Embed Size (px)
Citation preview

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 227, No. 2, December, pp. 416-424, 1983
Regulation of C4 Photosynthesis: Regulation of Activation and Inactivation
of NADP-Malate Dehydrogenase by NADP and NADPH
ANTHONY R. ASHTON AND M. D. HATCH’
LXvkian of Plant Industry CSIRO, P.O. Box 1600, Canberra City, A.C.T. 2601, Australia
Received April 25, 1983, and in revised form July 27, 1983
Inactive NADP-malate dehydrogenase (disulfide form) from chloroplasts of Zea mays is activated by reduced thioredoxin while the active enzyme (dithiol form) is inactivated by incubation with oxidized thioredoxin. This reductive activation of NADP-malate dehydrogenase is inhibited by over 95% in the presence of NADP and the Kd for this interaction of NADP with the inactive enzyme is about 3 pM. Other substrates of the enzyme (malate, oxaloacetate, or NADPH) do not effect the rate of enzyme activation but NADPH can reverse the inhibitory effect of NADP. It appears that NADPH (Kd = 250 PM) and NADP (Kd = 3 PM) compete for the same site, presumably the coenzyme- binding site at the active centre. Apparently the enzyme * NADP binary complex cannot be reduced by thioredoxin whereas the enzyme. NADPH complex is reduced at the same rate as is the free enzyme. Similarly the oxidative inactivation of reduced NADP- malate dehydrogenase is inhibited by up to 85% by NADP and NADPH completely reverses this inhibition. The Kd values of the active-reduced enzyme for NADP and NADPH were both estimated to be 30 j&M. From these data a model was constructed which predicts how changing NADPH/NADP levels in the chloroplast might change the steady-state level of NADP-malate dehydrogenase activity. The model indicates that at any fixed ratio of reduced to oxidized thioredoxin high proportions of active NADP-malate dehydrogenase and, hence, high rates of oxaloacetate reduction, can only occur with very high NADPH/NADP ratios.
In Zea rrutys and related C4 plants NADP-malate dehydrogenase catalyzes the reduction of oxaloacetate, the primary product of COz assimilation, to malate. In common with several enzymes of the pho- tosynthetic carbon-reduction cycle NADP- malate dehydrogenase is regulated in leaves by light-mediated activation and dark-mediated inactivation. These pro- cesses are believed to involve reduction of an enzyme disulfide in the light via thio- redoxin (1) or other factors (2,3) linked to photosynthetic electron transport and ox- idation of a dithiol via a thioredoxin-linked reaction in the dark (4,5). Operating alone,
this thioredoxin-dependent system would result in a coordinated regulation of pho- tosynthetic enzymes and, as a consequence, would limit the possibility of independent regulation of individual enzymes. There is now evidence for several of these enzymes that additional control of activity may be achieved by the specific binding of ligands modifying the rates of activation or in- activation. For instance substrates in- crease the rate of disulfide-bond reduction of fructose-1,6-bisphosphatase (6) and se- doheptulose-1,7-bisphosphatase (7) while ATP and ADP inhibit both reduction and oxidation of ribulose-5-phosphate kin- ase (8).
The present paper reports that the rate 1 To whom correspondence should be addressed. of activation and inactivation of purified
0003-9861/83 $3.00 Copyright 0 1983 by Academic Press. Inc. All rights of reproduction in any form reserved.
416
REGULATION OF NADP-MALATE DEHYDROGENASE 417
NADP-malate dehydrogenase from 2. mags is modified directly by NADP and indirectly by NADPH. The results provide a basis for quantitative models predicting how changes in the NADP and NADPH levels in chloroplasts may alter the steady- state levels of active enzyme under con- ditions where the level of reduced thio- redoxin remains unchanged.
MATERIALS AND METHODS
Materials. NADP-malate dehydrogenase and thio- redoxin were purified from 2. may.9 leaves as described previously (9). Biochemicals were obtained from Sigma Chemical Company.
Enzyme assays. The routine assay mixture for NADP-malate dehydrogenase (L-malate:NADP oxi- doreductase, EC 1.1.1.82) contained 2.5 mM oxaloac- etate, 0.2 mM NADPH, 25 mM Tricine*-KOH, 1 mM EDTA, adjusted to pH 8.3. The carryover of products and substrates from activation- or inactivation-re- action mixtures into the enzyme assay did not sig- nificantly alter enzyme activity.
Kinetics of activation and inactivation of NADP- malatedehydmgaase. All reaction mixtures contained 50 mM Tris-HCl, 10 mM KCI, 1 mM EDTA, 2 mM
MgClz, adjusted to pH 7.4 (referred to below as Tris- KC1 buffer) and were run at 25“C under Nz. Reactions were started by the addition of NADP-malate de- hydrogenase. At intervals 254 aliquots were removed and the reaction was stopped by dilution into enzyme- assay mixture. For activation the reaction also con- tained inactive enzyme and 100 mM dithiothreitol or 5 mM dithiothreitol plus thioredoxin. For inactivation the reaction also contained thioredoxin, 1 mrd oxidized glutathione and 25 ~1 of active NADP-malate dehy- drogenase (previously reduced by incubation overnight under Nr with 1 mM dithiothreitol and thioredoxin, see Ref. (9)). Malate dehydrogenase was added to give a concentration of between 1 and 2 units ml-’ reaction. Sufficient thioredoxin was added to give a tllz for ac- tivation of about 10 min and a tL/z for inactivation of about 2.5 min. Both activation and inactivation were largely dependent (more than 90% of the rate) on added thioredoxin.
