Embed Size (px)
Citation preview

Introduction____________________________________________________________________________
1
From the Neonatal Research Unit
Department of Women and Child Health
Astrid Lindgren Children Hospital
Karolinska Institutet
Stockholm, Sweden
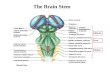
RESPIRATORY ACTIVITY IN
MEDULLA OBLONGATA
and its modulation by
ADENOSINE AND OPIOIDS
Eric HerleniusM.D.
Stockholm 1998

The brainstem and perinatal respiratory activity___________________________________________________________________________
2
This thesis is based on the following papers, which will be referred to in the text by
their Roman numerals:
I Eric Herlenius, Hugo Lagercrantz and Yuji Yamamoto. Adenosine modulates
inspiratory neurons and the respiratory pattern in the brainstem of neonatal rats.
Pediatric Research(1997), 42, 46-53.
II Koichi Takita, Eric Herlenius, Sten Lindahl and Yuji Yamamoto. Actions of
opioids on respiratory activity via activation of brainstemµ-, δ- andκ-receptors;
an in vitro study.Brain Research(1997), 778, 233-241
III Koichi Takita, Eric Herlenius, Sten Lindahl and Yuji Yamamoto. Age and
temperature dependent effects of opioids on medulla oblongata respiratory
activity: An in vitro study in newborn rat.Brain Research(1998), 800, 308-311.
IV Eric Herlenius, Ulrika Åden, Lie-Qi Tang and Hugo Lagercrantz. Adenosine in
the immature rat brain and effects of maternal caffeine intake: development of
respiratory control.Manuscript
V Eric Herlenius and Hugo Lagercrantz. Adenosinergic modulation of respiratory
neurones in thein vitro neonatal rat brainstem.Submitted

Introduction____________________________________________________________________________
3
ABBREVIATIONS
aCSF artificial cerebrospinal fluid
ATP Adenosine-5’-triphospate
Biphasic E Biphasic expiratory neurone (also called Pre-inspiratory)
C4 Cervical ventral root 4
CNS Central nervous system
CPG Central pattern generator
DAGO Try-D-Ala-Gly-[NMephe]-Gly-ol
DPCPX 8-cyclopentyl-1,3,dipropylxanthine
DPDPE [D-pen2,5]-enkephalin
Exp Expiratory related neurone
EPSP excitatory postsynaptic potential
Insp Inspiratory related neurone
Naloxone naloxone hydrochloride
Naloxanazine naloxanazine dihydrochloride
Nor-BNI nor-Binaltorphimine hydrochlorideNTS Nucleus tractus solitarius
PaO2 Partial pressure of oxygen in arterial blood
R-PIA adenosine A1-receptor agonist
(N6-(2-phenylisopropyl) adenosine, R(-) isomer)
Theophylline 1,3,-di-methylxanthine
TTX Tetrodotoxin
U50,488 488 ( trans - (±) - 3,4 - dichloro - N - methyl - N - ( 2 - [ 1 -
pyrrolidinyl] cyclohexyl) benzeneacetamide methanesulfonate)

The brainstem and perinatal respiratory activity___________________________________________________________________________
4
INTRODUCTION .......................................................................................................................................... 5
GENERAL BACKGROUND ...............................................................................................................................5
DEVELOPMENT OF RESPIRATORY CONTROL.................................................................................... 8
Respiratory depression during hypoxia.................................................................................................... 9
OPIOIDS AND RESPIRATORY DEPRESSION....................................................................................................... 11
ADENOSINE AND RESPIRATORY DEPRESSION................................................................................................. 12
THE BRAINSTEM SPINAL CORD PREPARATION................................................................................................ 14
THE PRESENT INVESTIGATION ............................................................................................................ 16
MATERIALS AND METHODS.................................................................................................................. 17
ANIMALS .................................................................................................................................................... 17
BRAINSTEM-SPINAL CORD PREPARATION...................................................................................................... 18
BLIND WHOLE CELL PATCH CLAMP ............................................................................................................... 19
HISTOLOGY ................................................................................................................................................ 21
BAROMETRIC PLETHYSMOGRAPH................................................................................................................. 21
DRUGS....................................................................................................................................................... 22
DATA ANALYSIS AND STATISTICS ................................................................................................................. 23
RESULTS AND DISCUSSION .................................................................................................................... 24
ASPECTS ON METHODOLOGHY..................................................................................................................... 24
Time dependency of opioid and adenosinergic drugs............................................................................. 25
Perinatal age and experiments .............................................................................................................. 25
OPIOIDS AND RESPIRATORY DEPRESSION....................................................................................................... 26
Opioids inhibit respiration byµ- includingµ-1 andκ-opioid receptors ................................................. 26
Temperature and respiratory control..................................................................................................... 27
Do opioids and adenosine interact in respiratory depression?............................................................... 30
ADENOSINE AND RESPIRATORY DEPRESSION................................................................................................. 30
Adenosine modulates respiratory activity by acting directly on brainstem respiration-related neuronal
networks................................................................................................................................................ 31
Adenosine modulates the synaptic activity in brainstem respiratory neurones....................................... 32
Adenosine and postsynaptic depression of respiration-related neurones................................................ 36
Inhibition of expiratory neurones does not abolish the respiratory rhythm............................................ 37
DEVELOPMENT OF RESPIRATORY CONTROL................................................................................................... 39
The neuronal network generating respiratory rhythm does not undergo major changes in the early
perinatal period. ................................................................................................................................... 39
The development of respiratory activity and chronic maternal intake of caffeine................................... 40
GENERAL DISCUSSION............................................................................................................................ 41
CONCLUSIONS........................................................................................................................................... 43
REFERENCES ............................................................................................................................................. 46
ABSTRACT……………………………………………………………………………………………………..56

