Embed Size (px)
Citation preview

Vol. 17, No. 5JOURNAL OF CLINICAL MICROBIOLOGY, May 1983, p. 791-7980095-1137/83/050791-08$02.00/0Copyright C 1983, American Society for Microbiology
Relative Substrate Affinity Index Values: a Method forIdentification of Beta-Lactamase Enzymes and Prediction of
Successful Beta-Lactam TherapyRICHARD JAMES
School of Biological Sciences, University of East Anglia, Norwich NR4 7TJ, Norfolk, England
Received 26 July 1982/Accepted 21 January 1983
Using a nitrocefin competition assay, I determined the relative substrate affinityindex (RSAI) values of nine clinically significant beta-lactamase enzymes againsta range of beta-lactams. Using selected beta-lactam substrates, I observed largedifferences in the RSAI values of the nine enzymes that were sufficient in manycases to positively identify specific enzymes. I made use of the unique RSAIvalues of SHV-1, TEM-1, and TEM-2 beta lactamases with cefoxitin to screen forthe presence of these enzymes in Klebsiella aerogenes clinical isolates. The RSAIvalues also allow for the prediction of the outcome of beta-lactam therapy againstspecific beta-lactamase-producing isolates.
Resistance to beta-lactam antibiotics is nowwidespread among many bacterial genera. Themost common mechanism of resistance is theproduction of beta-lactamases, enzymes whichhydrolyze the beta-lactam ring and inactivateantibiotics. Although many gram-negative bac-teria produce inducible, chromosomally codedbeta-lactamases, in all but a few cases their rolein resistance to beta-lactams appears to be opento doubt. In gram-negative bacteria, severalconstitutive, plasmid-coded beta-lactamaseshave been identified on the basis of their sub-strate profiles against a range of beta-lactamsand their isoelectric points (5). A variety ofmethods is available to determine substrate pro-files (6), but they suffer from a number ofdisadvantages. For example, hydrolysis of peni-cillins and cephalosporins may be monitored bya fall in UV absorption at a wavelength charac-teristic of each beta-lactam used, but it is readilyapparent that relatively pure enzyme prepara-tions have to be used in such assays to reducebackground absorption. In addition to beinglaborious to perform, many of the assay meth-ods are insensitive. Although capable of detect-ing gross differences in relative hydrolysis ratesof beta-lactams, substrate profiles often fail todistinguish between a low rate of hydrolysis dueto a low affinity of the enzyme for the substrateand one due to enzyme inhibition by the sub-strate. This distinction is becoming more impor-tant with the introduction of beta-lactamase in-hibitors, such as clavulanic acid.Given the large differences in the hydrolysis
rates of beta-lactams by different beta-lacta-mases, it is possible to predict that many beta-lactams are likely to be ineffective in vivo
against a pathogen producing specific beta-lacta-mases. Although a mass of evidence is availableon the trends of antibiotic susceptibility of clini-cal isolates (1), little practical information isavailable on the contribution of specific beta-lactamases, largely because of the difficulty ofidentification. The problem cannot be ignored,however, because beta-lactam antibiotics con-stitute our principal defense against pathogenicbacteria, and a policy for sensible antibiotic usedepends upon the avoidance of over-prescrip-tion of the newer cephalosporins when an older(and cheaper) beta-lactam would be effective. Itis impractical to test every clinical isolateagainst every possible beta-lactam and all possi-ble combinations of beta-lactamase inhibitorsand beta-lactams; therefore, a data base of thesensitivity of specific beta-lactamase-producingclinical isolates to a range of beta lactams isneeded.
I have developed a very sensitive assay whichallows the quantitation of the relative affinitiesof beta-lactamases for beta-lactam antibiotics.The assay is rapid, simple to perform, andamenable to automation. It yields a wealth ofinformation on the interactions between beta-lactamases and their substrates which is usefulin the prediction of successful beta-lactam thera-py. Differences in relative affinities of a varietyof clinically important beta-lactamases now alsoallow simple beta-lactamase identification and,in some cases, even bacterial identification.
MATERIALS AND METHODS
Bacterial strains and media. Escherichia coli C9 wasobtained from the Coli Genetic Stock Center, NewHaven, Conn. The transmissible plasmid RP4, which
791
on July 18, 2020 by guesthttp://jcm
.asm.org/
Dow
nloaded from

792 JAMES
codes for the TEM-2 beta-lactamase (2), was trans-ferred into E. coli C9 by plate mating. Enterobactercloacae was a laboratory strain which produces aconstitutive, chromosomally coded beta-lactamase. E.coli strains 2136E, 2008E, 1946E, 1527E, 2139E, and2140E, which produce TEM-1, SHV-1, HMS-1, OXA-1, OXA-2, and OXA-3 beta-lactamases, respectively,were a generous gift from the Glaxo Research Labora-tory, London, England.
For the isolation of the reference beta-lactamaseextracts, 300 ml of culture in a 1-liter flask was grownto stationary phase with shaking at 37°C in nutrientbroth (Oxoid Ltd., London, England). For screeningof beta-lactamases, 50 ml of culture was used toprepare the beta-lactamase.
