Embed Size (px)
Citation preview

Research ArticleThe Plant-Derived Chalcone 22101584051015840-TrihydroxychalconeProvides Neuroprotection against Toll-Like Receptor 4Triggered Inflammation in Microglia
Manasi Jiwrajka1 Alexandra Phillips2 Matt Butler2
Miriam Rossi3 and Jennifer M Pocock2
1Mayne Medical School 288 Herston Road Brisbane QLD 4006 Australia2Department of Neuroinflammation University College London Institute of Neurology 1 Wakefield Street London WC1N 1PK UK3Department of Chemistry Vassar College 124 Raymond Avenue Poughkeepsie NY 12604-0484 USA
Correspondence should be addressed to Jennifer M Pocock jpocockuclacuk
Received 20 July 2015 Accepted 30 September 2015
Academic Editor Felipe Dal Pizzol
Copyright copy 2016 Manasi Jiwrajka et alThis is an open access article distributed under the Creative CommonsAttribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Chalcones are plant metabolites with potential for therapeutic exploitation as antioxidant anti-inflammatory and antiproliferativeagents Here we explored the neuroprotective effects of 22101584051015840-trihydroxychalcone (225THC) a potent antioxidant with radical-scavenging properties 225THCwas found to be a potent inhibitor of apoptosis in stimulated primary rat neuronal culturesThiswaslikelymediated by an anti-inflammatory effect onmicroglial cells since 225THC inhibited LPS-stimulatedTNF-120572 and IL-6 secretionfrom primary rat microglia andmodulated the cytokinechemokine profile of BV2microglial cells Additionally 225THC inhibitedLPS-evoked inducible nitric oxide synthase expression but did not influence endogenous superoxide generation Microglial flowcytometric analyses indicated the 225THC treatment induced a shift fromanM1-like phenotype to amore downregulatedmicroglialprofile Taken together these data suggest that the chalcone 22101584051015840-trihydroxychalcone canmodulate neuroinflammatory activationin brain-derived microglia and holds promise as a therapeutic in neuroinflammatory conditions
1 Introduction
Plants produce secondarymetabolites that protect them fromtoxins and insects Some of these plant metabolites such aschalcones have significant antioxidant anti-inflammatoryand antiproliferative properties in a range of cell types [1ndash7] Chalcones are similar to other known antioxidants suchas resveratrol curcumin and ubiquinone and are the naturalprecursors of flavonoids and isoflavonoids in higher plants[4 8ndash10] In plants chalcones protect against UV expo-sure pathogens and insects and their antioxidant and anti-inflammatory properties make them of increasing interest inthe treatment of human conditions such as cancer inflamma-tion tuberculosis and malaria [2 7 11]
Stress and injury to cells can cause the production offree radicals and the release of cytokines In the brain such
substances are produced by the activation of microglia thebrainrsquos resident phagocytes leading to neurotoxicity [12ndash14]During ageing neurodegeneration ischaemia brain injuryor other neuropathologies there is enhanced production offree radicals and cytokines increased apoptosis and reducedexpression of synaptic or growth proteins [15ndash18]
In the brain the chalcone isoliquiritigenin has anxiolyticeffects [19] whilst two chalconoids from the desert plantPulicaria incisa inhibited the production of reactive oxygenspecies (ROS) by astrocytes and prevented their oxidant-induced cell death [20] One plant-derived chalcone 22101584051015840-trihydroxychalcone (225THC) demonstrated strong antioxi-dant and radical-scavenging properties in L-6 myoblasts andTHP-1 humanmonocytes [21] However the neuroprotectiveeffects of this particular chalcone on cells of the CNS areunknown and the subject of the present study
Hindawi Publishing CorporationOxidative Medicine and Cellular LongevityVolume 2016 Article ID 6301712 10 pageshttpdxdoiorg10115520166301712
2 Oxidative Medicine and Cellular Longevity
2 Materials and Methods
21 Cell Culture
211 BV2 Microglia The BV2 mouse microglial cell line wasa kind gift from Dr Claudie Hooper Institute of PsychiatryKings College London and was originally obtained from theDepartment of Life Sciences National Cheng Kung Univer-sity Taiwan The cells were cultured in RPMI-1640 medium(Gibco Life Technologies) plus 5 foetal bovine serum(FBS) and 50UmLpenicillin and 50120583gmL streptomycin (allInvitrogen httpwwwinvitrogencom) at 37∘C at 5 CO
2
and at a density of 2times 104 per 13mmglass coverslips in 24-wellplates or 1 times 105 per 6-well plate forWestern blotting or FACSanalyses and rested overnight at 37∘C plus 5 CO
2 Before
use cell medium was replaced with RPMI plus 1 FBS
212 Primary Cultures of Microglia and Cerebellar GranuleCell Neurons Sprague Dawley rat pups (postnatal day 5)were bred and reared in-house from stock animals (CharlesRiver UK Kent UK) and were sacrificed in accordancewith Schedule 1 of the Animals Scientific Procedures Act(1986) UK for the culture of primary microglia and cere-bellar neurons Microglia were cultured as described pre-viously [22] Briefly cells were plated on 13mm glass cov-erslips in 24 well plates at 05 times 105 cellswell and main-tained in culture medium consisting of MEM (Invitrogenhttpwwwinvitrogencom) supplemented with 10 FBS20mM KCl 20mM D-glucose 2mM D-glucose 25mMNaHCO
4 50UmL penicillin 50 120583gmL streptomycin and
6 120583gmL ampicillin The microglia were maintained at 37∘C6 CO
2and used 1 day after plating Where possible
experiments were carried out using primary microglia butwhere assays required higher cell numbers BV2 microgliawere used We have previously compared responses of BV2microglia and primarymicroglia and have found not obviousdifferences in responses Cerebellar granule cells (CGCs)were cultured as described previously [22] The cells wereplated on 13mm glass coverslips in 24 well plates at a densityof 8 times 105coverslip and maintained in culture medium(MEM plus 10 FBS 20mM KCl 30mM D-glucose 2mML-glutamine 25mM NaHCO
4 50UmL penicillin 50120583M
streptomycin and 6 120583gmL ampicillin) that was later supple-mented with 20120583M cytosine furanoarabinoside (Ara-C) tocontrol glial proliferation The cells were maintained at 37∘C6 CO
2and used after 8 days in vitro (8 DIV)
22 Chalcone Treatment The chalcone 22101584051015840-trihydroxy-chalcone (225THC) was purchased from Indofine ChemicalCo (Hillsborough NJ USA at 97 purity) and was appliedto microglia and neurons to test for any inherent toxicity225THC was added at (final concentrations) 1120583M 5 120583M10 120583M 25 120583M 50120583M 100 120583M or 500 120583M in 100 sterileDMSO (Sigma D2650 hybri-MAX) as well as a solventcontrol of 100 DMSO and incubated for 24 h Livedeadassays were carried out using Hoechst 33342 for total cellnumber and propidium iodide (PI) for dead cells as pre-viously described [22] Cells were visualised and imagescaptured with a Zeiss Axioskop 2 fluorescence microscope
(Oberkochen Germany) and images were captured usingZeiss Axiovision Imaging System 48 software The numberof livedead cells was counted manually or using ImageJ software At least 5 fields per coverslip 3 coverslips per225THC concentration from 3 separate cell platings wereanalysed
23 225THC Treatment of Surveillant and InflammatoryMicroglia The effects of 225THC on microglial activationwere tested by treating primary microglia or BV2 microgliawith 225THC (1120583M 5 120583M 10 120583M 50 120583M 100 120583M and500120583M) with or without 2120583gmL of the Toll-like receptor 4(TLR4) agonist lipopolysaccharide (LPS) for 24ndash48 h Thisconcentration of LPS was used as this produces maximaliNOS expression as determined in previous experimentsand published papers from our laboratory [14 23] Thecells were subsequently analysed by livedead assay Westernblot and cell supernatants analysed for cytokine secre-tion as described To test for protective effects of 225THCagainst neuroinflammation-driven neurotoxicity CGCs weredirectly treated with 225THC (5 120583M 50 120583M and 500 120583M)and then either 2120583gmL LPS or 10 ngmL IFN-120574 to activateresident microglia in the cultures Following 24 h CGCcultures were analysed by Hoechst 33342 staining to assessnuclear morphology as described previously [22]
24 Western Blot of Inducible Nitric Oxide Synthase Expres-sion Cells were treated for Western blotting using standardtechniques followed by blot visualisationwith ECL Beta- (120573-)actin was used as a loading control in all gels and proteinbands analysed following densitometry with Image J soft-ware Primary antibodies used were anti-iNOS 1 2500overnight followed by 1 5000 HRP conjugated goat anti-rabbit for 1 h and anti-120573 actin 1 10000 overnight fol-lowed by HRP conjugated goat anti-mouse 1 200000 for1 h Goat anti-rabbit peroxidase secondary antibodywas fromSigma (Poole UK) donkey anti-goat peroxidase secondaryantibody was from GeneTex (Insight Biotech WembleyUK) goat anti-arginase-1 was from Santa Cruz Biotech(httpwwwscbtcom) and rabbit anti-inducible nitricoxide synthase (iNOS)was fromBDBiosciences (httpwwwbdbiosciencescom)
25 Dihydroethidium Fluorescence Imaging of SuperoxideGeneration The superoxide sensitive fluorescent dye dihy-droethidium (dHEth) was used to assess microglial super-oxide generation and its regulation by the chalcone Dihy-droethidium is oxidised to 2-hydroxyethidium (2-OH-E+)upon exposure to superoxide specifically correlating with ashift in fluorescence from blue to red which is detectableby fluorescence microscopy [24] and we have used thispreviously to assess superoxide generation in microglia [22]BV2 microglia were treated with 225THC LPS or 10 nMphorbol 12-myristate 13-acetate (PMA) (the latter as a positivecontrol for the generation of superoxide by NADPH activity[22 25]) for 24 h and then incubated with 5120583M dHEthfor 40min to identify superoxide-producing microglia asdescribed previously [22] plus 06 120583gmL Hoechst 33342 for40min was used to counterstain all nuclei for total cell
Oxidative Medicine and Cellular Longevity 3
number Microglia were imaged by fluorescence microscopyas above and superoxide-positive microglia as indicated byred nuclei were counted and expressed as a percentage oftotal cell number To ensure that the red fluorescence was dueto superoxide production experiments were carried out byincubation of the cells with 10 120583M apocynin [22]
26 ELISA of Secreted Cytokines by Microglia TNF-120572 orIL-1120573 concentrations in primary rat microglia cell culturesupernatants were quantified using Quantikine Rat TNF-120572or IL-1120573 Immunoassay kit according to the manufacturerrsquosinstructions (RampD Systems Abingdon UK) Cytokine con-centrations in cell supernatants were determined against astandard curve of TNF-120572 or IL-1120573 In addition a rangeof inflammatory mediators were analysed in BV2 cell cul-ture supernatants with an Inflammatory ELISA strip assayaccording to the manufacturerrsquos instructions (Signosis IncCaltag MedSystems Ltd Buckingham UK) In this case allvalues were presented as a percentage of control values Foreach condition the cytokine content in supernatants wasanalysed from three coverslips of microglia in three inde-pendent cell platings with each sample assayed in duplicateAll other chemicals and reagents were from Sigma (PooleUK)
27 Flow Cytometric Analysis of Microglial InflammatoryMarkers BV2 cells were treated with 1 5 or 10 120583M of225THC for 2 hours prior to the addition of 2 120583gmL LPSfor a further 48 hours Cells were harvested by washing inDulbeccorsquos phosphate buffered saline (PBS without Ca2+ andMg2+) for 20 minutes at 37∘C and then resuspended in coldPBS containing 05 BSA005 NaN
3to metabolically fix
the cells Cells were stained with anti-mouse CD11b-FITCCD40-FITC CD54-FITC and CD68-FITC or appropriateFITC-conjugated isotype control antibodies (Miltenyi BiotecUK) as per the manufacturerrsquos instructions and cell sur-face staining was assessed by flow cytometry (FACSCaliburrunning CellQuest Pro Becton Dickinson UK) and anal-ysed using Flowing software v251 Data are presented asaverage fold change in mean fluorescence intensity (MFI)versus untreated cells plusmn SEM from 4 independent experi-ments
28 Statistical Analysis Statistical analyses were performedusing one-way ANOVA with Tukey post hoc analysis wherecomparisons were made between treatments and controluntreated cells and also between treatment groups as indi-cated Where stated Studentrsquos 119905-test was used and all datawere from 3 separate experiments lowast119901 lt 005 lowastlowast119901 lt 001and lowastlowastlowast119901 lt 0005 compared with controls or as indicated infiguresWhere imagingwas performed aminimumof 3 fieldsof viewwere analysed from 3 coverslips per condition Imagesof fluorescent fields of cells orWestern blots are representativeof analysed data from3 independent experimentsThedensit-ometry forWesternswas performedusing Image Jmeasuringthe optical density (OD) of each band and normalising to the120573-actin obtained for that particular sample For the statisticalanalysis of these data each treatment was compared withcontrol untreated cells by ANOVA analysis and appropriate
comparisons were also made between treatment groups plusmninhibitors using Studentrsquos 119905-test
3 Results and Discussion
31 Effects of 225THC in Primary Rat Cerebellar GranuleCell Neuronal Cultures We initially investigated whether the225THCwas directly toxic to neurons by incubating primaryrat cerebellar granule cell (CGC) neuronal culture neuronswith concentrations of 5ndash500120583M 225THC for 24 h Thisrange of concentrations was assessed as previous studiesusing similar compounds have identified activity within thisrange [20 26 27] Microscopic livedead analyses revealedno significant toxicity in these cultures with DMSO solventcontrol or with 5 50 or 500120583M 225THC (Figure 1(a))when compared with untreated cells We next investigatedthe potential neuroprotective properties of 225THC in thesecultures (which we have shown previously contain 2ndash5microglia) [28 29] A range of 225THC concentrations werepreincubated with CGCs for 1 hour followed by stimulationwith 2120583gmL LPS or 10 ngmL IFN-120574 to induce an inflam-matory phenotype within the CGC cultures After 24 h theCGCs were imaged with Hoechst 33342 to visualise apop-totic nuclei within the cultures LPS or IFN-120574 significantlyincreased the percentage of apoptotic cells in the neuronalcultures above basal (Figure 1(b)) whilst pretreatment with225THC at 50 or 500 120583Msignificantly reduced the percentageof apoptotic cells in cultures treated with LPS or IFN-120574 SinceLPS and IFN-120574 are potent activators of microglia with noknown effects on cerebellar neurons and since incubationwith chalcone directly to these cultures in the absence ofinflammation did not enhance basal levels of neuronal sur-vival it is likely that suppression of the neuroinflammatoryand neurotoxic effects seen here are due to suppressionof microglial responses Thus we determined how theseeffects might be mediated One proposed mechanism forthe neurotoxicity observed is that activated microglia releaseproinflammatory cytokines that cause cytotoxicity to theneurons [15 30 31]
32 225THC Inhibits Proinflammatory Cytokine Secretion byLPS-Stimulated Primary Rat Microglial Cells As a prelimi-nary assessment we again investigated the potential cellulartoxicity of 225THC when added to primary rat microglialcultures We observed that in three separate experiments aspecific concentration of 50120583M 225THC appeared to inducesignificant cell loss fromcoverslips in both nonstimulated andLPS-stimulated cells whilst concentrations above and belowthis did not (Figure 2(a)) This was most likely due to lossof contact rather than toxicity as there was no increase in PIstaining DMSO controls did not affect cell numbers (data notshown)
We next assessed cytokine secretion inmicroglial cell cul-ture supernatants under basal (nonstimulating conditions)and following LPS stimulation (2120583gmL) in the presenceof 50 or 500120583M 225THC (Figure 2(b)) The basal secretionof TNF-120572 IL-6 and IL-10 was not significantly modulatedby 5 120583M or 500120583M 225THC whilst LPS-evoked TNF-120572 andIL-6 secretion but not IL-10 was significantly inhibited by
4 Oxidative Medicine and Cellular Longevity
0
100
Live
neu
rons
fiel
d (
)
5 50 500Basal
ns
50
DMSO[225THC] (120583M)
(a)
Apop
totic
neu
rons
()
LPS IFN-1205740
25
50
75
ns
ns
DMSO only
[225THC]
lowastlowastlowastlowastlowastlowast
5120583M
500120583M50120583M
(b)
Figure 1 225THC is nontoxic and neuroprotective in stimulated primary rat neuronal cultures (cerebellar granule cell neurons CGCs)Effects of 225THC on primary rat CGC neurons (a) Toxicity assay of 225THC on primary rat CGC neurons Data from three separateexperiments represent the mean number of live neurons (as assessed by livedead assay) present per field of view following incubation for24 h with different concentrations of 225THC (0 5 50 and 500 120583M) for 24 h DMSO is the solvent control basal is CGCs with nothing addedStatistical analysis was performed by ANOVA with Bonferroni post hoc analysis and revealed no toxicity at the concentrations tested whenall concentrations were compared with basal The DMSO solvent control was also not toxic (b) Direct treatment of CGC cultures for 24 hwith 225THC (0 5 50 and 500120583M) and 2 120583gmL LPS or 10 ngmL IFN-120574 to activate resident microglia in the cultures The percentage ofapoptotic cells following Hoechst 33342 visualisation of nuclear condensation is shown for each treatment Statistical analysis was performedby a two-way ANOVA with Bonferroni post hoc analysis lowast lowast lowast indicates 119901 lt 0001 ns not significant when compared as indicated or withthe appropriate basal control
5 120583M or 500120583M 225THC (Figures 2(bi) 2(bii) and 2(biii)resp)
33 225THC Modulates Inflammatory Cytokine and Chemo-kine Responses in Murine BV2Microglial Cells To investigatethe microglial aspects further we used the BV2 mousemicroglial cell line which is a well validated substitute forprimary microglia [32] Further examination of a lowerrange of 225THC concentrations (1120583M 5 120583M and 10 120583M)following a 48 h LPS stimulation revealed selective inhi-bition of LPS-induced TNF-120572 in line with our findingsin primary microglia (Figure 3(a)) In addition the basalsecretion of monocyte chemoattractant protein-1 (MCP-1)was also significantly inhibited (Figure 3(b)) We did notobservemodulation of basal or LPS-evoked secretion of IFN-R IL-1120572 IL-1120573 (data not shown) or of RANTES (Figure 3(c))or MIP (Figure 3(d)) These data suggest that 225THC hasan effect on the secretion of inflammatory cytokines suchas TNF-120572 IL-6 and MCP-1 and the reduction of thesecytokines may aid neuroprotection Interestingly in line withour findings Herencia et al [26] report that the chalconeused in their study inhibited TNF-120572 mouse air pouch tissuelevels from leukocytes stimulated by zymosan whilst Bano etal [33] reported suppression of LPS-evoked TNF-120572 secretionfrom a mouse macrophage line with a synthetic chalcone Inaddition we found 225THC inhibited the secretion of twoother cytokines IL-6 and MCP-1 IL-6 has well recognised
roles in neuroinflammation and promotes microgliosis [34]whilst MCP-1 (also known as CCL-2) acts as a chemoattrac-tant to recruit monocytes and macrophages into the brain[35] Interestingly recent work suggests different chalconesmay selectively inhibit the secretion of different groups ofcytokines and inflammatory mediators [27] We did not seesignificant modulation by 225THC of RANTES MIP or IL-10 Other chalcones have been shown to promote the produc-tion of neurotrophic factors such as GDNF from astrocytes[20] and this is worth future exploration for microglia
34 Modulation of iNOS Expression and ROS Generationby 225THC The expression of iNOS was assessed in BV2microglia under basal conditions and following LPS stimu-lation (2 120583gmL) for 48 h in the presence of 225THC (1120583M5 120583M or 10 120583M) No obvious effects were observed on theexpression of iNOS in nonstimulated microglia that is225THC did not induce expression of iNOS (Figure 4(a))However LPS-evoked stimulation of iNOS expression wassignificantly inhibited by 225THC at concentrations above5 120583M(Figures 4(a) and 4(b))We next investigatedmicroglialsuperoxide generation using microscopic fluorescence imag-ing (Figure 4(c)) No obvious effects of the 225THC wereobserved on the basal cell numbers expressing superoxide-positive cell nuclei and whilst the number of superoxide-positive cells was increased following LPS stimulation thiswas not significantly regulated by 225THC concentrations
Oxidative Medicine and Cellular Longevity 5
0
200
400
Aver
age n
umbe
r of
cells
fiel
dco
nditi
on
Basa
l 1
1005025105 1
10050105
LPS 2 (120583gmL) [225THC] (120583M) [225THC] (120583M)
Exp 1Exp 2Exp 3
(a)
20
0LPS
0
02
IL-6
(ng
mL)
IL-1
0 (n
gm
L)
IL-6
LPS
IL-10
0
02
BasalBasalBasal LPS
TNF-120572
TNF-120572
(ng
mL)
lowastlowast
lowast
lowastlowast
(biii)(bii)(bi)
5120583M500120583M
5120583M500120583M
5120583M500120583M
(b)
Figure 2 225THC is nontoxic and inhibits inflammatory cytokine responses in stimulated primary rat microglial cultures Effects of 225THCon primary rat microglial cells (a) Toxicity assay of 225THC on primary cultured rat microglia Data shown from three separate experimentsrepresent the mean number of live primary microglia (as assessed by livedead assay) present per field of view following incubation for24 h with different concentrations of 225THC (0 1 5 10 50 and 100 120583M) for 24 h with or without coactivation of the microglia with LPS(2120583gmL) (b) ELISA assessment of secreted cytokines in LPS (2 120583gmL) activated primary rat microglia cultures treated with 225THC at theconcentrations indicated Statistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005lowastlowast119901 lt 001 when compared with basla or as indicated
from 1 to 100 120583M(Figure 4(c)) or at 500120583M(data not shown)The positive control of NADPH oxidase activation PMA(10 ngmL) induced a significant increase in superoxide-positive cells in line with previous findings [22]
The inhibition of iNOS expression in microglia is inagreement with findings for another chalcone 4-dimethyl-amino-3101584041015840-dimethoxychalcone on mouse macrophagesstimulated with zymosan [22] This chalcone inhibited iNOSexpression with a similar concentration (sim10 120583M) to the chal-cone used in the present study Herencia et al [26] proposedthat the effect on iNOS expression was via upstreammodula-tion of NADPH oxidase-mediated intracellular signalling
Since we did not detect modulation of microglial superoxidegeneration by 225THC it seems unlikely that this particularchalcone possesses this mechanism of action Other chal-cones have also been reported to modulate nitric oxide pro-duction or iNOS expression following LPS stimulation of amouse macrophage cell line [36] the BV2 microglial cell line[27] and a rat microglial cell line [37] within similar com-pound concentration ranges of 1ndash50 120583MThe exact signallingpathways inhibited by different chalcones may be diverseas some studies suggest modulation of nuclear factor-120581BMAPK or c-Jun pathways and others suggest STAT1 path-ways may be involved [26 37]
6 Oxidative Medicine and Cellular Longevity
60
220
Abov
e con
trol (
)
Basal LPS
ns
ns
lowastlowast
lowast
lowast lowast
1120583M5120583M10120583M
TNF-120572
(a)
MCP-1
60
220
LPS
ns
ns
lowastlowast
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(b)
Rantes
60
280
LPS
ns
lowast
lowastlowast
lowast
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(c)
LPS
MIP
80
170
ns
ns
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(d)
Figure 3 225THC modulates inflammatory cytokine and chemokine responses in murine BV2 microglial cells 225THC-modulation ofcytokinechemokine secretion by the BV2 microglial cell line ELISA assessment of cytokine ((a) TNF-120572) and chemokine ((b) MCP-1 (c)RANTES (d) MIP) secretion by BV2 microglial cells following stimulation with LPS (2 120583gmL) for 24 h in the presence of 225THC at theconcentrations indicated Statistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005lowastlowast119901 lt 001 ns not significant when compared with basal or as indicated