Introduction____________________________________________________________________________
5
INTRODUCTION
Breathing is a fundamental physiological process produced and controlled by the
nervous system which must be properly set into action from the moment of birth.
Currently it is not exactly known how breathing is initiated and basic mechanisms,
involved in the complex and fine-tuned regulation of respiration are not clearly
delineated yet. A better understanding of normal development of respiratory control
and its disturbances may lead to improved treatment and outcome of clinical
conditions such as repeated episodes of apnoea in preterm babies, sudden infant death
syndrome and postoperative hypoventilation. Hence, the study of mechanisms for
central pattern generation of breathing is of great concern, not least in the perinatal
period, which is the main theme in the present thesis focused on respiratory control
and its modulation by adenosine and opioids.
GENERAL BACKGROUND
It was not until the French revolution that it was understood, during the use of
the guillotine, that mammals do not need their heads to breathe. The French physician
Le Gallois (1770-1814) established that the respiratory centre is localised in the
medulla oblongata in a series of animal experiments (Le Gallois, 1812). This was the
first time that an area within a major subdivision of the brain had been defined
accurately by experimentation as having a specific function (Morton and Garrison,
1991). Le Gallois declared that life in an animal or in any of its organs depends on two
obligatory conditions. One is the integrity of the medulla oblongata and its nervous
output. The second is the circulation of arterial blood to the organ and the medulla
oblongata.
Since the discoveries of Le Gallois our understanding of the body, the brain and
the control of autonomic functions has increased enormously, for reviews see (von
Euler, 1991; Specket al., 1993; Bianchiet al., 1995).
In order to investigate different levels of neuronal networks or single neurones,
animal models have to be used. The anaesthetised or decerebrated cat was the model
of choice for the study of respiratory control from 1920 up to about 1985. Thesein
vivo studies localised the neuronal populations that contain the basic circuitry for
respiratory rhythm in the brainstem as well as the firing pattern, interconnection and
projections of respiratory neurones found in this region (Feldman, 1987; Bianchiet al.,

The brainstem and perinatal respiratory activity___________________________________________________________________________
6
1995). Based on such studies it was determined that the generation of breathing
rhythmogenesis does not critically depend on extrinsic feedback loops or reflexes,
provided the excitability of these mechanisms is kept sufficiently high by adequate
biasing inputs (von Euler, 1980).
During the last decade, thanks to the availability of novel techniques, especially
in vitro en bloc and slice preparations, our understanding has increased concerning
cellular and synaptic physiology of brainstem respiratory neurones. Thesein vitro
preparations make it feasible to examine cellular and sub-cellular levels of respiratory
control. Nevertheless, we still do not know how the neuronal networks in the medulla
oblongata generate respiratory rhythm even though several hypotheses have been
proposed. (Richteret al., 1992; Bianchiet al., 1995; Onimaruet al., 1997; Rekling and
Feldman, 1998). The classical concept of a reciprocal inhibition between two
symmetrical populations of inspiratory and expiratory premotor neurones, the “half-
centre model”, has been abandoned. Instead, the prevailing theories divide the
respiratory cycle into three phases (inspiratory, passive and active expiratory (Richter,
1982). Today, two main hypotheses exist: 1) Site hypothesis: the preBötzinger
complex is the site for respiratory rhythm generation (Smithet al., 1991) and 2)
rhythmogenesis hypothesis: pacemaker or group pacemaker neurones are the cellular
kernel for respiratory rhythm (Reklinget al., 1996) (Onimaruet al., 1988) The
preBötzinger complex in the rostral ventrolateral medulla oblongata seem to have an
obligatory role in respiratory rhythmogenesis, whereas more caudal and dorsal
medullary structures do not. Reciprocal synaptic inhibition between groups of
respiratory neurones is not the cellular basis for respiratory rhythm. This has led to that
neurones intrinsically capable of generating cyclic discharges, i.e., pacemaker
neurones have been proposed as candidates for rhythm generation (Onimaru, 1987) for
reviews see (Rekling and Feldman, 1998) and (Bianchiet al., 1995).
Thus, it has been established that a highly sophisticated central neuronal network
is responsible for the central control of breathing. This central network is able to
continuously adjust breathing to the requirements of the internal and external
environment. Central control implies that the central nervous system is intrinsically
capable of providing the proper timing of muscle activation. Fine-tuning of the central
output is attained by a wide range of vagal afferents from the airways and lungs
relaying sensory information to the nucleus tractus solitarius (NTS) in the dorsal
medulla oblongata. In addition, the arterial baro- and chemoreceptors, both those
relaying via the vagus from the aortic arch and those relaying via the IXth cranial

Introduction____________________________________________________________________________
7
nerve from the carotid bifurcation, send their fibres to the NTS. In addition,
descending inputs from cortex, diencephalon and the cerebellum also influence the
medullary network (Euler and Lagercrantz, 1987) Although suprapontine and sensory
input modulates respiratory rhythm and adapts breathing, these influences only
modulates the ongoing central respiratory rhythm generation. It is important, therefore,
to understand the processes underlying the central respiratory rhythm generation and
its intrinsic control. This thesis focuses on the intrinsic respiratory control system in
the brainstem.
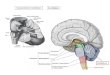
(Modified afterPaxinos and Watson, 1996 and refs.(Arataet al., 1990; Brockhauset
al., 1993).