Preparation of beta-lactamase extracts. All proce-dures were carried out at 4°C. Cell pellets wereobtained by centrifugation at 9,000 rpm for 15 min in aSorvall RC2B centrifuge. The pellets were washed in40 ml of 10 mM sodium phosphate buffer (pH 7.0) andcentrifuged again as described above. The washed cellpellets were suspended in 16 ml of the same buffer andsonicated in an Ultrasonics sealed-atmosphere type7530-5A treatment chamber for four periods of 30 seach with 60-s cooling periods. The sonicated extractswere centrifuged at 40,000 rpm for 40 min in a Beck-man L2 centrifuge. The resulting supernatants werestored in portions at -20°C until required.Assay of beta-lactamase activity. The assay mixture
(890 ,ul of 10 mM sodium phosphate buffer [pH 7.0 andprewarmed to 37°C], 100 p.l of the chromogenic sub-strate nitrocefin [7] made up in 10 mM phosphatebuffer [pH 7.0] at 500 ,ug/ml) was prepared in a 1-cmlight path spectrophotometer cuvette. After the tem-perature was equilibrated to 37°C in a heated cuvetteholder, the assay was started by the addition of 10 ,ulof the beta-lactamase extract (diluted if necessary in 10mM phosphate buffer [pH 7.0]).The change in optical density at a wavelength of 486
nm (OD486) was recorded over a 5-min period to follownitrocefin cleavage. The reference cuvette containedbuffer and nitrocefin but no enzyme. Data recording,graph plotting, and calculation of the percent inhibi-tion of nitrocefin cleavage were performed by anApple microcomputer interfaced to the spectropho-tometer via an analog-digital converter. In all assays, Iadjusted the amount of beta-lactamase extract to givean increase in the OD486 of the control of approximate-ly 0.15 to 0.25 per min.
Competition assays. For beta-lactam competitionassays, up to 100 p.1 of individual beta-lactam antibiot-ics made up in 10 mM phosphate buffer (pH 7.0) wassubstituted for an equivalent volume of buffer in theassay mixture. Concentrated stock solutions of thebeta-lactams were stored at -20°C for up to 4 weekswithout any significant loss of activity.
Preincubation assays were carried out by incubationof buffer, enzyme, and competing beta-lactam in acuvette at 37°C for various times before the addition ofnitrocefin. The percent inhibition of nitrocefin cleav-age was calculated for each concentration of compet-ing beta-lactam used, and a graph was plotted ofpercent inhibition against concentration.
Determination of MIC. For minimal inhibitory con-centration (MIC) determinations, doubling dilutions ofantibiotics were made in Oxoid nutrient broth inmicrotiter dishes. An inoculum of 106 cells of E. coli
J. CLIN. MICROBIOL.
C9 or E. coli C9RP4 was added and incubation wascarried out at 37°C for 16 h.
Isoelectric focusing. Volumes of 10 ml of the beta-lactamase extracts were loaded directly onto the sur-face of an Ampholine PAG plate (pH 3.5 to 9.5; LKBInstruments, Inc., Rockville, Md.), and isoelectricfocusing was performed according to the instructionsof the manufacturer. Beta-lactamase bands were visu-alized by staining with nitrocefin; pink bands appearedwithin a few minutes and were photographed with aPolaroid MP4 camera, Polaroid 665 film, and a KodakWratten 44 green filter.
Antibiotics. Cefoxitin was a generous gift fromMerck Sharp & Dohme Inc., West Point, Pa.; cefotax-ime was obtained from Hoechst-Roussel Pharmaceuti-cals Inc., Somerville, N.J.; ceftizoxime was obtainedfrom Fujisawa Pharmaceutical Co., Osaka, Japan;moxalactam was obtained from Eli Lilly & Co., India-napolis, Ind.; clavulanic acid, cefuroxime, ceftazi-dime, and nitrocefin were obtained from Glaxo Re-search Laboratory; SQ 14,359 was obtained fromE. R. Squibb & Sons, Princeton, N.J.; Ro 13-9904 wasobtained from Hoffmann-La Roche Inc., Nutley, N.J.;and CP 45,899, a beta-lactamase inhibitor, was ob-tained from Pfizer Inc., New York. All other antibiot-ics were purchased commercially.