35 Modulation ofMicroglial Inflammatory Phenotype UponLPS stimulationmicroglia adopt an inflammatory phenotypewhich is characterised by enhanced cell surface expression ofkey adhesion (CD11b CD54) and costimulatory molecules(CD40) We investigated the effects of 225THC treatment
on BV2 cell surface expression of CD11b CD40 CD54 andCD68 in basal and LPS-treated BV2 microglia (Figure 5)Flow cytometric analysis (FACs) of marker mean fluores-cence intensity (MFI) revealed no significant changes in basalCD40 CD54 CD68 or CD11b expression levels following
Oxidative Medicine and Cellular Longevity 7
iNOS
Basal 1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M)[225THC] (120583M)
120573-actin
(a)
0
07
1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
Den
sitom
etry
of i
NO
S120573
-act
in
lowast lowast
(b)
Supe
roxi
de-p
ositi
ve ce
lls(
tota
l)
50
01 5 10 100 1 5 10 100
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
lowastlowast lowastlowastlowastlowast lowastlowast
PMA10ngmL
lowast
lowast
(c)
1120583M LPS + 1120583M
10120583M
100120583M
LPS + 10120583M
LPS + 100120583M
Control LPS
(d)
Figure 4 225THC inhibits the LPS-induced expression of iNOS but not NADPH-driven superoxide production in BV2 microgliaModulation of BV2 iNOS expression but not superoxide production by 225THC Representative Western blots of iNOS expression and 120573-actin as a loading control (a) and densitometry (b) of three separate blots for BV2microglia exposed to 225THC at the stated concentrations plusmnLPS (2120583gmL) for 24 h Statistical analysis was performed by a two-way ANOVAwith Bonferroni post hoc analysis lowast indicates 119901 lt 005whencompared with basal or as indicated (c) Superoxide generation in BV2 microglia measured by dihydroethidium fluorescence following 24 hexposure to 225THC at the concentrations indicated plusmn LPS (2120583gmL) The phorbol PMA was used as a positive activator of superoxidegeneration (d) Representative fields of BV2 microglia generating intracellular superoxide following 24 h exposure to 225THC +minus LPSStatistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005 lowastlowast indicates 119901 lt 001when compared with basal or as indicated
225THC treatment for 48 h (data not shown) However fol-lowing stimulation with 2 120583gmL LPS CD40 (Figure 5(bii))CD54 (Figure 5(cii)) and CD68 (Figure 5(dii)) were signif-icantly upregulated and these changes in expression wereabrogated in the presence of 5 or 10 120583M 225THC CD40appeared to be the most potently regulatedmarker with both5 and 10 120583M 225THC completely inhibiting LPS-inducedupregulation (Figure 5(bii)) while the treatment had moremoderate effects on CD54 and CD68 marker expression As
in the basal state 225THC had no effect on CD11b expressionby LPS-stimulated microglia (Figure 5(aii))
Overall these data suggest that 225THC targets thesignalling pathways downstream of LPS binding and abro-gates the expression of genes such as TNF-120572 iNOS andCD40 Since these are all reported NF-120581B driven genesfuture work could investigate this further to identify theexact pathways modified by 225THC Abrogating this geneexpression produced a less neurotoxic microglial phenotype
8 Oxidative Medicine and Cellular Longevity
CD11b
(ai) (aii)
[225THC] (120583M)
FL1-H
CD11b
Basal LPS 1 5 10
111009080706050403020100
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(a)
CD40
(bi) (bii)
[225THC] (120583M)
FL1-H
CD40
Basal LPS 1 5 10
125
100
075
050
025
000
175
150
lowastlowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(b)
(ci) (cii)
[225THC] (120583M)
FL1-H
CD54 (ICAM-1)CD54 (ICAM-1)
Basal LPS 1 5 10
400
300
200
100
000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(c)
CD68
(di) (dii)
[225THC] (120583M)FL1-H
CD68
Basal LPS 1 5 10
160140120100080060040020000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(d)
Figure 5 225THC treatmentmodulates inflammatory cell surfacemarker expression in LPS-treated BV2microglia Flow cytometric analysisof 225THC-treated BV2 microglia following LPS stimulation (a) Representative histogram plots of BV2 cell surface CD11b CD40 CD54and CD68 expression following 2 120583gmL LPS-treatment for 48 hours (unfilled histograms) versus isotype control (black-filled histograms)and in the presence of 10 120583M 225THC (grey-filled histograms) (b) Summary data showing fold changes in CD11b CD40 CD54 or CD68expression by mean fluorescence intensity (MFI) from 4 independent experiments Data are mean values plusmn SEM where lowast indicates 119901 lt 005by Studentrsquos 119905-test
Oxidative Medicine and Cellular Longevity 9
which is likely to be beneficial for a number of neurodegen-erative diseases in which aberrant microglial inflammatoryreactivity is implicated Future work could pinpoint furtherthe microglial phenotype favoured by 225THC exposure andexplore the genotype of the microglia
Abbreviations
ANOVA Analysis of varianceDPPH 22-Diphenyl-1-picrylhydrazyldHEth DihydroethidiumCGCs Cerebellar granule neuronsDIV Days in vitroDAPI 410158406-Diamidino-2-phenylindoleDMSO DimethylsulphoxideECL ElectrochemiluminescenceELISA Enzyme-linked immunosorbent assayGABAA 120574-Aminobutyric acidIFN-120574 Interferon-120574IL-1120573 Interleukin-1120573IL-6 Interleukin-6IL-10 Interleukin-10iNOS Inducible nitric oxide synthaseLPS LipopolysaccharideMCP-1 CCL2monocyte chemoattractant
protein-1MEM Modified Eaglersquos mediumMIP Macrophage inhibitory proteinMCP-1 Monocyte chemoattractant protein-1NGS Normal goat serumPBS Phosphate buffered salinePFA Paraformaldehyde2-OH-E+ 2-HydroxyethidiumRANTES CCL5regulation on activation normal T
cell expressed and secretedROS Reactive oxygen speciesTNF-120572 Tumour necrosis factor-120572
Conflict of Interests
All authors declare no conflict of interests
Acknowledgments
Manasi Jiwrajka would like to thank Dr Kathleen Susman forher guidance This research was supported by the Universityof Queensland Advantage Grant Howard Hughes MedicalFoundation Grant 52006322 to Vassar College the ShermanFairchild Foundation and Vassar College and by generalfunds to the Pocock lab
References
[1] S Ducki R Forrest J A Hadfield et al ldquoPotent antimitotic andcell growth inhibitory properties of substituted chalconesrdquo Bio-organic and Medicinal Chemistry Letters vol 8 no 9 pp 1051ndash1056 1998
[2] M L F Ferreyra S P Rius and P Casati ldquoFlavonoids biosyn-thesis biological functions and biotechnological applicationsrdquoFrontiers in Plant Science vol 3 article 222 15 pages 2012
[3] G E-D A A Abuo-Rahma M Abdel-Aziz M A E Mouradand H H Farag ldquoSynthesis anti-inflammatory activity andulcerogenic liability of novel nitric oxide donatingchalconehybridsrdquo Bioorganic and Medicinal Chemistry vol 20 no 1 pp195ndash206 2012
[4] N K Sahu S S Balbhadra J Choudhary and D V KohlildquoExploring pharmacological significance of chalcone scaffoldrdquoCurrent Medicinal Chemistry vol 19 no 2 pp 209ndash225 2012
[5] H Wei X Zhang G Wu et al ldquoChalcone derivatives from thefern Cyclosorus parasiticus and their anti-proliferative activityrdquoFood and Chemical Toxicology vol 60 pp 147ndash152 2013
[6] C W Mai M Yaeghoobi N Abd-Rahman Y B Kang andM R Pichika ldquoChalcones with electron-withdrawing and ele-ctron-donating substituents anticancer activity against TRAILresistant cancer cells structure-activity relationship analysisand regulation of apoptotic proteinsrdquo European Journal ofMedicinal Chemistry vol 77 pp 378ndash387 2014
[7] Z Nowakowska ldquoA review of anti-infective and anti-inflam-matory chalconesrdquo European Journal of Medicinal Chemistryvol 42 no 2 pp 125ndash137 2007
[8] Y-M Lin Y Zhou M T Flavin L-M Zhou W Nie and F-CChen ldquoChalcones and flavonoids as anti-tuberculosis agentsrdquoBioorganic and Medicinal Chemistry vol 10 no 8 pp 2795ndash2802 2002
[9] JMojzis L Varinska GMojzisova I Kostova and LMirossayldquoAnti-angiogenic effects of flavonoids and chalconesrdquo Pharma-cological Research vol 57 no 4 pp 259ndash265 2008
[10] F Dal Piaz A Braca M A Belisario and N De TommasildquoThioredoxin systemmodulation by plant and fungal secondarymetabolitesrdquo Current Medicinal Chemistry vol 17 no 5 pp479ndash494 2010
[11] N Yadav S K Dixit A Bhattacharya et al ldquoAntimalarial activ-ity of newly synthesized chalcone derivatives in vitrordquo Chem-ical Biology and Drug Design vol 80 no 2 pp 340ndash347 2012
[12] F V DeFeudis ldquoNovel therapy for Alzheimerrsquos diseaserdquo Neuro-biology of Aging vol 10 no 5 pp 467ndash468 1989
[13] Y Nomura and Y Kitamura ldquoInducible nitric oxide synthasein glial cellsrdquo Neuroscience Research vol 18 no 2 pp 103ndash1071993
[14] P J Kingham M L Cuzner and J M Pocock ldquoApoptotic path-ways mobilized in microglia and neurones as a consequenceof chromogranin A-induced microglial activationrdquo Journal ofNeurochemistry vol 73 no 2 pp 538ndash547 1999
[15] S Singh S Swarnkar P Goswami and C Nath ldquoAstrocytes andmicroglia responses to neuropathological conditionsrdquo Inter-national Journal of Neuroscience vol 121 no 11 pp 589ndash5972011
[16] Y Y Deng J Lu E-A Ling and C Kaur ldquoRole of microglia inthe process of inflammation in the hypoxic developing brainrdquoFrontiers in Bioscience (Scholar Edition) vol 3 no 3 pp 884ndash900 2011
[17] M Dumont and M F Beal ldquoNeuroprotective strategies involv-ing ROS in Alzheimer diseaserdquo Free Radical Biology andMedicine vol 51 no 5 pp 1014ndash1026 2011
[18] AD Kraft andG JeanHarry ldquoFeatures ofmicroglia and neuro-inflammation relevant to environmental exposure and neuro-toxicityrdquo International Journal of Environmental Research andPublic Health vol 8 no 7 pp 2980ndash3018 2011
[19] S Cho S Kim Z Jin et al ldquoIsoliquiritigenin a chalcone com-pound is a positive allosteric modulator of GABA A recep-tors and shows hypnotic effectsrdquo Biochemical and BiophysicalResearch Communications vol 413 no 4 pp 637ndash642 2011
10 Oxidative Medicine and Cellular Longevity
[20] A Elmann A Telerman H Erlank et al ldquoProtective and anti-oxidant effects of a chalconoid from Pulicaria incisa on brainastrocytesrdquoOxidativeMedicine andCellular Longevity vol 2013Article ID 694398 10 pages 2013
[21] M Rossi F Caruso E J Crespi et al ldquoProbing antioxidantactivity of 21015840-hydroxychalcones crystal and molecular struc-tures in vitro antiproliferative studies and in vivo effects onglucose regulationrdquo Biochimie vol 95 no 10 pp 1954ndash19632013
[22] E LMead AMosley S Eaton L Dobson S J Heales and JMPocock ldquoMicroglial neurotransmitter receptors trigger super-oxide production in microglia consequences for microglial-neuronal interactionsrdquo Journal of Neurochemistry vol 121 no2 pp 287ndash301 2012
[23] C M Davenport I G Sevastou C Hooper and J M PocockldquoInhibiting p53 pathways in microglia attenuates microglial-evoked neurotoxicity following exposure to Alzheimer pep-tidesrdquo Journal of Neurochemistry vol 112 no 2 pp 552ndash5632010
[24] N C Garbett N B Hammond and D E Graves ldquoInfluence ofthe amino substituents in the interaction of ethidium bromidewith DNArdquo Biophysical Journal vol 87 no 6 pp 3974ndash39812004
[25] A Karlsson J B Nixon and L C McPhail ldquoPhorbol myristateacetate induces neutrophilNADPH-oxidase activity by two sep-arate signal transduction pathways dependent or independentof phosphatidylinositol 3-kinaserdquo Journal of Leukocyte Biologyvol 67 no 3 pp 396ndash404 2000
[26] F Herencia M L Ferrandiz A Ubeda et al ldquo4-Dimethyl-amino-3101584041015840-dimethoxychalcone down-regulates iNOS expres-sion and exerts anti-inflammatory effectsrdquo Free Radical Biologyamp Medicine vol 30 no 1 pp 43ndash50 2001
[27] N Mateeva M Gangapuram E Mazzio S Eyunni K FSoliman and K K Redda ldquoBiological evaluation of syntheticchalcone and flavone derivatives as anti-inflammatory agentsrdquoMedicinal Chemistry Research vol 24 no 4 pp 1672ndash16802015
[28] S C Morgan D L Taylor and J M Pocock ldquoMicroglia releaseactivators of neuronal proliferation mediated by activationof mitogen-activated protein kinase phosphatidylinositol-3-kinaseAkt and delta-Notch signalling cascadesrdquo Journal ofNeurochemistry vol 90 no 1 pp 89ndash101 2004
[29] J Jebelli T Piers and J Pocock ldquoSelective depletion of micro-glia from cerebellar granule cell cultures using L-leucinemethylesterrdquoThe Journal of Visualized Experiments no 101 Article IDe52983 2015
[30] R Von Bernhardi J E Tichauer and J Eugenın ldquoAging-dependent changes of microglial cells and their relevance forneurodegenerative disordersrdquo Journal of Neurochemistry vol112 no 5 pp 1099ndash1114 2010
[31] C Schwab and P L McGeer ldquoInflammatory aspects ofAlzheimer disease and other neurodegenerative disordersrdquoJournal of Alzheimerrsquos Disease vol 13 no 4 pp 359ndash369 2008
[32] A Henn S Lund M Hedtjarn A Schrattenholz P Porzgenand M Leist ldquoThe suitability of BV2 cells as alternative modelsystem for primary microglia cultures or for animal experi-ments examining brain inflammationrdquo Altex vol 26 no 2 pp83ndash94 2009
[33] S Bano K Javed S Ahmad et al ldquoSynthesis of some novel chal-cones flavanones and flavones and evaluation of their anti-inflammatory activityrdquo European Journal of Medicinal Chem-istry vol 65 pp 51ndash59 2013
[34] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[35] S L Deshmane S Kremlev S Amini and B E Sawaya ldquoMono-cyte chemoattractant protein-1 (MCP-1) an overviewrdquo Journalof Interferon and Cytokine Research vol 29 no 6 pp 313ndash3262009
[36] J Rojas J N Domınguez J E Charris G Lobo M Paya andM L Ferrandiz ldquoSynthesis and inhibitory activity of dimethyl-amino-chalcone derivatives on the induction of nitric oxidesynthaserdquo European Journal of Medicinal Chemistry vol 37 no8 pp 699ndash705 2002
[37] H Hara Y Nakamura M Ninomiya et al ldquoInhibitory effectsof chalcone glycosides isolated from Brassica rapa L lsquohidabenirsquoand their synthetic derivatives on LPS-induced NO productionin microgliardquo Bioorganic amp Medicinal Chemistry vol 19 no 18pp 5559ndash5568 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Oxidative Medicine and Cellular Longevity
2 Materials and Methods
21 Cell Culture
211 BV2 Microglia The BV2 mouse microglial cell line wasa kind gift from Dr Claudie Hooper Institute of PsychiatryKings College London and was originally obtained from theDepartment of Life Sciences National Cheng Kung Univer-sity Taiwan The cells were cultured in RPMI-1640 medium(Gibco Life Technologies) plus 5 foetal bovine serum(FBS) and 50UmLpenicillin and 50120583gmL streptomycin (allInvitrogen httpwwwinvitrogencom) at 37∘C at 5 CO
2
and at a density of 2times 104 per 13mmglass coverslips in 24-wellplates or 1 times 105 per 6-well plate forWestern blotting or FACSanalyses and rested overnight at 37∘C plus 5 CO
2 Before
use cell medium was replaced with RPMI plus 1 FBS
212 Primary Cultures of Microglia and Cerebellar GranuleCell Neurons Sprague Dawley rat pups (postnatal day 5)were bred and reared in-house from stock animals (CharlesRiver UK Kent UK) and were sacrificed in accordancewith Schedule 1 of the Animals Scientific Procedures Act(1986) UK for the culture of primary microglia and cere-bellar neurons Microglia were cultured as described pre-viously [22] Briefly cells were plated on 13mm glass cov-erslips in 24 well plates at 05 times 105 cellswell and main-tained in culture medium consisting of MEM (Invitrogenhttpwwwinvitrogencom) supplemented with 10 FBS20mM KCl 20mM D-glucose 2mM D-glucose 25mMNaHCO
4 50UmL penicillin 50 120583gmL streptomycin and
6 120583gmL ampicillin The microglia were maintained at 37∘C6 CO
2and used 1 day after plating Where possible
experiments were carried out using primary microglia butwhere assays required higher cell numbers BV2 microgliawere used We have previously compared responses of BV2microglia and primarymicroglia and have found not obviousdifferences in responses Cerebellar granule cells (CGCs)were cultured as described previously [22] The cells wereplated on 13mm glass coverslips in 24 well plates at a densityof 8 times 105coverslip and maintained in culture medium(MEM plus 10 FBS 20mM KCl 30mM D-glucose 2mML-glutamine 25mM NaHCO
4 50UmL penicillin 50120583M
streptomycin and 6 120583gmL ampicillin) that was later supple-mented with 20120583M cytosine furanoarabinoside (Ara-C) tocontrol glial proliferation The cells were maintained at 37∘C6 CO
2and used after 8 days in vitro (8 DIV)
22 Chalcone Treatment The chalcone 22101584051015840-trihydroxy-chalcone (225THC) was purchased from Indofine ChemicalCo (Hillsborough NJ USA at 97 purity) and was appliedto microglia and neurons to test for any inherent toxicity225THC was added at (final concentrations) 1120583M 5 120583M10 120583M 25 120583M 50120583M 100 120583M or 500 120583M in 100 sterileDMSO (Sigma D2650 hybri-MAX) as well as a solventcontrol of 100 DMSO and incubated for 24 h Livedeadassays were carried out using Hoechst 33342 for total cellnumber and propidium iodide (PI) for dead cells as pre-viously described [22] Cells were visualised and imagescaptured with a Zeiss Axioskop 2 fluorescence microscope
(Oberkochen Germany) and images were captured usingZeiss Axiovision Imaging System 48 software The numberof livedead cells was counted manually or using ImageJ software At least 5 fields per coverslip 3 coverslips per225THC concentration from 3 separate cell platings wereanalysed
23 225THC Treatment of Surveillant and InflammatoryMicroglia The effects of 225THC on microglial activationwere tested by treating primary microglia or BV2 microgliawith 225THC (1120583M 5 120583M 10 120583M 50 120583M 100 120583M and500120583M) with or without 2120583gmL of the Toll-like receptor 4(TLR4) agonist lipopolysaccharide (LPS) for 24ndash48 h Thisconcentration of LPS was used as this produces maximaliNOS expression as determined in previous experimentsand published papers from our laboratory [14 23] Thecells were subsequently analysed by livedead assay Westernblot and cell supernatants analysed for cytokine secre-tion as described To test for protective effects of 225THCagainst neuroinflammation-driven neurotoxicity CGCs weredirectly treated with 225THC (5 120583M 50 120583M and 500 120583M)and then either 2120583gmL LPS or 10 ngmL IFN-120574 to activateresident microglia in the cultures Following 24 h CGCcultures were analysed by Hoechst 33342 staining to assessnuclear morphology as described previously [22]
24 Western Blot of Inducible Nitric Oxide Synthase Expres-sion Cells were treated for Western blotting using standardtechniques followed by blot visualisationwith ECL Beta- (120573-)actin was used as a loading control in all gels and proteinbands analysed following densitometry with Image J soft-ware Primary antibodies used were anti-iNOS 1 2500overnight followed by 1 5000 HRP conjugated goat anti-rabbit for 1 h and anti-120573 actin 1 10000 overnight fol-lowed by HRP conjugated goat anti-mouse 1 200000 for1 h Goat anti-rabbit peroxidase secondary antibodywas fromSigma (Poole UK) donkey anti-goat peroxidase secondaryantibody was from GeneTex (Insight Biotech WembleyUK) goat anti-arginase-1 was from Santa Cruz Biotech(httpwwwscbtcom) and rabbit anti-inducible nitricoxide synthase (iNOS)was fromBDBiosciences (httpwwwbdbiosciencescom)
25 Dihydroethidium Fluorescence Imaging of SuperoxideGeneration The superoxide sensitive fluorescent dye dihy-droethidium (dHEth) was used to assess microglial super-oxide generation and its regulation by the chalcone Dihy-droethidium is oxidised to 2-hydroxyethidium (2-OH-E+)upon exposure to superoxide specifically correlating with ashift in fluorescence from blue to red which is detectableby fluorescence microscopy [24] and we have used thispreviously to assess superoxide generation in microglia [22]BV2 microglia were treated with 225THC LPS or 10 nMphorbol 12-myristate 13-acetate (PMA) (the latter as a positivecontrol for the generation of superoxide by NADPH activity[22 25]) for 24 h and then incubated with 5120583M dHEthfor 40min to identify superoxide-producing microglia asdescribed previously [22] plus 06 120583gmL Hoechst 33342 for40min was used to counterstain all nuclei for total cell
Oxidative Medicine and Cellular Longevity 3
number Microglia were imaged by fluorescence microscopyas above and superoxide-positive microglia as indicated byred nuclei were counted and expressed as a percentage oftotal cell number To ensure that the red fluorescence was dueto superoxide production experiments were carried out byincubation of the cells with 10 120583M apocynin [22]
26 ELISA of Secreted Cytokines by Microglia TNF-120572 orIL-1120573 concentrations in primary rat microglia cell culturesupernatants were quantified using Quantikine Rat TNF-120572or IL-1120573 Immunoassay kit according to the manufacturerrsquosinstructions (RampD Systems Abingdon UK) Cytokine con-centrations in cell supernatants were determined against astandard curve of TNF-120572 or IL-1120573 In addition a rangeof inflammatory mediators were analysed in BV2 cell cul-ture supernatants with an Inflammatory ELISA strip assayaccording to the manufacturerrsquos instructions (Signosis IncCaltag MedSystems Ltd Buckingham UK) In this case allvalues were presented as a percentage of control values Foreach condition the cytokine content in supernatants wasanalysed from three coverslips of microglia in three inde-pendent cell platings with each sample assayed in duplicateAll other chemicals and reagents were from Sigma (PooleUK)
27 Flow Cytometric Analysis of Microglial InflammatoryMarkers BV2 cells were treated with 1 5 or 10 120583M of225THC for 2 hours prior to the addition of 2 120583gmL LPSfor a further 48 hours Cells were harvested by washing inDulbeccorsquos phosphate buffered saline (PBS without Ca2+ andMg2+) for 20 minutes at 37∘C and then resuspended in coldPBS containing 05 BSA005 NaN
3to metabolically fix
the cells Cells were stained with anti-mouse CD11b-FITCCD40-FITC CD54-FITC and CD68-FITC or appropriateFITC-conjugated isotype control antibodies (Miltenyi BiotecUK) as per the manufacturerrsquos instructions and cell sur-face staining was assessed by flow cytometry (FACSCaliburrunning CellQuest Pro Becton Dickinson UK) and anal-ysed using Flowing software v251 Data are presented asaverage fold change in mean fluorescence intensity (MFI)versus untreated cells plusmn SEM from 4 independent experi-ments
28 Statistical Analysis Statistical analyses were performedusing one-way ANOVA with Tukey post hoc analysis wherecomparisons were made between treatments and controluntreated cells and also between treatment groups as indi-cated Where stated Studentrsquos 119905-test was used and all datawere from 3 separate experiments lowast119901 lt 005 lowastlowast119901 lt 001and lowastlowastlowast119901 lt 0005 compared with controls or as indicated infiguresWhere imagingwas performed aminimumof 3 fieldsof viewwere analysed from 3 coverslips per condition Imagesof fluorescent fields of cells orWestern blots are representativeof analysed data from3 independent experimentsThedensit-ometry forWesternswas performedusing Image Jmeasuringthe optical density (OD) of each band and normalising to the120573-actin obtained for that particular sample For the statisticalanalysis of these data each treatment was compared withcontrol untreated cells by ANOVA analysis and appropriate
comparisons were also made between treatment groups plusmninhibitors using Studentrsquos 119905-test
3 Results and Discussion
