plasmide pE194 est introduit dans la souche sauvage et dans une souche bactérienne présentant une mutation ole-2 de la protéine L17 ; cette mutation réduit l’affinité des ribosomes pour l’érythromycine. Les ARNm ermC sont analysés comme précédemment ; les résultats obtenus sont présentés figure n°6.
FIG. 6 : Comparison of ermC mRNA stability in ole-2 (Left) and ole+ (Right) hosts.
6

6- connaissant l’incidence de l’érythromycine sur la fonction des ribosomes, proposez un mécanisme de régulation de la traduction d’ermC lorsque les bactéries sont mises en présence de l’antibiotique et que les ribosomes ont été modifiés.
Exercice n°9 : protéine hsp, d’après Versteeg et al. (2003, J. Bact. 185, 466-474) et partie du sujet d’examen 1ère session 2002-2003 de Mr. Kahlem Tous les organismes réagissent à une élévation anormale de température par la synthèse de protéines de choc thermique ou HSP. Chez Escherichia coli, le gène htpG appartient au régulon HSP. Chez Bacillus subtilis, on constate que la synthèse de HtpG est augmentée 10 fois lors d'un choc thermique mais sa fonction reste mal comprise. S Versteeg et coll. ont analysé la régulation de ce gène. L'analyse du promoteur (figure n°l) révèle la présence des séquences classiques en -10 et -35 reconnues par le facteur σA (équivalent du σ70 d’E. coli), et de séquences directes ou inversées : RI, R2, R3 (tête de flèche).
FIG. 1. Localization of the regulatory sequence involved in the heat shock regulation of htpG. DNA sequence of the promoter region of htpG. One direct and two inverted repeats are indicated by arrowheads above the DNA sequence; the stop codon of the divergently transcribed yxcA and the putative start codon of htpG are indicated and given in boldface; the -35 and -10 regions of the potential σA -dependent promoter are indicated, given in boldface, and underlined; the transcriptional start site is underlined; the Shine-Dalgarno sequence of htpG is marked by asterisks above the DNA sequence.
Afin d'analyser la possible intervention de ces séquences dans la régulation, des gènes rapporteurs ont été construits et introduits par recombinaison, en un seul exemplaire, dans le locus amyE de B. subtilis. Ces gènes (figure n°2) comprennent le gène lacZ de la β-galactosidase sous le contrôle du promoteur de htpG (SS01) ou de promoteurs mutés par délétion totale ou partielle de séquences répétées ou par modifications ponctuelles d'une de ces séquences (SS02, SV02, SV03, SV09). La séquence GAAAAGG est déplacée, à proximité immédiate de la séquence promotrice -10 dans la construction SVI6. Cette même séquence est également introduite dans un gène rapporteur comportant le promoteur d'un gène qui n'appartient pas aux régulons HSP : le gène lepA, (SV27 – SV13).
FIG. 2. (A) Schematic representation of different promoter regions transcriptionally fused to the bgaB reporter gene. The repeats are indicated by arrows; the fusions in SS01, SS02, SV02, SV03, and SV09 contain htpG promoter regions (gray rectangles), and SV27 and SV13 contain the promoter of lepA (white rectangles). In strain SV13, part of the downstream region of htpG is fused to the lepA promoter. Asterisks mark the two point mutations (B) Schematic presentation of htpG-bgaB fusions; the repeats are indicated by arrows; point mutations are written in light gray letters and marked by asterisks.
L'activité β-galactosidasique des extraits bactériens est analysée à partir de cultures menées à 37°C ou ayant subi un choc thermique de 30 min à 50°C (figure n°3).
7

FIG. 3. Relative β-galactosidase activities of the transcriptional fusions presented in Fig.2. The enzymatic activity measured with the unstressed strain SS01 was set as one relative BgaB unit (equivalent to a specific activity of 0.010 ± 0.001 U/mg of protein). The reporter enzyme activities were measured from cultures without temperature shift (light gray columns) and 30 min after a shift from 37 to 50°C (dark gray columns).
Afin de préciser le mécanisme de la régulation, les transcrits de quelques clones ayant ou non subi un choc thermique, sont analysés après électrophorèse, transfert et hybridation des ARN avec une sonde correspondant au gène htpG (figure n°4).