Preparation of chloroplast extracts. Chloroplasts were prepared from 20 g of 2. muys leaves (predark- ened for 90 min) by slicing into approximately l-mm sections and then blending for 5 s in 0.4 M sorbitol, 20 mM Tris-HCl, pH 8.0,5 mM MgClz, and 5 mM sodium isoascorbate. The pellet obtained by centrifuging at 500~ for 3 min was washed by resuspending in the
* Abbreviation used: Tricine, N-[2-hydroxy-l,l- bis(hydroxymethyl)ethyl)glycine.
same medium without isoascorbate and recentrifug- ing. The chloroplasts were resuspended in 4 ml Tris- KC1 buffer (see above), frozen at -80°C and then thawed and centrifuged at 27,OOOg for 10 min. The supernatant solution was desalted on a Sephadex G- 25 column equilibrated with Tris-KC1 buffer. The NADP-malate dehydrogenase in the desalted chlo- roplast extract was activated with 100 mM dithio- threitol in the presence or absence of 1 mM NADP.
RESULTS
Regulation of the Activation of NADP- Malate Dehydrogenaee
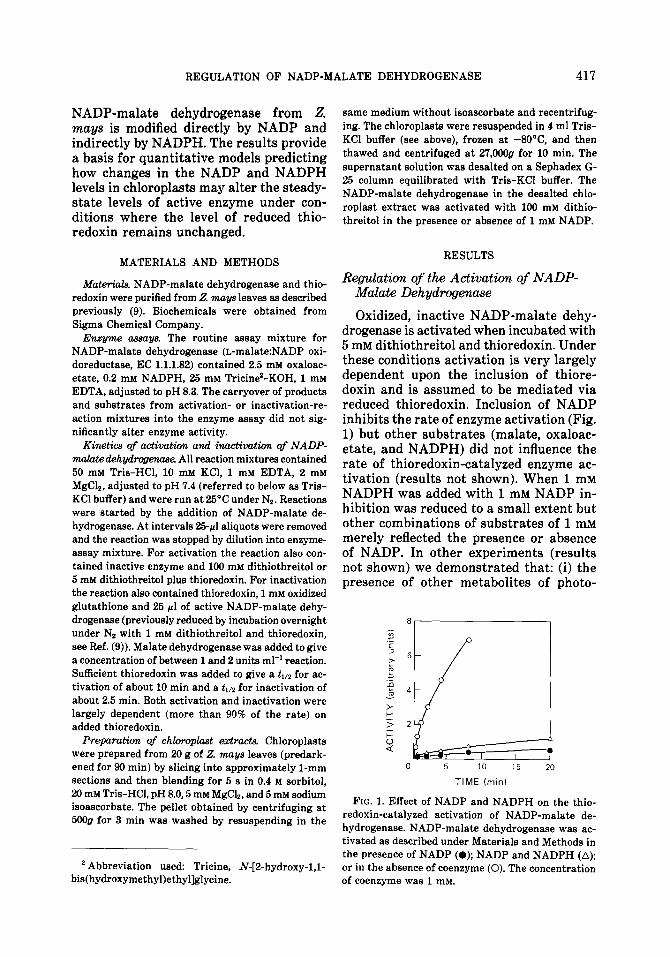
Oxidized, inactive NADP-malate dehy- drogenase is activated when incubated with 5 mM dithiothreitol and thioredoxin. Under these conditions activation is very largely dependent upon the inclusion of thiore- doxin and is assumed to be mediated via reduced thioredoxin. Inclusion of NADP inhibits the rate of enzyme activation (Fig. 1) but other substrates (malate, oxaloac- etate, and NADPH) did not influence the rate of thioredoxin-catalyzed enzyme ac- tivation (results not shown). When 1 mM NADPH was added with 1 mM NADP in- hibition was reduced to a small extent but other combinations of substrates of 1 mM merely reflected the presence or absence of NADP. In other experiments (results not shown) we demonstrated that: (i) the presence of other metabolites of photo-
TIME (min)
FIG. 1. Effect of NADP and NADPH on the thio- redoxin-catalyzed activation of NADP-malate de- hydrogenase. NADP-malate dehydrogenase was ac- tivated as described under Materials and Methods in the presence of NADP (0); NADP and NADPH (A); or in the absence of coenzyme (0). The concentration of coenzyme was 1 mM.
418 ASHTON AND HATCH
synthesis including ATP, ADP, AMP, Pi, 3-phosphoglycerate, pyruvate, and phos- phoenolpyruvate did not significantly in- fluence activation of NADP-malate dehy- drogenase at concentrations up to 1 mM; (ii) activation of NADP-malate dehydro- genase by dithiothreitol alone (no thiore- doxin) is also inhibited by NADP but not by NADPH, malate, or oxaloacetate; and (iii) NAD (1 mM) also inhibits the thio- redoxin-catalyzed activation of NADP- malate dehydrogenase but only about 5% as effectively as NADP. In this connection it is significant that the purified NADP- malate dehydrogenase shows weak activity with NADH as substrate. This activity with NADH is enhanced by reduction of the enzyme suggesting that it is an in- trinsic property of the NADP-malate de- hydrogenase.