31 Effects of 225THC in Primary Rat Cerebellar GranuleCell Neuronal Cultures We initially investigated whether the225THCwas directly toxic to neurons by incubating primaryrat cerebellar granule cell (CGC) neuronal culture neuronswith concentrations of 5ndash500120583M 225THC for 24 h Thisrange of concentrations was assessed as previous studiesusing similar compounds have identified activity within thisrange [20 26 27] Microscopic livedead analyses revealedno significant toxicity in these cultures with DMSO solventcontrol or with 5 50 or 500120583M 225THC (Figure 1(a))when compared with untreated cells We next investigatedthe potential neuroprotective properties of 225THC in thesecultures (which we have shown previously contain 2ndash5microglia) [28 29] A range of 225THC concentrations werepreincubated with CGCs for 1 hour followed by stimulationwith 2120583gmL LPS or 10 ngmL IFN-120574 to induce an inflam-matory phenotype within the CGC cultures After 24 h theCGCs were imaged with Hoechst 33342 to visualise apop-totic nuclei within the cultures LPS or IFN-120574 significantlyincreased the percentage of apoptotic cells in the neuronalcultures above basal (Figure 1(b)) whilst pretreatment with225THC at 50 or 500 120583Msignificantly reduced the percentageof apoptotic cells in cultures treated with LPS or IFN-120574 SinceLPS and IFN-120574 are potent activators of microglia with noknown effects on cerebellar neurons and since incubationwith chalcone directly to these cultures in the absence ofinflammation did not enhance basal levels of neuronal sur-vival it is likely that suppression of the neuroinflammatoryand neurotoxic effects seen here are due to suppressionof microglial responses Thus we determined how theseeffects might be mediated One proposed mechanism forthe neurotoxicity observed is that activated microglia releaseproinflammatory cytokines that cause cytotoxicity to theneurons [15 30 31]
32 225THC Inhibits Proinflammatory Cytokine Secretion byLPS-Stimulated Primary Rat Microglial Cells As a prelimi-nary assessment we again investigated the potential cellulartoxicity of 225THC when added to primary rat microglialcultures We observed that in three separate experiments aspecific concentration of 50120583M 225THC appeared to inducesignificant cell loss fromcoverslips in both nonstimulated andLPS-stimulated cells whilst concentrations above and belowthis did not (Figure 2(a)) This was most likely due to lossof contact rather than toxicity as there was no increase in PIstaining DMSO controls did not affect cell numbers (data notshown)
We next assessed cytokine secretion inmicroglial cell cul-ture supernatants under basal (nonstimulating conditions)and following LPS stimulation (2120583gmL) in the presenceof 50 or 500120583M 225THC (Figure 2(b)) The basal secretionof TNF-120572 IL-6 and IL-10 was not significantly modulatedby 5 120583M or 500120583M 225THC whilst LPS-evoked TNF-120572 andIL-6 secretion but not IL-10 was significantly inhibited by
4 Oxidative Medicine and Cellular Longevity
0
100
Live
neu
rons
fiel
d (
)
5 50 500Basal
ns
50
DMSO[225THC] (120583M)
(a)
Apop
totic
neu
rons
()
LPS IFN-1205740
25
50
75
ns
ns
DMSO only
[225THC]
lowastlowastlowastlowastlowastlowast
5120583M
500120583M50120583M
(b)
Figure 1 225THC is nontoxic and neuroprotective in stimulated primary rat neuronal cultures (cerebellar granule cell neurons CGCs)Effects of 225THC on primary rat CGC neurons (a) Toxicity assay of 225THC on primary rat CGC neurons Data from three separateexperiments represent the mean number of live neurons (as assessed by livedead assay) present per field of view following incubation for24 h with different concentrations of 225THC (0 5 50 and 500 120583M) for 24 h DMSO is the solvent control basal is CGCs with nothing addedStatistical analysis was performed by ANOVA with Bonferroni post hoc analysis and revealed no toxicity at the concentrations tested whenall concentrations were compared with basal The DMSO solvent control was also not toxic (b) Direct treatment of CGC cultures for 24 hwith 225THC (0 5 50 and 500120583M) and 2 120583gmL LPS or 10 ngmL IFN-120574 to activate resident microglia in the cultures The percentage ofapoptotic cells following Hoechst 33342 visualisation of nuclear condensation is shown for each treatment Statistical analysis was performedby a two-way ANOVA with Bonferroni post hoc analysis lowast lowast lowast indicates 119901 lt 0001 ns not significant when compared as indicated or withthe appropriate basal control
5 120583M or 500120583M 225THC (Figures 2(bi) 2(bii) and 2(biii)resp)
33 225THC Modulates Inflammatory Cytokine and Chemo-kine Responses in Murine BV2Microglial Cells To investigatethe microglial aspects further we used the BV2 mousemicroglial cell line which is a well validated substitute forprimary microglia [32] Further examination of a lowerrange of 225THC concentrations (1120583M 5 120583M and 10 120583M)following a 48 h LPS stimulation revealed selective inhi-bition of LPS-induced TNF-120572 in line with our findingsin primary microglia (Figure 3(a)) In addition the basalsecretion of monocyte chemoattractant protein-1 (MCP-1)was also significantly inhibited (Figure 3(b)) We did notobservemodulation of basal or LPS-evoked secretion of IFN-R IL-1120572 IL-1120573 (data not shown) or of RANTES (Figure 3(c))or MIP (Figure 3(d)) These data suggest that 225THC hasan effect on the secretion of inflammatory cytokines suchas TNF-120572 IL-6 and MCP-1 and the reduction of thesecytokines may aid neuroprotection Interestingly in line withour findings Herencia et al [26] report that the chalconeused in their study inhibited TNF-120572 mouse air pouch tissuelevels from leukocytes stimulated by zymosan whilst Bano etal [33] reported suppression of LPS-evoked TNF-120572 secretionfrom a mouse macrophage line with a synthetic chalcone Inaddition we found 225THC inhibited the secretion of twoother cytokines IL-6 and MCP-1 IL-6 has well recognised
roles in neuroinflammation and promotes microgliosis [34]whilst MCP-1 (also known as CCL-2) acts as a chemoattrac-tant to recruit monocytes and macrophages into the brain[35] Interestingly recent work suggests different chalconesmay selectively inhibit the secretion of different groups ofcytokines and inflammatory mediators [27] We did not seesignificant modulation by 225THC of RANTES MIP or IL-10 Other chalcones have been shown to promote the produc-tion of neurotrophic factors such as GDNF from astrocytes[20] and this is worth future exploration for microglia
34 Modulation of iNOS Expression and ROS Generationby 225THC The expression of iNOS was assessed in BV2microglia under basal conditions and following LPS stimu-lation (2 120583gmL) for 48 h in the presence of 225THC (1120583M5 120583M or 10 120583M) No obvious effects were observed on theexpression of iNOS in nonstimulated microglia that is225THC did not induce expression of iNOS (Figure 4(a))However LPS-evoked stimulation of iNOS expression wassignificantly inhibited by 225THC at concentrations above5 120583M(Figures 4(a) and 4(b))We next investigatedmicroglialsuperoxide generation using microscopic fluorescence imag-ing (Figure 4(c)) No obvious effects of the 225THC wereobserved on the basal cell numbers expressing superoxide-positive cell nuclei and whilst the number of superoxide-positive cells was increased following LPS stimulation thiswas not significantly regulated by 225THC concentrations
Oxidative Medicine and Cellular Longevity 5
0
200
400
Aver
age n
umbe
r of
cells
fiel
dco
nditi
on
Basa
l 1
1005025105 1
10050105
LPS 2 (120583gmL) [225THC] (120583M) [225THC] (120583M)
Exp 1Exp 2Exp 3
(a)
20
0LPS
0
02
IL-6
(ng
mL)
IL-1
0 (n
gm
L)
IL-6
LPS
IL-10
0
02
BasalBasalBasal LPS
TNF-120572
TNF-120572
(ng
mL)
lowastlowast
lowast
lowastlowast
(biii)(bii)(bi)
5120583M500120583M
5120583M500120583M
5120583M500120583M
(b)
Figure 2 225THC is nontoxic and inhibits inflammatory cytokine responses in stimulated primary rat microglial cultures Effects of 225THCon primary rat microglial cells (a) Toxicity assay of 225THC on primary cultured rat microglia Data shown from three separate experimentsrepresent the mean number of live primary microglia (as assessed by livedead assay) present per field of view following incubation for24 h with different concentrations of 225THC (0 1 5 10 50 and 100 120583M) for 24 h with or without coactivation of the microglia with LPS(2120583gmL) (b) ELISA assessment of secreted cytokines in LPS (2 120583gmL) activated primary rat microglia cultures treated with 225THC at theconcentrations indicated Statistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005lowastlowast119901 lt 001 when compared with basla or as indicated
from 1 to 100 120583M(Figure 4(c)) or at 500120583M(data not shown)The positive control of NADPH oxidase activation PMA(10 ngmL) induced a significant increase in superoxide-positive cells in line with previous findings [22]
The inhibition of iNOS expression in microglia is inagreement with findings for another chalcone 4-dimethyl-amino-3101584041015840-dimethoxychalcone on mouse macrophagesstimulated with zymosan [22] This chalcone inhibited iNOSexpression with a similar concentration (sim10 120583M) to the chal-cone used in the present study Herencia et al [26] proposedthat the effect on iNOS expression was via upstreammodula-tion of NADPH oxidase-mediated intracellular signalling
Since we did not detect modulation of microglial superoxidegeneration by 225THC it seems unlikely that this particularchalcone possesses this mechanism of action Other chal-cones have also been reported to modulate nitric oxide pro-duction or iNOS expression following LPS stimulation of amouse macrophage cell line [36] the BV2 microglial cell line[27] and a rat microglial cell line [37] within similar com-pound concentration ranges of 1ndash50 120583MThe exact signallingpathways inhibited by different chalcones may be diverseas some studies suggest modulation of nuclear factor-120581BMAPK or c-Jun pathways and others suggest STAT1 path-ways may be involved [26 37]
6 Oxidative Medicine and Cellular Longevity
60
220
Abov
e con
trol (
)
Basal LPS
ns
ns
lowastlowast
lowast
lowast lowast
1120583M5120583M10120583M
TNF-120572
(a)
MCP-1
60
220
LPS
ns
ns
lowastlowast
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(b)
Rantes
60
280
LPS
ns
lowast
lowastlowast
lowast
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(c)
LPS
MIP
80
170
ns
ns
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(d)
Figure 3 225THC modulates inflammatory cytokine and chemokine responses in murine BV2 microglial cells 225THC-modulation ofcytokinechemokine secretion by the BV2 microglial cell line ELISA assessment of cytokine ((a) TNF-120572) and chemokine ((b) MCP-1 (c)RANTES (d) MIP) secretion by BV2 microglial cells following stimulation with LPS (2 120583gmL) for 24 h in the presence of 225THC at theconcentrations indicated Statistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005lowastlowast119901 lt 001 ns not significant when compared with basal or as indicated
35 Modulation ofMicroglial Inflammatory Phenotype UponLPS stimulationmicroglia adopt an inflammatory phenotypewhich is characterised by enhanced cell surface expression ofkey adhesion (CD11b CD54) and costimulatory molecules(CD40) We investigated the effects of 225THC treatment
on BV2 cell surface expression of CD11b CD40 CD54 andCD68 in basal and LPS-treated BV2 microglia (Figure 5)Flow cytometric analysis (FACs) of marker mean fluores-cence intensity (MFI) revealed no significant changes in basalCD40 CD54 CD68 or CD11b expression levels following
Oxidative Medicine and Cellular Longevity 7
iNOS
Basal 1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M)[225THC] (120583M)
120573-actin
(a)
0
07
1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
Den
sitom
etry
of i
NO
S120573
-act
in
lowast lowast
(b)
Supe
roxi
de-p
ositi
ve ce
lls(
tota
l)
50
01 5 10 100 1 5 10 100
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
lowastlowast lowastlowastlowastlowast lowastlowast
PMA10ngmL
lowast
lowast
(c)
1120583M LPS + 1120583M
10120583M
100120583M
LPS + 10120583M
LPS + 100120583M
Control LPS
(d)
Figure 4 225THC inhibits the LPS-induced expression of iNOS but not NADPH-driven superoxide production in BV2 microgliaModulation of BV2 iNOS expression but not superoxide production by 225THC Representative Western blots of iNOS expression and 120573-actin as a loading control (a) and densitometry (b) of three separate blots for BV2microglia exposed to 225THC at the stated concentrations plusmnLPS (2120583gmL) for 24 h Statistical analysis was performed by a two-way ANOVAwith Bonferroni post hoc analysis lowast indicates 119901 lt 005whencompared with basal or as indicated (c) Superoxide generation in BV2 microglia measured by dihydroethidium fluorescence following 24 hexposure to 225THC at the concentrations indicated plusmn LPS (2120583gmL) The phorbol PMA was used as a positive activator of superoxidegeneration (d) Representative fields of BV2 microglia generating intracellular superoxide following 24 h exposure to 225THC +minus LPSStatistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005 lowastlowast indicates 119901 lt 001when compared with basal or as indicated
225THC treatment for 48 h (data not shown) However fol-lowing stimulation with 2 120583gmL LPS CD40 (Figure 5(bii))CD54 (Figure 5(cii)) and CD68 (Figure 5(dii)) were signif-icantly upregulated and these changes in expression wereabrogated in the presence of 5 or 10 120583M 225THC CD40appeared to be the most potently regulatedmarker with both5 and 10 120583M 225THC completely inhibiting LPS-inducedupregulation (Figure 5(bii)) while the treatment had moremoderate effects on CD54 and CD68 marker expression As
in the basal state 225THC had no effect on CD11b expressionby LPS-stimulated microglia (Figure 5(aii))
Overall these data suggest that 225THC targets thesignalling pathways downstream of LPS binding and abro-gates the expression of genes such as TNF-120572 iNOS andCD40 Since these are all reported NF-120581B driven genesfuture work could investigate this further to identify theexact pathways modified by 225THC Abrogating this geneexpression produced a less neurotoxic microglial phenotype
8 Oxidative Medicine and Cellular Longevity
CD11b
(ai) (aii)
[225THC] (120583M)
FL1-H
CD11b
Basal LPS 1 5 10
111009080706050403020100
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(a)
CD40
(bi) (bii)
[225THC] (120583M)
FL1-H
CD40
Basal LPS 1 5 10
125
100
075
050
025
000
175
150
lowastlowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(b)
(ci) (cii)
[225THC] (120583M)
FL1-H
CD54 (ICAM-1)CD54 (ICAM-1)
Basal LPS 1 5 10
400
300
200
100
000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(c)
CD68
(di) (dii)
[225THC] (120583M)FL1-H
CD68
Basal LPS 1 5 10
160140120100080060040020000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(d)
Figure 5 225THC treatmentmodulates inflammatory cell surfacemarker expression in LPS-treated BV2microglia Flow cytometric analysisof 225THC-treated BV2 microglia following LPS stimulation (a) Representative histogram plots of BV2 cell surface CD11b CD40 CD54and CD68 expression following 2 120583gmL LPS-treatment for 48 hours (unfilled histograms) versus isotype control (black-filled histograms)and in the presence of 10 120583M 225THC (grey-filled histograms) (b) Summary data showing fold changes in CD11b CD40 CD54 or CD68expression by mean fluorescence intensity (MFI) from 4 independent experiments Data are mean values plusmn SEM where lowast indicates 119901 lt 005by Studentrsquos 119905-test
Oxidative Medicine and Cellular Longevity 9
which is likely to be beneficial for a number of neurodegen-erative diseases in which aberrant microglial inflammatoryreactivity is implicated Future work could pinpoint furtherthe microglial phenotype favoured by 225THC exposure andexplore the genotype of the microglia
Abbreviations
ANOVA Analysis of varianceDPPH 22-Diphenyl-1-picrylhydrazyldHEth DihydroethidiumCGCs Cerebellar granule neuronsDIV Days in vitroDAPI 410158406-Diamidino-2-phenylindoleDMSO DimethylsulphoxideECL ElectrochemiluminescenceELISA Enzyme-linked immunosorbent assayGABAA 120574-Aminobutyric acidIFN-120574 Interferon-120574IL-1120573 Interleukin-1120573IL-6 Interleukin-6IL-10 Interleukin-10iNOS Inducible nitric oxide synthaseLPS LipopolysaccharideMCP-1 CCL2monocyte chemoattractant
protein-1MEM Modified Eaglersquos mediumMIP Macrophage inhibitory proteinMCP-1 Monocyte chemoattractant protein-1NGS Normal goat serumPBS Phosphate buffered salinePFA Paraformaldehyde2-OH-E+ 2-HydroxyethidiumRANTES CCL5regulation on activation normal T
cell expressed and secretedROS Reactive oxygen speciesTNF-120572 Tumour necrosis factor-120572
Conflict of Interests
All authors declare no conflict of interests
Acknowledgments
Manasi Jiwrajka would like to thank Dr Kathleen Susman forher guidance This research was supported by the Universityof Queensland Advantage Grant Howard Hughes MedicalFoundation Grant 52006322 to Vassar College the ShermanFairchild Foundation and Vassar College and by generalfunds to the Pocock lab
References
[1] S Ducki R Forrest J A Hadfield et al ldquoPotent antimitotic andcell growth inhibitory properties of substituted chalconesrdquo Bio-organic and Medicinal Chemistry Letters vol 8 no 9 pp 1051ndash1056 1998
[2] M L F Ferreyra S P Rius and P Casati ldquoFlavonoids biosyn-thesis biological functions and biotechnological applicationsrdquoFrontiers in Plant Science vol 3 article 222 15 pages 2012
[3] G E-D A A Abuo-Rahma M Abdel-Aziz M A E Mouradand H H Farag ldquoSynthesis anti-inflammatory activity andulcerogenic liability of novel nitric oxide donatingchalconehybridsrdquo Bioorganic and Medicinal Chemistry vol 20 no 1 pp195ndash206 2012
[4] N K Sahu S S Balbhadra J Choudhary and D V KohlildquoExploring pharmacological significance of chalcone scaffoldrdquoCurrent Medicinal Chemistry vol 19 no 2 pp 209ndash225 2012
[5] H Wei X Zhang G Wu et al ldquoChalcone derivatives from thefern Cyclosorus parasiticus and their anti-proliferative activityrdquoFood and Chemical Toxicology vol 60 pp 147ndash152 2013
[6] C W Mai M Yaeghoobi N Abd-Rahman Y B Kang andM R Pichika ldquoChalcones with electron-withdrawing and ele-ctron-donating substituents anticancer activity against TRAILresistant cancer cells structure-activity relationship analysisand regulation of apoptotic proteinsrdquo European Journal ofMedicinal Chemistry vol 77 pp 378ndash387 2014
[7] Z Nowakowska ldquoA review of anti-infective and anti-inflam-matory chalconesrdquo European Journal of Medicinal Chemistryvol 42 no 2 pp 125ndash137 2007
[8] Y-M Lin Y Zhou M T Flavin L-M Zhou W Nie and F-CChen ldquoChalcones and flavonoids as anti-tuberculosis agentsrdquoBioorganic and Medicinal Chemistry vol 10 no 8 pp 2795ndash2802 2002
[9] JMojzis L Varinska GMojzisova I Kostova and LMirossayldquoAnti-angiogenic effects of flavonoids and chalconesrdquo Pharma-cological Research vol 57 no 4 pp 259ndash265 2008
[10] F Dal Piaz A Braca M A Belisario and N De TommasildquoThioredoxin systemmodulation by plant and fungal secondarymetabolitesrdquo Current Medicinal Chemistry vol 17 no 5 pp479ndash494 2010
[11] N Yadav S K Dixit A Bhattacharya et al ldquoAntimalarial activ-ity of newly synthesized chalcone derivatives in vitrordquo Chem-ical Biology and Drug Design vol 80 no 2 pp 340ndash347 2012
[12] F V DeFeudis ldquoNovel therapy for Alzheimerrsquos diseaserdquo Neuro-biology of Aging vol 10 no 5 pp 467ndash468 1989
[13] Y Nomura and Y Kitamura ldquoInducible nitric oxide synthasein glial cellsrdquo Neuroscience Research vol 18 no 2 pp 103ndash1071993
[14] P J Kingham M L Cuzner and J M Pocock ldquoApoptotic path-ways mobilized in microglia and neurones as a consequenceof chromogranin A-induced microglial activationrdquo Journal ofNeurochemistry vol 73 no 2 pp 538ndash547 1999
[15] S Singh S Swarnkar P Goswami and C Nath ldquoAstrocytes andmicroglia responses to neuropathological conditionsrdquo Inter-national Journal of Neuroscience vol 121 no 11 pp 589ndash5972011
[16] Y Y Deng J Lu E-A Ling and C Kaur ldquoRole of microglia inthe process of inflammation in the hypoxic developing brainrdquoFrontiers in Bioscience (Scholar Edition) vol 3 no 3 pp 884ndash900 2011
[17] M Dumont and M F Beal ldquoNeuroprotective strategies involv-ing ROS in Alzheimer diseaserdquo Free Radical Biology andMedicine vol 51 no 5 pp 1014ndash1026 2011
[18] AD Kraft andG JeanHarry ldquoFeatures ofmicroglia and neuro-inflammation relevant to environmental exposure and neuro-toxicityrdquo International Journal of Environmental Research andPublic Health vol 8 no 7 pp 2980ndash3018 2011
[19] S Cho S Kim Z Jin et al ldquoIsoliquiritigenin a chalcone com-pound is a positive allosteric modulator of GABA A recep-tors and shows hypnotic effectsrdquo Biochemical and BiophysicalResearch Communications vol 413 no 4 pp 637ndash642 2011
10 Oxidative Medicine and Cellular Longevity
[20] A Elmann A Telerman H Erlank et al ldquoProtective and anti-oxidant effects of a chalconoid from Pulicaria incisa on brainastrocytesrdquoOxidativeMedicine andCellular Longevity vol 2013Article ID 694398 10 pages 2013
[21] M Rossi F Caruso E J Crespi et al ldquoProbing antioxidantactivity of 21015840-hydroxychalcones crystal and molecular struc-tures in vitro antiproliferative studies and in vivo effects onglucose regulationrdquo Biochimie vol 95 no 10 pp 1954ndash19632013
[22] E LMead AMosley S Eaton L Dobson S J Heales and JMPocock ldquoMicroglial neurotransmitter receptors trigger super-oxide production in microglia consequences for microglial-neuronal interactionsrdquo Journal of Neurochemistry vol 121 no2 pp 287ndash301 2012
[23] C M Davenport I G Sevastou C Hooper and J M PocockldquoInhibiting p53 pathways in microglia attenuates microglial-evoked neurotoxicity following exposure to Alzheimer pep-tidesrdquo Journal of Neurochemistry vol 112 no 2 pp 552ndash5632010
[24] N C Garbett N B Hammond and D E Graves ldquoInfluence ofthe amino substituents in the interaction of ethidium bromidewith DNArdquo Biophysical Journal vol 87 no 6 pp 3974ndash39812004
[25] A Karlsson J B Nixon and L C McPhail ldquoPhorbol myristateacetate induces neutrophilNADPH-oxidase activity by two sep-arate signal transduction pathways dependent or independentof phosphatidylinositol 3-kinaserdquo Journal of Leukocyte Biologyvol 67 no 3 pp 396ndash404 2000
[26] F Herencia M L Ferrandiz A Ubeda et al ldquo4-Dimethyl-amino-3101584041015840-dimethoxychalcone down-regulates iNOS expres-sion and exerts anti-inflammatory effectsrdquo Free Radical Biologyamp Medicine vol 30 no 1 pp 43ndash50 2001
[27] N Mateeva M Gangapuram E Mazzio S Eyunni K FSoliman and K K Redda ldquoBiological evaluation of syntheticchalcone and flavone derivatives as anti-inflammatory agentsrdquoMedicinal Chemistry Research vol 24 no 4 pp 1672ndash16802015
[28] S C Morgan D L Taylor and J M Pocock ldquoMicroglia releaseactivators of neuronal proliferation mediated by activationof mitogen-activated protein kinase phosphatidylinositol-3-kinaseAkt and delta-Notch signalling cascadesrdquo Journal ofNeurochemistry vol 90 no 1 pp 89ndash101 2004
[29] J Jebelli T Piers and J Pocock ldquoSelective depletion of micro-glia from cerebellar granule cell cultures using L-leucinemethylesterrdquoThe Journal of Visualized Experiments no 101 Article IDe52983 2015
[30] R Von Bernhardi J E Tichauer and J Eugenın ldquoAging-dependent changes of microglial cells and their relevance forneurodegenerative disordersrdquo Journal of Neurochemistry vol112 no 5 pp 1099ndash1114 2010
[31] C Schwab and P L McGeer ldquoInflammatory aspects ofAlzheimer disease and other neurodegenerative disordersrdquoJournal of Alzheimerrsquos Disease vol 13 no 4 pp 359ndash369 2008
[32] A Henn S Lund M Hedtjarn A Schrattenholz P Porzgenand M Leist ldquoThe suitability of BV2 cells as alternative modelsystem for primary microglia cultures or for animal experi-ments examining brain inflammationrdquo Altex vol 26 no 2 pp83ndash94 2009
[33] S Bano K Javed S Ahmad et al ldquoSynthesis of some novel chal-cones flavanones and flavones and evaluation of their anti-inflammatory activityrdquo European Journal of Medicinal Chem-istry vol 65 pp 51ndash59 2013
[34] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[35] S L Deshmane S Kremlev S Amini and B E Sawaya ldquoMono-cyte chemoattractant protein-1 (MCP-1) an overviewrdquo Journalof Interferon and Cytokine Research vol 29 no 6 pp 313ndash3262009
[36] J Rojas J N Domınguez J E Charris G Lobo M Paya andM L Ferrandiz ldquoSynthesis and inhibitory activity of dimethyl-amino-chalcone derivatives on the induction of nitric oxidesynthaserdquo European Journal of Medicinal Chemistry vol 37 no8 pp 699ndash705 2002
[37] H Hara Y Nakamura M Ninomiya et al ldquoInhibitory effectsof chalcone glycosides isolated from Brassica rapa L lsquohidabenirsquoand their synthetic derivatives on LPS-induced NO productionin microgliardquo Bioorganic amp Medicinal Chemistry vol 19 no 18pp 5559ndash5568 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 3