RESULTS
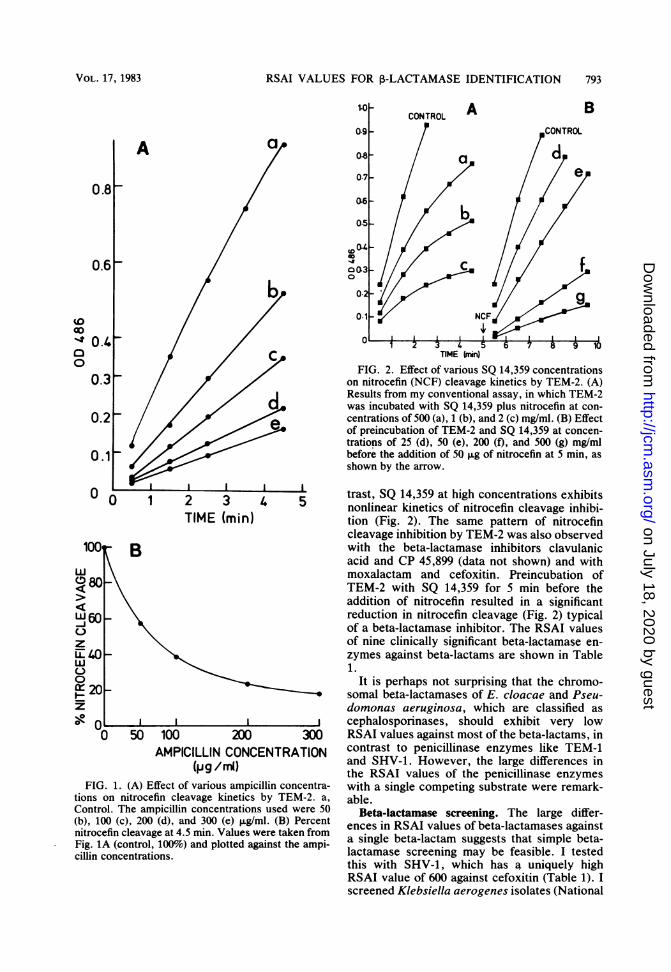
Determination of RSAI values. The chromo-genic substrate nitrocefin is readily cleaved byall of the beta-lactamases I used. Beta-lactamantibiotics which are substrates of a beta-lacta-mase enzyme readily compete with nitrocefin inmy assay. The effect of various concentrationsof ampicillin on the cleavage of nitrocefin byTEM-2 is shown in Fig. 1A. From a plot ofpercent nitrocefin cleavage against ampicillinconcentration, the ampicillin concentrationwhich results in 50% inhibition of the activity ofTEM-2 (Fig. 1B) can be determined. This ampi-cillin concentration is a direct estimate of therelative affinity of TEM-2 for ampicillin andnitrocefin. For ease of comparison of the affinityvalues of a specific beta-lactamase against dif-ferent beta-lactam antibiotics, I propose theterm relative substrate affinity index (RSAI),which is the ratio of the beta-lactam concentra-tion giving 50% inhibition of nitrocefin cleavageto the nitrocefin concentration used in the assay.In the case of ampicillin, the RSAI value is 1.3.The effect of competition by beta-lactams on
nitrocefin cleavage kinetics also yields informa-tion on the mechanism of interaction between anantibiotic and a beta-lactamase. For example,most of the beta-lactams tested against TEM-2show a pattern of inhibition similar to that ofampicillin (Fig. 1A); i.e., increasing the beta-lactam concentration reduces the rate of nitroce-fin cleavage (shown as a reduced slope of theincrease in OD486 value), but the plots are stilllinear. This is characteristic of competitive inhi-bition between ampicillin and nitrocefin. In con-
on July 18, 2020 by guesthttp://jcm
.asm.org/
Dow
nloaded from
RSAI VALUES FOR ,-LACTAMASE IDENTIFICATION
L.
0.8L
QD-t 0.400
0.3
0.2h
0.1
0 1 2 3TIME (min)
B
I I
0 50 100 200 300AMPICILLIN CONCENTRATION
(pg /mi)FIG. 1. (A) Effect of various ampicillin concentra-
tions on nitrocefin cleavage kinetics by TEM-2. a,Control. The ampicillin concentrations used were 50(b), 100 (c), 200 (d), and 300 (e) ,ug/ml. (B) Percentnitrocefin cleavage at 4.5 min. Values were taken fromFig. 1A (control, 100%) and plotted against the ampi-cillin concentrations.
0
100
CD80
W 60
zii 40w
0
'20LI-z
C TIME hin)FIG. 2. Effect of various SQ 14,359 concentrations
on nitrocefin (NCF) cleavage kinetics by TEM-2. (A)Results from my conventional assay, in which TEM-2
d was incubated with SQ 14,359 plus nitrocefin at con-9. centrations of 500 (a), 1 (b), and 2 (c) mg/ml. (B) Effect
of preincubation of TEM-2 and SQ 14,359 at concen-trations of 25 (d), 50 (e), 200 (f), and 500 (g) mg/mlbefore the addition of 50 ,ug of nitrocefin at 5 min, asshown by the arrow.
trast, SQ 14,359 at high concentrations exhibits4 5 nonlinear kinetics of nitrocefin cleavage inhibi-
tion (Fig. 2). The same pattern of nitrocefincleavage inhibition by TEM-2 was also observedwith the beta-lactamase inhibitors clavulanicacid and CP 45,899 (data not shown) and withmoxalactam and cefoxitin. Preincubation ofTEM-2 with SQ 14,359 for 5 min before theaddition of nitrocefin resulted in a significantreduction in nitrocefin cleavage (Fig. 2) typicalof a beta-lactamase inhibitor. The RSAI valuesof nine clinically significant beta-lactamase en-zymes against beta-lactams are shown in Table1.
It is perhaps not surprising that the chromo-somal beta-lactamases of E. cloacae and Pseu-domonas aeruginosa, which are classified ascephalosporinases, should exhibit very lowRSAI values against most of the beta-lactams, incontrast to penicillinase enzymes like TEM-1and SHV-1. However, the large differences inthe RSAI values of the penicillinase enzymeswith a single competing substrate were remark-able.