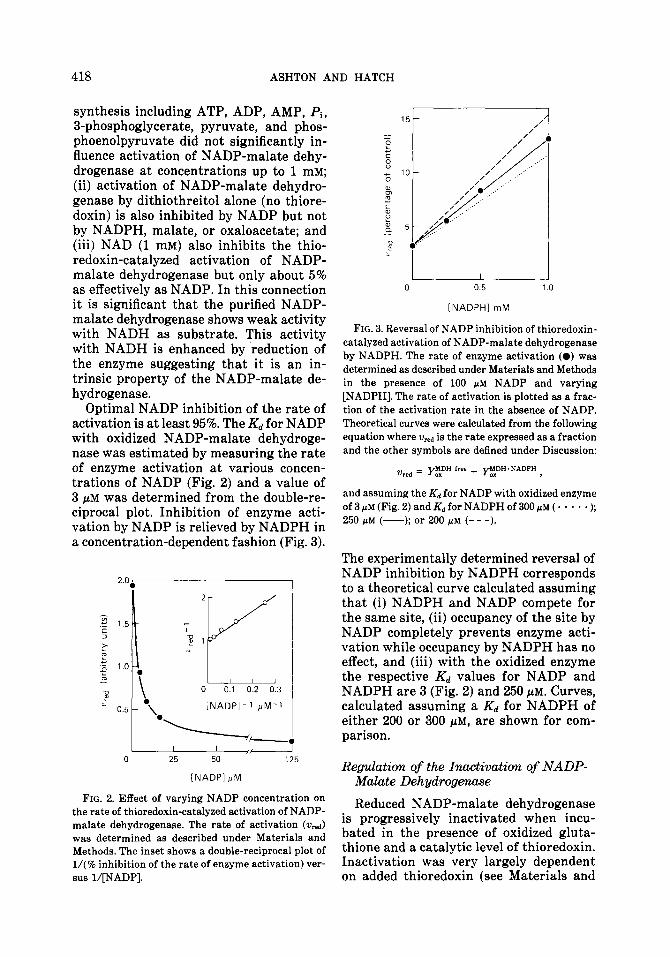
Optimal NADP inhibition of the rate of activation is at least 95%. The Kd for NADP with oxidized NADP-malate dehydroge- nase was estimated by measuring the rate of enzyme activation at various concen- trations of NADP (Fig. 2) and a value of 3 PM was determined from the double-re- ciprocal plot. Inhibition of enzyme acti- vation by NADP is relieved by NADPH in a concentration-dependent fashion (Fig. 3).
0 WA 125
INADPI us
FIG. 2. Effect of varying NADP concentration on the rate of thioredoxin-catalyzed activation of NADP- malate dehydrogenase. The rate of activation (w,J was determined as described under Materials and Methods. The inset shows a double-reciprocal plot of l/(70 inhibition of the rate of enzyme activation) ver- sus l/[NADP].
15- /’
= /’ P /’
g /
/
o lo- / ..’
z ,’ ,..’
,’ :’
P /’ // l ,..:‘.
t; z /
,,’ .:.”
k 5 ,,;.... ”
; ”
la -7
,/. ”
P l ..:”
I; I I
0 0.5 1 .o
[NADPHI mM
FIG. 3. Reversal of NADP inhibition of thioredoxin- catalyzed activation of NADP-malate dehydrogenase by NADPH. The rate of enzyme activation (0) was determined as described under Materials and Methods in the presence of 100 PM NADP and varying [NADPH]. The rate of activation is plotted as a frac- tion of the activation rate in the absence of NADP. Theoretical curves were calculated from the following equation where w,,,, is the rate expressed as a fraction and the other symbols are defined under Discussion:
and assuming the Kd for NADP with oxidized enzyme of 3 ELM (Fig. 2) and Kd for NADPH of 300 pM ( * * * * - );
250 pM (-); or 200 PM (- - -).
The experimentally determined reversal of NADP inhibition by NADPH corresponds to a theoretical curve calculated assuming that (i) NADPH and NADP compete for the same site, (ii) occupancy of the site by NADP completely prevents enzyme acti- vation while occupancy by NADPH has no effect, and (iii) with the oxidized enzyme the respective Kd values for NADP and NADPH are 3 (Fig. 2) and 250 pM. Curves, calculated assuming a Kd for NADPH of either 200 or 300 PM, are shown for com- parison.
Regulation of the Inactivation of NADP- Malate Dehydrogenme
Reduced NADP-malate dehydrogenase is progressively inactivated when incu- bated in the presence of oxidized gluta- thione and a catalytic level of thioredoxin. Inactivation was very largely dependent on added thioredoxin (see Materials and
REGULATION OF NADP-MALATE DEHYDROGENASE 419
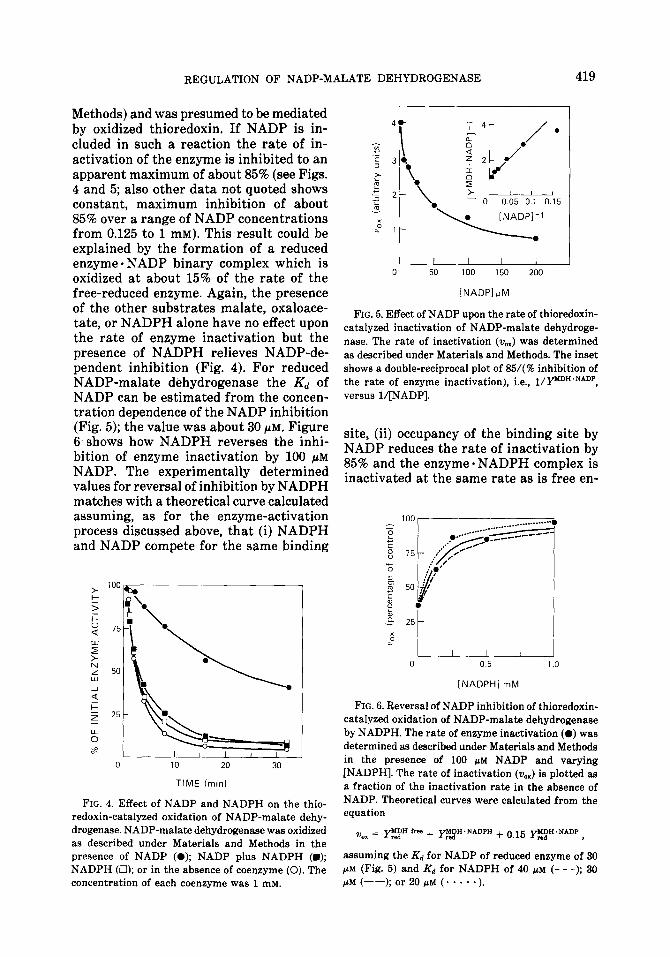
Methods) and was presumed to be mediated by oxidized thioredoxin. If NADP is in- cluded in such a reaction the rate of in- activation of the enzyme is inhibited to an apparent maximum of about 85% (see Figs. 4 and 5; also other data not quoted shows constant, maximum inhibition of about 85% over a range of NADP concentrations from 0.125 to 1 mM). This result could be explained by the formation of a reduced enzyme 0 NADP binary complex which is oxidized at about 15% of the rate of the free-reduced enzyme. Again, the presence of the other substrates malate, oxaloace- tate, or NADPH alone have no effect upon the rate of enzyme inactivation but the presence of NADPH relieves NADP-de- pendent inhibition (Fig. 4). For reduced NADP-malate dehydrogenase the Kd of NADP can be estimated from the concen- tration dependence of the NADP inhibition (Fig. 5); the value was about 30 PM. Figure 6 shows how NADPH reverses the inhi- bition of enzyme inactivation by 100 PM NADP. The experimentally determined values for reversal of inhibition by NADPH matches with a theoretical curve calculated assuming, as for the enzyme-activation process discussed above, that (i) NADPH and NADP compete for the same binding
0 10 20 30
TIME (mln)
FIG. 4. Effect of NADP and NADPH on the thio- redoxin-catalyzed oxidation of NADP-malate dehy- drogenase. NADP-malate dehydrogenase was oxidized as described under Materials and Methods in the presence of NADP (0); NADP plus NADPH (m); NADPH (Cl); or in the absence of coenzyme (0). The concentration of each coenzyme was 1 mM.