number Microglia were imaged by fluorescence microscopyas above and superoxide-positive microglia as indicated byred nuclei were counted and expressed as a percentage oftotal cell number To ensure that the red fluorescence was dueto superoxide production experiments were carried out byincubation of the cells with 10 120583M apocynin [22]
26 ELISA of Secreted Cytokines by Microglia TNF-120572 orIL-1120573 concentrations in primary rat microglia cell culturesupernatants were quantified using Quantikine Rat TNF-120572or IL-1120573 Immunoassay kit according to the manufacturerrsquosinstructions (RampD Systems Abingdon UK) Cytokine con-centrations in cell supernatants were determined against astandard curve of TNF-120572 or IL-1120573 In addition a rangeof inflammatory mediators were analysed in BV2 cell cul-ture supernatants with an Inflammatory ELISA strip assayaccording to the manufacturerrsquos instructions (Signosis IncCaltag MedSystems Ltd Buckingham UK) In this case allvalues were presented as a percentage of control values Foreach condition the cytokine content in supernatants wasanalysed from three coverslips of microglia in three inde-pendent cell platings with each sample assayed in duplicateAll other chemicals and reagents were from Sigma (PooleUK)
27 Flow Cytometric Analysis of Microglial InflammatoryMarkers BV2 cells were treated with 1 5 or 10 120583M of225THC for 2 hours prior to the addition of 2 120583gmL LPSfor a further 48 hours Cells were harvested by washing inDulbeccorsquos phosphate buffered saline (PBS without Ca2+ andMg2+) for 20 minutes at 37∘C and then resuspended in coldPBS containing 05 BSA005 NaN
3to metabolically fix
the cells Cells were stained with anti-mouse CD11b-FITCCD40-FITC CD54-FITC and CD68-FITC or appropriateFITC-conjugated isotype control antibodies (Miltenyi BiotecUK) as per the manufacturerrsquos instructions and cell sur-face staining was assessed by flow cytometry (FACSCaliburrunning CellQuest Pro Becton Dickinson UK) and anal-ysed using Flowing software v251 Data are presented asaverage fold change in mean fluorescence intensity (MFI)versus untreated cells plusmn SEM from 4 independent experi-ments
28 Statistical Analysis Statistical analyses were performedusing one-way ANOVA with Tukey post hoc analysis wherecomparisons were made between treatments and controluntreated cells and also between treatment groups as indi-cated Where stated Studentrsquos 119905-test was used and all datawere from 3 separate experiments lowast119901 lt 005 lowastlowast119901 lt 001and lowastlowastlowast119901 lt 0005 compared with controls or as indicated infiguresWhere imagingwas performed aminimumof 3 fieldsof viewwere analysed from 3 coverslips per condition Imagesof fluorescent fields of cells orWestern blots are representativeof analysed data from3 independent experimentsThedensit-ometry forWesternswas performedusing Image Jmeasuringthe optical density (OD) of each band and normalising to the120573-actin obtained for that particular sample For the statisticalanalysis of these data each treatment was compared withcontrol untreated cells by ANOVA analysis and appropriate
comparisons were also made between treatment groups plusmninhibitors using Studentrsquos 119905-test
3 Results and Discussion
31 Effects of 225THC in Primary Rat Cerebellar GranuleCell Neuronal Cultures We initially investigated whether the225THCwas directly toxic to neurons by incubating primaryrat cerebellar granule cell (CGC) neuronal culture neuronswith concentrations of 5ndash500120583M 225THC for 24 h Thisrange of concentrations was assessed as previous studiesusing similar compounds have identified activity within thisrange [20 26 27] Microscopic livedead analyses revealedno significant toxicity in these cultures with DMSO solventcontrol or with 5 50 or 500120583M 225THC (Figure 1(a))when compared with untreated cells We next investigatedthe potential neuroprotective properties of 225THC in thesecultures (which we have shown previously contain 2ndash5microglia) [28 29] A range of 225THC concentrations werepreincubated with CGCs for 1 hour followed by stimulationwith 2120583gmL LPS or 10 ngmL IFN-120574 to induce an inflam-matory phenotype within the CGC cultures After 24 h theCGCs were imaged with Hoechst 33342 to visualise apop-totic nuclei within the cultures LPS or IFN-120574 significantlyincreased the percentage of apoptotic cells in the neuronalcultures above basal (Figure 1(b)) whilst pretreatment with225THC at 50 or 500 120583Msignificantly reduced the percentageof apoptotic cells in cultures treated with LPS or IFN-120574 SinceLPS and IFN-120574 are potent activators of microglia with noknown effects on cerebellar neurons and since incubationwith chalcone directly to these cultures in the absence ofinflammation did not enhance basal levels of neuronal sur-vival it is likely that suppression of the neuroinflammatoryand neurotoxic effects seen here are due to suppressionof microglial responses Thus we determined how theseeffects might be mediated One proposed mechanism forthe neurotoxicity observed is that activated microglia releaseproinflammatory cytokines that cause cytotoxicity to theneurons [15 30 31]
32 225THC Inhibits Proinflammatory Cytokine Secretion byLPS-Stimulated Primary Rat Microglial Cells As a prelimi-nary assessment we again investigated the potential cellulartoxicity of 225THC when added to primary rat microglialcultures We observed that in three separate experiments aspecific concentration of 50120583M 225THC appeared to inducesignificant cell loss fromcoverslips in both nonstimulated andLPS-stimulated cells whilst concentrations above and belowthis did not (Figure 2(a)) This was most likely due to lossof contact rather than toxicity as there was no increase in PIstaining DMSO controls did not affect cell numbers (data notshown)
We next assessed cytokine secretion inmicroglial cell cul-ture supernatants under basal (nonstimulating conditions)and following LPS stimulation (2120583gmL) in the presenceof 50 or 500120583M 225THC (Figure 2(b)) The basal secretionof TNF-120572 IL-6 and IL-10 was not significantly modulatedby 5 120583M or 500120583M 225THC whilst LPS-evoked TNF-120572 andIL-6 secretion but not IL-10 was significantly inhibited by
4 Oxidative Medicine and Cellular Longevity
0
100
Live
neu
rons
fiel
d (
)
5 50 500Basal
ns
50
DMSO[225THC] (120583M)
(a)
Apop
totic
neu
rons
()
LPS IFN-1205740
25
50
75
ns
ns
DMSO only
[225THC]
lowastlowastlowastlowastlowastlowast
5120583M
500120583M50120583M
(b)
Figure 1 225THC is nontoxic and neuroprotective in stimulated primary rat neuronal cultures (cerebellar granule cell neurons CGCs)Effects of 225THC on primary rat CGC neurons (a) Toxicity assay of 225THC on primary rat CGC neurons Data from three separateexperiments represent the mean number of live neurons (as assessed by livedead assay) present per field of view following incubation for24 h with different concentrations of 225THC (0 5 50 and 500 120583M) for 24 h DMSO is the solvent control basal is CGCs with nothing addedStatistical analysis was performed by ANOVA with Bonferroni post hoc analysis and revealed no toxicity at the concentrations tested whenall concentrations were compared with basal The DMSO solvent control was also not toxic (b) Direct treatment of CGC cultures for 24 hwith 225THC (0 5 50 and 500120583M) and 2 120583gmL LPS or 10 ngmL IFN-120574 to activate resident microglia in the cultures The percentage ofapoptotic cells following Hoechst 33342 visualisation of nuclear condensation is shown for each treatment Statistical analysis was performedby a two-way ANOVA with Bonferroni post hoc analysis lowast lowast lowast indicates 119901 lt 0001 ns not significant when compared as indicated or withthe appropriate basal control
5 120583M or 500120583M 225THC (Figures 2(bi) 2(bii) and 2(biii)resp)
33 225THC Modulates Inflammatory Cytokine and Chemo-kine Responses in Murine BV2Microglial Cells To investigatethe microglial aspects further we used the BV2 mousemicroglial cell line which is a well validated substitute forprimary microglia [32] Further examination of a lowerrange of 225THC concentrations (1120583M 5 120583M and 10 120583M)following a 48 h LPS stimulation revealed selective inhi-bition of LPS-induced TNF-120572 in line with our findingsin primary microglia (Figure 3(a)) In addition the basalsecretion of monocyte chemoattractant protein-1 (MCP-1)was also significantly inhibited (Figure 3(b)) We did notobservemodulation of basal or LPS-evoked secretion of IFN-R IL-1120572 IL-1120573 (data not shown) or of RANTES (Figure 3(c))or MIP (Figure 3(d)) These data suggest that 225THC hasan effect on the secretion of inflammatory cytokines suchas TNF-120572 IL-6 and MCP-1 and the reduction of thesecytokines may aid neuroprotection Interestingly in line withour findings Herencia et al [26] report that the chalconeused in their study inhibited TNF-120572 mouse air pouch tissuelevels from leukocytes stimulated by zymosan whilst Bano etal [33] reported suppression of LPS-evoked TNF-120572 secretionfrom a mouse macrophage line with a synthetic chalcone Inaddition we found 225THC inhibited the secretion of twoother cytokines IL-6 and MCP-1 IL-6 has well recognised
roles in neuroinflammation and promotes microgliosis [34]whilst MCP-1 (also known as CCL-2) acts as a chemoattrac-tant to recruit monocytes and macrophages into the brain[35] Interestingly recent work suggests different chalconesmay selectively inhibit the secretion of different groups ofcytokines and inflammatory mediators [27] We did not seesignificant modulation by 225THC of RANTES MIP or IL-10 Other chalcones have been shown to promote the produc-tion of neurotrophic factors such as GDNF from astrocytes[20] and this is worth future exploration for microglia
34 Modulation of iNOS Expression and ROS Generationby 225THC The expression of iNOS was assessed in BV2microglia under basal conditions and following LPS stimu-lation (2 120583gmL) for 48 h in the presence of 225THC (1120583M5 120583M or 10 120583M) No obvious effects were observed on theexpression of iNOS in nonstimulated microglia that is225THC did not induce expression of iNOS (Figure 4(a))However LPS-evoked stimulation of iNOS expression wassignificantly inhibited by 225THC at concentrations above5 120583M(Figures 4(a) and 4(b))We next investigatedmicroglialsuperoxide generation using microscopic fluorescence imag-ing (Figure 4(c)) No obvious effects of the 225THC wereobserved on the basal cell numbers expressing superoxide-positive cell nuclei and whilst the number of superoxide-positive cells was increased following LPS stimulation thiswas not significantly regulated by 225THC concentrations
Oxidative Medicine and Cellular Longevity 5
0
200
400
Aver
age n
umbe
r of
cells
fiel
dco
nditi
on
Basa
l 1
1005025105 1
10050105
LPS 2 (120583gmL) [225THC] (120583M) [225THC] (120583M)
Exp 1Exp 2Exp 3
(a)
20
0LPS
0
02
IL-6
(ng
mL)
IL-1
0 (n
gm
L)
IL-6
LPS
IL-10
0
02
BasalBasalBasal LPS
TNF-120572
TNF-120572
(ng
mL)
lowastlowast
lowast
lowastlowast
(biii)(bii)(bi)
5120583M500120583M
5120583M500120583M
5120583M500120583M
(b)
Figure 2 225THC is nontoxic and inhibits inflammatory cytokine responses in stimulated primary rat microglial cultures Effects of 225THCon primary rat microglial cells (a) Toxicity assay of 225THC on primary cultured rat microglia Data shown from three separate experimentsrepresent the mean number of live primary microglia (as assessed by livedead assay) present per field of view following incubation for24 h with different concentrations of 225THC (0 1 5 10 50 and 100 120583M) for 24 h with or without coactivation of the microglia with LPS(2120583gmL) (b) ELISA assessment of secreted cytokines in LPS (2 120583gmL) activated primary rat microglia cultures treated with 225THC at theconcentrations indicated Statistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005lowastlowast119901 lt 001 when compared with basla or as indicated
from 1 to 100 120583M(Figure 4(c)) or at 500120583M(data not shown)The positive control of NADPH oxidase activation PMA(10 ngmL) induced a significant increase in superoxide-positive cells in line with previous findings [22]
The inhibition of iNOS expression in microglia is inagreement with findings for another chalcone 4-dimethyl-amino-3101584041015840-dimethoxychalcone on mouse macrophagesstimulated with zymosan [22] This chalcone inhibited iNOSexpression with a similar concentration (sim10 120583M) to the chal-cone used in the present study Herencia et al [26] proposedthat the effect on iNOS expression was via upstreammodula-tion of NADPH oxidase-mediated intracellular signalling
Since we did not detect modulation of microglial superoxidegeneration by 225THC it seems unlikely that this particularchalcone possesses this mechanism of action Other chal-cones have also been reported to modulate nitric oxide pro-duction or iNOS expression following LPS stimulation of amouse macrophage cell line [36] the BV2 microglial cell line[27] and a rat microglial cell line [37] within similar com-pound concentration ranges of 1ndash50 120583MThe exact signallingpathways inhibited by different chalcones may be diverseas some studies suggest modulation of nuclear factor-120581BMAPK or c-Jun pathways and others suggest STAT1 path-ways may be involved [26 37]
6 Oxidative Medicine and Cellular Longevity
60
220
Abov
e con
trol (
)
Basal LPS
ns
ns
lowastlowast
lowast
lowast lowast
1120583M5120583M10120583M
TNF-120572
(a)
MCP-1
60
220
LPS
ns
ns
lowastlowast
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(b)
Rantes
60
280
LPS
ns
lowast
lowastlowast
lowast
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(c)
LPS
MIP
80
170
ns
ns
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(d)
Figure 3 225THC modulates inflammatory cytokine and chemokine responses in murine BV2 microglial cells 225THC-modulation ofcytokinechemokine secretion by the BV2 microglial cell line ELISA assessment of cytokine ((a) TNF-120572) and chemokine ((b) MCP-1 (c)RANTES (d) MIP) secretion by BV2 microglial cells following stimulation with LPS (2 120583gmL) for 24 h in the presence of 225THC at theconcentrations indicated Statistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005lowastlowast119901 lt 001 ns not significant when compared with basal or as indicated
35 Modulation ofMicroglial Inflammatory Phenotype UponLPS stimulationmicroglia adopt an inflammatory phenotypewhich is characterised by enhanced cell surface expression ofkey adhesion (CD11b CD54) and costimulatory molecules(CD40) We investigated the effects of 225THC treatment
on BV2 cell surface expression of CD11b CD40 CD54 andCD68 in basal and LPS-treated BV2 microglia (Figure 5)Flow cytometric analysis (FACs) of marker mean fluores-cence intensity (MFI) revealed no significant changes in basalCD40 CD54 CD68 or CD11b expression levels following
Oxidative Medicine and Cellular Longevity 7
iNOS
Basal 1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M)[225THC] (120583M)
120573-actin
(a)
0
07
1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
Den
sitom
etry
of i
NO
S120573
-act
in
lowast lowast
(b)
Supe
roxi
de-p
ositi
ve ce
lls(
tota
l)
50
01 5 10 100 1 5 10 100
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
lowastlowast lowastlowastlowastlowast lowastlowast
PMA10ngmL
lowast
lowast
(c)
1120583M LPS + 1120583M
10120583M
100120583M
LPS + 10120583M
LPS + 100120583M
Control LPS
(d)
Figure 4 225THC inhibits the LPS-induced expression of iNOS but not NADPH-driven superoxide production in BV2 microgliaModulation of BV2 iNOS expression but not superoxide production by 225THC Representative Western blots of iNOS expression and 120573-actin as a loading control (a) and densitometry (b) of three separate blots for BV2microglia exposed to 225THC at the stated concentrations plusmnLPS (2120583gmL) for 24 h Statistical analysis was performed by a two-way ANOVAwith Bonferroni post hoc analysis lowast indicates 119901 lt 005whencompared with basal or as indicated (c) Superoxide generation in BV2 microglia measured by dihydroethidium fluorescence following 24 hexposure to 225THC at the concentrations indicated plusmn LPS (2120583gmL) The phorbol PMA was used as a positive activator of superoxidegeneration (d) Representative fields of BV2 microglia generating intracellular superoxide following 24 h exposure to 225THC +minus LPSStatistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005 lowastlowast indicates 119901 lt 001when compared with basal or as indicated
225THC treatment for 48 h (data not shown) However fol-lowing stimulation with 2 120583gmL LPS CD40 (Figure 5(bii))CD54 (Figure 5(cii)) and CD68 (Figure 5(dii)) were signif-icantly upregulated and these changes in expression wereabrogated in the presence of 5 or 10 120583M 225THC CD40appeared to be the most potently regulatedmarker with both5 and 10 120583M 225THC completely inhibiting LPS-inducedupregulation (Figure 5(bii)) while the treatment had moremoderate effects on CD54 and CD68 marker expression As
in the basal state 225THC had no effect on CD11b expressionby LPS-stimulated microglia (Figure 5(aii))
Overall these data suggest that 225THC targets thesignalling pathways downstream of LPS binding and abro-gates the expression of genes such as TNF-120572 iNOS andCD40 Since these are all reported NF-120581B driven genesfuture work could investigate this further to identify theexact pathways modified by 225THC Abrogating this geneexpression produced a less neurotoxic microglial phenotype
8 Oxidative Medicine and Cellular Longevity
CD11b
(ai) (aii)
[225THC] (120583M)
FL1-H
CD11b
Basal LPS 1 5 10
111009080706050403020100
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(a)
CD40
(bi) (bii)
[225THC] (120583M)
FL1-H
CD40
Basal LPS 1 5 10
125
100
075
050
025
000
175
150
lowastlowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(b)
(ci) (cii)
[225THC] (120583M)
FL1-H
CD54 (ICAM-1)CD54 (ICAM-1)
Basal LPS 1 5 10
400
300
200
100
000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(c)
CD68
(di) (dii)
[225THC] (120583M)FL1-H
CD68
Basal LPS 1 5 10
160140120100080060040020000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(d)
Figure 5 225THC treatmentmodulates inflammatory cell surfacemarker expression in LPS-treated BV2microglia Flow cytometric analysisof 225THC-treated BV2 microglia following LPS stimulation (a) Representative histogram plots of BV2 cell surface CD11b CD40 CD54and CD68 expression following 2 120583gmL LPS-treatment for 48 hours (unfilled histograms) versus isotype control (black-filled histograms)and in the presence of 10 120583M 225THC (grey-filled histograms) (b) Summary data showing fold changes in CD11b CD40 CD54 or CD68expression by mean fluorescence intensity (MFI) from 4 independent experiments Data are mean values plusmn SEM where lowast indicates 119901 lt 005by Studentrsquos 119905-test
Oxidative Medicine and Cellular Longevity 9
which is likely to be beneficial for a number of neurodegen-erative diseases in which aberrant microglial inflammatoryreactivity is implicated Future work could pinpoint furtherthe microglial phenotype favoured by 225THC exposure andexplore the genotype of the microglia
Abbreviations
ANOVA Analysis of varianceDPPH 22-Diphenyl-1-picrylhydrazyldHEth DihydroethidiumCGCs Cerebellar granule neuronsDIV Days in vitroDAPI 410158406-Diamidino-2-phenylindoleDMSO DimethylsulphoxideECL ElectrochemiluminescenceELISA Enzyme-linked immunosorbent assayGABAA 120574-Aminobutyric acidIFN-120574 Interferon-120574IL-1120573 Interleukin-1120573IL-6 Interleukin-6IL-10 Interleukin-10iNOS Inducible nitric oxide synthaseLPS LipopolysaccharideMCP-1 CCL2monocyte chemoattractant
protein-1MEM Modified Eaglersquos mediumMIP Macrophage inhibitory proteinMCP-1 Monocyte chemoattractant protein-1NGS Normal goat serumPBS Phosphate buffered salinePFA Paraformaldehyde2-OH-E+ 2-HydroxyethidiumRANTES CCL5regulation on activation normal T
cell expressed and secretedROS Reactive oxygen speciesTNF-120572 Tumour necrosis factor-120572
Conflict of Interests
All authors declare no conflict of interests
Acknowledgments
Manasi Jiwrajka would like to thank Dr Kathleen Susman forher guidance This research was supported by the Universityof Queensland Advantage Grant Howard Hughes MedicalFoundation Grant 52006322 to Vassar College the ShermanFairchild Foundation and Vassar College and by generalfunds to the Pocock lab
References
[1] S Ducki R Forrest J A Hadfield et al ldquoPotent antimitotic andcell growth inhibitory properties of substituted chalconesrdquo Bio-organic and Medicinal Chemistry Letters vol 8 no 9 pp 1051ndash1056 1998
[2] M L F Ferreyra S P Rius and P Casati ldquoFlavonoids biosyn-thesis biological functions and biotechnological applicationsrdquoFrontiers in Plant Science vol 3 article 222 15 pages 2012
[3] G E-D A A Abuo-Rahma M Abdel-Aziz M A E Mouradand H H Farag ldquoSynthesis anti-inflammatory activity andulcerogenic liability of novel nitric oxide donatingchalconehybridsrdquo Bioorganic and Medicinal Chemistry vol 20 no 1 pp195ndash206 2012
[4] N K Sahu S S Balbhadra J Choudhary and D V KohlildquoExploring pharmacological significance of chalcone scaffoldrdquoCurrent Medicinal Chemistry vol 19 no 2 pp 209ndash225 2012
[5] H Wei X Zhang G Wu et al ldquoChalcone derivatives from thefern Cyclosorus parasiticus and their anti-proliferative activityrdquoFood and Chemical Toxicology vol 60 pp 147ndash152 2013
[6] C W Mai M Yaeghoobi N Abd-Rahman Y B Kang andM R Pichika ldquoChalcones with electron-withdrawing and ele-ctron-donating substituents anticancer activity against TRAILresistant cancer cells structure-activity relationship analysisand regulation of apoptotic proteinsrdquo European Journal ofMedicinal Chemistry vol 77 pp 378ndash387 2014
[7] Z Nowakowska ldquoA review of anti-infective and anti-inflam-matory chalconesrdquo European Journal of Medicinal Chemistryvol 42 no 2 pp 125ndash137 2007
[8] Y-M Lin Y Zhou M T Flavin L-M Zhou W Nie and F-CChen ldquoChalcones and flavonoids as anti-tuberculosis agentsrdquoBioorganic and Medicinal Chemistry vol 10 no 8 pp 2795ndash2802 2002
[9] JMojzis L Varinska GMojzisova I Kostova and LMirossayldquoAnti-angiogenic effects of flavonoids and chalconesrdquo Pharma-cological Research vol 57 no 4 pp 259ndash265 2008
[10] F Dal Piaz A Braca M A Belisario and N De TommasildquoThioredoxin systemmodulation by plant and fungal secondarymetabolitesrdquo Current Medicinal Chemistry vol 17 no 5 pp479ndash494 2010
[11] N Yadav S K Dixit A Bhattacharya et al ldquoAntimalarial activ-ity of newly synthesized chalcone derivatives in vitrordquo Chem-ical Biology and Drug Design vol 80 no 2 pp 340ndash347 2012
[12] F V DeFeudis ldquoNovel therapy for Alzheimerrsquos diseaserdquo Neuro-biology of Aging vol 10 no 5 pp 467ndash468 1989
[13] Y Nomura and Y Kitamura ldquoInducible nitric oxide synthasein glial cellsrdquo Neuroscience Research vol 18 no 2 pp 103ndash1071993
[14] P J Kingham M L Cuzner and J M Pocock ldquoApoptotic path-ways mobilized in microglia and neurones as a consequenceof chromogranin A-induced microglial activationrdquo Journal ofNeurochemistry vol 73 no 2 pp 538ndash547 1999
[15] S Singh S Swarnkar P Goswami and C Nath ldquoAstrocytes andmicroglia responses to neuropathological conditionsrdquo Inter-national Journal of Neuroscience vol 121 no 11 pp 589ndash5972011
[16] Y Y Deng J Lu E-A Ling and C Kaur ldquoRole of microglia inthe process of inflammation in the hypoxic developing brainrdquoFrontiers in Bioscience (Scholar Edition) vol 3 no 3 pp 884ndash900 2011
[17] M Dumont and M F Beal ldquoNeuroprotective strategies involv-ing ROS in Alzheimer diseaserdquo Free Radical Biology andMedicine vol 51 no 5 pp 1014ndash1026 2011
[18] AD Kraft andG JeanHarry ldquoFeatures ofmicroglia and neuro-inflammation relevant to environmental exposure and neuro-toxicityrdquo International Journal of Environmental Research andPublic Health vol 8 no 7 pp 2980ndash3018 2011
[19] S Cho S Kim Z Jin et al ldquoIsoliquiritigenin a chalcone com-pound is a positive allosteric modulator of GABA A recep-tors and shows hypnotic effectsrdquo Biochemical and BiophysicalResearch Communications vol 413 no 4 pp 637ndash642 2011
10 Oxidative Medicine and Cellular Longevity
[20] A Elmann A Telerman H Erlank et al ldquoProtective and anti-oxidant effects of a chalconoid from Pulicaria incisa on brainastrocytesrdquoOxidativeMedicine andCellular Longevity vol 2013Article ID 694398 10 pages 2013