FIG. 4. Northern blot analyses of different fusions as indicated. Total RNA was prepared before (-) and 10 min after (+) a heat shock from 37 to50°C. Each lane contained 2 µg of total RNA. The bgaB transcript was visualized with DIG-labeled antisense bgaB RNA.
1) Rappelez les principales fonctions des HSP. 2) Analysez les résultats obtenus Fig. 3 avec les constructions SS01, SS02, SV02, SV03 (Fig. 2) et tirez-en
une conclusion sur le rôle éventuel des séquences répétées dans la régulation de la synthèse de HtpG. 3) Comparez les résultats obtenus Fig.3 avec les constructions SV03, SV09, SV27, SV13 (Fig. l) et
proposez un ou plusieurs modèles de régulation. 4) Analysez la figure 4 et le résultat de la construction SV16 (Fig. l), et montrez qu'il est en accord avec
un modèle de régulation. 5) Ce modèle reste incomplet. Proposez une hypothèse pour compléter ce modèle et proposez une ou
plusieurs méthodes susceptibles d'atteindre cet objectif. 6) Le mécanisme moléculaire de la perception du signal thermique reste ici inconnue. Expliquez
pourquoi les auteurs ont tenté d'induire (sans succès) la synthèse de HtpG en traitant les bactéries avec de l'éthanol ou de la puromycine.
Sujet d’examen (1 h), session 1, 2004-05 La protéine SecB est un chaperon moléculaire qui participe à l’exportation de protéines au travers des membranes cytoplasmiques chez E. coli. La synthèse de la protéine SecB est contrôlée par la teneur en source carbonée ; ainsi la quantité intracellulaire en SecB est diminuée en présence de glucose ; le complexe CRP-AMPc joue également un rôle dans la synthèse de cette protéine. Seoh & Tai ont étudié plus en détail le mode de régulation de l’expression de SecB (J. Bacteriol. (1999) 181 : 1892-99).
1- rappeler le principe de la régulation exercée par le complexe CRP-AMPc ; présenter un exemple.
Les auteurs ont tout d’abord cherché à identifier le promoteur du gène secB. Pour cela ils ont réalisé différentes fusions transcriptionnelles secB-lacZ ; la mise en évidence d’une activité transcriptionnelle est évaluée par mesure de l’activité β-galactosidase. La réponse du glycérol ou du glucose et l’effet de l’AMPc ont été évalués. Les résultats obtenus sont présentés figure n°1.
8

Figure n°1 : A- schéma de l’organisation de la région du génome d’E. coli contenant le cistron secB. B- recherche du promoteur du gène secB : réalisation de fusions secB-lacZ ; les traits fins représentent les différentes
régions en amont du site EcoRV ; trait épais : gène lacZ sans son promoteur ; détermination de l’activité β-galactosidase sur cellules renfermant les différents plasmides de fusion lorsque la culture est menée en présence de glycérol, ou sur du glucose, ou encore sur du glucose avec apport d’AMPc. L’activité mesurée (unité arbitraire) représente la moyenne de 3 mesures avec une erreur standard de +/- 10%. Le schéma n’est pas à l’ échelle.
2- analyser la figure n°1, en déduire la (ou les) région(s) promotrice(s) du gène secB
Par la suite les auteurs ont localisé les sites d’initiation de la transcription. Afin de préciser le rôle exact joué par le complexe CRP-AMPc dans la régulation de la transcription, des expériences d’empreinte ont été réalisées afin de mesurer les interactions possibles entre le complexe CRP-AMPc et le promoteur. Il apparaît que le motif reconnu par la CRP présente une séquence assez différente des sites reconnus habituellement par cette protéine. Afin de lever toute ambiguïté, des expériences de délétions et des mutations ont été réalisées (figure n°2).
Figure n°2 :
A- Les lettres en gras indiquent les bases qui ont été remplacées. B- Fragments d’ADN utilisés pour les expériences de fixation de la CRP
Des quantités croissantes de CRP sont mélangées avec le fragment choisi (préalablement radio-marqué) ; les complexes formés sont analysés par électrophorèse. Les résultats obtenus avec les différents fragments utilisés sont présentés figure n°3.
Pr-46 Pr-47 Pr-60 - Figure n°3 : analyse de la fixation de la CRP sur différentes constructions Electrophorèse des mélanges réalisés, 3 concentrations différentes en CRP ont été utilisées.