r
I I I I 0 50 100 150 200
[NADPI~M
FIG. 5. Effect of NADP upon the rate of thioredoxin- catalyzed inactivation of NADP-malate dehydroge- nase. The rate of inactivation (v,,) was determined as described under Materials and Methods. The inset shows a double-reciprocal plot of 85/(% inhibition of the rate of enzyme inactivation), i.e., l/Ym’H*N*DP, versus l/[NADP].
site, (ii) occupancy of the binding site by NADP reduces the rate of inactivation by 85% and the enzyme l NADPH complex is inactivated at the same rate as is free en-
0 0.5 1 .o
[NADPHI mM
FIG. 6. Reversal of NADP inhibition of thioredoxin- catalyzed oxidation of NADP-malate dehydrogenase by NADPH. The rate of enzyme inactivation (0) was determined as described under Materials and Methods in the presence of 100 PM NADP and varying [NADPH]. The rate of inactivation (v.,) is plotted as a fraction of the inactivation rate in the absence of NADP. Theoretical curves were calculated from the equation
assuming the Kd for NADP of reduced enzyme of 30 @hi (Fig. 5) and Kd for NADPH of 40 PM (- - -); 30 /AM (-); or 20 PM ( * * * * * ).
420 ASHTON AND HATCH
zyme, and (iii) the Kd values of reduced NADP-malate dehydrogenase for both NADP and NADPH are 30 PM. Also shown are curves calculated assuming a Kd for NADPH of either 20 or 40 PM.
Alternutive Fm of NADP-Malate Dehydrogenase It should be noted that with some prep-
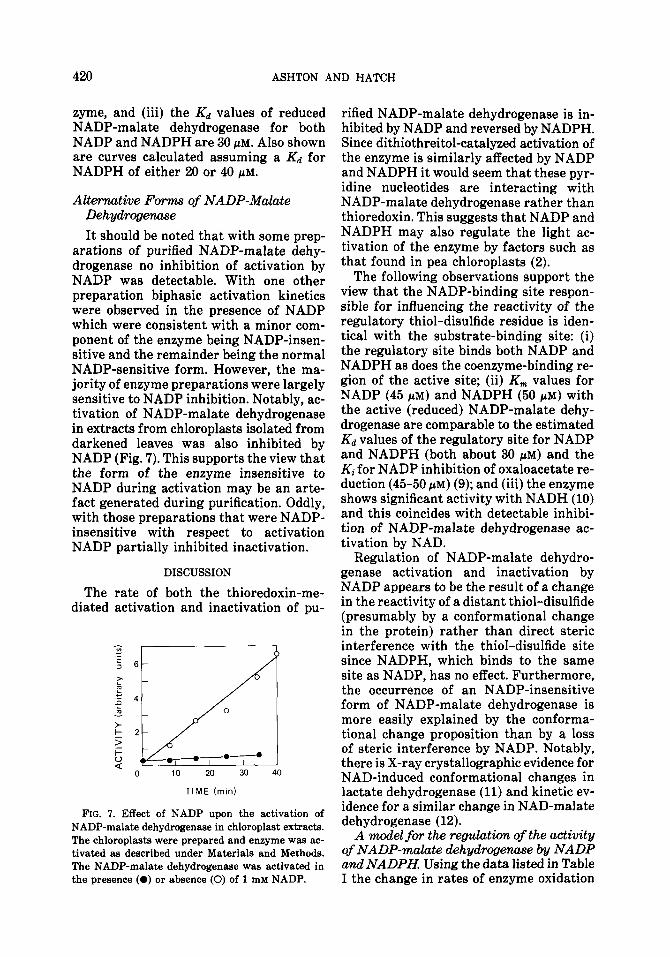
arations of purified NADP-malate dehy- drogenase no inhibition of activation by NADP was detectable. With one other preparation biphasic activation kinetics were observed in the presence of NADP which were consistent with a minor com- ponent of the enzyme being NADP-insen- sitive and the remainder being the normal NADP-sensitive form. However, the ma- jority of enzyme preparations were largely sensitive to NADP inhibition. Notably, ac- tivation of NADP-malate dehydrogenase in extracts from chloroplasts isolated from darkened leaves was also inhibited by NADP (Fig. 7). This supports the view that the form of the enzyme insensitive to NADP during activation may be an arte- fact generated during purification. Oddly, with those preparations that were NADP- insensitive with respect to activation NADP partially inhibited inactivation.