Beta-lactamase screening. The large differ-ences in RSAI values of beta-lactamases againsta single beta-lactam suggests that simple beta-lactamase screening may be feasible. I testedthis with SHV-1, which has a uniquely highRSAI value of 600 against cefoxitin (Table 1). Iscreened Klebsiella aerogenes isolates (National
VOL. 17, 1983 793
0.6r
on July 18, 2020 by guesthttp://jcm
.asm.org/
Dow
nloaded from

TABLE 1. RSAI values of nine clinically significant beta-lactamasesRSAI value of following beta-lactamasea:
Beta-lactamP. aeruginosa E. cloacae OXA-3 OXA-2 OXA-1 HMS-1 SHV-1 TEM-2 TEM-1
Penicillin 0.13 0.08 0.4 0.6 0.2 0.5 1 1 3Ampicillin 0.12 0.02 0.2 10 4 0.5 3 2.6 4Cloxacillin 0.00017 0.00004 0.01 30 0.6 0.02 2 0.5 0.75Cephalothin 0.7 0.6 0.4 0.7 4 1 7 15 10Cephalozolin 15 17 3.4 34 10 0.8 15 48 30Cephaloridine 1.5 16 4.4 4 60 6 30 20 40Cefoxitin 0.003 0.01 0.004 0.2 20 60 600 60* 60*Cefuroxime 0.0001 0.0002 0.003 3.6 10 16 20 40 50Cefotaxime 0.004 0.0006 15 12 9 11 400 600 400SQ 14,359 0.004 0.002 0.6* 0.7* 50* 1.5* 45* 15* 10*Moxalactam 0.0006* 0.0001* 1* 3* 4* 0.3* 280 60* 40*Cefoperazone 0.5 1.3 2 2 4.5 0.2 4.5 10 8Carbenicillin 0.008 0.0003* 12 15 0.5 0.09 0.5 0.4 0.6Cefamandole 5 0.03 0.75 1.7 1.5 0.5 6.6 11 11Cephradine 1 1.8 100 86 420 25 15 50 35Mecillinam 68 1.3* 30 100 400 2 80 80 90Ceftizoxime 500 0.1 200 110 50 66 400 1,000 1,000Ceftazidime 0.46* 0.1* >400 900 >2,000 10 1,140 1,500 >400
a *, Nonlinear inhibition kinetics.
Collection of Type Cultures, Central PublicHealth Laboratory, London NW9 5HT, En-gland) for the presence of SHV-1 by using aconcentration of 30 mg of cefoxitin per ml in thestandard nitrocefin competition assay. All butone of the K. aerogenes isolates contained abeta-lactamase, as judged by nitrocefin cleav-age, The percent inhibition of nitrocefin cleav-age by cefoxitin in all other isolates was 50 +8%, with the exception of NCTC 1073, whichwas inhibited by only 23%. Isoelectric focusingof the extracts of each isolate confirmed thepresence of a single major band in each, whichhad the same isoelectric point as the SHV-1control enzyme (pl = 7.6), except in the case ofNCTC 1073 (pl = 6.5). The RSAI values of theNCTC 1073 beta-lactamase are shown in Table2. It is of interest that this enzyme does notexhibit nonlinear inhibition kinetics with any ofthe beta-lactams tested, in contrast to SHV-1.
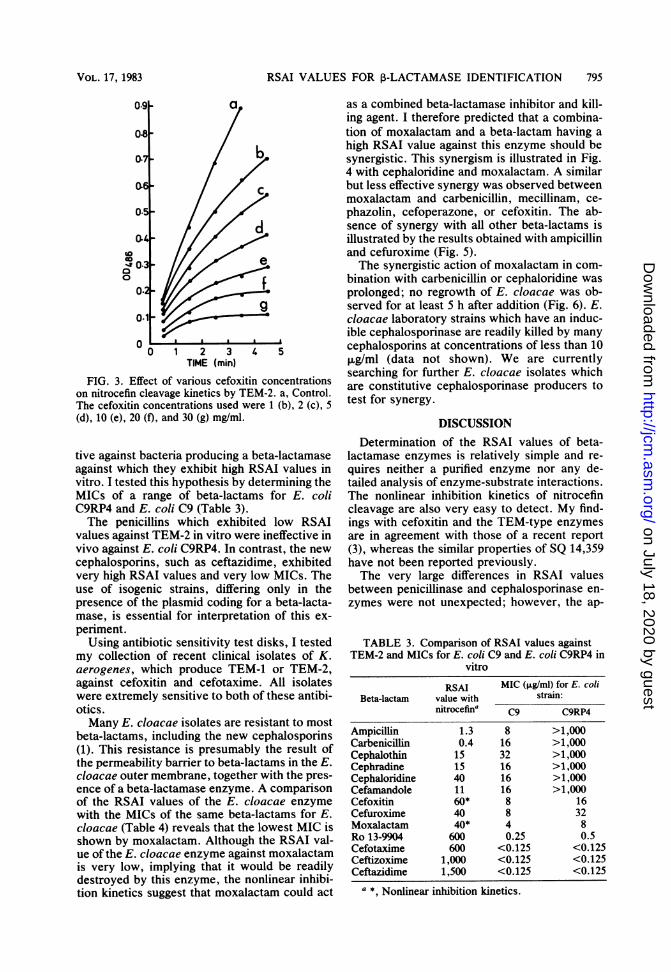
Cefoxitin at a concentration of 30 mg/ml isalso useful to screen for the presence of TEM-1or TEM-2, owing to the characteristic nonlinearinhibition kinetics of nitrocefin cleavage (Fig. 3).I screened a large number of recent clinicalisolates of K. aerogenes (Public Health Labora-tory, Norwich, Norfolk, England) with cefoxi-tin. Over 60% exhibited the characteristic non-linear inhibition kinetics of nitrocefin cleavagewith cefoxitin. Isoelectric focusing with controlSHV-1, TEM-1, and TEM-2 confirmed the pres-ence of TEM-1 or TEM-2 in those isolatespredicted from the above test. In all of theisolates which produced a TEM-type enzyme,we also detected SHV-1 on the isoelectric focus-ing gels when they were overloaded with respectto TEM-1 or TEM-2.