+
0,2 0,8 3,2 0,2 0,8 3,2 0,2 0,8 3,2 [CRP] (nM) 3- analysez la figure n°3 ; à quoi correspondent les différentes bandes observées ? quel résultat attend-on avec le fragment Pr-102 ? (faire un schéma)
Les mutations effectuées sur 4 bases du site reconnu par la CRP (cf. figure n°3A) abolissent complètement la fixation de la protéine CRP.
4- schématiser l’autoradiogramme attendu (y placer les « bons » témoins)
9

La dernière série d’expériences menées correspond à l’étude in vivo de l’effet de mutations sur le site reconnu par la CRP sur l’expression de SecB par l’utilisation de la fusion secB-lacZ sur la souche sauvage (ZK4) et sur une souche mutante crp-déficiente (RH77). Les principaux résultats obtenus sont présentés figure n°4.
Figure n°4 : effet de la fixation de la CRP sur l’activité β-gal Etude in vivo sur une souche sauvage (ZK4) et une souche mutante (RH77) crp-déficiente
5- analyser la figure n°4 6- conclure sous la forme d’un schéma récapitulatif de la régulation
Sujet d’examen (1 h), session 2, 2004-05 La régulation de l’expression de l’opéron pyr de Bacillus subtilis implique la protéine PyrR, qui agit associée à des pyrimidines. Cette protéine est codée par le premier gène de cet opéron, et elle agit au niveau de trois séquences atténuatrices situées, pour la première, au sein d’une séquence leader, pour la seconde, dans la région intercistronique entre le 1er et le 2ème gène et pour la troisième, dans la région intercistronique entre le 2ème et le 3ème gène de l’opéron (voir figure n° 1 A). Chacun des transcrits obtenus à partir des 3 atténuateurs peuvent se structurer et constituer selon les cas des terminateurs ou des antiterminateurs de la transcription. Diverses expériences antérieures ont montré que la délétion du gène pyrR entraîne l’expression constitutive et à fort taux des gènes de l’opéron pyr.
1- rappeler (à l’aide d’un schéma) le principe de la régulation par atténuation transcriptionnelle ; donner un exemple (8 pts)
Un modèle de régulation a été proposé par Lu & Switzer (1996 J. Bacteriol. 178 : 7206) sur la base des résultats obtenus après reconstitution du phénomène d’atténuation in vitro. Ce système comprend l’ARN polymérase, un ADN matrice (une région de l’opéron pyr), des NTP (dont de l’ATP radiomarqué), la protéine PyrR et de l’UMP. Une analyse de la longueur des transcrits obtenus a été effectuée. Pour cela différents ADN matrices (présentés figure n°1A) ont été utilisés ; les transcrits obtenus sont analysés sur gel et leurs tailles déterminées (figure n°2).
10
(A) pyrR pyrP pyrB’
3 1 2
(B) Matrice 1 2 3
Taille de l’ADN matrice (pb)
591 560 622
Taille attendue pour le transcrit 1 (nt)
450 324 387
Taille attendue pour le transcrit 2 (nt)
124 154 167
Figure n°1 : A- matrices utilisées pour les expériences de transcription in vitro. Le haut du schéma représente la région 5’ de l’opéron pyr avec 3 de ses gènes ; les régions non traduites sont en trait fin ; les ORF se présentent sous la forme de rectangle ; le bas du schéma présente les 3 segments d’ADN utilisés comme matrice ; la flèche représente le point de départ de la transcription ; dans le cas des matrices 2 & 3 en amont de la région à analyser se trouve un promoteur. B- Tableau récapitulatif des tailles des ADN matrices utilisés et des transcrits 1 & et 2 obtenus. Le schéma n’est pas à l’échelle.
2- à quoi correspondent les 2 types de produits de transcription obtenus à partir de chaque matrice? (3
pts)

Figure n°2 : effets de la protéine PyrR et l’UMP sur la transcription in vitro ; analyse électrophorétique des transcrits obtenus. (-) indique absence du composé (+) indique présence du composé
3- quel ADN matrice a été utilisé dans l’expérience de transcription in vitro présenté figure n°2 ? quelle est l’influence de la protéine PyrR et l’UMP sur la transcription ? (3 pts)
Cette même expérience de transcription in vitro réalisée avec les deux autres matrices a donné les mêmes résultats. Cependant avec la matrice 3, l’effet provoqué par PyrR et l’UMP est moins net. Les auteurs émettent l’hypothèse que le transcrit le plus court qui se forme doit présenter une structure secondaire particulièrement stable.