The brainstem and perinatal respiratory activity___________________________________________________________________________
8
DEVELOPMENT OF RESPIRATORY CONTROL
The ability of central networks to produce rhythmic respiratory motor behaviour
is a well conserved property of the brainstem reticular formation between species
(Borday et al., 1997). Since 1991 an area in the ventral group of respiration related
neurones, the pre-Bötzinger complex, has been considered by most researchers to be
the crucial site of this rhythmogenesis (Smithet al., 1991). This area was identified by
a series of transection studies in thein vitro brainstem spinal cord preparation from
neonatal rats that is the main tool in this thesis. During recent years this area has been
reported to also be importantin vivo and in other species (Paton 1997, (Pierreficheet
al., 1998; Ramirezet al., 1998c). The ventral group of respiratory neurones (VRG)
extends through the whole length of the ventrolateral medulla, partly along the nucleus
ambiguous (Ezure, 1990) but it is mainly in the rostral VRG where bulbospinal
neurones recide (Fig 1). The nucleus tractus solitarius in the dorsal group of
respiratory neurones (DRG) receive afferent input from the lungs and also receive
input from peripheral baro and chemoreceptor. It is thus an important relay for
integrating afferent input that can influence respiration. However, in the rat the DRG
makes little or no contribution to the respiratory drive transmitted from medulla to
spinal motoneurones (Bianchi 1995).
Respiratory rhythm generation is established long before birth, and in the human
foetus respiratory movements already occur at the 11th week of gestation (de Vrieset
al., 1982). These movements are necessary for normal lung development (Kitterman,
1996). Foetal breathing movements are episodic and are progressively more inhibited
towards the end of pregnancy when periods without respiratory movements (apnoea)
dominate (Maloney, 1975 However, from the dramatic moment of birth, these
respiratory neuronal networks need to function and continuously generate a respiratory
rhythm to sustain oxygenation and metabolism. The mechanisms of this transition from
episodic to continuous breathing is still unclear. Respiration must also have the
capacity to respond to changes in the external and internal environments to maintain
body homeostasis. The rhythmic respiratory movements in foetuses, as well as the
breathing in neonates and adults, are governed from neuronal networks in the
brainstem which generate the respiratory rhythm. Thus, during the last decades
researchers have tried to understand how thisnoed vital develops and functions

Introduction____________________________________________________________________________
9
through different experimental approaches, for review see (Mitchell, 1997) and (Speck
et al., 1993).
Respiratory depression during hypoxia
Breathing movements before birth are inhibited by hypoxia (Boddyet al., 1974;
Johnston, 1991). This is functional in foetal life when processes consuming oxygen
have to be turned off if they are not vital for survival (Johnston, 1991; Hanson, 1996).
However, this apnoeic response to hypoxia may be less appropriate after birth. During
the early postnatal period respiratory control is still different than in adults. Both adult
and new-born animals respond to hypoxia with a biphasic change of respiration
(Schwieler, 1968; Lawson and Long, 1983; Runoldet al.Fredholm, 1989). There is an
initial increase of respiration due to stimulation of the peripheral chemoreceptors
(Schwieler, 1968), activation of nervous structures localised rostral to the brainstem
(Eldridge et al., 1981) and structures within the brainstem (Ramirezet al. 1998b).
However, in newborn animals the response to hypoxia is similar to that of the foetus
and the secondary depression of respiration is pronounced and ventilation falls below
normoxic levels within minutes after initiation of hypoxic exposure. During severe
hypoxia or anoxia there are progressive changes in the ventilatory pattern, from
hyperpnoea to a sustained pause of respiration, which is then, after a period of
expiratory apnoea, followed by gasping. Respiratory gasping is characterised by a
series of brief bursts of phrenic activity with sudden onsets followed by a rapid decline
and absence of expiratory activity (Guntheroth and Kawabori, 1975).
What are the brain structure(s) and neuromodulators that are involved in the
hypoxia-induced respiratory depression? Several possibilities have been considered as
mechanisms behind the secondary depression of respiration, for review see (Lawson
and Long, 1983; Neubaueret al., 1990; England, 1993). These hypotheses can be
divided into those stressing inhibitory supramedullary input to the brainstem as vital
for the depressioni.e. (Johnston, 1991; Waiteset al., 1996) (Hanson, 1996; Okadaet
al., 1998) and those emphasising local neurochemical processes in the brainstem
(Moss et al., 1986). Recent findings indicate that even in the reducedin vitro
preparation the hypoxic response is biphasic both at the motor output level and within
the respiratory network (Brockhauset al., 1993; Ramirezet al., 1998a; Ramirezet al.,
1998b). Both afferent and supramedullary input as well as intrinsic brainstem
processes are thus probably involved in this complex behaviour. Diverse structures at

The brainstem and perinatal respiratory activity___________________________________________________________________________
10
several CNS levels are thus involved in the response to hypoxia but the basic
mechanism(s) behind the central inhibition of breathing are still unclear. A proposed
intrinsic brainstem mechanism is a depression of central respiratory neurones mediated
by the release of several neurotransmitters/modulators. These include GABA
(Neubaueret al., 1990) and prostaglandinsPGE2 (REF). Naloxone and theophylline
partially abolish the respiratory inhibition caused by asphyxia, suggesting that opioids
and adenosine are involved (Grunsteinet al., 1981; Chernick and Craig, 1982; Hedner
et al., 1984; Runoldet al., 1989)
Figure 2 Depicts the biphasic ventilatory response to hypoxia (10-15 % O2 in N2).
The inhibitory phase is more pronounced in immature animals. Naloxone or
theophylline can attenuate the inhibition, suggesting that opioids and adenosine are
involved in the secondary inhibition. Administration of dipyridamole, which augments
the endogenous levels of adenosine induces a more pronounced hypoxia-induced
inhibition. (Modified after Lagercrantz, 1987, Grunstein, 1981, Darnall 1985).
0
0,5
1
1,5
-2 3 8
Control
Naloxone /Theophylline
Dipyridamole
Rel
ativ
eve
ntil
ation
Hypoxia min