TABLE 2. Comparison of RSAI values of SHV-1and K1073
RSAI value of following beta-Beta-lactam lactamasea:
SHV-1 K1073
Ampicillin 3 6.7Cephazolin 15 16Cephaloridine 30 19.2Cefoxitin 600 1,020SQ 14,359 45* 60Ceftizoxime 1,400 720CP 45,899 0.018* 0.18
a*, Nonlinear inhibition kinetics.
A third example of the use of RSAI values inbeta-lactamase identification involves an ampi-cillin-resistant isolate obtained from a watersample from the River Wensum next to theUniversity of East Anglia, Norwich, Norfolk,England. After confirming the presence of abeta-lactamase which had an RSAI value of 0.02against cefoxitin, I suspected the E. cloacaebeta-lactamase. This suspicion was confirmedby determining the RSAI value of the extractagainst cephaloridine (RSAI value, 15.6) andthat against clavulanic acid (RSAI value, 48).My identification of this river isolate as E.cloacae was confirmed by a blind bacterial iden-tification test kindly performed for me by thePublic Health Laboratory.RSAI values and MICs. A high RSAI value
implies that the beta-lactam is a poor substratefor that beta-lactamase. Therefore, in the ab-sence of any permeability barrier to their pene-tration, beta-lactams should be clinically effec-
794 JAMES J. CLIN. MICROBIOL.
on July 18, 2020 by guesthttp://jcm
.asm.org/
Dow
nloaded from
RSAI VALUES FOR P-LACTAMASE IDENTIFICATION 795
0-9F048-
0.71
O*61
0.51
0a4D
o
021
0.1
0 L.0 1 2 3 4
TIME (min)5
FIG. 3. Effect of various cefoxitin concentrationson nitrocefin cleavage kinetics by TEM-2. a, Control.The cefoxitin concentrations used were 1 (b), 2 (c), 5(d), 10 (e), 20 (f), and 30 (g) mg/ml.
tive against bacteria producing a beta-lactamaseagainst which they exhibit high RSAI values invitro. I tested this hypothesis by determining theMICs of a range of beta-lactams for E. coliC9RP4 and E. coli C9 (Table 3).The penicillins which exhibited low RSAI
values against TEM-2 in vitro were ineffective invivo against E. coli C9RP4. In contrast, the newcephalosporins, such as ceftazidime, exhibitedvery high RSAI values and very low MICs. Theuse of isogenic strains, differing only in thepresence of the plasmid coding for a beta-lacta-mase, is essential for interpretation of this ex-