4- schématiser l’autoradiogramme attendu dans le cas où la matrice 3 entière (de 622 pb) est utilisée (2 pts)
5- comment peut-on vérifier expérimentalement l’hypothèse formulée par les auteurs ? (2 pts) 6- faire un schéma récapitulatif de la régulation exercée par PyrR. (2 pts)
11

Sujet d’examen 1ère session 2006-2007 (1h) Les gènes requis pour le développement de la bioluminescence (lux) ont été isolés de la bactérie marine Vibrio fischeri et exprimés dans E. coli ; la séquence nucléotidique du régulon lux en entier a été déterminée. Les gènes lux sont organisés en 2 opérons, transcrits de manière divergente à partir d’une région régulatrice commune, séparant ces 2 opérons. L’opéron D, de droite, comprend 6 gènes (luxICDABE) codant entre autre l’enzyme requise pour la synthèse de l’AI (lux I), les sous unités a & b de la luciférase (luxAB). L’opéron G, à gauche, comprend un seul gène, luxR qui code la protéine régulatrice LuxR. Faire un schéma de l’organisation de ce régulon. La bioluminescence chez V. fischeri est régulée par un mécanisme singulier dépendant de la densité cellulaire, appelé auto-induction. Ce mécanisme repose sur la production d’une petite molécule appelée auto-inducteur (AI). Le modèle de l’induction de la bioluminescence est le suivant : à un niveau basal d’expression de l’opéron D, il y a production à faible taux de l’AI par la protéine LuxI ; à forte densité cellulaire, l’AI s’accumule et interagit avec la protéine LuxR pour former un complexe se fixant à l’opérateur lux et stimulant la transcription de l’opéron R. La séquence cible du complexe est palindromique. Certaines mutations réalisées dans la séquence palindromique entraînent un blocage de l’activation de la transcription par le complexe LuxR-AI. A l’aide d’un schéma, proposer un modèle pour la fixation du complexe LuxR-AI sur sa séquence cible ; quel peut être l’effet de l’AI sur la structure de LuxR? Des souches d’E. coli déficientes soit dans leur capacité à synthétiser l’AMPc, soit dans leur capacité à synthétiser la protéine CAP ne sont plus capables de réguler l’opéron G. Rappeler le principe de ce mode de régulation. De plus, la transcription de l’opéron G est régulée négativement par la protéine LuxR selon un mécanisme qui dépend de l’AI. Shadel G.S. & T.O. Baldwin (J. Bacteriol. 1991) ont réalisé une série d’expériences afin de mieux comprendre ce mécanisme de régulation. Un premier plasmide (appelé pGS1) est préparé : il comprend la région régulatrice derrière laquelle sont placés les gènes codant la luciférase. Les cellules qui renferment un tel plasmide ne peuvent produire de l’AI. Un second plasmide (pPD7) est préparé ; il permet de déclencher la production de la protéine LuxR après l’addition d’IPTG. Schématiser l’organisation du plasmide pPD7. L’expression de la luciférase est suivie au cours du temps sur une souche renfermant les 2 plasmides pGS1 et pPD7 ; les effets de l’AI et de l’IPTG sur la transcription sont présentés figure n°1.
12
Figure n°1 : Suivi de la luminescence produite par la souche d’E. coli transformée par les 2 plasmides pGS1 et pPD7 au cours de la croissance ; l’IPTG et l’AI sont ajoutés au moment indiqué par la flèche.