Introduction____________________________________________________________________________
11
OPIOIDS AND RESPIRATORY DEPRESSION
Opioid-induced respiratory depression is well known from clinical situations as
well as from experimental studies (Yeadon and Kitchen, 1989). Administration of
exogenous opioids is associated with depression of central respiratory activity in
mammals.
Naloxone reverses neonatal depression caused by foetal asphyxia (Chernick and
Craig, 1982). Thus, opioids are thought to be involved in hypoxia-induced respiratory
depression (Mosset al., 1987) Nevertheless, the role of endogenous opioids in basal
respiratory control has not yet been completely established. Some studies using
naloxone have demonstrated that endogenous opioids do not have any significant
influence on the basal regulation of breathing in rats (Steinbrooket al., 1984; Olson,
1987). Others have come to the opposite conclusion (Isom and Elshowihy, 1982).
Recent findings by Greeret al. (Greeret al., 1995) indicate that naloxone has no effect
on medullary respiratory control duringin vivo or in vitro experimental conditions in
newborn rats.
Most studies thus indicate that endogenous opioids are not involved during
eupneic breathing but have an important role during hypoxia and postoperative
hypoventilation. Opioids have also been proposed to be involved in the pathogenesis
of sudden infant death syndrome (Orlowski, 1986; Morinet al., 1992).
Three major classes of opioid receptorsµ, δ, and κ have currently, been
identified, characterised and cloned, all with putative receptor subtypes. All are seven-
transmembrane proteins and members of the G-protein coupled receptor superfamily.
Endogenous opioid peptides with distinctive selectivity profiles exist namely the
enkephalin (µ), endorphin (δ) and dynorphin (κ) groups.µ-receptor binding sites are
present during mid-foetal time and have a high density in cardiorespiratory-related
brainstem nuclei, whereas theδ-opioid receptors primarily appear during the postnatal
period in rats (Xia and Haddad, 1991).
Opioid-induced respiratory depression has been suggested to be caused by direct
actions on the brainstem (Flórez and Hurlé, 1993). Most previousin vivo studies have
suggested thatµ- and δ-opioid receptors participate in opioid-induced respiratory
depression, whileκ-opioid receptors are not involved (Shooket al., 1990; Flórez and
Hurlé, 1993). However,κ- and µ-opioid receptors have a similar distribution in the
brainstem (Mansouret al., 1988) and due to the somewhat unselective receptor
agonists used, it is still debated if onlyµ-receptors are involved in the respiratory
depression caused by opioids. Based on a subdivision intoµ-1 andµ-2 isoreceptors

The brainstem and perinatal respiratory activity___________________________________________________________________________
12
(Pasternaket al., 1980), some authors have claimed that opioid-induced respiratory
depression is mediated byµ-2 receptors (Linget al., 1985; Ling et al., 1986).
However, the exact actions of opioids on respiratory activity via activation ofµ-, δ-
andκ-opioid receptors in the medulla oblongata remain to be elucidated.
ADENOSINE AND RESPIRATORY DEPRESSION
As early as 1929, Drury and Szent-Györygi reported that adenosine can inhibit
respiratory and intestinal movements as well as decrease heart (Drury and Szent-
Gyrörgyi, 1929)). Adenosine is a constituent of all body fluids, including the
extracellular space of the central nervous system. It has multiple effects on organs
and cells of the body (Berne, 1986) Thus, its levels are tightly regulated by a series
of enzymatic steps (Fredholm, 1995). Adenosine can be regarded more as a
neuromodulator in that it does not seem to be stored in vesicles with a regulated
release from nerve terminals. Adenosine is produced by dephosphorylation of
adenosine monophosphate (AMP) by 5`nucleotidase, an enzyme occurring in both
membrane-bound and cytosolic forms (Brundege and Dunwiddie, 1997). Degradation
of intra- and extracellular ATP is the main source of extracellular adenosine
(Dunwiddie and Fredholm, 1997). Specific bi-directional transporters maintain intra-
and extracellular concentrations of adenosine at similar levels. During basal
conditions adenosine levels are 30-300 nM and can rise following stimuli that cause
an imbalance between ATP synthesis and ATP breakdown. Thus, the levels during
ischemia or hypoxia can rise 100-fold (Winnet al., 1981; Fredholm, 1995).
[ATP] i
[AMP] ⇐⇐⇐⇐ [ATP] e.
Inosine⇐⇐⇐⇐ [Adenosine]i ⇔⇔⇔⇔ [Adenosine]e .
bidirectionaltransporter
Figure 3. Schematic representation of the degradation of ATP to adenosine

Introduction____________________________________________________________________________
13
Hypoxia can trigger apnoea in the human neonate (Rigattoet al., 1972; Miller
and Martin, 1992), whereas the adenosine antagonist theophylline blocks hypoxia-
induced depression of breathing in rats (Neylon and Marshall, 1991), rabbits (Runold
et al., 1989) and piglets (Mosset al., 1987; Lopeset al., 1994). The therapeutic effect
of theophylline on neonatal apnoea has thus been suggested to be due to its
antagonistic action on adenosine receptors (Lagercrantz Yet al., 1984; Darnall, 1985;
Hedneret al., 1985). Several lines of evidence indicate that adenosine can inhibit
respiration-related neurones in the brainstem (Eldridgeet al., 1983; Thomaset al. ,
1994). However, adenosine also decreases metabolism and oxygen consumption
through inhibiting lipolysis and non-shivering thermogenesis (Ballet al., 1996).
Neonatal animals respond to hypoxia with decreased body temperature. Thus, it has
been suggested that the main effect of adenosine in depressing breathing is indirect by
a decrease of oxygen consumption (Lagercrantzet al., 1986).
To date four distinct receptor subtypes have been identified by cloning, denoted
A1, A2a, A2b and A3-adenosine receptors. All subtypes belong to the family of rhodopsin
G-protein-coupled receptors (Fredholm, 1995). Basal levels of adenosine can act on
A1- and A2a–receptors whereas A2b-receptors only are activated at pathological
adenosine levels. A2a receptors are coupled to GTP-binding (G) proteins classified as
Gs, because of their stimulatory effect on adenylyl cyclas, and mainly expressed in
dopamine-rich regions such as the striatum. Adenosine A1-receptors are coupled to Gi(1-
3) (inhibitory to adenylyl cyclas) or Go (no effect on adenylyl cyclas) proteins and are
ubiquitously expressed in the CNS with higher expression in regions such as the
cortex, hippocampus and cerebellum (Fredholm, 1995). A1-receptors have been
characterised by behavioural, biochemical and anatomical studies, for a review see
(Dunwiddie and Fredholm, 1997). A1-receptors can modulate synaptic transmission in
the CNS through several different mechanisms. Among these are decreased
neurotransmitter release (Mynlieff and Beam, 1994) and reduction of neuronal
excitability through hyperpolarisation of neurones (Thompsonet al., 1992). Adenosine
have been demonstrated to modulate cardiovascular control through its action in the
rostral ventrolateral medulla (Thomas and Soyer, 1996). Responses to adenosine are
different even in neuronal populations using the same transmitter, which makes it
difficult to directly extrapolate data obtained from one group of neurones to another.
Thus, a modulation shown for parts of a neuronal network may not be applicable when
all its components are examined.
Adenosine is also an important neuromodulator in foetal life (Bissonetteet al.,
1990; Koos and Matsuda, 1990) and at birth (Irestedtet al., 1989). Changes of