periment.Using antibiotic sensitivity test disks, I tested
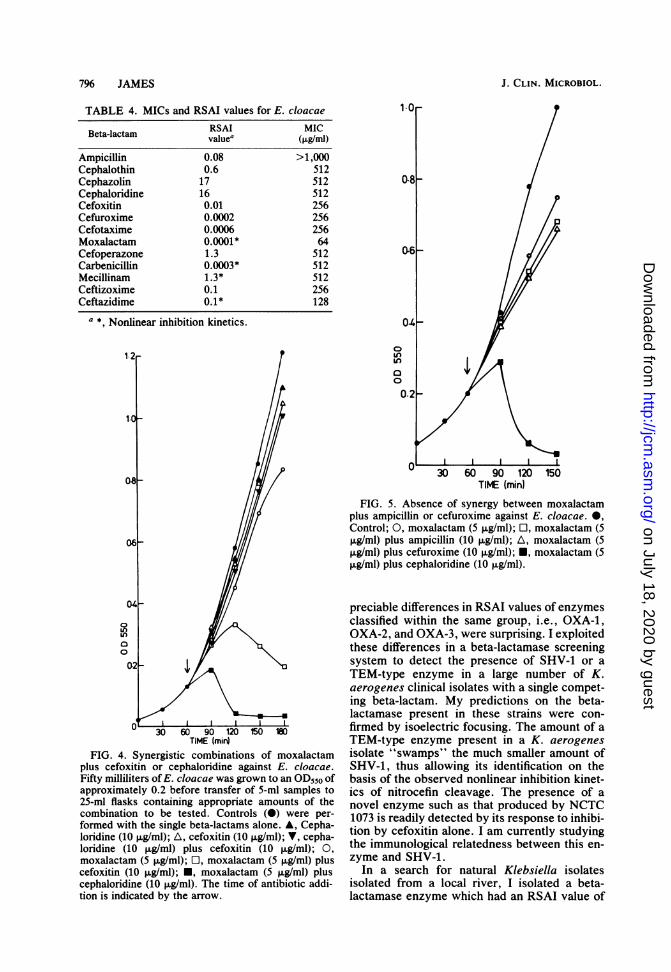
my collection of recent clinical isolates of K.aerogenes, which produce TEM-1 or TEM-2,against cefoxitin and cefotaxime. All isolateswere extremely sensitive to both of these antibi-otics.Many E. cloacae isolates are resistant to most
beta-lactams, including the new cephalosporins(1). This resistance is presumably the result ofthe permeability barrier to beta-lactams in the E.cloacae outer membrane, together with the pres-ence of a beta-lactamase enzyme. A comparisonof the RSAI values of the E. cloacae enzymewith the MICs of the same beta-lactams for E.cloacae (Table 4) reveals that the lowest MIC isshown by moxalactam. Although the RSAI val-ue of the E. cloacae enzyme against moxalactamis very low, implying that it would be readilydestroyed by this enzyme, the nonlinear inhibi-tion kinetics suggest that moxalactam could act
as a combined beta-lactamase inhibitor and kill-ing agent. I therefore predicted that a combina-tion of moxalactam and a beta-lactam having ahigh RSAI value against this enzyme should besynergistic. This synergism is illustrated in Fig.4 with cephaloridine and moxalactam. A similarbut less effective synergy was observed betweenmoxalactam and carbenicillin, mecillinam, ce-phazolin, cefoperazone, or cefoxitin. The ab-sence of synergy with all other beta-lactams isillustrated by the results obtained with ampicillinand cefuroxime (Fig. 5).The synergistic action of moxalactam in com-
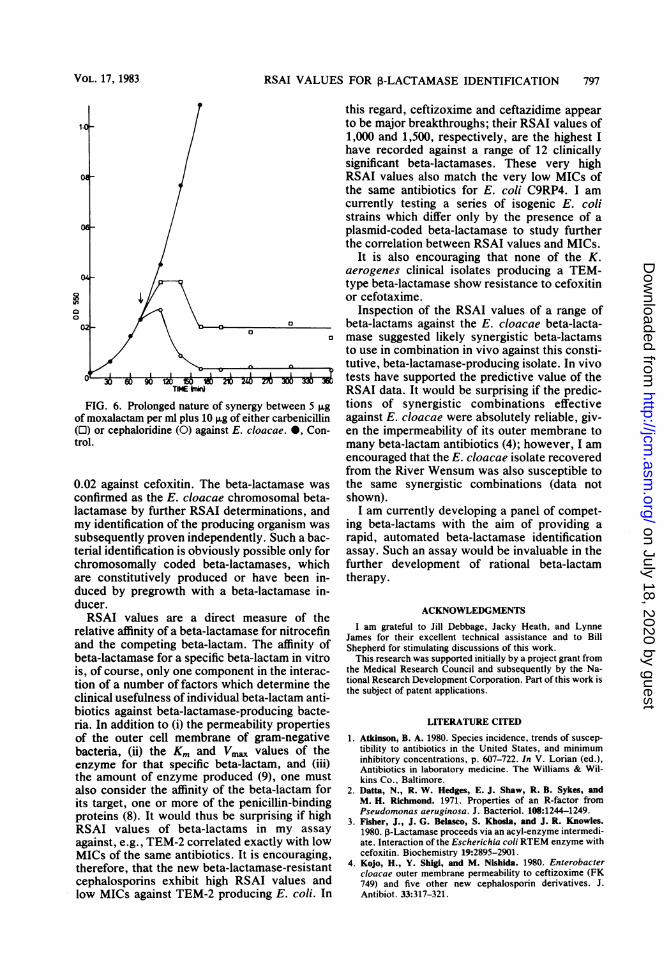
bination with carbenicillin or cephaloridine wasprolonged; no regrowth of E. cloacae was ob-served for at least 5 h after addition (Fig. 6). E.cloacae laboratory strains which have an induc-ible cephalosporinase are readily killed by manycephalosporins at concentrations of less than 10,ug/ml (data not shown). We are currentlysearching for further E. cloacae isolates whichare constitutive cephalosporinase producers totest for synergy.