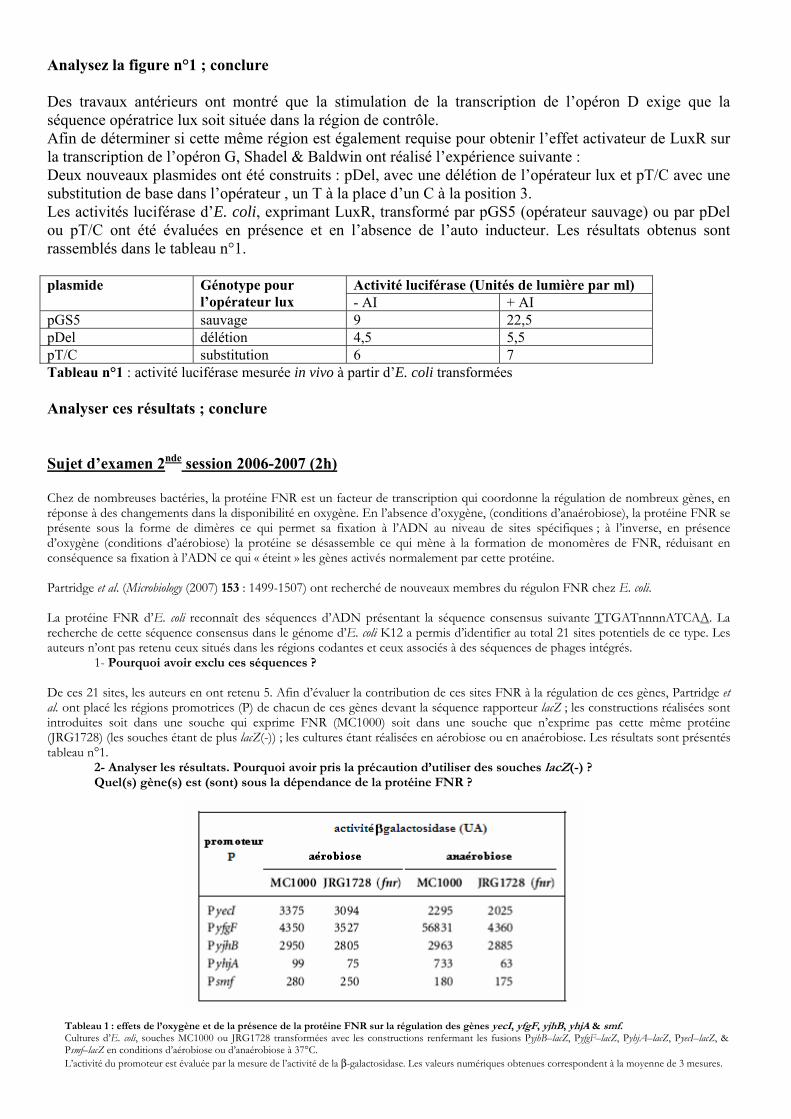
Analysez la figure n°1 ; conclure Des travaux antérieurs ont montré que la stimulation de la transcription de l’opéron D exige que la séquence opératrice lux soit située dans la région de contrôle. Afin de déterminer si cette même région est également requise pour obtenir l’effet activateur de LuxR sur la transcription de l’opéron G, Shadel & Baldwin ont réalisé l’expérience suivante : Deux nouveaux plasmides ont été construits : pDel, avec une délétion de l’opérateur lux et pT/C avec une substitution de base dans l’opérateur , un T à la place d’un C à la position 3. Les activités luciférase d’E. coli, exprimant LuxR, transformé par pGS5 (opérateur sauvage) ou par pDel ou pT/C ont été évaluées en présence et en l’absence de l’auto inducteur. Les résultats obtenus sont rassemblés dans le tableau n°1.
Activité luciférase (Unités de lumière par ml) plasmide Génotype pour l’opérateur lux - AI + AI
pGS5 sauvage 9 22,5 pDel délétion 4,5 5,5 pT/C substitution 6 7 Tableau n°1 : activité luciférase mesurée in vivo à partir d’E. coli transformées Analyser ces résultats ; conclure Sujet d’examen 2nde session 2006-2007 (2h)
Chez de nombreuses bactéries, la protéine FNR est un facteur de transcription qui coordonne la régulation de nombreux gènes, en réponse à des changements dans la disponibilité en oxygène. En l’absence d’oxygène, (conditions d’anaérobiose), la protéine FNR se présente sous la forme de dimères ce qui permet sa fixation à l’ADN au niveau de sites spécifiques ; à l’inverse, en présence d’oxygène (conditions d’aérobiose) la protéine se désassemble ce qui mène à la formation de monomères de FNR, réduisant en conséquence sa fixation à l’ADN ce qui « éteint » les gènes activés normalement par cette protéine. Partridge et al. (Microbiology (2007) 153 : 1499-1507) ont recherché de nouveaux membres du régulon FNR chez E. coli. La protéine FNR d’E. coli reconnaît des séquences d’ADN présentant la séquence consensus suivante TTGATnnnnATCAA. La recherche de cette séquence consensus dans le génome d’E. coli K12 a permis d’identifier au total 21 sites potentiels de ce type. Les auteurs n’ont pas retenu ceux situés dans les régions codantes et ceux associés à des séquences de phages intégrés. 1- Pourquoi avoir exclu ces séquences ?