DISCUSSION
The rate of both the thioredoxin-me- diated activation and inactivation of pu-
TIME (min)
FIG. 7. Effect of NADP upon the activation of NADP-malate dehydrogenase in chloroplast extracts. The chloroplasts were prepared and enzyme was ac- tivated as described under Materials and Methods. The NADP-malate dehydrogenase was activated in the presence (0) or absence (0) of 1 mM NADP.
rified NADP-malate dehydrogenase is in- hibited by NADP and reversed by NADPH. Since dithiothreitol-catalyzed activation of the enzyme is similarly affected by NADP and NADPH it would seem that these pyr- idine nucleotides are interacting with NADP-malate dehydrogenase rather than thioredoxin. This suggests that NADP and NADPH may also regulate the light ac- tivation of the enzyme by factors such as that found in pea chloroplasts (2).
The following observations support the view that the NADP-binding site respon- sible for influencing the reactivity of the regulatory thiol-disulfide residue is iden- tical with the substrate-binding site: (i) the regulatory site binds both NADP and NADPH as does the coenzyme-binding re- gion of the active site; (ii) Km values for NADP (45 PM) and NADPH (50 PM) with the active (reduced) NADP-malate dehy- drogenase are comparable to the estimated Kd values of the regulatory site for NADP and NADPH (both about 30 PM) and the Ki for NADP inhibition of oxaloacetate re- duction (45-50 PM) (9); and (iii) the enzyme shows significant activity with NADH (10) and this coincides with detectable inhibi- tion of NADP-malate dehydrogenase ac- tivation by NAD.
Regulation of NADP-malate dehydro- genase activation and inactivation by NADP appears to be the result of a change in the reactivity of a distant thiol-disulfide (presumably by a conformational change in the protein) rather than direct steric interference with the thiol-disulfide site since NADPH, which binds to the same site as NADP, has no effect. Furthermore, the occurrence of an NADP-insensitive form of NADP-malate dehydrogenase is more easily explained by the conforma- tional change proposition than by a loss of steric interference by NADP. Notably, there is X-ray crystallographic evidence for NAD-induced conformational changes in lactate dehydrogenase (11) and kinetic ev- idence for a similar change in NAD-malate dehydrogenase (12).
A model for the regulation of the activity of NADP-malate dehydrogenase & NADP and NADPH. Using the data listed in Table I the change in rates of enzyme oxidation

REGULATION OF NADP-MALATE DEHYDROGENASE 421
TABLE I
COENZYME-BINDING PROPERTIES AND DITHIOL- DISULFIDE REACTIVITY OF INACTIVE AND
ACTIVE NADP-MALATE DEHYDROGENASE”
NADP NADPH
Inactive enzyme & (PM) 3 250 %d 0 1
Active enzyme & (PM) 36 30 210, 0.15 1
a K,+ values were obtained by analysis of the data of Figs. 2, 3, 5, and 6 while the relative rates of re- duction (v,& and oxidation (v,) of the enzyme-coen- zyme binary complex relative to that of the free en- zyme were obtained from Figs. 2 and 5.
and reduction in response to the neces- sarily simultaneous changes in NADP and NADPH levels that would occur in the me- sophyll chloroplast may be calculated as follows: Let Y represent the fraction of ac- tive (or reduced) and inactive (or oxidized) NADP-malate dehydrogenase ( YEiH, YzDH) or thioredoxion (c)z, Et”). Also let the fractional saturation of NADP-ma- late dehydrogenase with NADP or NADPH be represented by YMDHeNADP and YMDHV NADPH respectively, while YMDHfree repre- sents the fraction of enzyme not containing bound coenzyme. Assuming that the total NADP + NADPH content as well as the enzyme and thioredoxin content of the chloroplast is constant, and that the con- centration of [NADP] + [NADPH] is 1 mM (close to the values reported for chloro- plasts, see Refs. (13, 14)), it follows that yAl$]- + BADPHI = 1 mM, c,$?
- 1 and Yf”,fH + YEDH = 1. Also sincz acti& and inactive NADP-malatd dehydrogenase can each bind coenzyme,
YgyH = YE?” free + yzlH.NADP + yglH.NADPH,
and yMDH =
ox yMDH free
ox
+ yKDH.NADP + y,M,DH.NADPH,
The fractional saturation of enzyme NADP ( YMDH’NADP) is given by
by
KyADP - [NADPH] -’ + KFADPH.[NADP] ’ I
Since free enzyme and enzyme * NADPH but not enzyme. NADP can be activated, the overall rate of reduction of NADP-ma- late dehydrogenase in the presence of NADP and NADPH, expressed as a frac- tion of the maximum rate, is given by ~u,,d = y:DH free + yzDH. NADPH
= I _ yMDH.NADP ox t
and, similarly, the relative rate of oxida- tion of NADP malate dehydrogenase is given by
VOX = yE:Hfree + y!lH .NADPH
+ 0.15 Yf”:“‘NADP = 1 - 0 85 yf”,v”+JA=‘P ,
since our data shows that the reduced en- zyme . NADP complex is oxidized at about 15% of the rate of free enzyme.