DISCUSSIONDetermination of the RSAI values of beta-
lactamase enzymes is relatively simple and re-quires neither a purified enzyme nor any de-tailed analysis of enzyme-substrate interactions.The nonlinear inhibition kinetics of nitrocefincleavage are also very easy to detect. My find-ings with cefoxitin and the TEM-type enzymesare in agreement with those of a recent report(3), whereas the similar properties of SQ 14,359have not been reported previously.The very large differences in RSAI values
between penicillinase and cephalosporinase en-zymes were not unexpected; however, the ap-
TABLE 3. Comparison of RSAI values againstTEM-2 and MICs for E. coli C9 and E. coli C9RP4 in
vitro
RSAI MIC (sLg/ml) for E. coliBeta-lactam value with strain:
nitrocefina C9 C9RP4
Ampicillin 1.3 8 >1,000Carbenicillin 0.4 16 >1,000Cephalothin 15 32 >1,000Cephradine 15 16 >1,000Cephaloridine 40 16 >1,000Cefamandole 11 16 >1,000Cefoxitin 60* 8 16Cefuroxime 40 8 32Moxalactam 40* 4 8Ro 13-9904 600 0.25 0.5Cefotaxime 600 <0.125 <0.125Ceftizoxime 1,000 <0.125 <0.125Ceftazidime 1,500 <0.125 <0.125
a *, Nonlinear inhibition kinetics.
VOL. 17, 1983
on July 18, 2020 by guesthttp://jcm
.asm.org/
Dow
nloaded from
J. CLIN. MICROBIOL.
TABLE 4. MICs and RSAI values for E. cloacaeRSAI MICBeta-lactam vAl mI)value' (,ug/ml)
Ampicillin 0.08 >1,000Cephalothin 0.6 512Cephazolin 17 512Cephaloridine 16 512Cefoxitin 0.01 256Cefuroxime 0.0002 256Cefotaxime 0.0006 256Moxalactam 0.0001* 64Cefoperazone 1.3 512Carbenicillin 0.0003* 512Mecillinam 1.3* 512Ceftizoxime 0.1 256Ceftazidime 0.1* 128
a, Nonlinear inhibition kinetics.
30 60 90 120 150 180TIME (minl
FIG. 4. Synergistic combinations of moxalactamplus cefoxitin or cephaloridine against E. cloacae.Fifty milliliters of E. cloacae was grown to an OD550 ofapproximately 0.2 before transfer of 5-ml samples to25-ml flasks containing appropriate amounts of thecombination to be tested. Controls (0) were per-formed with the single beta-lactams alone. A, Cepha-loridine (10 ,ug/ml); A, cefoxitin (10 FLg/ml); V, cepha-loridine (10 pg/ml) plus cefoxitin (10 ,ug/ml); 0,
moxalactam (5 ,g/ml); O, moxalactam (5 ,ug/ml) pluscefoxitin (10 ,ug/ml); *, moxalactam (5 ,ug/ml) pluscephaloridine (10 ,ug/ml). The time of antibiotic addi-tion is indicated by the arrow.
30 60 90 120 150TIME (min)
FIG. 5. Absence of synergy between moxalactamplus ampicillin or cefuroxime against E. cloacae. 0,Control; 0, moxalactam (5 ,ug/ml); O, moxalactam (5,ug/ml) plus ampicillin (10 ,ug/ml); A, moxalactam (5,ug/ml) plus cefuroxime (10 ,ug/ml); *, moxalactam (5,ug/ml) plus cephaloridine (10 ,ug/ml).
preciable differences in RSAI values of enzymesclassified within the same group, i.e., OXA-1,OXA-2, and OXA-3, were surprising. I exploitedthese differences in a beta-lactamase screeningsystem to detect the presence of SHV-1 or aTEM-type enzyme in a large number of K.aerogenes clinical isolates with a single compet-ing beta-lactam. My predictions on the beta-lactamase present in these strains were con-firmed by isoelectric focusing. The amount of aTEM-type enzyme present in a K. aerogenesisolate "swamps" the much smaller amount ofSHV-1, thus allowing its identification on thebasis of the observed nonlinear inhibition kinet-ics of nitrocefin cleavage. The presence of anovel enzyme such as that produced by NCTC1073 is readily detected by its response to inhibi-tion by cefoxitin alone. I am currently studyingthe immunological relatedness between this en-zyme and SHV-1.
In a search for natural Klebsiella isolatesisolated from a local river, I isolated a beta-lactamase enzyme which had an RSAI value of
796 JAMES
on July 18, 2020 by guesthttp://jcm
.asm.org/
Dow
nloaded from
RSAI VALUES FOR 13-LACTAMASE IDENTIFICATION 797
1tO
04
04
0
I
cl0
00
a
I
X0U 91U90 S LU2V 2 MTIME nin
FIG. 6. Prolonged nature of synergy between 5 Fgof moxalactam per ml plus 10 ,ug of either carbenicillin(O) or cephaloridine (0) against E. cloacae. 0, Con-trol.