De ces 21 sites, les auteurs en ont retenu 5. Afin d’évaluer la contribution de ces sites FNR à la régulation de ces gènes, Partridge et al. ont placé les régions promotrices (P) de chacun de ces gènes devant la séquence rapporteur lacZ ; les constructions réalisées sont introduites soit dans une souche qui exprime FNR (MC1000) soit dans une souche que n’exprime pas cette même protéine (JRG1728) (les souches étant de plus lacZ(-)) ; les cultures étant réalisées en aérobiose ou en anaérobiose. Les résultats sont présentés tableau n°1.
2- Analyser les résultats. Pourquoi avoir pris la précaution d’utiliser des souches lacZ(-) ? Quel(s) gène(s) est (sont) sous la dépendance de la protéine FNR ?
13
Tableau 1 : effets de l’oxygène et de la présence de la protéine FNR sur la régulation des gènes yecI, yfgF, yjhB, yhjA & smf. Cultures d’E. coli, souches MC1000 ou JRG1728 transformées avec les constructions renfermant les fusions PyjhB–lacZ, PyfgF–lacZ, PyhjA–lacZ, PyecI–lacZ, & Psmf–lacZ en conditions d’aérobiose ou d’anaérobiose à 37°C. L’activité du promoteur est évaluée par la mesure de l’activité de la β-galactosidase. Les valeurs numériques obtenues correspondent à la moyenne de 3 mesures.
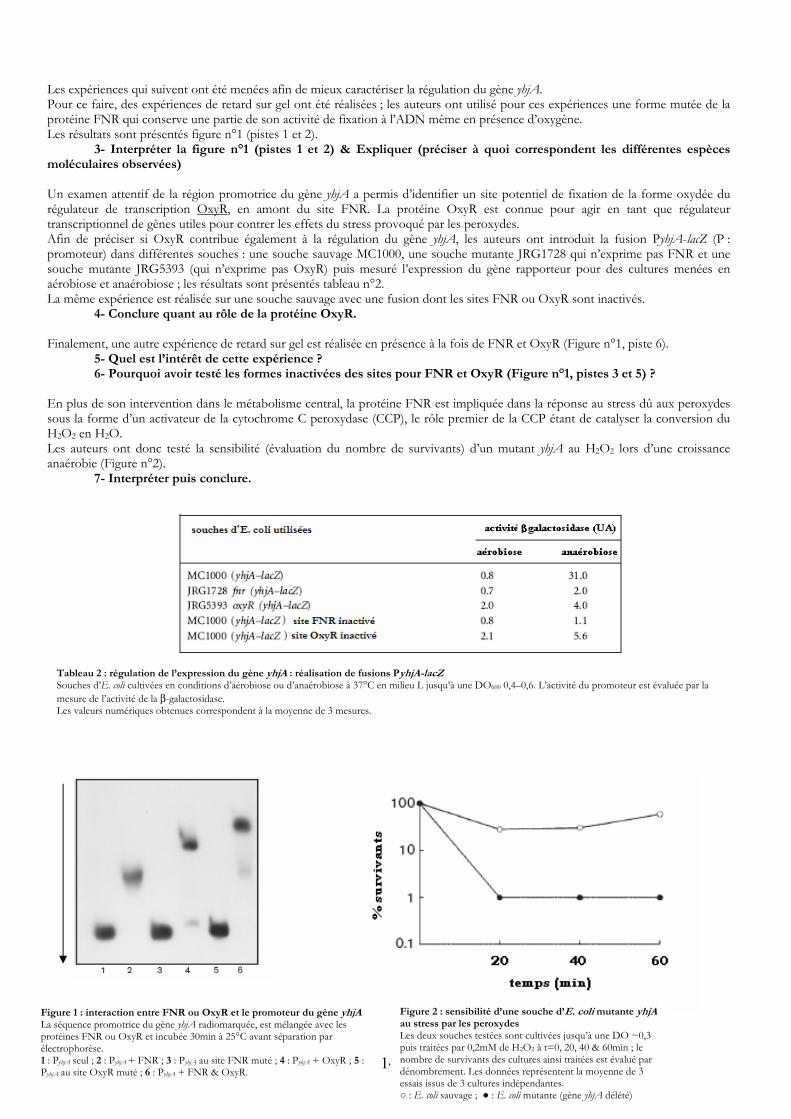
Les expériences qui suivent ont été menées afin de mieux caractériser la régulation du gène yhjA. Pour ce faire, des expériences de retard sur gel ont été réalisées ; les auteurs ont utilisé pour ces expériences une forme mutée de la protéine FNR qui conserve une partie de son activité de fixation à l’ADN même en présence d’oxygène. Les résultats sont présentés figure n°1 (pistes 1 et 2).