The brainstem and perinatal respiratory activity___________________________________________________________________________
14
adenosinergic tonic activity due to caffeine, an adenosine antagonist, may interfere in
the communication between developing neurones leading to long term sequele.
Caffeine, is the most widely consumed neuroactive substance worldwide. Negative
effects on fertility, birthweight, risk for prematurity and congenital malformations of
high doses of caffeine have been demonstrated in animals, for review see (Nehlig and
Debry, 1994). There is however little evidence that normal human caffeine
consumption has any of these consequences.
Does maternal intake of adenosine antagonists, such as caffeine affect the
development of respiratory rhythm generation and control during foetal and neonatal
life? High doses of caffeine intake during gestation or the early postnatal period affect
postnatal behaviour in rats (Holloway and Thor, 1982; Guillet, 1990; Devoeet al.,
1993; Guillet and Dunham, 1995). Effects of more moderate prenatal caffeine intake
on postnatal behaviour have also been studied, and several investigators have
described behavioural hyperactivity in developing rodents (Sobotka, 1979; Sinton,
1981; Holloway Jr, 1982; Holloway Jr and Thor, 1982). If low doses of caffeine
during gestation can induce long-term changes in adenosine receptor expression and if
the development of respiratory control is affected this may have important clinical
implications.
THE BRAINSTEM SPINAL CORD PREPARATION
Studies on respiratory control and the highly sophisticated neuronal networks
that generate this complex behaviour can be performed at different levels: whole-body,
organ, neuronal networks, cellular, synaptic and molecular. During the last 15 yearsin
vitro experimental preparations have proved invaluable in unravelling the mechanisms
of central respiratory control. Among the powerful advantages ofin vitro preparations
are: 1) direct access to neurones of interest; 2) mechanical stability, allowing
intracellular recordings with whole-cell patch techniques; and 3) improved control of
the environment surrounding the neurones.
A brainstem-spinal cord preparation from neonatal rats was first used by Suzue
and co-workers (Suzue, 1984). This en bloc brainstem spinal cord preparation
generates respiration-related rhythmic activity for several hours when perfused with
oxygenated artificial cerebrospinal fluid (aCSF). Compared to thein vivo situation, the
respiratory frequency is low. Inspiratory-phase dischargein vitro is rapidly peaking-
slowly decrementing instead of a short duration, augmenting discharge envelope
(Smithet al., 1990). The transformation of the respiratory motor patternin vivo to that

Introduction____________________________________________________________________________
15
of the isolated and reduced brainstem spinal cord preparation can be explained by
several facts: 1) it does not have suprapontine inputs and can be further reduced by
removing the pons; 2) removal of vagal mechanosensory afferent input; 3) low
temperature (25-32oC) (Smithet al., 1990; Onimaru, 1995a).
Respiratory rhythm generation in neonatalin vitro models differs from that of
adult in vivo models. In this neonatal model Chloride-mediated inhibition is not
necessary for the respiratory rhythm whereas it probably is in the adult (OnimaruArata
and Homma, 1990; Feldman Smith and Let al.iu, 1991; Onimaruet al., 1997;
Ramirezet al., 1997). However, the isolated respiratory network in the brainstem is
sufficient to generate the essential features of respiratory activity at the cellular and
system levels that have previously been described for the intact respiratory system.
The relevance of neuronal mechanisms studied in this reduced preparation to
those operating in the intact neonatal or adult rat has been determined by several
studies during the last decade, for reviews see (Smithet al., 1990; Onimaruet al.,
1997).
Synaptic events during the respiratory cycle have not been thoroughly analysed
in any neonatal mammalian species, so the correspondence between the synaptic
events producing the cycle phasing in the neonatal and adult system remains to be
established (Smithet al., 1990) The studies that have been performed reveal
remarkable similarities in the synaptic interactions between respiratory neurones
although recordings fromin vivo cat andin vitro rodents were compared. (Ramirezet
al., 1997).
Neonatal rats, as all mammals, are born with a functional central pattern
generator for respiration. However, the CNS and its connections with the internal and
external environments develops before and after birth. As described in the previous
pages it is known that both adenosine and opioids have different effects on respiration
in the neonate compared to the adult mammal. Are these changes in modulatory
effects initiated at the moment of birth, during the first postnatal days or are they
dependent on progressive maturation of neuronal networks, receptor expression and
affinity? We have in the following studies used the brainstem spinal cord preparation,
complemented within vivo andin situ techniques, to address some of these questions.