[21] M Rossi F Caruso E J Crespi et al ldquoProbing antioxidantactivity of 21015840-hydroxychalcones crystal and molecular struc-tures in vitro antiproliferative studies and in vivo effects onglucose regulationrdquo Biochimie vol 95 no 10 pp 1954ndash19632013
[22] E LMead AMosley S Eaton L Dobson S J Heales and JMPocock ldquoMicroglial neurotransmitter receptors trigger super-oxide production in microglia consequences for microglial-neuronal interactionsrdquo Journal of Neurochemistry vol 121 no2 pp 287ndash301 2012
[23] C M Davenport I G Sevastou C Hooper and J M PocockldquoInhibiting p53 pathways in microglia attenuates microglial-evoked neurotoxicity following exposure to Alzheimer pep-tidesrdquo Journal of Neurochemistry vol 112 no 2 pp 552ndash5632010
[24] N C Garbett N B Hammond and D E Graves ldquoInfluence ofthe amino substituents in the interaction of ethidium bromidewith DNArdquo Biophysical Journal vol 87 no 6 pp 3974ndash39812004
[25] A Karlsson J B Nixon and L C McPhail ldquoPhorbol myristateacetate induces neutrophilNADPH-oxidase activity by two sep-arate signal transduction pathways dependent or independentof phosphatidylinositol 3-kinaserdquo Journal of Leukocyte Biologyvol 67 no 3 pp 396ndash404 2000
[26] F Herencia M L Ferrandiz A Ubeda et al ldquo4-Dimethyl-amino-3101584041015840-dimethoxychalcone down-regulates iNOS expres-sion and exerts anti-inflammatory effectsrdquo Free Radical Biologyamp Medicine vol 30 no 1 pp 43ndash50 2001
[27] N Mateeva M Gangapuram E Mazzio S Eyunni K FSoliman and K K Redda ldquoBiological evaluation of syntheticchalcone and flavone derivatives as anti-inflammatory agentsrdquoMedicinal Chemistry Research vol 24 no 4 pp 1672ndash16802015
[28] S C Morgan D L Taylor and J M Pocock ldquoMicroglia releaseactivators of neuronal proliferation mediated by activationof mitogen-activated protein kinase phosphatidylinositol-3-kinaseAkt and delta-Notch signalling cascadesrdquo Journal ofNeurochemistry vol 90 no 1 pp 89ndash101 2004
[29] J Jebelli T Piers and J Pocock ldquoSelective depletion of micro-glia from cerebellar granule cell cultures using L-leucinemethylesterrdquoThe Journal of Visualized Experiments no 101 Article IDe52983 2015
[30] R Von Bernhardi J E Tichauer and J Eugenın ldquoAging-dependent changes of microglial cells and their relevance forneurodegenerative disordersrdquo Journal of Neurochemistry vol112 no 5 pp 1099ndash1114 2010
[31] C Schwab and P L McGeer ldquoInflammatory aspects ofAlzheimer disease and other neurodegenerative disordersrdquoJournal of Alzheimerrsquos Disease vol 13 no 4 pp 359ndash369 2008
[32] A Henn S Lund M Hedtjarn A Schrattenholz P Porzgenand M Leist ldquoThe suitability of BV2 cells as alternative modelsystem for primary microglia cultures or for animal experi-ments examining brain inflammationrdquo Altex vol 26 no 2 pp83ndash94 2009
[33] S Bano K Javed S Ahmad et al ldquoSynthesis of some novel chal-cones flavanones and flavones and evaluation of their anti-inflammatory activityrdquo European Journal of Medicinal Chem-istry vol 65 pp 51ndash59 2013
[34] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[35] S L Deshmane S Kremlev S Amini and B E Sawaya ldquoMono-cyte chemoattractant protein-1 (MCP-1) an overviewrdquo Journalof Interferon and Cytokine Research vol 29 no 6 pp 313ndash3262009
[36] J Rojas J N Domınguez J E Charris G Lobo M Paya andM L Ferrandiz ldquoSynthesis and inhibitory activity of dimethyl-amino-chalcone derivatives on the induction of nitric oxidesynthaserdquo European Journal of Medicinal Chemistry vol 37 no8 pp 699ndash705 2002
[37] H Hara Y Nakamura M Ninomiya et al ldquoInhibitory effectsof chalcone glycosides isolated from Brassica rapa L lsquohidabenirsquoand their synthetic derivatives on LPS-induced NO productionin microgliardquo Bioorganic amp Medicinal Chemistry vol 19 no 18pp 5559ndash5568 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
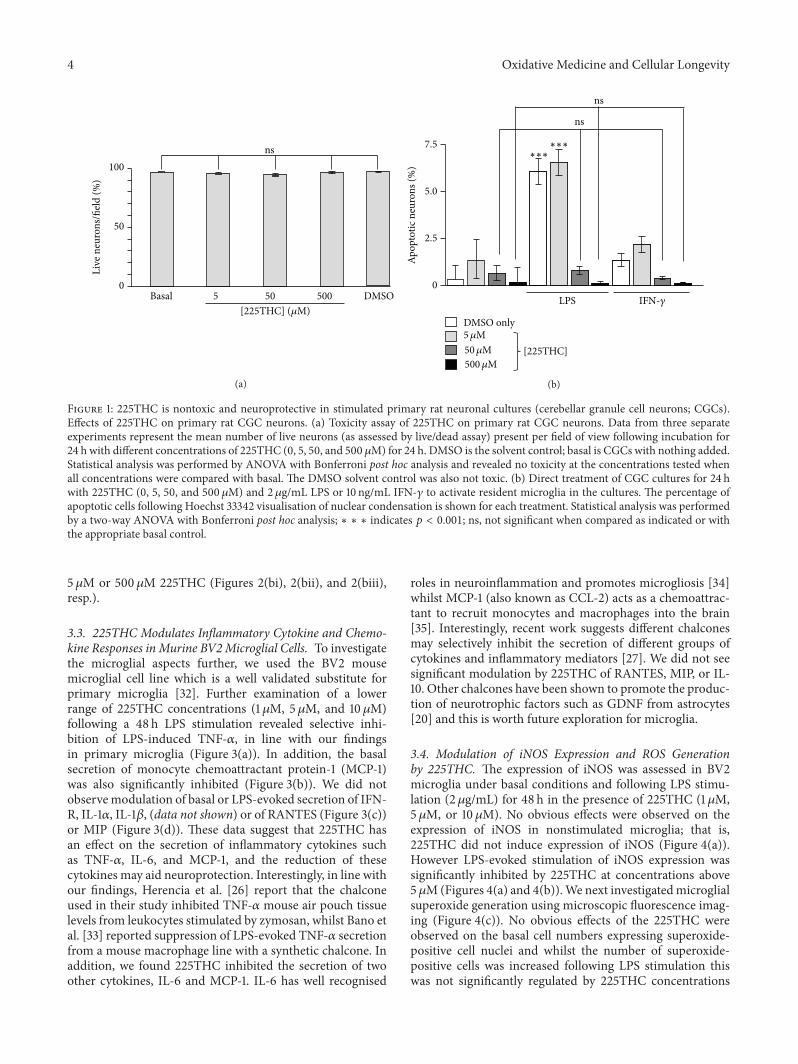
4 Oxidative Medicine and Cellular Longevity
0
100
Live
neu
rons
fiel
d (
)
5 50 500Basal
ns
50
DMSO[225THC] (120583M)
(a)
Apop
totic
neu
rons
()
LPS IFN-1205740
25
50
75
ns
ns
DMSO only
[225THC]
lowastlowastlowastlowastlowastlowast
5120583M
500120583M50120583M
(b)
Figure 1 225THC is nontoxic and neuroprotective in stimulated primary rat neuronal cultures (cerebellar granule cell neurons CGCs)Effects of 225THC on primary rat CGC neurons (a) Toxicity assay of 225THC on primary rat CGC neurons Data from three separateexperiments represent the mean number of live neurons (as assessed by livedead assay) present per field of view following incubation for24 h with different concentrations of 225THC (0 5 50 and 500 120583M) for 24 h DMSO is the solvent control basal is CGCs with nothing addedStatistical analysis was performed by ANOVA with Bonferroni post hoc analysis and revealed no toxicity at the concentrations tested whenall concentrations were compared with basal The DMSO solvent control was also not toxic (b) Direct treatment of CGC cultures for 24 hwith 225THC (0 5 50 and 500120583M) and 2 120583gmL LPS or 10 ngmL IFN-120574 to activate resident microglia in the cultures The percentage ofapoptotic cells following Hoechst 33342 visualisation of nuclear condensation is shown for each treatment Statistical analysis was performedby a two-way ANOVA with Bonferroni post hoc analysis lowast lowast lowast indicates 119901 lt 0001 ns not significant when compared as indicated or withthe appropriate basal control
5 120583M or 500120583M 225THC (Figures 2(bi) 2(bii) and 2(biii)resp)
33 225THC Modulates Inflammatory Cytokine and Chemo-kine Responses in Murine BV2Microglial Cells To investigatethe microglial aspects further we used the BV2 mousemicroglial cell line which is a well validated substitute forprimary microglia [32] Further examination of a lowerrange of 225THC concentrations (1120583M 5 120583M and 10 120583M)following a 48 h LPS stimulation revealed selective inhi-bition of LPS-induced TNF-120572 in line with our findingsin primary microglia (Figure 3(a)) In addition the basalsecretion of monocyte chemoattractant protein-1 (MCP-1)was also significantly inhibited (Figure 3(b)) We did notobservemodulation of basal or LPS-evoked secretion of IFN-R IL-1120572 IL-1120573 (data not shown) or of RANTES (Figure 3(c))or MIP (Figure 3(d)) These data suggest that 225THC hasan effect on the secretion of inflammatory cytokines suchas TNF-120572 IL-6 and MCP-1 and the reduction of thesecytokines may aid neuroprotection Interestingly in line withour findings Herencia et al [26] report that the chalconeused in their study inhibited TNF-120572 mouse air pouch tissuelevels from leukocytes stimulated by zymosan whilst Bano etal [33] reported suppression of LPS-evoked TNF-120572 secretionfrom a mouse macrophage line with a synthetic chalcone Inaddition we found 225THC inhibited the secretion of twoother cytokines IL-6 and MCP-1 IL-6 has well recognised
roles in neuroinflammation and promotes microgliosis [34]whilst MCP-1 (also known as CCL-2) acts as a chemoattrac-tant to recruit monocytes and macrophages into the brain[35] Interestingly recent work suggests different chalconesmay selectively inhibit the secretion of different groups ofcytokines and inflammatory mediators [27] We did not seesignificant modulation by 225THC of RANTES MIP or IL-10 Other chalcones have been shown to promote the produc-tion of neurotrophic factors such as GDNF from astrocytes[20] and this is worth future exploration for microglia
34 Modulation of iNOS Expression and ROS Generationby 225THC The expression of iNOS was assessed in BV2microglia under basal conditions and following LPS stimu-lation (2 120583gmL) for 48 h in the presence of 225THC (1120583M5 120583M or 10 120583M) No obvious effects were observed on theexpression of iNOS in nonstimulated microglia that is225THC did not induce expression of iNOS (Figure 4(a))However LPS-evoked stimulation of iNOS expression wassignificantly inhibited by 225THC at concentrations above5 120583M(Figures 4(a) and 4(b))We next investigatedmicroglialsuperoxide generation using microscopic fluorescence imag-ing (Figure 4(c)) No obvious effects of the 225THC wereobserved on the basal cell numbers expressing superoxide-positive cell nuclei and whilst the number of superoxide-positive cells was increased following LPS stimulation thiswas not significantly regulated by 225THC concentrations
Oxidative Medicine and Cellular Longevity 5
0
200
400
Aver
age n
umbe
r of
cells
fiel
dco
nditi
on
Basa
l 1
1005025105 1
10050105
LPS 2 (120583gmL) [225THC] (120583M) [225THC] (120583M)
Exp 1Exp 2Exp 3
(a)
20
0LPS
0
02
IL-6
(ng
mL)
IL-1
0 (n
gm
L)
IL-6
LPS
IL-10
0
02
BasalBasalBasal LPS
TNF-120572
TNF-120572
(ng
mL)
lowastlowast
lowast
lowastlowast
(biii)(bii)(bi)
5120583M500120583M
5120583M500120583M
5120583M500120583M
(b)
Figure 2 225THC is nontoxic and inhibits inflammatory cytokine responses in stimulated primary rat microglial cultures Effects of 225THCon primary rat microglial cells (a) Toxicity assay of 225THC on primary cultured rat microglia Data shown from three separate experimentsrepresent the mean number of live primary microglia (as assessed by livedead assay) present per field of view following incubation for24 h with different concentrations of 225THC (0 1 5 10 50 and 100 120583M) for 24 h with or without coactivation of the microglia with LPS(2120583gmL) (b) ELISA assessment of secreted cytokines in LPS (2 120583gmL) activated primary rat microglia cultures treated with 225THC at theconcentrations indicated Statistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005lowastlowast119901 lt 001 when compared with basla or as indicated
from 1 to 100 120583M(Figure 4(c)) or at 500120583M(data not shown)The positive control of NADPH oxidase activation PMA(10 ngmL) induced a significant increase in superoxide-positive cells in line with previous findings [22]
The inhibition of iNOS expression in microglia is inagreement with findings for another chalcone 4-dimethyl-amino-3101584041015840-dimethoxychalcone on mouse macrophagesstimulated with zymosan [22] This chalcone inhibited iNOSexpression with a similar concentration (sim10 120583M) to the chal-cone used in the present study Herencia et al [26] proposedthat the effect on iNOS expression was via upstreammodula-tion of NADPH oxidase-mediated intracellular signalling
Since we did not detect modulation of microglial superoxidegeneration by 225THC it seems unlikely that this particularchalcone possesses this mechanism of action Other chal-cones have also been reported to modulate nitric oxide pro-duction or iNOS expression following LPS stimulation of amouse macrophage cell line [36] the BV2 microglial cell line[27] and a rat microglial cell line [37] within similar com-pound concentration ranges of 1ndash50 120583MThe exact signallingpathways inhibited by different chalcones may be diverseas some studies suggest modulation of nuclear factor-120581BMAPK or c-Jun pathways and others suggest STAT1 path-ways may be involved [26 37]
6 Oxidative Medicine and Cellular Longevity
60
220
Abov
e con
trol (
)
Basal LPS
ns
ns
lowastlowast
lowast
lowast lowast
1120583M5120583M10120583M
TNF-120572
(a)
MCP-1
60
220
LPS
ns
ns
lowastlowast
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(b)
Rantes
60
280
LPS
ns
lowast
lowastlowast
lowast
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(c)
LPS
MIP
80
170
ns
ns
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(d)
Figure 3 225THC modulates inflammatory cytokine and chemokine responses in murine BV2 microglial cells 225THC-modulation ofcytokinechemokine secretion by the BV2 microglial cell line ELISA assessment of cytokine ((a) TNF-120572) and chemokine ((b) MCP-1 (c)RANTES (d) MIP) secretion by BV2 microglial cells following stimulation with LPS (2 120583gmL) for 24 h in the presence of 225THC at theconcentrations indicated Statistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005lowastlowast119901 lt 001 ns not significant when compared with basal or as indicated
35 Modulation ofMicroglial Inflammatory Phenotype UponLPS stimulationmicroglia adopt an inflammatory phenotypewhich is characterised by enhanced cell surface expression ofkey adhesion (CD11b CD54) and costimulatory molecules(CD40) We investigated the effects of 225THC treatment
on BV2 cell surface expression of CD11b CD40 CD54 andCD68 in basal and LPS-treated BV2 microglia (Figure 5)Flow cytometric analysis (FACs) of marker mean fluores-cence intensity (MFI) revealed no significant changes in basalCD40 CD54 CD68 or CD11b expression levels following
Oxidative Medicine and Cellular Longevity 7
iNOS
Basal 1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M)[225THC] (120583M)
120573-actin
(a)
0
07
1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
Den
sitom
etry
of i
NO
S120573
-act
in
lowast lowast
(b)
Supe
roxi
de-p
ositi
ve ce
lls(
tota
l)
50
01 5 10 100 1 5 10 100
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
lowastlowast lowastlowastlowastlowast lowastlowast
PMA10ngmL
lowast
lowast
(c)
1120583M LPS + 1120583M
10120583M
100120583M
LPS + 10120583M
LPS + 100120583M
Control LPS
(d)
Figure 4 225THC inhibits the LPS-induced expression of iNOS but not NADPH-driven superoxide production in BV2 microgliaModulation of BV2 iNOS expression but not superoxide production by 225THC Representative Western blots of iNOS expression and 120573-actin as a loading control (a) and densitometry (b) of three separate blots for BV2microglia exposed to 225THC at the stated concentrations plusmnLPS (2120583gmL) for 24 h Statistical analysis was performed by a two-way ANOVAwith Bonferroni post hoc analysis lowast indicates 119901 lt 005whencompared with basal or as indicated (c) Superoxide generation in BV2 microglia measured by dihydroethidium fluorescence following 24 hexposure to 225THC at the concentrations indicated plusmn LPS (2120583gmL) The phorbol PMA was used as a positive activator of superoxidegeneration (d) Representative fields of BV2 microglia generating intracellular superoxide following 24 h exposure to 225THC +minus LPSStatistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005 lowastlowast indicates 119901 lt 001when compared with basal or as indicated
225THC treatment for 48 h (data not shown) However fol-lowing stimulation with 2 120583gmL LPS CD40 (Figure 5(bii))CD54 (Figure 5(cii)) and CD68 (Figure 5(dii)) were signif-icantly upregulated and these changes in expression wereabrogated in the presence of 5 or 10 120583M 225THC CD40appeared to be the most potently regulatedmarker with both5 and 10 120583M 225THC completely inhibiting LPS-inducedupregulation (Figure 5(bii)) while the treatment had moremoderate effects on CD54 and CD68 marker expression As
in the basal state 225THC had no effect on CD11b expressionby LPS-stimulated microglia (Figure 5(aii))
Overall these data suggest that 225THC targets thesignalling pathways downstream of LPS binding and abro-gates the expression of genes such as TNF-120572 iNOS andCD40 Since these are all reported NF-120581B driven genesfuture work could investigate this further to identify theexact pathways modified by 225THC Abrogating this geneexpression produced a less neurotoxic microglial phenotype
8 Oxidative Medicine and Cellular Longevity
CD11b
(ai) (aii)
[225THC] (120583M)
FL1-H
CD11b
Basal LPS 1 5 10
111009080706050403020100
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(a)
CD40
(bi) (bii)
[225THC] (120583M)
FL1-H
CD40
Basal LPS 1 5 10
125
100
075
050
025
000
175
150
lowastlowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(b)
(ci) (cii)
[225THC] (120583M)
FL1-H
CD54 (ICAM-1)CD54 (ICAM-1)
Basal LPS 1 5 10
400
300
200
100
000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(c)
CD68
(di) (dii)
[225THC] (120583M)FL1-H
CD68
Basal LPS 1 5 10
160140120100080060040020000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(d)
Figure 5 225THC treatmentmodulates inflammatory cell surfacemarker expression in LPS-treated BV2microglia Flow cytometric analysisof 225THC-treated BV2 microglia following LPS stimulation (a) Representative histogram plots of BV2 cell surface CD11b CD40 CD54and CD68 expression following 2 120583gmL LPS-treatment for 48 hours (unfilled histograms) versus isotype control (black-filled histograms)and in the presence of 10 120583M 225THC (grey-filled histograms) (b) Summary data showing fold changes in CD11b CD40 CD54 or CD68expression by mean fluorescence intensity (MFI) from 4 independent experiments Data are mean values plusmn SEM where lowast indicates 119901 lt 005by Studentrsquos 119905-test
Oxidative Medicine and Cellular Longevity 9
which is likely to be beneficial for a number of neurodegen-erative diseases in which aberrant microglial inflammatoryreactivity is implicated Future work could pinpoint furtherthe microglial phenotype favoured by 225THC exposure andexplore the genotype of the microglia
Abbreviations
ANOVA Analysis of varianceDPPH 22-Diphenyl-1-picrylhydrazyldHEth DihydroethidiumCGCs Cerebellar granule neuronsDIV Days in vitroDAPI 410158406-Diamidino-2-phenylindoleDMSO DimethylsulphoxideECL ElectrochemiluminescenceELISA Enzyme-linked immunosorbent assayGABAA 120574-Aminobutyric acidIFN-120574 Interferon-120574IL-1120573 Interleukin-1120573IL-6 Interleukin-6IL-10 Interleukin-10iNOS Inducible nitric oxide synthaseLPS LipopolysaccharideMCP-1 CCL2monocyte chemoattractant
protein-1MEM Modified Eaglersquos mediumMIP Macrophage inhibitory proteinMCP-1 Monocyte chemoattractant protein-1NGS Normal goat serumPBS Phosphate buffered salinePFA Paraformaldehyde2-OH-E+ 2-HydroxyethidiumRANTES CCL5regulation on activation normal T
cell expressed and secretedROS Reactive oxygen speciesTNF-120572 Tumour necrosis factor-120572
Conflict of Interests
All authors declare no conflict of interests
Acknowledgments
Manasi Jiwrajka would like to thank Dr Kathleen Susman forher guidance This research was supported by the Universityof Queensland Advantage Grant Howard Hughes MedicalFoundation Grant 52006322 to Vassar College the ShermanFairchild Foundation and Vassar College and by generalfunds to the Pocock lab
References
[1] S Ducki R Forrest J A Hadfield et al ldquoPotent antimitotic andcell growth inhibitory properties of substituted chalconesrdquo Bio-organic and Medicinal Chemistry Letters vol 8 no 9 pp 1051ndash1056 1998
[2] M L F Ferreyra S P Rius and P Casati ldquoFlavonoids biosyn-thesis biological functions and biotechnological applicationsrdquoFrontiers in Plant Science vol 3 article 222 15 pages 2012
[3] G E-D A A Abuo-Rahma M Abdel-Aziz M A E Mouradand H H Farag ldquoSynthesis anti-inflammatory activity andulcerogenic liability of novel nitric oxide donatingchalconehybridsrdquo Bioorganic and Medicinal Chemistry vol 20 no 1 pp195ndash206 2012
[4] N K Sahu S S Balbhadra J Choudhary and D V KohlildquoExploring pharmacological significance of chalcone scaffoldrdquoCurrent Medicinal Chemistry vol 19 no 2 pp 209ndash225 2012
[5] H Wei X Zhang G Wu et al ldquoChalcone derivatives from thefern Cyclosorus parasiticus and their anti-proliferative activityrdquoFood and Chemical Toxicology vol 60 pp 147ndash152 2013
[6] C W Mai M Yaeghoobi N Abd-Rahman Y B Kang andM R Pichika ldquoChalcones with electron-withdrawing and ele-ctron-donating substituents anticancer activity against TRAILresistant cancer cells structure-activity relationship analysisand regulation of apoptotic proteinsrdquo European Journal ofMedicinal Chemistry vol 77 pp 378ndash387 2014
[7] Z Nowakowska ldquoA review of anti-infective and anti-inflam-matory chalconesrdquo European Journal of Medicinal Chemistryvol 42 no 2 pp 125ndash137 2007
[8] Y-M Lin Y Zhou M T Flavin L-M Zhou W Nie and F-CChen ldquoChalcones and flavonoids as anti-tuberculosis agentsrdquoBioorganic and Medicinal Chemistry vol 10 no 8 pp 2795ndash2802 2002
[9] JMojzis L Varinska GMojzisova I Kostova and LMirossayldquoAnti-angiogenic effects of flavonoids and chalconesrdquo Pharma-cological Research vol 57 no 4 pp 259ndash265 2008
[10] F Dal Piaz A Braca M A Belisario and N De TommasildquoThioredoxin systemmodulation by plant and fungal secondarymetabolitesrdquo Current Medicinal Chemistry vol 17 no 5 pp479ndash494 2010
[11] N Yadav S K Dixit A Bhattacharya et al ldquoAntimalarial activ-ity of newly synthesized chalcone derivatives in vitrordquo Chem-ical Biology and Drug Design vol 80 no 2 pp 340ndash347 2012
[12] F V DeFeudis ldquoNovel therapy for Alzheimerrsquos diseaserdquo Neuro-biology of Aging vol 10 no 5 pp 467ndash468 1989
[13] Y Nomura and Y Kitamura ldquoInducible nitric oxide synthasein glial cellsrdquo Neuroscience Research vol 18 no 2 pp 103ndash1071993
[14] P J Kingham M L Cuzner and J M Pocock ldquoApoptotic path-ways mobilized in microglia and neurones as a consequenceof chromogranin A-induced microglial activationrdquo Journal ofNeurochemistry vol 73 no 2 pp 538ndash547 1999
[15] S Singh S Swarnkar P Goswami and C Nath ldquoAstrocytes andmicroglia responses to neuropathological conditionsrdquo Inter-national Journal of Neuroscience vol 121 no 11 pp 589ndash5972011
[16] Y Y Deng J Lu E-A Ling and C Kaur ldquoRole of microglia inthe process of inflammation in the hypoxic developing brainrdquoFrontiers in Bioscience (Scholar Edition) vol 3 no 3 pp 884ndash900 2011
[17] M Dumont and M F Beal ldquoNeuroprotective strategies involv-ing ROS in Alzheimer diseaserdquo Free Radical Biology andMedicine vol 51 no 5 pp 1014ndash1026 2011
[18] AD Kraft andG JeanHarry ldquoFeatures ofmicroglia and neuro-inflammation relevant to environmental exposure and neuro-toxicityrdquo International Journal of Environmental Research andPublic Health vol 8 no 7 pp 2980ndash3018 2011
[19] S Cho S Kim Z Jin et al ldquoIsoliquiritigenin a chalcone com-pound is a positive allosteric modulator of GABA A recep-tors and shows hypnotic effectsrdquo Biochemical and BiophysicalResearch Communications vol 413 no 4 pp 637ndash642 2011
10 Oxidative Medicine and Cellular Longevity
[20] A Elmann A Telerman H Erlank et al ldquoProtective and anti-oxidant effects of a chalconoid from Pulicaria incisa on brainastrocytesrdquoOxidativeMedicine andCellular Longevity vol 2013Article ID 694398 10 pages 2013
[21] M Rossi F Caruso E J Crespi et al ldquoProbing antioxidantactivity of 21015840-hydroxychalcones crystal and molecular struc-tures in vitro antiproliferative studies and in vivo effects onglucose regulationrdquo Biochimie vol 95 no 10 pp 1954ndash19632013
[22] E LMead AMosley S Eaton L Dobson S J Heales and JMPocock ldquoMicroglial neurotransmitter receptors trigger super-oxide production in microglia consequences for microglial-neuronal interactionsrdquo Journal of Neurochemistry vol 121 no2 pp 287ndash301 2012
[23] C M Davenport I G Sevastou C Hooper and J M PocockldquoInhibiting p53 pathways in microglia attenuates microglial-evoked neurotoxicity following exposure to Alzheimer pep-tidesrdquo Journal of Neurochemistry vol 112 no 2 pp 552ndash5632010
[24] N C Garbett N B Hammond and D E Graves ldquoInfluence ofthe amino substituents in the interaction of ethidium bromidewith DNArdquo Biophysical Journal vol 87 no 6 pp 3974ndash39812004
[25] A Karlsson J B Nixon and L C McPhail ldquoPhorbol myristateacetate induces neutrophilNADPH-oxidase activity by two sep-arate signal transduction pathways dependent or independentof phosphatidylinositol 3-kinaserdquo Journal of Leukocyte Biologyvol 67 no 3 pp 396ndash404 2000
[26] F Herencia M L Ferrandiz A Ubeda et al ldquo4-Dimethyl-amino-3101584041015840-dimethoxychalcone down-regulates iNOS expres-sion and exerts anti-inflammatory effectsrdquo Free Radical Biologyamp Medicine vol 30 no 1 pp 43ndash50 2001
[27] N Mateeva M Gangapuram E Mazzio S Eyunni K FSoliman and K K Redda ldquoBiological evaluation of syntheticchalcone and flavone derivatives as anti-inflammatory agentsrdquoMedicinal Chemistry Research vol 24 no 4 pp 1672ndash16802015
[28] S C Morgan D L Taylor and J M Pocock ldquoMicroglia releaseactivators of neuronal proliferation mediated by activationof mitogen-activated protein kinase phosphatidylinositol-3-kinaseAkt and delta-Notch signalling cascadesrdquo Journal ofNeurochemistry vol 90 no 1 pp 89ndash101 2004
[29] J Jebelli T Piers and J Pocock ldquoSelective depletion of micro-glia from cerebellar granule cell cultures using L-leucinemethylesterrdquoThe Journal of Visualized Experiments no 101 Article IDe52983 2015