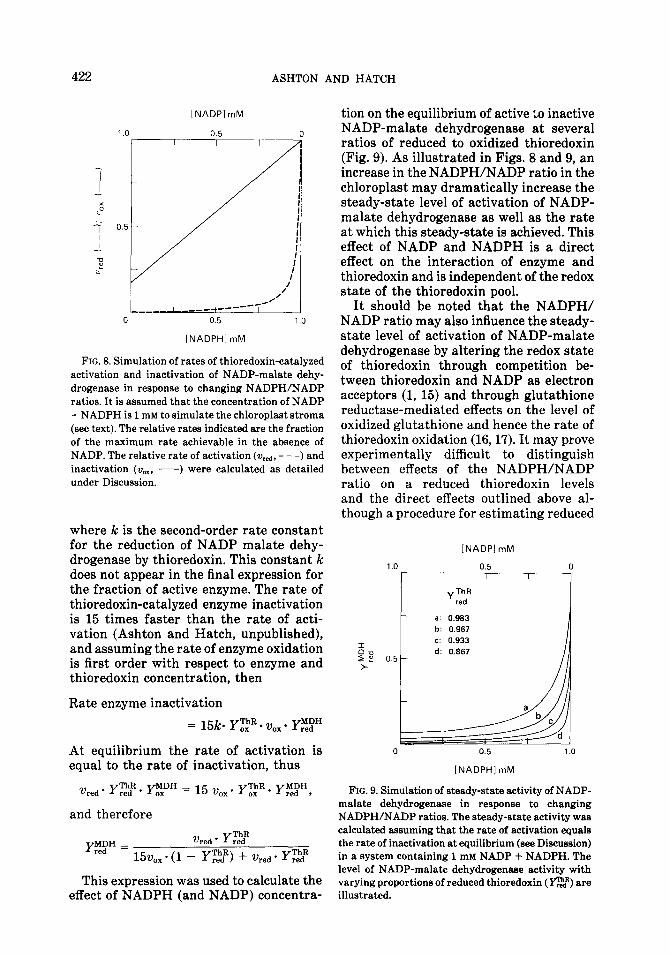
The change in the relative rate of re- duction (v,,~) and oxidation (v,,) of enzyme as [NADPH] increases to 1 mM (and the [NADP] simultaneously declines) is illus- trated in Fig. 8. The marked changes in %&,d compared to vOX as the NADP/NADPH ratio alters is indicative of the weak bind- ing of NADPH to the high-affinity NADP- binding site of inactive NADP-malate de- hydrogenase.
The steady-state level of active NADP- malate dehydrogenase will depend upon the relative rates of the opposing processes of activation and inactivation. Using the rates of thioredoxin-catalyzed inactivation and activation of NADP-malate dehy- drogenase shown in Fig. 8 the extent of activation of the enzyme at the steady state can be estimated as a function of the con- centration of NADPH and NADP as fol- lows: The rate of enzyme activation is first order with respect to thioredoxin and en- zyme concentrations (Ashton and Hatch, unpublished). Thus Rate of enzyme activation
422 ASHTON AND HATCH
[NADPI mM
1 .o 0.5 0
I
J .
1 I
I
P AL
_--- /’ --- _-- --
0 0.5 1 .o
[NADPHI mM
FIG. 8. Simulation of rates of thioredoxin-catalyzed activation and inactivation of NADP-malate dehy- drogenase in response to changing NADPH/NADP ratios. It is assumed that the concentration of NADP + NADPH is 1 mM to simulate the chloroplast stroma (see text). The relative rates indicated are the fraction of the maximum rate achievable in the absence of NADP. The relative rate of activation (v,~, - - -) and inactivation (v,,, -) were calculated as detailed under Discussion.
where k is the second-order rate constant for the reduction of NADP malate dehy- drogenase by thioredoxin. This constant k does not appear in the final expression for the fraction of active enzyme. The rate of thioredoxin-catalyzed enzyme inactivation is 15 times faster than the rate of acti- vation (Ashton and Hatch, unpublished), and assuming the rate of enzyme oxidation is first order with respect to enzyme and thioredoxin concentration, then
Rate enzyme inactivation
= 15k. Y ::‘a vex. Y?edDH
At equilibrium the rate of activation is equal to the rate of inactivation, thus
yMDH ox = 15 vu,, - OX yThR. yfffH f
and therefore
This expression was used to calculate the effect of NADPH (and NADP) concentra-
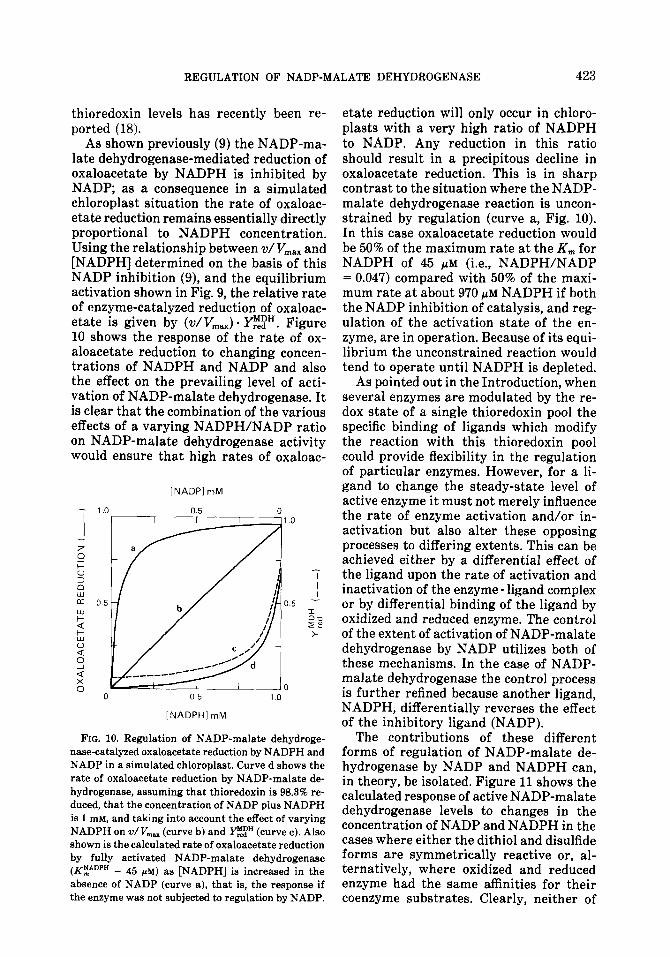
tion on the equilibrium of active to inactive NADP-malate dehydrogenase at several ratios of reduced to oxidized thioredoxin (Fig. 9). As illustrated in Figs. 8 and 9, an increase in the NADPH/NADP ratio in the chloroplast may dramatically increase the steady-state level of activation of NADP- malate dehydrogenase as well as the rate at which this steady-state is achieved. This effect of NADP and NADPH is a direct effect on the interaction of enzyme and thioredoxin and is independent of the redox state of the thioredoxin pool.