The PresentInvestigation
_____________________________________________________________________
16
THE PRESENT INVESTIGATION
The overall aim of the present study was to further elucidate the perinatal development
of central respiratory activity and its control. Specifically, the aims where to:
• investigate if there is a transition at birth of the properties of the isolated neuronal
network that generate respiratory rhythm. (IV)
• determine if adenosine and opioids inhibit respiration by acting directly on the
central neuronal networks generating respiratory activity. (I-V)
• determine if the adenosinergic and opioidergic modulation of respiratory activity in
the medulla oblongata changes during the first days after birth. (I, III-V)
• investigate if chronic maternal intake of caffeine (a typical adenosine antagonist),
during gestation and early postnatal life, alters the development of respiratory
control. (IV)
• investigate the mechanisms behind and at which level adenosinergic modulation of
central respiratory activity occurs, i.e. pre- vs. postsynaptically.(I and V)

Material and Methods____________________________________________________________________________
17
MATERIALS AND METHODS
ANIMALSRats were bred by certified animal supplier and stored at the animal facilities
according to local regulations. Neonatal rats were kept with their mothers until the
experiments started. Pups were examinedin vitro at postnatal age 0-4 days and at
embryonic (E) day 18-21. Rat brains were examined at E14, E18, E 21, exactly 2
hours and 24 hours after vaginal delivery and postnatal day (P)3 and P7.
In the experiments performed in paper IV, examining the development of
respiratory control, timed pregnant rats were used. The day when a vaginal plug was
found was designed as embryonic day 0. The regional animal ethics committee
approved the experiments, which followed the European Community regulations.
Paper Questions n Methods
I Early postnatal adenosinergic
modulation of respiratory activity62 Brainstem spinal cord
preparation (BSP)
II Opioid-receptors 156 BSP
III Opiods-temperature and age 273 BSP
IV Development of respiratory control
and its adenosinergic modulation
Development of adenosine A1-
receptors and maternal caffeine
80
40
200
BSP
Barometric plethysmograph
In situ hybridisation and
receptor autoradiography
V Mechanisms of central adenosinergic
respiratory modulation?111 BSP
Blind whole cell patch clamp
An extended description of the blind whole-cell patch clamp technique, the
accompanying histological method as well as thein vivo experiments follows below.
The other methods used are described in the papers included.

The brainstem and perinatal respiratory activity____________________________________________________________________________
18
BRAINSTEM-SPINAL CORD PREPARATION
The in vitro experiments were performed on the brainstems and spinal cords of
newborn (0- to 4-day-old) Bkl:S-D or Wistar rats. Under deep ether anaesthesia, the
brainstem-spinal cord was dissected and isolated as described previously (Suzue,
1984). The brainstem was then rostrally decerebrated between the VIth cranial nerve
roots and the lower border of the trapezoid body. This preparation was subsequently
transferred to a 2-ml chamber, where it was continuously perfused at a rate of 3.0-3.5
ml min-1 with artificial cerebrospinal fluid (aCSF) equilibrated with 95% O2 and 5%
CO2at 26-28oC to give a pH of 7.4. Foetal rats were obtained from plugged dams by
caesarean section. Respiratory activity was recorded employing suction electrodes
applied to the proximal ends of cut C4 or C5 ventral roots containing respiratory
motoneurone axons that innervate the intercostal muscles and the ribcagein vivo. (Fig.
2). The phasic C4/C5 activity corresponds to phrenic nerve discharges (Suzue, 1984;
Smith et al., 1990). The regularity of respiratory activity was quantified as coefficient
of variation (CV) of the interval between C4 burst discharges.
For further details see paper I-V
Figure 4 Brainstem-spinal cord preparation

Material and Methods____________________________________________________________________________
19
BLIND WHOLE CELL PATCH CLAMP
Whole cell recordings from respiratory neurones were obtained using the
modified blind patch clamp technique (Blantonet al., 1989; Onimaru and Homma,
1992). Briefly, patch pipettes were pulled in one step from borosilicate glass
(GC100TF-10, outer diameter 1.0 mm, with a filament; Clark Electromed, Reading,
UK) using a vertical puller (PE-2; Narishige, Tokyo, Japan). The electrode tips had
inner diameters of 1.2-2.0µm and a DC resistance of 4-8 MΩ. The electrodes were
filled with a solution consisting of (mM): potassium gluconate 120,
ethylenebis(oxonitrilo)-tetraacetate (EGTA) 10, 4-(2-hydroxyethyl)-1-
piperazineethane-sulphonic acid (HEPES) 10, CaCl2 1, MgCl2 1 and Na2-ATP 1. KOH
was used to adjust the pH to 7.3. In a number of experiments this electrode solution
also contained 0.2 % Lucifer Yellow (LY; Sigma, St. Louis, Mo, USA). The solution
was routinely filtered through a Millipore filter (pore size 0.45µm) immediately
before its introduction into the electrodes.
A small area of thepia mater was removed using a glass needle and the
electrodes were inserted through this area into the ventral medulla. A slight positive
pressure (5-15 cm H2O) was applied during advancement of the electrode to keep the
electrode tip clean. Extracellular signals and intracellular membrane potentials were
measured with a voltage clamp amplifier (Nihon Koden, CEZ-311). Respiration-
related neurones were sought in RVL (Arataet al., 1990) in the region of the ventral
respiratory group (Richteret al., 1992), while monitoring extracellular signals
amplified with a loudspeaker (Onimaru, 1995b). In whole cell experiments, when such
a respiration-related neurone was found, a slight negative pressure was applied. The
resulting formation of a gigaohm seal (>1GΩ) was monitored and confirmed by
applying a hyperpolarising current pulse (0.1 nA; duration, 30 ms 2 Hz). When this
seal had been established, the pressure was returned to zero. Rupture of the cell
membrane was achieved by applying negative pressure (50-80 cm H2O) often together
with a single hyperpolarising current pulse (0.5-0.8 nA; duration 30 ms). When whole
cell recording had been established the pressure was rapidly returned to zero ±2-3 cm
H2O. Series resistance (20 - 50 MΩ) was compensated by a bridge balance circuit. The
voltage deflections induced by 1 Hz 30 ms hyperpolarising pulses were monitored and
compensated for by eye.