[30] R Von Bernhardi J E Tichauer and J Eugenın ldquoAging-dependent changes of microglial cells and their relevance forneurodegenerative disordersrdquo Journal of Neurochemistry vol112 no 5 pp 1099ndash1114 2010
[31] C Schwab and P L McGeer ldquoInflammatory aspects ofAlzheimer disease and other neurodegenerative disordersrdquoJournal of Alzheimerrsquos Disease vol 13 no 4 pp 359ndash369 2008
[32] A Henn S Lund M Hedtjarn A Schrattenholz P Porzgenand M Leist ldquoThe suitability of BV2 cells as alternative modelsystem for primary microglia cultures or for animal experi-ments examining brain inflammationrdquo Altex vol 26 no 2 pp83ndash94 2009
[33] S Bano K Javed S Ahmad et al ldquoSynthesis of some novel chal-cones flavanones and flavones and evaluation of their anti-inflammatory activityrdquo European Journal of Medicinal Chem-istry vol 65 pp 51ndash59 2013
[34] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[35] S L Deshmane S Kremlev S Amini and B E Sawaya ldquoMono-cyte chemoattractant protein-1 (MCP-1) an overviewrdquo Journalof Interferon and Cytokine Research vol 29 no 6 pp 313ndash3262009
[36] J Rojas J N Domınguez J E Charris G Lobo M Paya andM L Ferrandiz ldquoSynthesis and inhibitory activity of dimethyl-amino-chalcone derivatives on the induction of nitric oxidesynthaserdquo European Journal of Medicinal Chemistry vol 37 no8 pp 699ndash705 2002
[37] H Hara Y Nakamura M Ninomiya et al ldquoInhibitory effectsof chalcone glycosides isolated from Brassica rapa L lsquohidabenirsquoand their synthetic derivatives on LPS-induced NO productionin microgliardquo Bioorganic amp Medicinal Chemistry vol 19 no 18pp 5559ndash5568 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
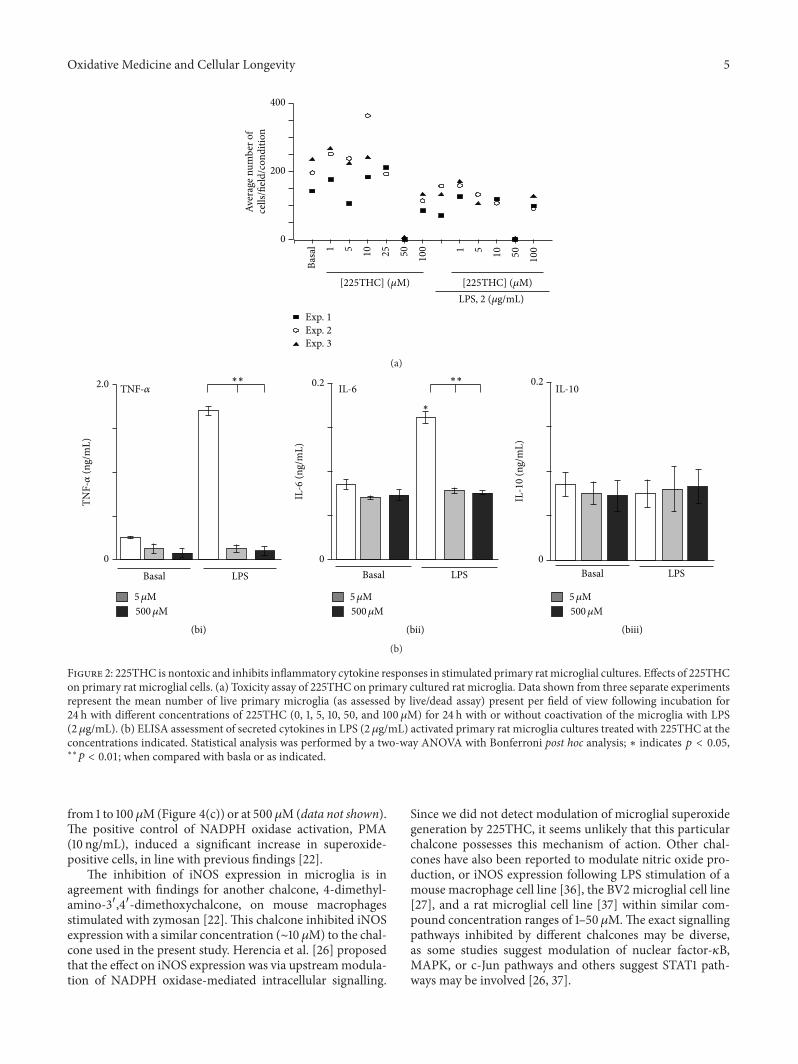
Oxidative Medicine and Cellular Longevity 5
0
200
400
Aver
age n
umbe
r of
cells
fiel
dco
nditi
on
Basa
l 1
1005025105 1
10050105
LPS 2 (120583gmL) [225THC] (120583M) [225THC] (120583M)
Exp 1Exp 2Exp 3
(a)
20
0LPS
0
02
IL-6
(ng
mL)
IL-1
0 (n
gm
L)
IL-6
LPS
IL-10
0
02
BasalBasalBasal LPS
TNF-120572
TNF-120572
(ng
mL)
lowastlowast
lowast
lowastlowast
(biii)(bii)(bi)
5120583M500120583M
5120583M500120583M
5120583M500120583M
(b)
Figure 2 225THC is nontoxic and inhibits inflammatory cytokine responses in stimulated primary rat microglial cultures Effects of 225THCon primary rat microglial cells (a) Toxicity assay of 225THC on primary cultured rat microglia Data shown from three separate experimentsrepresent the mean number of live primary microglia (as assessed by livedead assay) present per field of view following incubation for24 h with different concentrations of 225THC (0 1 5 10 50 and 100 120583M) for 24 h with or without coactivation of the microglia with LPS(2120583gmL) (b) ELISA assessment of secreted cytokines in LPS (2 120583gmL) activated primary rat microglia cultures treated with 225THC at theconcentrations indicated Statistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005lowastlowast119901 lt 001 when compared with basla or as indicated
from 1 to 100 120583M(Figure 4(c)) or at 500120583M(data not shown)The positive control of NADPH oxidase activation PMA(10 ngmL) induced a significant increase in superoxide-positive cells in line with previous findings [22]
The inhibition of iNOS expression in microglia is inagreement with findings for another chalcone 4-dimethyl-amino-3101584041015840-dimethoxychalcone on mouse macrophagesstimulated with zymosan [22] This chalcone inhibited iNOSexpression with a similar concentration (sim10 120583M) to the chal-cone used in the present study Herencia et al [26] proposedthat the effect on iNOS expression was via upstreammodula-tion of NADPH oxidase-mediated intracellular signalling
Since we did not detect modulation of microglial superoxidegeneration by 225THC it seems unlikely that this particularchalcone possesses this mechanism of action Other chal-cones have also been reported to modulate nitric oxide pro-duction or iNOS expression following LPS stimulation of amouse macrophage cell line [36] the BV2 microglial cell line[27] and a rat microglial cell line [37] within similar com-pound concentration ranges of 1ndash50 120583MThe exact signallingpathways inhibited by different chalcones may be diverseas some studies suggest modulation of nuclear factor-120581BMAPK or c-Jun pathways and others suggest STAT1 path-ways may be involved [26 37]
6 Oxidative Medicine and Cellular Longevity
60
220
Abov
e con
trol (
)
Basal LPS
ns
ns
lowastlowast
lowast
lowast lowast
1120583M5120583M10120583M
TNF-120572
(a)
MCP-1
60
220
LPS
ns
ns
lowastlowast
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(b)
Rantes
60
280
LPS
ns
lowast
lowastlowast
lowast
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(c)
LPS
MIP
80
170
ns
ns
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(d)
Figure 3 225THC modulates inflammatory cytokine and chemokine responses in murine BV2 microglial cells 225THC-modulation ofcytokinechemokine secretion by the BV2 microglial cell line ELISA assessment of cytokine ((a) TNF-120572) and chemokine ((b) MCP-1 (c)RANTES (d) MIP) secretion by BV2 microglial cells following stimulation with LPS (2 120583gmL) for 24 h in the presence of 225THC at theconcentrations indicated Statistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005lowastlowast119901 lt 001 ns not significant when compared with basal or as indicated
35 Modulation ofMicroglial Inflammatory Phenotype UponLPS stimulationmicroglia adopt an inflammatory phenotypewhich is characterised by enhanced cell surface expression ofkey adhesion (CD11b CD54) and costimulatory molecules(CD40) We investigated the effects of 225THC treatment
on BV2 cell surface expression of CD11b CD40 CD54 andCD68 in basal and LPS-treated BV2 microglia (Figure 5)Flow cytometric analysis (FACs) of marker mean fluores-cence intensity (MFI) revealed no significant changes in basalCD40 CD54 CD68 or CD11b expression levels following
Oxidative Medicine and Cellular Longevity 7
iNOS
Basal 1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M)[225THC] (120583M)
120573-actin
(a)
0
07
1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
Den
sitom
etry
of i
NO
S120573
-act
in
lowast lowast
(b)
Supe
roxi
de-p
ositi
ve ce
lls(
tota
l)
50
01 5 10 100 1 5 10 100
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
lowastlowast lowastlowastlowastlowast lowastlowast
PMA10ngmL
lowast
lowast
(c)
1120583M LPS + 1120583M
10120583M
100120583M
LPS + 10120583M
LPS + 100120583M
Control LPS
(d)
Figure 4 225THC inhibits the LPS-induced expression of iNOS but not NADPH-driven superoxide production in BV2 microgliaModulation of BV2 iNOS expression but not superoxide production by 225THC Representative Western blots of iNOS expression and 120573-actin as a loading control (a) and densitometry (b) of three separate blots for BV2microglia exposed to 225THC at the stated concentrations plusmnLPS (2120583gmL) for 24 h Statistical analysis was performed by a two-way ANOVAwith Bonferroni post hoc analysis lowast indicates 119901 lt 005whencompared with basal or as indicated (c) Superoxide generation in BV2 microglia measured by dihydroethidium fluorescence following 24 hexposure to 225THC at the concentrations indicated plusmn LPS (2120583gmL) The phorbol PMA was used as a positive activator of superoxidegeneration (d) Representative fields of BV2 microglia generating intracellular superoxide following 24 h exposure to 225THC +minus LPSStatistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005 lowastlowast indicates 119901 lt 001when compared with basal or as indicated
225THC treatment for 48 h (data not shown) However fol-lowing stimulation with 2 120583gmL LPS CD40 (Figure 5(bii))CD54 (Figure 5(cii)) and CD68 (Figure 5(dii)) were signif-icantly upregulated and these changes in expression wereabrogated in the presence of 5 or 10 120583M 225THC CD40appeared to be the most potently regulatedmarker with both5 and 10 120583M 225THC completely inhibiting LPS-inducedupregulation (Figure 5(bii)) while the treatment had moremoderate effects on CD54 and CD68 marker expression As
in the basal state 225THC had no effect on CD11b expressionby LPS-stimulated microglia (Figure 5(aii))
Overall these data suggest that 225THC targets thesignalling pathways downstream of LPS binding and abro-gates the expression of genes such as TNF-120572 iNOS andCD40 Since these are all reported NF-120581B driven genesfuture work could investigate this further to identify theexact pathways modified by 225THC Abrogating this geneexpression produced a less neurotoxic microglial phenotype
8 Oxidative Medicine and Cellular Longevity
CD11b
(ai) (aii)
[225THC] (120583M)
FL1-H
CD11b
Basal LPS 1 5 10
111009080706050403020100
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(a)
CD40
(bi) (bii)
[225THC] (120583M)
FL1-H
CD40
Basal LPS 1 5 10
125
100
075
050
025
000
175
150
lowastlowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(b)
(ci) (cii)
[225THC] (120583M)
FL1-H
CD54 (ICAM-1)CD54 (ICAM-1)
Basal LPS 1 5 10
400
300
200
100
000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(c)
CD68
(di) (dii)
[225THC] (120583M)FL1-H
CD68
Basal LPS 1 5 10
160140120100080060040020000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(d)
Figure 5 225THC treatmentmodulates inflammatory cell surfacemarker expression in LPS-treated BV2microglia Flow cytometric analysisof 225THC-treated BV2 microglia following LPS stimulation (a) Representative histogram plots of BV2 cell surface CD11b CD40 CD54and CD68 expression following 2 120583gmL LPS-treatment for 48 hours (unfilled histograms) versus isotype control (black-filled histograms)and in the presence of 10 120583M 225THC (grey-filled histograms) (b) Summary data showing fold changes in CD11b CD40 CD54 or CD68expression by mean fluorescence intensity (MFI) from 4 independent experiments Data are mean values plusmn SEM where lowast indicates 119901 lt 005by Studentrsquos 119905-test
Oxidative Medicine and Cellular Longevity 9
which is likely to be beneficial for a number of neurodegen-erative diseases in which aberrant microglial inflammatoryreactivity is implicated Future work could pinpoint furtherthe microglial phenotype favoured by 225THC exposure andexplore the genotype of the microglia
Abbreviations
ANOVA Analysis of varianceDPPH 22-Diphenyl-1-picrylhydrazyldHEth DihydroethidiumCGCs Cerebellar granule neuronsDIV Days in vitroDAPI 410158406-Diamidino-2-phenylindoleDMSO DimethylsulphoxideECL ElectrochemiluminescenceELISA Enzyme-linked immunosorbent assayGABAA 120574-Aminobutyric acidIFN-120574 Interferon-120574IL-1120573 Interleukin-1120573IL-6 Interleukin-6IL-10 Interleukin-10iNOS Inducible nitric oxide synthaseLPS LipopolysaccharideMCP-1 CCL2monocyte chemoattractant
protein-1MEM Modified Eaglersquos mediumMIP Macrophage inhibitory proteinMCP-1 Monocyte chemoattractant protein-1NGS Normal goat serumPBS Phosphate buffered salinePFA Paraformaldehyde2-OH-E+ 2-HydroxyethidiumRANTES CCL5regulation on activation normal T
cell expressed and secretedROS Reactive oxygen speciesTNF-120572 Tumour necrosis factor-120572
Conflict of Interests
All authors declare no conflict of interests
Acknowledgments
Manasi Jiwrajka would like to thank Dr Kathleen Susman forher guidance This research was supported by the Universityof Queensland Advantage Grant Howard Hughes MedicalFoundation Grant 52006322 to Vassar College the ShermanFairchild Foundation and Vassar College and by generalfunds to the Pocock lab
References
[1] S Ducki R Forrest J A Hadfield et al ldquoPotent antimitotic andcell growth inhibitory properties of substituted chalconesrdquo Bio-organic and Medicinal Chemistry Letters vol 8 no 9 pp 1051ndash1056 1998
[2] M L F Ferreyra S P Rius and P Casati ldquoFlavonoids biosyn-thesis biological functions and biotechnological applicationsrdquoFrontiers in Plant Science vol 3 article 222 15 pages 2012
[3] G E-D A A Abuo-Rahma M Abdel-Aziz M A E Mouradand H H Farag ldquoSynthesis anti-inflammatory activity andulcerogenic liability of novel nitric oxide donatingchalconehybridsrdquo Bioorganic and Medicinal Chemistry vol 20 no 1 pp195ndash206 2012
[4] N K Sahu S S Balbhadra J Choudhary and D V KohlildquoExploring pharmacological significance of chalcone scaffoldrdquoCurrent Medicinal Chemistry vol 19 no 2 pp 209ndash225 2012
[5] H Wei X Zhang G Wu et al ldquoChalcone derivatives from thefern Cyclosorus parasiticus and their anti-proliferative activityrdquoFood and Chemical Toxicology vol 60 pp 147ndash152 2013
[6] C W Mai M Yaeghoobi N Abd-Rahman Y B Kang andM R Pichika ldquoChalcones with electron-withdrawing and ele-ctron-donating substituents anticancer activity against TRAILresistant cancer cells structure-activity relationship analysisand regulation of apoptotic proteinsrdquo European Journal ofMedicinal Chemistry vol 77 pp 378ndash387 2014
[7] Z Nowakowska ldquoA review of anti-infective and anti-inflam-matory chalconesrdquo European Journal of Medicinal Chemistryvol 42 no 2 pp 125ndash137 2007
[8] Y-M Lin Y Zhou M T Flavin L-M Zhou W Nie and F-CChen ldquoChalcones and flavonoids as anti-tuberculosis agentsrdquoBioorganic and Medicinal Chemistry vol 10 no 8 pp 2795ndash2802 2002
[9] JMojzis L Varinska GMojzisova I Kostova and LMirossayldquoAnti-angiogenic effects of flavonoids and chalconesrdquo Pharma-cological Research vol 57 no 4 pp 259ndash265 2008
[10] F Dal Piaz A Braca M A Belisario and N De TommasildquoThioredoxin systemmodulation by plant and fungal secondarymetabolitesrdquo Current Medicinal Chemistry vol 17 no 5 pp479ndash494 2010
[11] N Yadav S K Dixit A Bhattacharya et al ldquoAntimalarial activ-ity of newly synthesized chalcone derivatives in vitrordquo Chem-ical Biology and Drug Design vol 80 no 2 pp 340ndash347 2012
[12] F V DeFeudis ldquoNovel therapy for Alzheimerrsquos diseaserdquo Neuro-biology of Aging vol 10 no 5 pp 467ndash468 1989
[13] Y Nomura and Y Kitamura ldquoInducible nitric oxide synthasein glial cellsrdquo Neuroscience Research vol 18 no 2 pp 103ndash1071993
[14] P J Kingham M L Cuzner and J M Pocock ldquoApoptotic path-ways mobilized in microglia and neurones as a consequenceof chromogranin A-induced microglial activationrdquo Journal ofNeurochemistry vol 73 no 2 pp 538ndash547 1999
[15] S Singh S Swarnkar P Goswami and C Nath ldquoAstrocytes andmicroglia responses to neuropathological conditionsrdquo Inter-national Journal of Neuroscience vol 121 no 11 pp 589ndash5972011
[16] Y Y Deng J Lu E-A Ling and C Kaur ldquoRole of microglia inthe process of inflammation in the hypoxic developing brainrdquoFrontiers in Bioscience (Scholar Edition) vol 3 no 3 pp 884ndash900 2011
[17] M Dumont and M F Beal ldquoNeuroprotective strategies involv-ing ROS in Alzheimer diseaserdquo Free Radical Biology andMedicine vol 51 no 5 pp 1014ndash1026 2011
[18] AD Kraft andG JeanHarry ldquoFeatures ofmicroglia and neuro-inflammation relevant to environmental exposure and neuro-toxicityrdquo International Journal of Environmental Research andPublic Health vol 8 no 7 pp 2980ndash3018 2011
[19] S Cho S Kim Z Jin et al ldquoIsoliquiritigenin a chalcone com-pound is a positive allosteric modulator of GABA A recep-tors and shows hypnotic effectsrdquo Biochemical and BiophysicalResearch Communications vol 413 no 4 pp 637ndash642 2011
10 Oxidative Medicine and Cellular Longevity
[20] A Elmann A Telerman H Erlank et al ldquoProtective and anti-oxidant effects of a chalconoid from Pulicaria incisa on brainastrocytesrdquoOxidativeMedicine andCellular Longevity vol 2013Article ID 694398 10 pages 2013
[21] M Rossi F Caruso E J Crespi et al ldquoProbing antioxidantactivity of 21015840-hydroxychalcones crystal and molecular struc-tures in vitro antiproliferative studies and in vivo effects onglucose regulationrdquo Biochimie vol 95 no 10 pp 1954ndash19632013
[22] E LMead AMosley S Eaton L Dobson S J Heales and JMPocock ldquoMicroglial neurotransmitter receptors trigger super-oxide production in microglia consequences for microglial-neuronal interactionsrdquo Journal of Neurochemistry vol 121 no2 pp 287ndash301 2012
[23] C M Davenport I G Sevastou C Hooper and J M PocockldquoInhibiting p53 pathways in microglia attenuates microglial-evoked neurotoxicity following exposure to Alzheimer pep-tidesrdquo Journal of Neurochemistry vol 112 no 2 pp 552ndash5632010
[24] N C Garbett N B Hammond and D E Graves ldquoInfluence ofthe amino substituents in the interaction of ethidium bromidewith DNArdquo Biophysical Journal vol 87 no 6 pp 3974ndash39812004
[25] A Karlsson J B Nixon and L C McPhail ldquoPhorbol myristateacetate induces neutrophilNADPH-oxidase activity by two sep-arate signal transduction pathways dependent or independentof phosphatidylinositol 3-kinaserdquo Journal of Leukocyte Biologyvol 67 no 3 pp 396ndash404 2000
[26] F Herencia M L Ferrandiz A Ubeda et al ldquo4-Dimethyl-amino-3101584041015840-dimethoxychalcone down-regulates iNOS expres-sion and exerts anti-inflammatory effectsrdquo Free Radical Biologyamp Medicine vol 30 no 1 pp 43ndash50 2001
[27] N Mateeva M Gangapuram E Mazzio S Eyunni K FSoliman and K K Redda ldquoBiological evaluation of syntheticchalcone and flavone derivatives as anti-inflammatory agentsrdquoMedicinal Chemistry Research vol 24 no 4 pp 1672ndash16802015
[28] S C Morgan D L Taylor and J M Pocock ldquoMicroglia releaseactivators of neuronal proliferation mediated by activationof mitogen-activated protein kinase phosphatidylinositol-3-kinaseAkt and delta-Notch signalling cascadesrdquo Journal ofNeurochemistry vol 90 no 1 pp 89ndash101 2004
[29] J Jebelli T Piers and J Pocock ldquoSelective depletion of micro-glia from cerebellar granule cell cultures using L-leucinemethylesterrdquoThe Journal of Visualized Experiments no 101 Article IDe52983 2015
[30] R Von Bernhardi J E Tichauer and J Eugenın ldquoAging-dependent changes of microglial cells and their relevance forneurodegenerative disordersrdquo Journal of Neurochemistry vol112 no 5 pp 1099ndash1114 2010
[31] C Schwab and P L McGeer ldquoInflammatory aspects ofAlzheimer disease and other neurodegenerative disordersrdquoJournal of Alzheimerrsquos Disease vol 13 no 4 pp 359ndash369 2008
[32] A Henn S Lund M Hedtjarn A Schrattenholz P Porzgenand M Leist ldquoThe suitability of BV2 cells as alternative modelsystem for primary microglia cultures or for animal experi-ments examining brain inflammationrdquo Altex vol 26 no 2 pp83ndash94 2009
[33] S Bano K Javed S Ahmad et al ldquoSynthesis of some novel chal-cones flavanones and flavones and evaluation of their anti-inflammatory activityrdquo European Journal of Medicinal Chem-istry vol 65 pp 51ndash59 2013
[34] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[35] S L Deshmane S Kremlev S Amini and B E Sawaya ldquoMono-cyte chemoattractant protein-1 (MCP-1) an overviewrdquo Journalof Interferon and Cytokine Research vol 29 no 6 pp 313ndash3262009
[36] J Rojas J N Domınguez J E Charris G Lobo M Paya andM L Ferrandiz ldquoSynthesis and inhibitory activity of dimethyl-amino-chalcone derivatives on the induction of nitric oxidesynthaserdquo European Journal of Medicinal Chemistry vol 37 no8 pp 699ndash705 2002
[37] H Hara Y Nakamura M Ninomiya et al ldquoInhibitory effectsof chalcone glycosides isolated from Brassica rapa L lsquohidabenirsquoand their synthetic derivatives on LPS-induced NO productionin microgliardquo Bioorganic amp Medicinal Chemistry vol 19 no 18pp 5559ndash5568 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
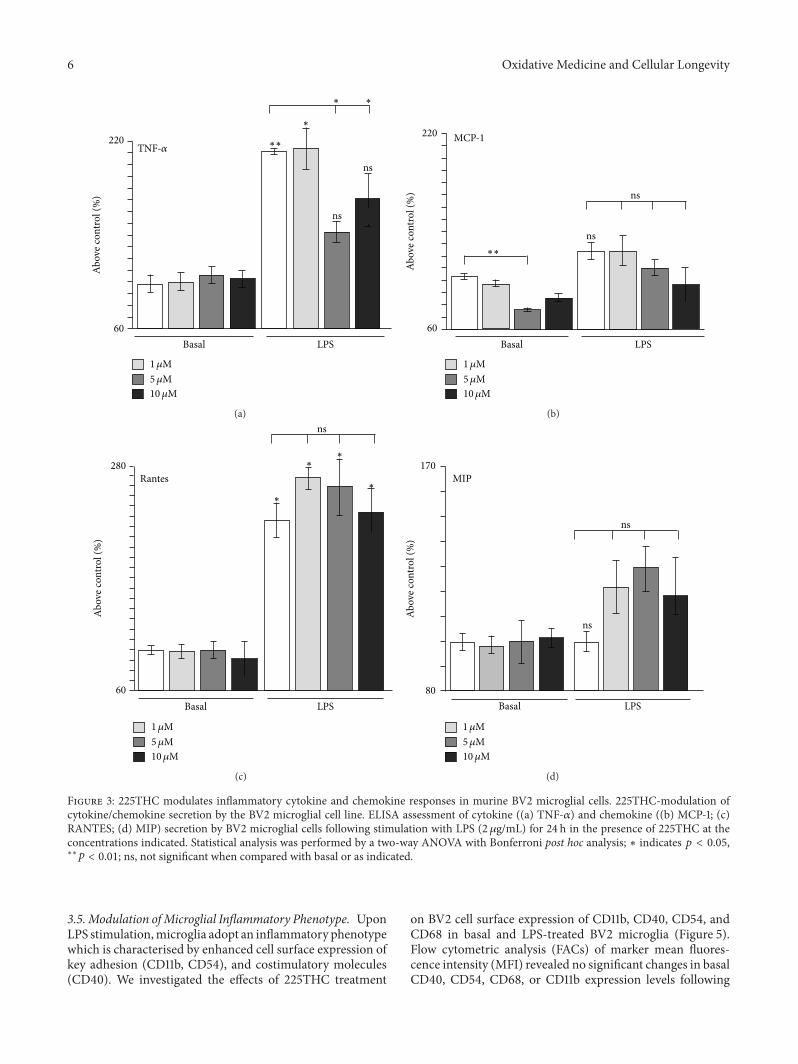
6 Oxidative Medicine and Cellular Longevity
60
220
Abov
e con
trol (
)
Basal LPS
ns
ns
lowastlowast
lowast
lowast lowast
1120583M5120583M10120583M
TNF-120572
(a)
MCP-1
60
220
LPS
ns
ns
lowastlowast
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(b)
Rantes
60
280
LPS
ns
lowast
lowastlowast
lowast
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(c)
LPS
MIP
80
170
ns
ns
Abov
e con
trol (
)
Basal
1120583M5120583M10120583M
(d)
Figure 3 225THC modulates inflammatory cytokine and chemokine responses in murine BV2 microglial cells 225THC-modulation ofcytokinechemokine secretion by the BV2 microglial cell line ELISA assessment of cytokine ((a) TNF-120572) and chemokine ((b) MCP-1 (c)RANTES (d) MIP) secretion by BV2 microglial cells following stimulation with LPS (2 120583gmL) for 24 h in the presence of 225THC at theconcentrations indicated Statistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005lowastlowast119901 lt 001 ns not significant when compared with basal or as indicated
35 Modulation ofMicroglial Inflammatory Phenotype UponLPS stimulationmicroglia adopt an inflammatory phenotypewhich is characterised by enhanced cell surface expression ofkey adhesion (CD11b CD54) and costimulatory molecules(CD40) We investigated the effects of 225THC treatment
on BV2 cell surface expression of CD11b CD40 CD54 andCD68 in basal and LPS-treated BV2 microglia (Figure 5)Flow cytometric analysis (FACs) of marker mean fluores-cence intensity (MFI) revealed no significant changes in basalCD40 CD54 CD68 or CD11b expression levels following
Oxidative Medicine and Cellular Longevity 7
iNOS
Basal 1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M)[225THC] (120583M)
120573-actin
(a)
0
07
1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
Den
sitom
etry
of i
NO
S120573
-act
in
lowast lowast
(b)
Supe
roxi
de-p
ositi
ve ce
lls(
tota
l)
50
01 5 10 100 1 5 10 100
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
lowastlowast lowastlowastlowastlowast lowastlowast
PMA10ngmL
lowast
lowast
(c)
1120583M LPS + 1120583M
10120583M
100120583M
LPS + 10120583M
LPS + 100120583M
Control LPS
(d)
Figure 4 225THC inhibits the LPS-induced expression of iNOS but not NADPH-driven superoxide production in BV2 microgliaModulation of BV2 iNOS expression but not superoxide production by 225THC Representative Western blots of iNOS expression and 120573-actin as a loading control (a) and densitometry (b) of three separate blots for BV2microglia exposed to 225THC at the stated concentrations plusmnLPS (2120583gmL) for 24 h Statistical analysis was performed by a two-way ANOVAwith Bonferroni post hoc analysis lowast indicates 119901 lt 005whencompared with basal or as indicated (c) Superoxide generation in BV2 microglia measured by dihydroethidium fluorescence following 24 hexposure to 225THC at the concentrations indicated plusmn LPS (2120583gmL) The phorbol PMA was used as a positive activator of superoxidegeneration (d) Representative fields of BV2 microglia generating intracellular superoxide following 24 h exposure to 225THC +minus LPSStatistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005 lowastlowast indicates 119901 lt 001when compared with basal or as indicated
225THC treatment for 48 h (data not shown) However fol-lowing stimulation with 2 120583gmL LPS CD40 (Figure 5(bii))CD54 (Figure 5(cii)) and CD68 (Figure 5(dii)) were signif-icantly upregulated and these changes in expression wereabrogated in the presence of 5 or 10 120583M 225THC CD40appeared to be the most potently regulatedmarker with both5 and 10 120583M 225THC completely inhibiting LPS-inducedupregulation (Figure 5(bii)) while the treatment had moremoderate effects on CD54 and CD68 marker expression As
in the basal state 225THC had no effect on CD11b expressionby LPS-stimulated microglia (Figure 5(aii))
Overall these data suggest that 225THC targets thesignalling pathways downstream of LPS binding and abro-gates the expression of genes such as TNF-120572 iNOS andCD40 Since these are all reported NF-120581B driven genesfuture work could investigate this further to identify theexact pathways modified by 225THC Abrogating this geneexpression produced a less neurotoxic microglial phenotype
8 Oxidative Medicine and Cellular Longevity
CD11b
(ai) (aii)
[225THC] (120583M)
FL1-H
CD11b
Basal LPS 1 5 10
111009080706050403020100
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(a)
CD40
(bi) (bii)
[225THC] (120583M)
FL1-H
CD40
Basal LPS 1 5 10
125
100
075
050
025
000
175
150
lowastlowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(b)
(ci) (cii)
[225THC] (120583M)
FL1-H
CD54 (ICAM-1)CD54 (ICAM-1)
Basal LPS 1 5 10
400
300
200
100
000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(c)
CD68
(di) (dii)
[225THC] (120583M)FL1-H
CD68
Basal LPS 1 5 10
160140120100080060040020000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(d)