It should be noted that the NADPH/ NADP ratio may also influence the steady- state level of activation of NADP-malate dehydrogenase by altering the redox state of thioredoxin through competition be- tween thioredoxin and NADP as electron acceptors (1, 15) and through glutathione reductase-mediated effects on the level of oxidized glutathione and hence the rate of thioredoxin oxidation (16,17). It may prove experimentally difficult to distinguish between effects of the NADPH/NADP ratio on a reduced thioredoxin levels and the direct effects outlined above al- though a procedure for estimating reduced
[NADPI mM
1 .o 0.5 0 I I I
yThR red
a: 0.983 I
b: 0.967
c:
6,
0.933
d: 0.867 2: 0.5 - z
0 0.5 1 .o
[NADPH] mM
FIG. 9. Simulation of steady-state activity of NADP- malate dehydrogenase in response to changing NADPH/NADP ratios. The steady-state activity was calculated assuming that the rate of activation equals the rate of inactivation at equilibrium (see Discussion) in a system containing 1 mM NADP + NADPH. The level of NADP-malate dehydrogenase activity with varying proportions of reduced thioredoxin ( ptir‘) are illustrated.
REGULATION OF NADP-MALATE DEHYDROGENASE 423
thioredoxin levels has recently been re- ported (18).
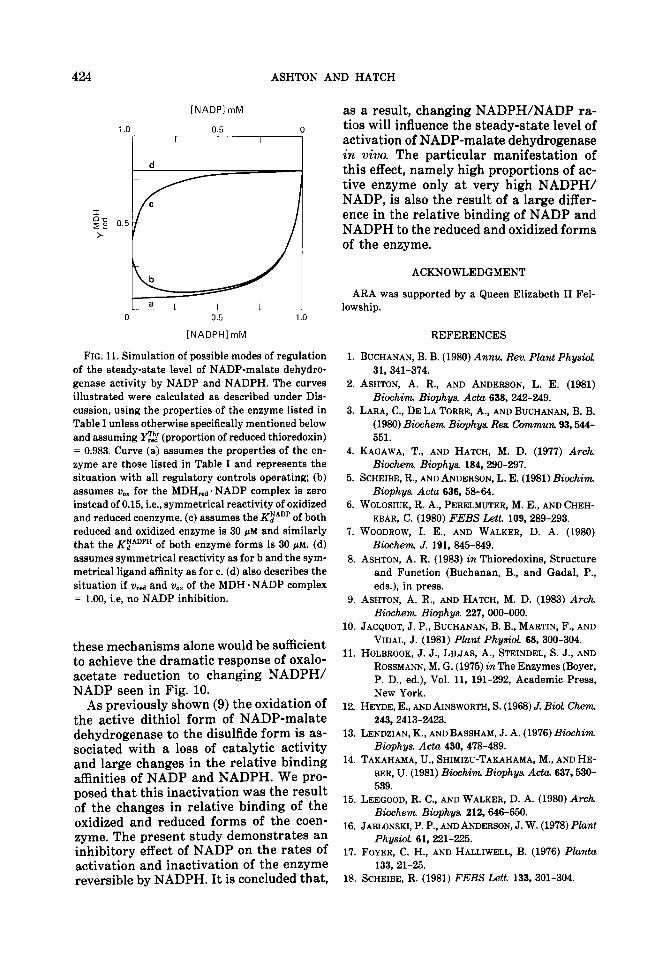
As shown previously (9) the NADP-ma- late dehydrogenase-mediated reduction of oxaloacetate by NADPH is inhibited by NADP; as a consequence in a simulated chloroplast situation the rate of oxaloac- etate reduction remains essentially directly proportional to NADPH concentration. Using the relationship between v/V,,, and [NADPH] determined on the basis of this NADP inhibition (9), and the equilibrium activation shown in Fig. 9, the relative rate of enzyme-catalyzed reduction of oxaloac- etate is given by (v/Vmax) . YE?“. Figure 10 shows the response of the rate of ox- aloacetate reduction to changing concen- trations of NADPH and NADP and also the effect on the prevailing level of acti- vation of NADP-malate dehydrogenase. It is clear that the combination of the various effects of a varying NADPH/NADP ratio on NADP-malate dehydrogenase activity would ensure that high rates of oxaloac-
[NADPI mM
0 0.5 1 .o
[NADPHI mM
FIG. 10. Regulation of NADP-malate dehydroge- nase-catalyzed oxaloacetate reduction by NADPH and NADP in a simulated chloroplast. Curve d shows the rate of oxaloacetate reduction by NADP-malate de- hydrogenase, assuming that thioredoxin is 98.3% re- duced, that the concentration of NADP plus NADPH is 1 mM, and taking into account the effect of varying NADPH on v/V,, (curve b) and Y”z” (curve c). Also shown is the calculated rate of oxaloacetate reduction by fully activated NADP-malate dehydrogenase (KEAoPH = 45 pM) as [NADPH] is increased in the absence of NADP (curve a), that is, the response if the enzyme was not subjected to regulation by NADP.
etate reduction will only occur in chloro- plasts with a very high ratio of NADPH to NADP. Any reduction in this ratio should result in a precipitous decline in oxaloacetate reduction. This is in sharp contrast to the situation where the NADP- malate dehydrogenase reaction is uncon- strained by regulation (curve a, Fig. 10). In this case oxaloacetate reduction would be 50% of the maximum rate at the Km for NADPH of 45 PM (i.e., NADPH/NADP = 0.047) compared with 50% of the maxi- mum rate at about 970 PM NADPH if both the NADP inhibition of catalysis, and reg- ulation of the activation state of the en- zyme, are in operation. Because of its equi- librium the unconstrained reaction would tend to operate until NADPH is depleted.