The brainstem and perinatal respiratory activity____________________________________________________________________________
20
Neurones were identified and classified on the basis of their characteristic firing
patterns and the temporal correlation of this activity to the respiratory cycle of C4/C5
activity. Inspiratory neurones (Insp) receive excitatory synaptic input and discharge
action potentials during inspiratory phrenic (C4/C5) activity. Expiratory neurones
(Exp) discharge action potentials between the inspiratory phases and are inhibited
during the inspiratory phase (Shao and Feldman, 1997). Biphasic expiratory neurones
(also classified as Preinspiratory by Onimaruet al.) are characterised by pre- and
postinspiratory excitation and inspiratory-related inhibition (Onimaruet al., 1990;
Smithet al., 1990).
Inspiratory neurones were further classified into three subtypes, according to
previous classifications performed with this preparation (Onimaruet al., 1996;
Onimaru et al., 1997). Type I neurones (Insp I) receive excitatory postsynaptic
potentials (EPSPs) prior to the onset, as well as after the termination of C4 activity,
whereas type III neurones (Insp III) are hyperpolarised by synchronised inhibitory
postsynaptic potentials (IPSPs) during the pre- and postinspiratory phases. Insp
neurones, which only exhibited EPSPs during the inspiratory phase and no
hyperpolarisation during the pre- or postinspiratory phase, were classified as type II
neurones (Insp II ).
Respiratory cycle
Neuronal type Inspiratory Peri-inspiratory Expiratory-I Expiratory-II
Insp type-I ++ (+) 0 0Insp type-II ++ 0 0 (+)Insp type-III ++ - 0 0Expiratory-t - + + +Expiratory-l - - 0 +Biphasic E - + 0 0
Table 1 Respiration-related neurons in the brainstem spinal cord preparation, here
categorised in relation to the C4 inspiratory activity according to Onimaruet al.
(Onimaru 1997)- = hyperpolarised, Inspiratory (Insp), Biphasic expiratory (Biphasic
E), tonic Expiratory (Expiratory-t), late Expiratory (Expiratory-l)
The current-voltage (I-V) relationship was determined by injection of an inward
current (0.02-0.08 nA; duration 100 ms) during silent phases between bursts or during

Material and Methods____________________________________________________________________________
21
negative holding potentials (-50 mV) in the case of certain expiratory neurones. Input
resistance (Rm) was calculated from the slope of a least-squares regression line fitted to
the data. In Exp neurones, the frequency (action potentials / min) of expiratory
neuronal activity was examined. During the experiments, signals were displayed on a
chart recorder, monitored using an oscilloscope, digitalised (Digidata 1200B, Axon
Instr., Foster, CA, U.S.) and stored on DAT tape (RD-120TE, TEAC, Tokyo, Japan)
or a hard disc for off-line analysis.
HISTOLOGYAfter completing thein vitro recordings, those preparations containing neurones
filled with Lucifer Yellow were removed from the recording chamber and placed in a
solution of cold (4OC) Lillie solution (10% formalin in phosphate buffer, pH 7.0) for
at least 48 hours. The preparation was then rinsed and placed serially for 10 min each
in 5, 10 and 15% sucrose in 0.1 M phosphate buffer (PB, Sorensen-Gomori, pH 7.2).
The sample was subsequently stored in the 15% solution at 4oC for at least 12 hours
before 70-µm sections were cut on a cryostat. Sections were mounted on glass slides,
coated with PBS and propriogallate. The intracellularly marked Lucifer Yellow-
neurones were photographed and reconstructed with the aid of a camera lucida
attached to a fluorescence microscope (Olympus, Tokyo, Japan). Counterstaining was
performed with 2% neutral red. After dehydration in a series of solutions containing
increasing ethanol concentrations, sections were clarified with xylene and a cover was
placed on them.
BAROMETRIC PLETHYSMOGRAPHIn vivo ventilation was monitored with a barometric plethysmograph, a procedure
which allows non-invasive recording of respiration in unanesthetized animals
(Drorbaugh and Fenn, 1955). This method is based on the fact that warming a gas
maintained at a constant volume increases its pressure. Thus, during inspiration,
pressure inside the chamber increases and during expiration it decreases. The animals
were placed in a plexiglass chamber (160 ml in volume) which, together with an
identical reference chamber, was connected to a highly sensitive differential pressure
transducer (Validyne DP103). Pressurised gas from gas tubes was continuously fed to
the recording chamber and removed with a vacuum pump. The resistance of the inlet
and outlet tubing was adjusted with needle valves in order to maintain approximately
atmospheric pressure in the recording chamber. The rate of gas flow through the
recording chamber was about 1.2 l min-1. The time constant of the recording system

The brainstem and perinatal respiratory activity____________________________________________________________________________
22
was 0.2 seconds. The pressure signal was amplified and recorded by an on-line
computer. The measurements of breathing were only semiquantitative because core
body temperature was not measured and because of the short time constant of the
recording system. Animal and chamber temperatures were recorded using digital
thermometers with an accuracy of 0.1oC. At the onset of the experiment the chamber
temperatur was kept at 29.5 ± 0.5oC via a thermoradiator and a heating pad.
DRUGSThe pH was assessed and adjusted to 7.4 with 95% O2 and 5% CO2 prior to bath
application of drugs. Sufficient time for achievement of the steady-state drug response,
determined for each drug used was allowed to elapse before determination of the drug
response. In paper IV dams were given caffeine (0 or 0.3g/L) in the drinking water from
embryonic day 2 (E2) throughout gestation and postnatal life. The daily intake of water
was measured in all litters. Blood from five litters were collected in heparinized plastic
tubes and centrifuged. Plasma concentrations of caffeine and its metabolites theophylline,
theobromine and paraxanthine were analysed using HPLC. In paper II 17 animals received
a subcutaneous injection (s.c.) of naloxanazine 35 mg/kg ( =5 ml naloxanazine solution/kg)
and 17 animals received saline 5 ml/kg s.c. 22-26 hours after these injectionsin vitro
studies were performed. See Table 3 for drugs used.