Figure 5 225THC treatmentmodulates inflammatory cell surfacemarker expression in LPS-treated BV2microglia Flow cytometric analysisof 225THC-treated BV2 microglia following LPS stimulation (a) Representative histogram plots of BV2 cell surface CD11b CD40 CD54and CD68 expression following 2 120583gmL LPS-treatment for 48 hours (unfilled histograms) versus isotype control (black-filled histograms)and in the presence of 10 120583M 225THC (grey-filled histograms) (b) Summary data showing fold changes in CD11b CD40 CD54 or CD68expression by mean fluorescence intensity (MFI) from 4 independent experiments Data are mean values plusmn SEM where lowast indicates 119901 lt 005by Studentrsquos 119905-test
Oxidative Medicine and Cellular Longevity 9
which is likely to be beneficial for a number of neurodegen-erative diseases in which aberrant microglial inflammatoryreactivity is implicated Future work could pinpoint furtherthe microglial phenotype favoured by 225THC exposure andexplore the genotype of the microglia
Abbreviations
ANOVA Analysis of varianceDPPH 22-Diphenyl-1-picrylhydrazyldHEth DihydroethidiumCGCs Cerebellar granule neuronsDIV Days in vitroDAPI 410158406-Diamidino-2-phenylindoleDMSO DimethylsulphoxideECL ElectrochemiluminescenceELISA Enzyme-linked immunosorbent assayGABAA 120574-Aminobutyric acidIFN-120574 Interferon-120574IL-1120573 Interleukin-1120573IL-6 Interleukin-6IL-10 Interleukin-10iNOS Inducible nitric oxide synthaseLPS LipopolysaccharideMCP-1 CCL2monocyte chemoattractant
protein-1MEM Modified Eaglersquos mediumMIP Macrophage inhibitory proteinMCP-1 Monocyte chemoattractant protein-1NGS Normal goat serumPBS Phosphate buffered salinePFA Paraformaldehyde2-OH-E+ 2-HydroxyethidiumRANTES CCL5regulation on activation normal T
cell expressed and secretedROS Reactive oxygen speciesTNF-120572 Tumour necrosis factor-120572
Conflict of Interests
All authors declare no conflict of interests
Acknowledgments
Manasi Jiwrajka would like to thank Dr Kathleen Susman forher guidance This research was supported by the Universityof Queensland Advantage Grant Howard Hughes MedicalFoundation Grant 52006322 to Vassar College the ShermanFairchild Foundation and Vassar College and by generalfunds to the Pocock lab
References
[1] S Ducki R Forrest J A Hadfield et al ldquoPotent antimitotic andcell growth inhibitory properties of substituted chalconesrdquo Bio-organic and Medicinal Chemistry Letters vol 8 no 9 pp 1051ndash1056 1998
[2] M L F Ferreyra S P Rius and P Casati ldquoFlavonoids biosyn-thesis biological functions and biotechnological applicationsrdquoFrontiers in Plant Science vol 3 article 222 15 pages 2012
[3] G E-D A A Abuo-Rahma M Abdel-Aziz M A E Mouradand H H Farag ldquoSynthesis anti-inflammatory activity andulcerogenic liability of novel nitric oxide donatingchalconehybridsrdquo Bioorganic and Medicinal Chemistry vol 20 no 1 pp195ndash206 2012
[4] N K Sahu S S Balbhadra J Choudhary and D V KohlildquoExploring pharmacological significance of chalcone scaffoldrdquoCurrent Medicinal Chemistry vol 19 no 2 pp 209ndash225 2012
[5] H Wei X Zhang G Wu et al ldquoChalcone derivatives from thefern Cyclosorus parasiticus and their anti-proliferative activityrdquoFood and Chemical Toxicology vol 60 pp 147ndash152 2013
[6] C W Mai M Yaeghoobi N Abd-Rahman Y B Kang andM R Pichika ldquoChalcones with electron-withdrawing and ele-ctron-donating substituents anticancer activity against TRAILresistant cancer cells structure-activity relationship analysisand regulation of apoptotic proteinsrdquo European Journal ofMedicinal Chemistry vol 77 pp 378ndash387 2014
[7] Z Nowakowska ldquoA review of anti-infective and anti-inflam-matory chalconesrdquo European Journal of Medicinal Chemistryvol 42 no 2 pp 125ndash137 2007
[8] Y-M Lin Y Zhou M T Flavin L-M Zhou W Nie and F-CChen ldquoChalcones and flavonoids as anti-tuberculosis agentsrdquoBioorganic and Medicinal Chemistry vol 10 no 8 pp 2795ndash2802 2002
[9] JMojzis L Varinska GMojzisova I Kostova and LMirossayldquoAnti-angiogenic effects of flavonoids and chalconesrdquo Pharma-cological Research vol 57 no 4 pp 259ndash265 2008
[10] F Dal Piaz A Braca M A Belisario and N De TommasildquoThioredoxin systemmodulation by plant and fungal secondarymetabolitesrdquo Current Medicinal Chemistry vol 17 no 5 pp479ndash494 2010
[11] N Yadav S K Dixit A Bhattacharya et al ldquoAntimalarial activ-ity of newly synthesized chalcone derivatives in vitrordquo Chem-ical Biology and Drug Design vol 80 no 2 pp 340ndash347 2012
[12] F V DeFeudis ldquoNovel therapy for Alzheimerrsquos diseaserdquo Neuro-biology of Aging vol 10 no 5 pp 467ndash468 1989
[13] Y Nomura and Y Kitamura ldquoInducible nitric oxide synthasein glial cellsrdquo Neuroscience Research vol 18 no 2 pp 103ndash1071993
[14] P J Kingham M L Cuzner and J M Pocock ldquoApoptotic path-ways mobilized in microglia and neurones as a consequenceof chromogranin A-induced microglial activationrdquo Journal ofNeurochemistry vol 73 no 2 pp 538ndash547 1999
[15] S Singh S Swarnkar P Goswami and C Nath ldquoAstrocytes andmicroglia responses to neuropathological conditionsrdquo Inter-national Journal of Neuroscience vol 121 no 11 pp 589ndash5972011
[16] Y Y Deng J Lu E-A Ling and C Kaur ldquoRole of microglia inthe process of inflammation in the hypoxic developing brainrdquoFrontiers in Bioscience (Scholar Edition) vol 3 no 3 pp 884ndash900 2011
[17] M Dumont and M F Beal ldquoNeuroprotective strategies involv-ing ROS in Alzheimer diseaserdquo Free Radical Biology andMedicine vol 51 no 5 pp 1014ndash1026 2011
[18] AD Kraft andG JeanHarry ldquoFeatures ofmicroglia and neuro-inflammation relevant to environmental exposure and neuro-toxicityrdquo International Journal of Environmental Research andPublic Health vol 8 no 7 pp 2980ndash3018 2011
[19] S Cho S Kim Z Jin et al ldquoIsoliquiritigenin a chalcone com-pound is a positive allosteric modulator of GABA A recep-tors and shows hypnotic effectsrdquo Biochemical and BiophysicalResearch Communications vol 413 no 4 pp 637ndash642 2011
10 Oxidative Medicine and Cellular Longevity
[20] A Elmann A Telerman H Erlank et al ldquoProtective and anti-oxidant effects of a chalconoid from Pulicaria incisa on brainastrocytesrdquoOxidativeMedicine andCellular Longevity vol 2013Article ID 694398 10 pages 2013
[21] M Rossi F Caruso E J Crespi et al ldquoProbing antioxidantactivity of 21015840-hydroxychalcones crystal and molecular struc-tures in vitro antiproliferative studies and in vivo effects onglucose regulationrdquo Biochimie vol 95 no 10 pp 1954ndash19632013
[22] E LMead AMosley S Eaton L Dobson S J Heales and JMPocock ldquoMicroglial neurotransmitter receptors trigger super-oxide production in microglia consequences for microglial-neuronal interactionsrdquo Journal of Neurochemistry vol 121 no2 pp 287ndash301 2012
[23] C M Davenport I G Sevastou C Hooper and J M PocockldquoInhibiting p53 pathways in microglia attenuates microglial-evoked neurotoxicity following exposure to Alzheimer pep-tidesrdquo Journal of Neurochemistry vol 112 no 2 pp 552ndash5632010
[24] N C Garbett N B Hammond and D E Graves ldquoInfluence ofthe amino substituents in the interaction of ethidium bromidewith DNArdquo Biophysical Journal vol 87 no 6 pp 3974ndash39812004
[25] A Karlsson J B Nixon and L C McPhail ldquoPhorbol myristateacetate induces neutrophilNADPH-oxidase activity by two sep-arate signal transduction pathways dependent or independentof phosphatidylinositol 3-kinaserdquo Journal of Leukocyte Biologyvol 67 no 3 pp 396ndash404 2000
[26] F Herencia M L Ferrandiz A Ubeda et al ldquo4-Dimethyl-amino-3101584041015840-dimethoxychalcone down-regulates iNOS expres-sion and exerts anti-inflammatory effectsrdquo Free Radical Biologyamp Medicine vol 30 no 1 pp 43ndash50 2001
[27] N Mateeva M Gangapuram E Mazzio S Eyunni K FSoliman and K K Redda ldquoBiological evaluation of syntheticchalcone and flavone derivatives as anti-inflammatory agentsrdquoMedicinal Chemistry Research vol 24 no 4 pp 1672ndash16802015
[28] S C Morgan D L Taylor and J M Pocock ldquoMicroglia releaseactivators of neuronal proliferation mediated by activationof mitogen-activated protein kinase phosphatidylinositol-3-kinaseAkt and delta-Notch signalling cascadesrdquo Journal ofNeurochemistry vol 90 no 1 pp 89ndash101 2004
[29] J Jebelli T Piers and J Pocock ldquoSelective depletion of micro-glia from cerebellar granule cell cultures using L-leucinemethylesterrdquoThe Journal of Visualized Experiments no 101 Article IDe52983 2015
[30] R Von Bernhardi J E Tichauer and J Eugenın ldquoAging-dependent changes of microglial cells and their relevance forneurodegenerative disordersrdquo Journal of Neurochemistry vol112 no 5 pp 1099ndash1114 2010
[31] C Schwab and P L McGeer ldquoInflammatory aspects ofAlzheimer disease and other neurodegenerative disordersrdquoJournal of Alzheimerrsquos Disease vol 13 no 4 pp 359ndash369 2008
[32] A Henn S Lund M Hedtjarn A Schrattenholz P Porzgenand M Leist ldquoThe suitability of BV2 cells as alternative modelsystem for primary microglia cultures or for animal experi-ments examining brain inflammationrdquo Altex vol 26 no 2 pp83ndash94 2009
[33] S Bano K Javed S Ahmad et al ldquoSynthesis of some novel chal-cones flavanones and flavones and evaluation of their anti-inflammatory activityrdquo European Journal of Medicinal Chem-istry vol 65 pp 51ndash59 2013
[34] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[35] S L Deshmane S Kremlev S Amini and B E Sawaya ldquoMono-cyte chemoattractant protein-1 (MCP-1) an overviewrdquo Journalof Interferon and Cytokine Research vol 29 no 6 pp 313ndash3262009
[36] J Rojas J N Domınguez J E Charris G Lobo M Paya andM L Ferrandiz ldquoSynthesis and inhibitory activity of dimethyl-amino-chalcone derivatives on the induction of nitric oxidesynthaserdquo European Journal of Medicinal Chemistry vol 37 no8 pp 699ndash705 2002
[37] H Hara Y Nakamura M Ninomiya et al ldquoInhibitory effectsof chalcone glycosides isolated from Brassica rapa L lsquohidabenirsquoand their synthetic derivatives on LPS-induced NO productionin microgliardquo Bioorganic amp Medicinal Chemistry vol 19 no 18pp 5559ndash5568 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
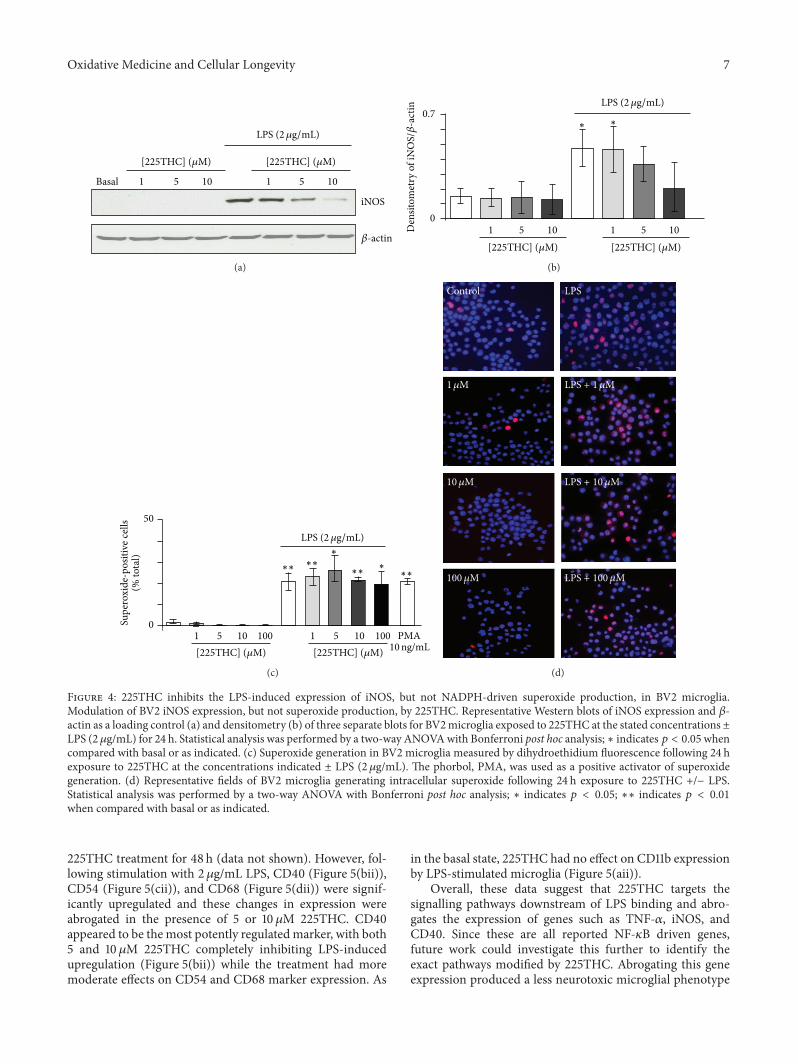
Oxidative Medicine and Cellular Longevity 7
iNOS
Basal 1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M)[225THC] (120583M)
120573-actin
(a)
0
07
1 5 10 1 5 10
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
Den
sitom
etry
of i
NO
S120573
-act
in
lowast lowast
(b)
Supe
roxi
de-p
ositi
ve ce
lls(
tota
l)
50
01 5 10 100 1 5 10 100
LPS (2 120583gmL)
[225THC] (120583M) [225THC] (120583M)
lowastlowast lowastlowastlowastlowast lowastlowast
PMA10ngmL
lowast
lowast
(c)
1120583M LPS + 1120583M
10120583M
100120583M
LPS + 10120583M
LPS + 100120583M
Control LPS
(d)
Figure 4 225THC inhibits the LPS-induced expression of iNOS but not NADPH-driven superoxide production in BV2 microgliaModulation of BV2 iNOS expression but not superoxide production by 225THC Representative Western blots of iNOS expression and 120573-actin as a loading control (a) and densitometry (b) of three separate blots for BV2microglia exposed to 225THC at the stated concentrations plusmnLPS (2120583gmL) for 24 h Statistical analysis was performed by a two-way ANOVAwith Bonferroni post hoc analysis lowast indicates 119901 lt 005whencompared with basal or as indicated (c) Superoxide generation in BV2 microglia measured by dihydroethidium fluorescence following 24 hexposure to 225THC at the concentrations indicated plusmn LPS (2120583gmL) The phorbol PMA was used as a positive activator of superoxidegeneration (d) Representative fields of BV2 microglia generating intracellular superoxide following 24 h exposure to 225THC +minus LPSStatistical analysis was performed by a two-way ANOVA with Bonferroni post hoc analysis lowast indicates 119901 lt 005 lowastlowast indicates 119901 lt 001when compared with basal or as indicated
225THC treatment for 48 h (data not shown) However fol-lowing stimulation with 2 120583gmL LPS CD40 (Figure 5(bii))CD54 (Figure 5(cii)) and CD68 (Figure 5(dii)) were signif-icantly upregulated and these changes in expression wereabrogated in the presence of 5 or 10 120583M 225THC CD40appeared to be the most potently regulatedmarker with both5 and 10 120583M 225THC completely inhibiting LPS-inducedupregulation (Figure 5(bii)) while the treatment had moremoderate effects on CD54 and CD68 marker expression As
in the basal state 225THC had no effect on CD11b expressionby LPS-stimulated microglia (Figure 5(aii))
Overall these data suggest that 225THC targets thesignalling pathways downstream of LPS binding and abro-gates the expression of genes such as TNF-120572 iNOS andCD40 Since these are all reported NF-120581B driven genesfuture work could investigate this further to identify theexact pathways modified by 225THC Abrogating this geneexpression produced a less neurotoxic microglial phenotype
8 Oxidative Medicine and Cellular Longevity
CD11b
(ai) (aii)
[225THC] (120583M)
FL1-H
CD11b
Basal LPS 1 5 10
111009080706050403020100
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(a)
CD40
(bi) (bii)
[225THC] (120583M)
FL1-H
CD40
Basal LPS 1 5 10
125
100
075
050
025
000
175
150
lowastlowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(b)
(ci) (cii)
[225THC] (120583M)
FL1-H
CD54 (ICAM-1)CD54 (ICAM-1)
Basal LPS 1 5 10
400
300
200
100
000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(c)
CD68
(di) (dii)
[225THC] (120583M)FL1-H
CD68
Basal LPS 1 5 10
160140120100080060040020000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(d)
Figure 5 225THC treatmentmodulates inflammatory cell surfacemarker expression in LPS-treated BV2microglia Flow cytometric analysisof 225THC-treated BV2 microglia following LPS stimulation (a) Representative histogram plots of BV2 cell surface CD11b CD40 CD54and CD68 expression following 2 120583gmL LPS-treatment for 48 hours (unfilled histograms) versus isotype control (black-filled histograms)and in the presence of 10 120583M 225THC (grey-filled histograms) (b) Summary data showing fold changes in CD11b CD40 CD54 or CD68expression by mean fluorescence intensity (MFI) from 4 independent experiments Data are mean values plusmn SEM where lowast indicates 119901 lt 005by Studentrsquos 119905-test
Oxidative Medicine and Cellular Longevity 9
which is likely to be beneficial for a number of neurodegen-erative diseases in which aberrant microglial inflammatoryreactivity is implicated Future work could pinpoint furtherthe microglial phenotype favoured by 225THC exposure andexplore the genotype of the microglia
Abbreviations
ANOVA Analysis of varianceDPPH 22-Diphenyl-1-picrylhydrazyldHEth DihydroethidiumCGCs Cerebellar granule neuronsDIV Days in vitroDAPI 410158406-Diamidino-2-phenylindoleDMSO DimethylsulphoxideECL ElectrochemiluminescenceELISA Enzyme-linked immunosorbent assayGABAA 120574-Aminobutyric acidIFN-120574 Interferon-120574IL-1120573 Interleukin-1120573IL-6 Interleukin-6IL-10 Interleukin-10iNOS Inducible nitric oxide synthaseLPS LipopolysaccharideMCP-1 CCL2monocyte chemoattractant
protein-1MEM Modified Eaglersquos mediumMIP Macrophage inhibitory proteinMCP-1 Monocyte chemoattractant protein-1NGS Normal goat serumPBS Phosphate buffered salinePFA Paraformaldehyde2-OH-E+ 2-HydroxyethidiumRANTES CCL5regulation on activation normal T
cell expressed and secretedROS Reactive oxygen speciesTNF-120572 Tumour necrosis factor-120572
Conflict of Interests
All authors declare no conflict of interests
Acknowledgments
Manasi Jiwrajka would like to thank Dr Kathleen Susman forher guidance This research was supported by the Universityof Queensland Advantage Grant Howard Hughes MedicalFoundation Grant 52006322 to Vassar College the ShermanFairchild Foundation and Vassar College and by generalfunds to the Pocock lab
References
[1] S Ducki R Forrest J A Hadfield et al ldquoPotent antimitotic andcell growth inhibitory properties of substituted chalconesrdquo Bio-organic and Medicinal Chemistry Letters vol 8 no 9 pp 1051ndash1056 1998
[2] M L F Ferreyra S P Rius and P Casati ldquoFlavonoids biosyn-thesis biological functions and biotechnological applicationsrdquoFrontiers in Plant Science vol 3 article 222 15 pages 2012
[3] G E-D A A Abuo-Rahma M Abdel-Aziz M A E Mouradand H H Farag ldquoSynthesis anti-inflammatory activity andulcerogenic liability of novel nitric oxide donatingchalconehybridsrdquo Bioorganic and Medicinal Chemistry vol 20 no 1 pp195ndash206 2012
[4] N K Sahu S S Balbhadra J Choudhary and D V KohlildquoExploring pharmacological significance of chalcone scaffoldrdquoCurrent Medicinal Chemistry vol 19 no 2 pp 209ndash225 2012
[5] H Wei X Zhang G Wu et al ldquoChalcone derivatives from thefern Cyclosorus parasiticus and their anti-proliferative activityrdquoFood and Chemical Toxicology vol 60 pp 147ndash152 2013
[6] C W Mai M Yaeghoobi N Abd-Rahman Y B Kang andM R Pichika ldquoChalcones with electron-withdrawing and ele-ctron-donating substituents anticancer activity against TRAILresistant cancer cells structure-activity relationship analysisand regulation of apoptotic proteinsrdquo European Journal ofMedicinal Chemistry vol 77 pp 378ndash387 2014
[7] Z Nowakowska ldquoA review of anti-infective and anti-inflam-matory chalconesrdquo European Journal of Medicinal Chemistryvol 42 no 2 pp 125ndash137 2007
[8] Y-M Lin Y Zhou M T Flavin L-M Zhou W Nie and F-CChen ldquoChalcones and flavonoids as anti-tuberculosis agentsrdquoBioorganic and Medicinal Chemistry vol 10 no 8 pp 2795ndash2802 2002
[9] JMojzis L Varinska GMojzisova I Kostova and LMirossayldquoAnti-angiogenic effects of flavonoids and chalconesrdquo Pharma-cological Research vol 57 no 4 pp 259ndash265 2008
[10] F Dal Piaz A Braca M A Belisario and N De TommasildquoThioredoxin systemmodulation by plant and fungal secondarymetabolitesrdquo Current Medicinal Chemistry vol 17 no 5 pp479ndash494 2010
[11] N Yadav S K Dixit A Bhattacharya et al ldquoAntimalarial activ-ity of newly synthesized chalcone derivatives in vitrordquo Chem-ical Biology and Drug Design vol 80 no 2 pp 340ndash347 2012
[12] F V DeFeudis ldquoNovel therapy for Alzheimerrsquos diseaserdquo Neuro-biology of Aging vol 10 no 5 pp 467ndash468 1989
[13] Y Nomura and Y Kitamura ldquoInducible nitric oxide synthasein glial cellsrdquo Neuroscience Research vol 18 no 2 pp 103ndash1071993
[14] P J Kingham M L Cuzner and J M Pocock ldquoApoptotic path-ways mobilized in microglia and neurones as a consequenceof chromogranin A-induced microglial activationrdquo Journal ofNeurochemistry vol 73 no 2 pp 538ndash547 1999
[15] S Singh S Swarnkar P Goswami and C Nath ldquoAstrocytes andmicroglia responses to neuropathological conditionsrdquo Inter-national Journal of Neuroscience vol 121 no 11 pp 589ndash5972011
[16] Y Y Deng J Lu E-A Ling and C Kaur ldquoRole of microglia inthe process of inflammation in the hypoxic developing brainrdquoFrontiers in Bioscience (Scholar Edition) vol 3 no 3 pp 884ndash900 2011
[17] M Dumont and M F Beal ldquoNeuroprotective strategies involv-ing ROS in Alzheimer diseaserdquo Free Radical Biology andMedicine vol 51 no 5 pp 1014ndash1026 2011
[18] AD Kraft andG JeanHarry ldquoFeatures ofmicroglia and neuro-inflammation relevant to environmental exposure and neuro-toxicityrdquo International Journal of Environmental Research andPublic Health vol 8 no 7 pp 2980ndash3018 2011
[19] S Cho S Kim Z Jin et al ldquoIsoliquiritigenin a chalcone com-pound is a positive allosteric modulator of GABA A recep-tors and shows hypnotic effectsrdquo Biochemical and BiophysicalResearch Communications vol 413 no 4 pp 637ndash642 2011
10 Oxidative Medicine and Cellular Longevity
[20] A Elmann A Telerman H Erlank et al ldquoProtective and anti-oxidant effects of a chalconoid from Pulicaria incisa on brainastrocytesrdquoOxidativeMedicine andCellular Longevity vol 2013Article ID 694398 10 pages 2013
[21] M Rossi F Caruso E J Crespi et al ldquoProbing antioxidantactivity of 21015840-hydroxychalcones crystal and molecular struc-tures in vitro antiproliferative studies and in vivo effects onglucose regulationrdquo Biochimie vol 95 no 10 pp 1954ndash19632013
[22] E LMead AMosley S Eaton L Dobson S J Heales and JMPocock ldquoMicroglial neurotransmitter receptors trigger super-oxide production in microglia consequences for microglial-neuronal interactionsrdquo Journal of Neurochemistry vol 121 no2 pp 287ndash301 2012
[23] C M Davenport I G Sevastou C Hooper and J M PocockldquoInhibiting p53 pathways in microglia attenuates microglial-evoked neurotoxicity following exposure to Alzheimer pep-tidesrdquo Journal of Neurochemistry vol 112 no 2 pp 552ndash5632010
[24] N C Garbett N B Hammond and D E Graves ldquoInfluence ofthe amino substituents in the interaction of ethidium bromidewith DNArdquo Biophysical Journal vol 87 no 6 pp 3974ndash39812004
[25] A Karlsson J B Nixon and L C McPhail ldquoPhorbol myristateacetate induces neutrophilNADPH-oxidase activity by two sep-arate signal transduction pathways dependent or independentof phosphatidylinositol 3-kinaserdquo Journal of Leukocyte Biologyvol 67 no 3 pp 396ndash404 2000
[26] F Herencia M L Ferrandiz A Ubeda et al ldquo4-Dimethyl-amino-3101584041015840-dimethoxychalcone down-regulates iNOS expres-sion and exerts anti-inflammatory effectsrdquo Free Radical Biologyamp Medicine vol 30 no 1 pp 43ndash50 2001
[27] N Mateeva M Gangapuram E Mazzio S Eyunni K FSoliman and K K Redda ldquoBiological evaluation of syntheticchalcone and flavone derivatives as anti-inflammatory agentsrdquoMedicinal Chemistry Research vol 24 no 4 pp 1672ndash16802015
[28] S C Morgan D L Taylor and J M Pocock ldquoMicroglia releaseactivators of neuronal proliferation mediated by activationof mitogen-activated protein kinase phosphatidylinositol-3-kinaseAkt and delta-Notch signalling cascadesrdquo Journal ofNeurochemistry vol 90 no 1 pp 89ndash101 2004
[29] J Jebelli T Piers and J Pocock ldquoSelective depletion of micro-glia from cerebellar granule cell cultures using L-leucinemethylesterrdquoThe Journal of Visualized Experiments no 101 Article IDe52983 2015
[30] R Von Bernhardi J E Tichauer and J Eugenın ldquoAging-dependent changes of microglial cells and their relevance forneurodegenerative disordersrdquo Journal of Neurochemistry vol112 no 5 pp 1099ndash1114 2010
[31] C Schwab and P L McGeer ldquoInflammatory aspects ofAlzheimer disease and other neurodegenerative disordersrdquoJournal of Alzheimerrsquos Disease vol 13 no 4 pp 359ndash369 2008
[32] A Henn S Lund M Hedtjarn A Schrattenholz P Porzgenand M Leist ldquoThe suitability of BV2 cells as alternative modelsystem for primary microglia cultures or for animal experi-ments examining brain inflammationrdquo Altex vol 26 no 2 pp83ndash94 2009
[33] S Bano K Javed S Ahmad et al ldquoSynthesis of some novel chal-cones flavanones and flavones and evaluation of their anti-inflammatory activityrdquo European Journal of Medicinal Chem-istry vol 65 pp 51ndash59 2013
[34] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[35] S L Deshmane S Kremlev S Amini and B E Sawaya ldquoMono-cyte chemoattractant protein-1 (MCP-1) an overviewrdquo Journalof Interferon and Cytokine Research vol 29 no 6 pp 313ndash3262009
[36] J Rojas J N Domınguez J E Charris G Lobo M Paya andM L Ferrandiz ldquoSynthesis and inhibitory activity of dimethyl-amino-chalcone derivatives on the induction of nitric oxidesynthaserdquo European Journal of Medicinal Chemistry vol 37 no8 pp 699ndash705 2002
[37] H Hara Y Nakamura M Ninomiya et al ldquoInhibitory effectsof chalcone glycosides isolated from Brassica rapa L lsquohidabenirsquoand their synthetic derivatives on LPS-induced NO productionin microgliardquo Bioorganic amp Medicinal Chemistry vol 19 no 18pp 5559ndash5568 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
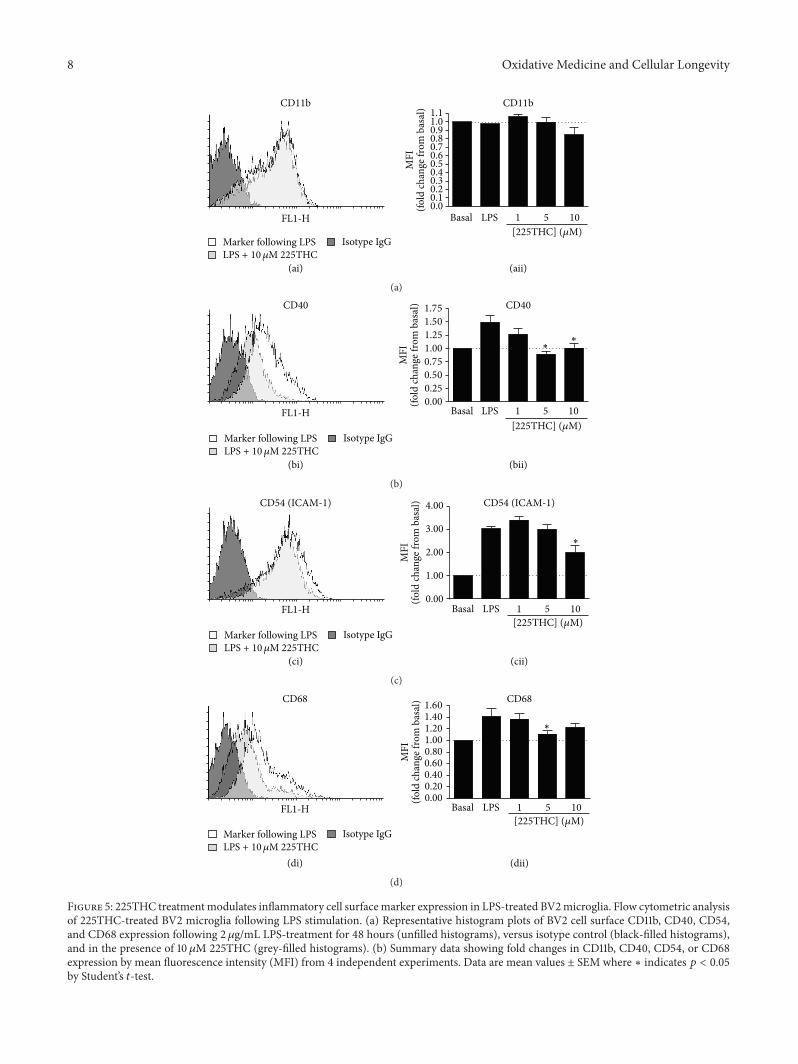
8 Oxidative Medicine and Cellular Longevity
CD11b
(ai) (aii)
[225THC] (120583M)
FL1-H
CD11b
Basal LPS 1 5 10
111009080706050403020100
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(a)
CD40
(bi) (bii)
[225THC] (120583M)
FL1-H
CD40
Basal LPS 1 5 10
125
100
075
050
025
000
175
150
lowastlowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(b)
(ci) (cii)
[225THC] (120583M)
FL1-H
CD54 (ICAM-1)CD54 (ICAM-1)
Basal LPS 1 5 10
400
300
200
100
000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(c)
CD68
(di) (dii)
[225THC] (120583M)FL1-H