3- Interpréter la figure n°1 (pistes 1 et 2) & Expliquer (préciser à quoi correspondent les différentes espèces moléculaires observées)
Un examen attentif de la région promotrice du gène yhjA a permis d’identifier un site potentiel de fixation de la forme oxydée du régulateur de transcription OxyR, en amont du site FNR. La protéine OxyR est connue pour agir en tant que régulateur transcriptionnel de gènes utiles pour contrer les effets du stress provoqué par les peroxydes. Afin de préciser si OxyR contribue également à la régulation du gène yhjA, les auteurs ont introduit la fusion PyhjA-lacZ (P : promoteur) dans différentes souches : une souche sauvage MC1000, une souche mutante JRG1728 qui n’exprime pas FNR et une souche mutante JRG5393 (qui n’exprime pas OxyR) puis mesuré l’expression du gène rapporteur pour des cultures menées en aérobiose et anaérobiose ; les résultats sont présentés tableau n°2. La même expérience est réalisée sur une souche sauvage avec une fusion dont les sites FNR ou OxyR sont inactivés.
4- Conclure quant au rôle de la protéine OxyR.
Finalement, une autre expérience de retard sur gel est réalisée en présence à la fois de FNR et OxyR (Figure n°1, piste 6). 5- Quel est l’intérêt de cette expérience ?
6- Pourquoi avoir testé les formes inactivées des sites pour FNR et OxyR (Figure n°1, pistes 3 et 5) ? En plus de son intervention dans le métabolisme central, la protéine FNR est impliquée dans la réponse au stress dû aux peroxydes sous la forme d’un activateur de la cytochrome C peroxydase (CCP), le rôle premier de la CCP étant de catalyser la conversion du H2O2 en H2O. Les auteurs ont donc testé la sensibilité (évaluation du nombre de survivants) d’un mutant yhjA au H2O2 lors d’une croissance anaérobie (Figure n°2). 7- Interpréter puis conclure.
FiLapréle1 :Py
Tableau 2 : régulation de l’expression du gène yhjA : réalisation de fusions PyhjA lacZ -Souches d’E. coli cultivées en conditions d’aérobiose ou d’anaérobiose à 37°C en milieu L jusqu’à une DO600 0,4–0,6. L’activité du promoteur est évaluée par la mesure de l’activité de la β-galactosidase. Les valeurs numériques obtenues correspondent à la moyenne de 3 mesures.
gure 1 : interaction entre séquence promotrice du gotéines FNR ou OxyR et inctrophorèse. PyhjA seul ; 2 : PyhjA + FNR hjA au site OxyR muté ; 6 : P
FNR ou OxyR et le promoteur du gène yène yhjA radiomarquée, est mélangée avec lescubée 30min à 25°C avant séparation par
; 3 : PyhjA au site FNR muté ; 4 : PyhjA + OxyRyhjA + FNR & OxyR.
14
Figure 2 : sensibilité d’une souche d’E. coli mutante yhjA au stress par les peroxydes Les deux souches testées sont cultivées jusqu’à une DO ~0,3 puis traitées par 0,2mM de H2O2 à t=0, 20, 40 & 60min ; le nombre de survivants des cultures ainsi traitées est évalué par dénombrement. Les données représentent la moyenne de 3 essais issus de 3 cultures indépendantes. ○ : E. coli sauvage ; ● : E. coli mutante (gène yhjA délété)
hjA
; 5 :





