As pointed out in the Introduction, when several enzymes are modulated by the re- dox state of a single thioredoxin pool the specific binding of ligands which modify the reaction with this thioredoxin pool could provide flexibility in the regulation of particular enzymes. However, for a li- gand to change the steady-state level of active enzyme it must not merely influence the rate of enzyme activation and/or in- activation but also alter these opposing processes to differing extents. This can be achieved either by a differential effect of the ligand upon the rate of activation and inactivation of the enzyme * ligand complex or by differential binding of the ligand by oxidized and reduced enzyme. The control of the extent of activation of NADP-malate dehydrogenase by NADP utilizes both of these mechanisms. In the case of NADP- malate dehydrogenase the control process is further refined because another ligand, NADPH, differentially reverses the effect of the inhibitory ligand (NADP).
The contributions of these different forms of regulation of NADP-malate de- hydrogenase by NADP and NADPH can, in theory, be isolated. Figure 11 shows the calculated response of active NADP-malate dehydrogenase levels to changes in the concentration of NADP and NADPH in the cases where either the dithiol and disulfide forms are symmetrically reactive or, al- ternatively, where oxidized and reduced enzyme had the same affinities for their coenzyme substrates. Clearly, neither of
424 ASHTON AND HATCH
[NADPI mM
1.0 0.5
l----i d
a , I I 0 0.5 1.0
[NADPHI mM
FIG. 11. Simulation of possible modes of regulation of the steady-state level of NADP-malate dehydro- genase activity by NADP and NADPH. The curves illustrated were calculated as described under Dis- cussion, using the properties of the enzyme listed in Table I unless otherwise specifically mentioned below and assuming cs (proportion of reduced thioredoxin) = 0.983. Curve (a) assumes the properties of the en- zyme are those listed in Table I and represents the situation with all regulatory controls operating; (b) assumes uO:, for the MDH,,d. NADP complex is zero instead of 0.15, i.e., symmetrical reactivity of oxidized and reduced coenzyme. (c) assumes the K!ADp of both reduced and oxidized enzyme is 30 PM and similarly that the K2ADPH of both enzyme forms is 30 FM. (d) assumes symmetrical reactivity as for b and the sym- metrical ligand affinity as for c. (d) also describes the situation if 2r,d and w,, of the MDH * NADP complex = 1.00, i.e, no NADP inhibition.
these mechanisms alone would be sufficient to achieve the dramatic response of oxalo- acetate reduction to changing NADPH/ NADP seen in Fig. 10.
As previously shown (9) the oxidation of the active dithiol form of NADP-malate dehydrogenase to the disulfide form is as- sociated with a loss of catalytic activity and large changes in the relative binding affinities of NADP and NADPH. We pro- posed that this inactivation was the result of the changes in relative binding of the oxidized and reduced forms of the coen- zyme. The present study demonstrates an inhibitory effect of NADP on the rates of activation and inactivation of the enzyme reversible by NADPH. It is concluded that,
as a result, changing NADPH/NADP ra- tios will influence the steady-state level of activation of NADP-malate dehydrogenase in Go. The particular manifestation of this effect, namely high proportions of ac- tive enzyme only at very high NADPH/ NADP, is also the result of a large differ- ence in the relative binding of NADP and NADPH to the reduced and oxidized forms of the enzyme.
ACKNOWLEDGMENT
ARA was supported by a Queen Elizabeth II Fel- lowship.
REFERENCES
1. BUCHANAN, B. B. (1980) Annu. Rev. Plant Physiol 31.341-374.
2. ASHTON, A. R., AND ANDERSON, L. E. (1981) B&him. Biophys. Acta 638.242-249.
3. LARA, C., DE LA TORRE, A., AND BUCHANAN, B. B. (1980) Biochem Biophys Res Commun 93,544- 551.
4. KAGAWA, T., AND HATCH, M. D. (1977) Arch
Biochem Biophys. 184,290-29’7. 5. SCHEIBE, R., AND ANDERSON, L. E. (1981) B&him
Biophys. Acta 636, 58-64. 6. WOLOSIUK, R. A., PERELMUTER, M. E., AND CHEH-
EBAR, c. (1980) FEBS L&t. 109,289-293. ‘7. WOODROW, I. E., AND WALKER, D. A. (1980)
Biochem. J. 191,845~849. 8. ASHTON, A. R. (1983) in Thioredoxins, Structure
and Function (Buchanan, B., and Gadal, P., eds.), in press.
9. ASHTON, A. R., AND HATCH, M. D. (1983) Arch. Biochxm Biophys. 227,000-000.
10. JACQUOT, J. P., BUCHANAN, B. B., MARTIN, F., AND VIDAL, J. (1981) Plant Physiol 68, 300-304.
11. HOLBROOK, J. J., LILJAS, A., STEINDEL, S. J., AND ROSSMANN, M. G. (1975) in The Enzymes (Boyer, P. D., ed.), Vol. 11, 191-292, Academic Press, New York.
12. HEYDE, E., AND AINSWORTH, S. (1968) J. Biol Chen~ 243,2413-2423.
13. LENDZIAN, K., AND BASSHAM, J. A. (1976) Biochim. Biophys. Acta 430,478-489.
14. TAKAHAMA, U., SHIMIZU-TAKAHAMA, M., AND HE- BER, U. (1981) Biochim Biophys Acta 637,530- 539.
15. LEEGOOD, R. C., AND WALKER, D. A. (1980) Arch Biochem Biophys. 212,646-650.
16. JABLONSKI, P. P., AND ANDERSON, J. W. (1978) Plant Physiol 61,221-225.
17. FOYER, C. H., AND HALLIWELL, B. (1976) Planta 133, 21-25.
18. SCHEIBE, R. (1981) FEBS L&t. 133.301-304.





