0.02 against cefoxitin. The beta-lactamase wasconfirmed as the E. cloacae chromosomal beta-lactamase by further RSAI determinations, andmy identification of the producing organism was
subsequently proven independently. Such a bac-terial identification is obviously possible only forchromosomally coded beta-lactamases, whichare constitutively produced or have been in-duced by pregrowth with a beta-lactamase in-ducer.RSAI values are a direct measure of the
relative affinity of a beta-lactamase for nitrocefinand the competing beta-lactam. The affinity ofbeta-lactamase for a specific beta-lactam in vitrois, of course, only one component in the interac-tion of a number of factors which determine theclinical usefulness of individual beta-lactam anti-biotics against beta-lactamase-producing bacte-ria. In addition to (i) the permeability propertiesof the outer cell membrane of gram-negativebacteria, (ii) the Km and Vm. values of theenzyme for that specific beta-lactam, and (iii)the amount of enzyme produced (9), one mustalso consider the affinity of the beta-lactam forits target, one or more of the penicillin-bindingproteins (8). It would thus be surprising if highRSAI values of beta-lactams in my assayagainst, e.g., TEM-2 correlated exactly with lowMICs of the same antibiotics. It is encouraging,therefore, that the new beta-lactamase-resistantcephalosporins exhibit high RSAI values andlow MICs against TEM-2 producing E. coli. In
this regard, ceftizoxime and ceftazidime appearto be major breakthroughs; their RSAI values of1,000 and 1,500, respectively, are the highest Ihave recorded against a range of 12 clinicallysignificant beta-lactamases. These very highRSAI values also match the very low MICs ofthe same antibiotics for E. coli C9RP4. I amcurrently testing a series of isogenic E. colistrains which differ only by the presence of aplasmid-coded beta-lactamase to study furtherthe correlation between RSAI values and MICs.
It is also encouraging that none of the K.aerogenes clinical isolates producing a TEM-type beta-lactamase show resistance to cefoxitinor cefotaxime.
Inspection of the RSAI values of a range ofbeta-lactams against the E. cloacae beta-lacta-mase suggested likely synergistic beta-lactamsto use in combination in vivo against this consti-tutive, beta-lactamase-producing isolate. In vivotests have supported the predictive value of theRSAI data. It would be surprising if the predic-tions of synergistic combinations effectiveagainst E. cloacae were absolutely reliable, giv-en the impermeability of its outer membrane tomany beta-lactam antibiotics (4); however, I amencouraged that the E. cloacae isolate recoveredfrom the River Wensum was also susceptible tothe same synergistic combinations (data notshown).
I am currently developing a panel of compet-ing beta-lactams with the aim of providing arapid, automated beta-lactamase identificationassay. Such an assay would be invaluable in thefurther development of rational beta-lactamtherapy.
ACKNOWLEDGMENTS
I am grateful to Jill Debbage, Jacky Heath, and LynneJames for their excellent technical assistance and to BillShepherd for stimulating discussions of this work.This research was supported initially by a project grant from
the Medical Research Council and subsequently by the Na-tional Research Development Corporation. Part of this work isthe subject of patent applications.
LITERATURE CITED
1. Atkinson, B. A. 1980. Species incidence, trends of suscep-tibility to antibiotics in the United States, and minimuminhibitory concentrations, p. 607-722. In V. Lorian (ed.),Antibiotics in laboratory medicine. The Williams & Wil-kins Co., Baltimore.
2. Datta, N., R. W. Hedges, E. J. Shaw, R. B. Sykes, andM. H. Richmond. 1971. Properties of an R-factor fromPseudomonas aeruginosa. J. Bacteriol. 108:1244-1249.
3. Fisher, J., J. G. Belasco, S. Khosba, and J. R. Knowles.1980. ,B-Lactamase proceeds via an acyl-enzyme intermedi-ate. Interaction of the Escherichia coli RTEM enzyme withcefoxitin. Biochemistry 19:2895-2901.
4. Kojo, H., Y. Shigi, and M. Nishida. 1980. Enterobactercloacae outer membrane permeability to ceftizoxime (FK749) and five other new cephalosporin derivatives. J.Antibiot. 33:317-321.
VOL. 17, 1983
on July 18, 2020 by guesthttp://jcm
.asm.org/
Dow
nloaded from

798 JAMES J. CLIN. MICROBIOL.
5. Matthew, M., R. W. Hedges, and J. T. Smith. 1979. Typesof beta-lactamase determined by plasmids in gram-negativebacteria. J. Bacteriol. 138:657-662.
6. Neu, H. C. 1980. Antibiotic inactivating enzymes and bac-terial resistance, p. 454-473. In V. Lorian (ed.), Antibioticsin laboratory medicine. The Williams & Wilkins Co.,Baltimore.
7. O'Callaghan, C. H., A. Morris, S. M. Kirby, and A. N.Shingler. 1972. Novel method for detection of beta-lacta-
mase by using a chromogenic cephalosporin substrate.Antimicrob. Agents Chemother. 1:283-288.
8. Spratt, B. G. 1975. Distinct penicillin binding proteinsinvolved in the division, elongation and shape of Escherich-ia coli K12. Proc. Natl. Acad. Sci. U.S.A. 72:2999-3003.
9. Sykes, R. B., and M. Matthew. 1976. The beta-lactamasesof Gram-negative bacteria and their role in resistance tobeta-lactam antibiotics. J. Antimicrob. Chemother. 2:115-157.
on July 18, 2020 by guesthttp://jcm
.asm.org/
Dow
nloaded from





