CD68
Basal LPS 1 5 10
160140120100080060040020000
lowast
Isotype IgGMarker following LPS LPS + 10120583M 225THC
MFI
(fold
chan
ge fr
om b
asal
)
(d)
Figure 5 225THC treatmentmodulates inflammatory cell surfacemarker expression in LPS-treated BV2microglia Flow cytometric analysisof 225THC-treated BV2 microglia following LPS stimulation (a) Representative histogram plots of BV2 cell surface CD11b CD40 CD54and CD68 expression following 2 120583gmL LPS-treatment for 48 hours (unfilled histograms) versus isotype control (black-filled histograms)and in the presence of 10 120583M 225THC (grey-filled histograms) (b) Summary data showing fold changes in CD11b CD40 CD54 or CD68expression by mean fluorescence intensity (MFI) from 4 independent experiments Data are mean values plusmn SEM where lowast indicates 119901 lt 005by Studentrsquos 119905-test
Oxidative Medicine and Cellular Longevity 9
which is likely to be beneficial for a number of neurodegen-erative diseases in which aberrant microglial inflammatoryreactivity is implicated Future work could pinpoint furtherthe microglial phenotype favoured by 225THC exposure andexplore the genotype of the microglia
Abbreviations
ANOVA Analysis of varianceDPPH 22-Diphenyl-1-picrylhydrazyldHEth DihydroethidiumCGCs Cerebellar granule neuronsDIV Days in vitroDAPI 410158406-Diamidino-2-phenylindoleDMSO DimethylsulphoxideECL ElectrochemiluminescenceELISA Enzyme-linked immunosorbent assayGABAA 120574-Aminobutyric acidIFN-120574 Interferon-120574IL-1120573 Interleukin-1120573IL-6 Interleukin-6IL-10 Interleukin-10iNOS Inducible nitric oxide synthaseLPS LipopolysaccharideMCP-1 CCL2monocyte chemoattractant
protein-1MEM Modified Eaglersquos mediumMIP Macrophage inhibitory proteinMCP-1 Monocyte chemoattractant protein-1NGS Normal goat serumPBS Phosphate buffered salinePFA Paraformaldehyde2-OH-E+ 2-HydroxyethidiumRANTES CCL5regulation on activation normal T
cell expressed and secretedROS Reactive oxygen speciesTNF-120572 Tumour necrosis factor-120572
Conflict of Interests
All authors declare no conflict of interests
Acknowledgments
Manasi Jiwrajka would like to thank Dr Kathleen Susman forher guidance This research was supported by the Universityof Queensland Advantage Grant Howard Hughes MedicalFoundation Grant 52006322 to Vassar College the ShermanFairchild Foundation and Vassar College and by generalfunds to the Pocock lab
References
[1] S Ducki R Forrest J A Hadfield et al ldquoPotent antimitotic andcell growth inhibitory properties of substituted chalconesrdquo Bio-organic and Medicinal Chemistry Letters vol 8 no 9 pp 1051ndash1056 1998
[2] M L F Ferreyra S P Rius and P Casati ldquoFlavonoids biosyn-thesis biological functions and biotechnological applicationsrdquoFrontiers in Plant Science vol 3 article 222 15 pages 2012
[3] G E-D A A Abuo-Rahma M Abdel-Aziz M A E Mouradand H H Farag ldquoSynthesis anti-inflammatory activity andulcerogenic liability of novel nitric oxide donatingchalconehybridsrdquo Bioorganic and Medicinal Chemistry vol 20 no 1 pp195ndash206 2012
[4] N K Sahu S S Balbhadra J Choudhary and D V KohlildquoExploring pharmacological significance of chalcone scaffoldrdquoCurrent Medicinal Chemistry vol 19 no 2 pp 209ndash225 2012
[5] H Wei X Zhang G Wu et al ldquoChalcone derivatives from thefern Cyclosorus parasiticus and their anti-proliferative activityrdquoFood and Chemical Toxicology vol 60 pp 147ndash152 2013
[6] C W Mai M Yaeghoobi N Abd-Rahman Y B Kang andM R Pichika ldquoChalcones with electron-withdrawing and ele-ctron-donating substituents anticancer activity against TRAILresistant cancer cells structure-activity relationship analysisand regulation of apoptotic proteinsrdquo European Journal ofMedicinal Chemistry vol 77 pp 378ndash387 2014
[7] Z Nowakowska ldquoA review of anti-infective and anti-inflam-matory chalconesrdquo European Journal of Medicinal Chemistryvol 42 no 2 pp 125ndash137 2007
[8] Y-M Lin Y Zhou M T Flavin L-M Zhou W Nie and F-CChen ldquoChalcones and flavonoids as anti-tuberculosis agentsrdquoBioorganic and Medicinal Chemistry vol 10 no 8 pp 2795ndash2802 2002
[9] JMojzis L Varinska GMojzisova I Kostova and LMirossayldquoAnti-angiogenic effects of flavonoids and chalconesrdquo Pharma-cological Research vol 57 no 4 pp 259ndash265 2008
[10] F Dal Piaz A Braca M A Belisario and N De TommasildquoThioredoxin systemmodulation by plant and fungal secondarymetabolitesrdquo Current Medicinal Chemistry vol 17 no 5 pp479ndash494 2010
[11] N Yadav S K Dixit A Bhattacharya et al ldquoAntimalarial activ-ity of newly synthesized chalcone derivatives in vitrordquo Chem-ical Biology and Drug Design vol 80 no 2 pp 340ndash347 2012
[12] F V DeFeudis ldquoNovel therapy for Alzheimerrsquos diseaserdquo Neuro-biology of Aging vol 10 no 5 pp 467ndash468 1989
[13] Y Nomura and Y Kitamura ldquoInducible nitric oxide synthasein glial cellsrdquo Neuroscience Research vol 18 no 2 pp 103ndash1071993
[14] P J Kingham M L Cuzner and J M Pocock ldquoApoptotic path-ways mobilized in microglia and neurones as a consequenceof chromogranin A-induced microglial activationrdquo Journal ofNeurochemistry vol 73 no 2 pp 538ndash547 1999
[15] S Singh S Swarnkar P Goswami and C Nath ldquoAstrocytes andmicroglia responses to neuropathological conditionsrdquo Inter-national Journal of Neuroscience vol 121 no 11 pp 589ndash5972011
[16] Y Y Deng J Lu E-A Ling and C Kaur ldquoRole of microglia inthe process of inflammation in the hypoxic developing brainrdquoFrontiers in Bioscience (Scholar Edition) vol 3 no 3 pp 884ndash900 2011
[17] M Dumont and M F Beal ldquoNeuroprotective strategies involv-ing ROS in Alzheimer diseaserdquo Free Radical Biology andMedicine vol 51 no 5 pp 1014ndash1026 2011
[18] AD Kraft andG JeanHarry ldquoFeatures ofmicroglia and neuro-inflammation relevant to environmental exposure and neuro-toxicityrdquo International Journal of Environmental Research andPublic Health vol 8 no 7 pp 2980ndash3018 2011
[19] S Cho S Kim Z Jin et al ldquoIsoliquiritigenin a chalcone com-pound is a positive allosteric modulator of GABA A recep-tors and shows hypnotic effectsrdquo Biochemical and BiophysicalResearch Communications vol 413 no 4 pp 637ndash642 2011
10 Oxidative Medicine and Cellular Longevity
[20] A Elmann A Telerman H Erlank et al ldquoProtective and anti-oxidant effects of a chalconoid from Pulicaria incisa on brainastrocytesrdquoOxidativeMedicine andCellular Longevity vol 2013Article ID 694398 10 pages 2013
[21] M Rossi F Caruso E J Crespi et al ldquoProbing antioxidantactivity of 21015840-hydroxychalcones crystal and molecular struc-tures in vitro antiproliferative studies and in vivo effects onglucose regulationrdquo Biochimie vol 95 no 10 pp 1954ndash19632013
[22] E LMead AMosley S Eaton L Dobson S J Heales and JMPocock ldquoMicroglial neurotransmitter receptors trigger super-oxide production in microglia consequences for microglial-neuronal interactionsrdquo Journal of Neurochemistry vol 121 no2 pp 287ndash301 2012
[23] C M Davenport I G Sevastou C Hooper and J M PocockldquoInhibiting p53 pathways in microglia attenuates microglial-evoked neurotoxicity following exposure to Alzheimer pep-tidesrdquo Journal of Neurochemistry vol 112 no 2 pp 552ndash5632010
[24] N C Garbett N B Hammond and D E Graves ldquoInfluence ofthe amino substituents in the interaction of ethidium bromidewith DNArdquo Biophysical Journal vol 87 no 6 pp 3974ndash39812004
[25] A Karlsson J B Nixon and L C McPhail ldquoPhorbol myristateacetate induces neutrophilNADPH-oxidase activity by two sep-arate signal transduction pathways dependent or independentof phosphatidylinositol 3-kinaserdquo Journal of Leukocyte Biologyvol 67 no 3 pp 396ndash404 2000
[26] F Herencia M L Ferrandiz A Ubeda et al ldquo4-Dimethyl-amino-3101584041015840-dimethoxychalcone down-regulates iNOS expres-sion and exerts anti-inflammatory effectsrdquo Free Radical Biologyamp Medicine vol 30 no 1 pp 43ndash50 2001
[27] N Mateeva M Gangapuram E Mazzio S Eyunni K FSoliman and K K Redda ldquoBiological evaluation of syntheticchalcone and flavone derivatives as anti-inflammatory agentsrdquoMedicinal Chemistry Research vol 24 no 4 pp 1672ndash16802015
[28] S C Morgan D L Taylor and J M Pocock ldquoMicroglia releaseactivators of neuronal proliferation mediated by activationof mitogen-activated protein kinase phosphatidylinositol-3-kinaseAkt and delta-Notch signalling cascadesrdquo Journal ofNeurochemistry vol 90 no 1 pp 89ndash101 2004
[29] J Jebelli T Piers and J Pocock ldquoSelective depletion of micro-glia from cerebellar granule cell cultures using L-leucinemethylesterrdquoThe Journal of Visualized Experiments no 101 Article IDe52983 2015
[30] R Von Bernhardi J E Tichauer and J Eugenın ldquoAging-dependent changes of microglial cells and their relevance forneurodegenerative disordersrdquo Journal of Neurochemistry vol112 no 5 pp 1099ndash1114 2010
[31] C Schwab and P L McGeer ldquoInflammatory aspects ofAlzheimer disease and other neurodegenerative disordersrdquoJournal of Alzheimerrsquos Disease vol 13 no 4 pp 359ndash369 2008
[32] A Henn S Lund M Hedtjarn A Schrattenholz P Porzgenand M Leist ldquoThe suitability of BV2 cells as alternative modelsystem for primary microglia cultures or for animal experi-ments examining brain inflammationrdquo Altex vol 26 no 2 pp83ndash94 2009
[33] S Bano K Javed S Ahmad et al ldquoSynthesis of some novel chal-cones flavanones and flavones and evaluation of their anti-inflammatory activityrdquo European Journal of Medicinal Chem-istry vol 65 pp 51ndash59 2013
[34] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[35] S L Deshmane S Kremlev S Amini and B E Sawaya ldquoMono-cyte chemoattractant protein-1 (MCP-1) an overviewrdquo Journalof Interferon and Cytokine Research vol 29 no 6 pp 313ndash3262009
[36] J Rojas J N Domınguez J E Charris G Lobo M Paya andM L Ferrandiz ldquoSynthesis and inhibitory activity of dimethyl-amino-chalcone derivatives on the induction of nitric oxidesynthaserdquo European Journal of Medicinal Chemistry vol 37 no8 pp 699ndash705 2002
[37] H Hara Y Nakamura M Ninomiya et al ldquoInhibitory effectsof chalcone glycosides isolated from Brassica rapa L lsquohidabenirsquoand their synthetic derivatives on LPS-induced NO productionin microgliardquo Bioorganic amp Medicinal Chemistry vol 19 no 18pp 5559ndash5568 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 9
which is likely to be beneficial for a number of neurodegen-erative diseases in which aberrant microglial inflammatoryreactivity is implicated Future work could pinpoint furtherthe microglial phenotype favoured by 225THC exposure andexplore the genotype of the microglia
Abbreviations
ANOVA Analysis of varianceDPPH 22-Diphenyl-1-picrylhydrazyldHEth DihydroethidiumCGCs Cerebellar granule neuronsDIV Days in vitroDAPI 410158406-Diamidino-2-phenylindoleDMSO DimethylsulphoxideECL ElectrochemiluminescenceELISA Enzyme-linked immunosorbent assayGABAA 120574-Aminobutyric acidIFN-120574 Interferon-120574IL-1120573 Interleukin-1120573IL-6 Interleukin-6IL-10 Interleukin-10iNOS Inducible nitric oxide synthaseLPS LipopolysaccharideMCP-1 CCL2monocyte chemoattractant
protein-1MEM Modified Eaglersquos mediumMIP Macrophage inhibitory proteinMCP-1 Monocyte chemoattractant protein-1NGS Normal goat serumPBS Phosphate buffered salinePFA Paraformaldehyde2-OH-E+ 2-HydroxyethidiumRANTES CCL5regulation on activation normal T
cell expressed and secretedROS Reactive oxygen speciesTNF-120572 Tumour necrosis factor-120572
Conflict of Interests
All authors declare no conflict of interests
Acknowledgments
Manasi Jiwrajka would like to thank Dr Kathleen Susman forher guidance This research was supported by the Universityof Queensland Advantage Grant Howard Hughes MedicalFoundation Grant 52006322 to Vassar College the ShermanFairchild Foundation and Vassar College and by generalfunds to the Pocock lab
References
[1] S Ducki R Forrest J A Hadfield et al ldquoPotent antimitotic andcell growth inhibitory properties of substituted chalconesrdquo Bio-organic and Medicinal Chemistry Letters vol 8 no 9 pp 1051ndash1056 1998
[2] M L F Ferreyra S P Rius and P Casati ldquoFlavonoids biosyn-thesis biological functions and biotechnological applicationsrdquoFrontiers in Plant Science vol 3 article 222 15 pages 2012
[3] G E-D A A Abuo-Rahma M Abdel-Aziz M A E Mouradand H H Farag ldquoSynthesis anti-inflammatory activity andulcerogenic liability of novel nitric oxide donatingchalconehybridsrdquo Bioorganic and Medicinal Chemistry vol 20 no 1 pp195ndash206 2012
[4] N K Sahu S S Balbhadra J Choudhary and D V KohlildquoExploring pharmacological significance of chalcone scaffoldrdquoCurrent Medicinal Chemistry vol 19 no 2 pp 209ndash225 2012
[5] H Wei X Zhang G Wu et al ldquoChalcone derivatives from thefern Cyclosorus parasiticus and their anti-proliferative activityrdquoFood and Chemical Toxicology vol 60 pp 147ndash152 2013
[6] C W Mai M Yaeghoobi N Abd-Rahman Y B Kang andM R Pichika ldquoChalcones with electron-withdrawing and ele-ctron-donating substituents anticancer activity against TRAILresistant cancer cells structure-activity relationship analysisand regulation of apoptotic proteinsrdquo European Journal ofMedicinal Chemistry vol 77 pp 378ndash387 2014
[7] Z Nowakowska ldquoA review of anti-infective and anti-inflam-matory chalconesrdquo European Journal of Medicinal Chemistryvol 42 no 2 pp 125ndash137 2007
[8] Y-M Lin Y Zhou M T Flavin L-M Zhou W Nie and F-CChen ldquoChalcones and flavonoids as anti-tuberculosis agentsrdquoBioorganic and Medicinal Chemistry vol 10 no 8 pp 2795ndash2802 2002
[9] JMojzis L Varinska GMojzisova I Kostova and LMirossayldquoAnti-angiogenic effects of flavonoids and chalconesrdquo Pharma-cological Research vol 57 no 4 pp 259ndash265 2008
[10] F Dal Piaz A Braca M A Belisario and N De TommasildquoThioredoxin systemmodulation by plant and fungal secondarymetabolitesrdquo Current Medicinal Chemistry vol 17 no 5 pp479ndash494 2010
[11] N Yadav S K Dixit A Bhattacharya et al ldquoAntimalarial activ-ity of newly synthesized chalcone derivatives in vitrordquo Chem-ical Biology and Drug Design vol 80 no 2 pp 340ndash347 2012
[12] F V DeFeudis ldquoNovel therapy for Alzheimerrsquos diseaserdquo Neuro-biology of Aging vol 10 no 5 pp 467ndash468 1989
[13] Y Nomura and Y Kitamura ldquoInducible nitric oxide synthasein glial cellsrdquo Neuroscience Research vol 18 no 2 pp 103ndash1071993
[14] P J Kingham M L Cuzner and J M Pocock ldquoApoptotic path-ways mobilized in microglia and neurones as a consequenceof chromogranin A-induced microglial activationrdquo Journal ofNeurochemistry vol 73 no 2 pp 538ndash547 1999
[15] S Singh S Swarnkar P Goswami and C Nath ldquoAstrocytes andmicroglia responses to neuropathological conditionsrdquo Inter-national Journal of Neuroscience vol 121 no 11 pp 589ndash5972011
[16] Y Y Deng J Lu E-A Ling and C Kaur ldquoRole of microglia inthe process of inflammation in the hypoxic developing brainrdquoFrontiers in Bioscience (Scholar Edition) vol 3 no 3 pp 884ndash900 2011
[17] M Dumont and M F Beal ldquoNeuroprotective strategies involv-ing ROS in Alzheimer diseaserdquo Free Radical Biology andMedicine vol 51 no 5 pp 1014ndash1026 2011
[18] AD Kraft andG JeanHarry ldquoFeatures ofmicroglia and neuro-inflammation relevant to environmental exposure and neuro-toxicityrdquo International Journal of Environmental Research andPublic Health vol 8 no 7 pp 2980ndash3018 2011
[19] S Cho S Kim Z Jin et al ldquoIsoliquiritigenin a chalcone com-pound is a positive allosteric modulator of GABA A recep-tors and shows hypnotic effectsrdquo Biochemical and BiophysicalResearch Communications vol 413 no 4 pp 637ndash642 2011
10 Oxidative Medicine and Cellular Longevity
[20] A Elmann A Telerman H Erlank et al ldquoProtective and anti-oxidant effects of a chalconoid from Pulicaria incisa on brainastrocytesrdquoOxidativeMedicine andCellular Longevity vol 2013Article ID 694398 10 pages 2013
[21] M Rossi F Caruso E J Crespi et al ldquoProbing antioxidantactivity of 21015840-hydroxychalcones crystal and molecular struc-tures in vitro antiproliferative studies and in vivo effects onglucose regulationrdquo Biochimie vol 95 no 10 pp 1954ndash19632013
[22] E LMead AMosley S Eaton L Dobson S J Heales and JMPocock ldquoMicroglial neurotransmitter receptors trigger super-oxide production in microglia consequences for microglial-neuronal interactionsrdquo Journal of Neurochemistry vol 121 no2 pp 287ndash301 2012
[23] C M Davenport I G Sevastou C Hooper and J M PocockldquoInhibiting p53 pathways in microglia attenuates microglial-evoked neurotoxicity following exposure to Alzheimer pep-tidesrdquo Journal of Neurochemistry vol 112 no 2 pp 552ndash5632010
[24] N C Garbett N B Hammond and D E Graves ldquoInfluence ofthe amino substituents in the interaction of ethidium bromidewith DNArdquo Biophysical Journal vol 87 no 6 pp 3974ndash39812004
[25] A Karlsson J B Nixon and L C McPhail ldquoPhorbol myristateacetate induces neutrophilNADPH-oxidase activity by two sep-arate signal transduction pathways dependent or independentof phosphatidylinositol 3-kinaserdquo Journal of Leukocyte Biologyvol 67 no 3 pp 396ndash404 2000
[26] F Herencia M L Ferrandiz A Ubeda et al ldquo4-Dimethyl-amino-3101584041015840-dimethoxychalcone down-regulates iNOS expres-sion and exerts anti-inflammatory effectsrdquo Free Radical Biologyamp Medicine vol 30 no 1 pp 43ndash50 2001
[27] N Mateeva M Gangapuram E Mazzio S Eyunni K FSoliman and K K Redda ldquoBiological evaluation of syntheticchalcone and flavone derivatives as anti-inflammatory agentsrdquoMedicinal Chemistry Research vol 24 no 4 pp 1672ndash16802015
[28] S C Morgan D L Taylor and J M Pocock ldquoMicroglia releaseactivators of neuronal proliferation mediated by activationof mitogen-activated protein kinase phosphatidylinositol-3-kinaseAkt and delta-Notch signalling cascadesrdquo Journal ofNeurochemistry vol 90 no 1 pp 89ndash101 2004
[29] J Jebelli T Piers and J Pocock ldquoSelective depletion of micro-glia from cerebellar granule cell cultures using L-leucinemethylesterrdquoThe Journal of Visualized Experiments no 101 Article IDe52983 2015
[30] R Von Bernhardi J E Tichauer and J Eugenın ldquoAging-dependent changes of microglial cells and their relevance forneurodegenerative disordersrdquo Journal of Neurochemistry vol112 no 5 pp 1099ndash1114 2010
[31] C Schwab and P L McGeer ldquoInflammatory aspects ofAlzheimer disease and other neurodegenerative disordersrdquoJournal of Alzheimerrsquos Disease vol 13 no 4 pp 359ndash369 2008
[32] A Henn S Lund M Hedtjarn A Schrattenholz P Porzgenand M Leist ldquoThe suitability of BV2 cells as alternative modelsystem for primary microglia cultures or for animal experi-ments examining brain inflammationrdquo Altex vol 26 no 2 pp83ndash94 2009
[33] S Bano K Javed S Ahmad et al ldquoSynthesis of some novel chal-cones flavanones and flavones and evaluation of their anti-inflammatory activityrdquo European Journal of Medicinal Chem-istry vol 65 pp 51ndash59 2013
[34] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[35] S L Deshmane S Kremlev S Amini and B E Sawaya ldquoMono-cyte chemoattractant protein-1 (MCP-1) an overviewrdquo Journalof Interferon and Cytokine Research vol 29 no 6 pp 313ndash3262009
[36] J Rojas J N Domınguez J E Charris G Lobo M Paya andM L Ferrandiz ldquoSynthesis and inhibitory activity of dimethyl-amino-chalcone derivatives on the induction of nitric oxidesynthaserdquo European Journal of Medicinal Chemistry vol 37 no8 pp 699ndash705 2002
[37] H Hara Y Nakamura M Ninomiya et al ldquoInhibitory effectsof chalcone glycosides isolated from Brassica rapa L lsquohidabenirsquoand their synthetic derivatives on LPS-induced NO productionin microgliardquo Bioorganic amp Medicinal Chemistry vol 19 no 18pp 5559ndash5568 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Oxidative Medicine and Cellular Longevity
[20] A Elmann A Telerman H Erlank et al ldquoProtective and anti-oxidant effects of a chalconoid from Pulicaria incisa on brainastrocytesrdquoOxidativeMedicine andCellular Longevity vol 2013Article ID 694398 10 pages 2013
[21] M Rossi F Caruso E J Crespi et al ldquoProbing antioxidantactivity of 21015840-hydroxychalcones crystal and molecular struc-tures in vitro antiproliferative studies and in vivo effects onglucose regulationrdquo Biochimie vol 95 no 10 pp 1954ndash19632013
[22] E LMead AMosley S Eaton L Dobson S J Heales and JMPocock ldquoMicroglial neurotransmitter receptors trigger super-oxide production in microglia consequences for microglial-neuronal interactionsrdquo Journal of Neurochemistry vol 121 no2 pp 287ndash301 2012
[23] C M Davenport I G Sevastou C Hooper and J M PocockldquoInhibiting p53 pathways in microglia attenuates microglial-evoked neurotoxicity following exposure to Alzheimer pep-tidesrdquo Journal of Neurochemistry vol 112 no 2 pp 552ndash5632010
[24] N C Garbett N B Hammond and D E Graves ldquoInfluence ofthe amino substituents in the interaction of ethidium bromidewith DNArdquo Biophysical Journal vol 87 no 6 pp 3974ndash39812004
[25] A Karlsson J B Nixon and L C McPhail ldquoPhorbol myristateacetate induces neutrophilNADPH-oxidase activity by two sep-arate signal transduction pathways dependent or independentof phosphatidylinositol 3-kinaserdquo Journal of Leukocyte Biologyvol 67 no 3 pp 396ndash404 2000
[26] F Herencia M L Ferrandiz A Ubeda et al ldquo4-Dimethyl-amino-3101584041015840-dimethoxychalcone down-regulates iNOS expres-sion and exerts anti-inflammatory effectsrdquo Free Radical Biologyamp Medicine vol 30 no 1 pp 43ndash50 2001
[27] N Mateeva M Gangapuram E Mazzio S Eyunni K FSoliman and K K Redda ldquoBiological evaluation of syntheticchalcone and flavone derivatives as anti-inflammatory agentsrdquoMedicinal Chemistry Research vol 24 no 4 pp 1672ndash16802015
[28] S C Morgan D L Taylor and J M Pocock ldquoMicroglia releaseactivators of neuronal proliferation mediated by activationof mitogen-activated protein kinase phosphatidylinositol-3-kinaseAkt and delta-Notch signalling cascadesrdquo Journal ofNeurochemistry vol 90 no 1 pp 89ndash101 2004
[29] J Jebelli T Piers and J Pocock ldquoSelective depletion of micro-glia from cerebellar granule cell cultures using L-leucinemethylesterrdquoThe Journal of Visualized Experiments no 101 Article IDe52983 2015
[30] R Von Bernhardi J E Tichauer and J Eugenın ldquoAging-dependent changes of microglial cells and their relevance forneurodegenerative disordersrdquo Journal of Neurochemistry vol112 no 5 pp 1099ndash1114 2010
[31] C Schwab and P L McGeer ldquoInflammatory aspects ofAlzheimer disease and other neurodegenerative disordersrdquoJournal of Alzheimerrsquos Disease vol 13 no 4 pp 359ndash369 2008
[32] A Henn S Lund M Hedtjarn A Schrattenholz P Porzgenand M Leist ldquoThe suitability of BV2 cells as alternative modelsystem for primary microglia cultures or for animal experi-ments examining brain inflammationrdquo Altex vol 26 no 2 pp83ndash94 2009
[33] S Bano K Javed S Ahmad et al ldquoSynthesis of some novel chal-cones flavanones and flavones and evaluation of their anti-inflammatory activityrdquo European Journal of Medicinal Chem-istry vol 65 pp 51ndash59 2013
[34] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[35] S L Deshmane S Kremlev S Amini and B E Sawaya ldquoMono-cyte chemoattractant protein-1 (MCP-1) an overviewrdquo Journalof Interferon and Cytokine Research vol 29 no 6 pp 313ndash3262009
[36] J Rojas J N Domınguez J E Charris G Lobo M Paya andM L Ferrandiz ldquoSynthesis and inhibitory activity of dimethyl-amino-chalcone derivatives on the induction of nitric oxidesynthaserdquo European Journal of Medicinal Chemistry vol 37 no8 pp 699ndash705 2002
[37] H Hara Y Nakamura M Ninomiya et al ldquoInhibitory effectsof chalcone glycosides isolated from Brassica rapa L lsquohidabenirsquoand their synthetic derivatives on LPS-induced NO productionin microgliardquo Bioorganic amp Medicinal Chemistry vol 19 no 18pp 5559ndash5568 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom







![Supporting Information - Royal Society of Chemistry · S1 Supporting Information Base-promoted [1,4]-Wittig Rearrangement of Chalcone-derived Allylic Ethers Leading to Aromatic -Benzyl](https://img.pdfslide.net/doc/110x75/5e5dc637753a70122b21ac24/supporting-information-royal-society-of-s1-supporting-information-base-promoted.jpg)





