Material and Methods____________________________________________________________________________
23
Table 3. Major pharmacological tools used in this thesis
Compound Selectivity
Adenosinergic
R-PIA A1-adenosine receptor agonist
Dipyridamol nucleoside transport inhibitor
DPCPX A1-adenosine receptor antagonist
Theophylline Adenosine receptor antagonist
Caffeine Adenosine receptor antagonist
Opioidergic
Morphine opioid receptor agonist
DAGO µ-opioid receptor agonist
DPDPE δ-opioid receptor agonist
U50,488 κ-opioid receptor agonist
Naloxone opioid receptor antagonist
Nor-BNI κ-opioid receptor antagonist
Naloxanazine µ-1 opioid receptor antagonist
DATA ANALYSIS AND STATISTICS
Off-line analysis was performed using a personal computer and the commercially
available programs Axoscope (Axon Inc., Foster, CA, USA), Origin (Microcal
Software Inc., Northampton, MA, USA) and JMP (SAS Inst. Inc., Cary, NC, USA).
The results are presented as means ± standard deviations or standard errors of the
mean when small or non-normally distributed data are presented. After analysis of the
variance by theF-test, statistical analysis was performed using the multivariate
analysis of variance (MANOVA) repeated measure design, two-tailed paired Student’s
t-test or Wilcoxon's signed-rank test (when variances were unequal). The differences
between several independent means were analysed by comparing all pairs using the
Tukey-Kramer HSD test. Spearman rank nonparametric correlation was performed on
measured variables with respect to postnatal age, in order to evaluate a possible
dependency of the results on age. A P value of <0.05 was considered to be statistically
significant.

Brainstem perinatal respiratory activity___________________________________________________________________________
24
RESULTS AND DISCUSSION
ASPECTS ON METHODOLOGHY
The region where respiration-related neurones in the neonatal rat are located is in
the ventrolateral reticular formation at depths of 50-500µm. In the brainstem spinal
cord preparation there are substantial gradients of PO2, PCO2, K+ and pH from the
surface (Brockhauset al., 1993; Okadaet al., 1993; Voipio and Ballanyi, 1997) to the
respiratory network but the oxygenation permits the neurones to operate under
conditions of aerobic metabolism (Brockhauset al., 1993). Furthermore, the brainstem
spinal cord preparation is sensitive to changes in pH and oxygenation and responds in
a similar manner at the neuronal and motor output levels, as do animalsin vivo
(Ramirezet al., 1998b). The preparation is thus used for studying cellular mechanisms
and the localisation of central chemosensitivity as well as the biphasic respiratory
response to hypoxia (Kawaiet al., 1996; Ramirezet al., 1998b). The preparation is
also suited for detailed pharmacological studies of the respiratory centre in
anaesthesia-free conditions. Drugs can be applied at defined concentrations to the
regions of interest and simultaneous recordings of respiratory output and from
individual neurones made.
Although it has several advantages, thein vitro system is a reduced preparation
with altered parameters of neuronal function including CNS temperature (25-30°C).
Several parameters of neuronal function, including the kinetics of membrane currents
(Hille, 1984) are temperature dependent (Smithet al., 1990). Hypothermia can
attenuate or enhance the action of neuromodulators/drugs affecting respiration (Vitez
et al., 1974; Puiget al., 1987). The temperature therefore has to be considered when
performing experiments using this preparation.The size of thein vitro preparation is a serious limitation since substantial
gradients for oxygen, carbon dioxide and pH may alter the condition of the tissue and
its viability (Mitchell, 1993). Thein vitro preparation is highly reduced by the removal
of afferents and suprapontine inputs. This is desirable in that it excludes several
compensatory mechanisms, i.e., cardiovascular changes, that may complicate data
interpretation. However, this also alters the state of the central pattern generator by
removing tonic inhibitory and excitatory input. Likewise, because of the artificial

Results and Discussion__________________________________________________________________________
25
environment of the neurones, variables such as pH, oxygenation and steady state have
to be taken into consideration when determining effects of drugs.
Thus, data have to be cautiously interpreted and compared with data fromin vivo
models to determine the relationship between the nervous system behaviourin vitro
with that of in vivo systems. We demonstrate below how results depend on time,
temperature as well as perinatal age when performing experiments in this model using
mammals still in a developmental phase.
Time dependency of opioid and adenosinergic drugs
The drug effect has been evaluated after 5 minutes in several of the previous
studies. We demonstrate in the present studies (I-III and V) that steady state effect of
both adenosinergic and opioidergic drugs can take as long as 10-20 minutes to establish.
There are several possible explanations for this:a) Drug kinetics: R-PIA is highly
lipophilic, and thus it is possible that R-PIA binds to fat-rich glia before a gradient and
slow diffusion establishes steady state concentrations within the region of the respiratory
neurones, 50-600µm below the ventral surface;b) Receptor recruitment: recent studies
indicate that some G-protein linked receptors increase at the cell surface when
stimulated. A1-receptors are recruited from the cytoplasm to the surface of neurones after
stimulation (Ciruelaet al., 1997). In addition, it has recently been reported that activation
of dopamine D1-receptor, also G-protein coupled, induces a time- and dose-dependent
recruitment of D1-receptors to the cell surface (Brismaret al., 1998). Thus, an alternative
explanation for the time dependency could be that the full effect of ligands first is
reached when ligand-induced aggregation of receptors to the cell surface is completed.
However, our studies cannot determine the mechanisms for this time dependency, but
nevertheless underline the importance of taking time into account when determining drug
effects in this preparation.
Perinatal age and experiments
The inhibitory effect of morphine on the respiratory rhythmic activity increased
with postnatal age (P0-P4) (III) while the depressant effect of adenosine on respiration
decreased during both foetal (E18-E21) and postnatal life (P0-P4) (I, IV and V). Age
has a significant effect on both control respiratory activity (I and IV) and the effects of

Brainstem perinatal respiratory activity___________________________________________________________________________
26
drugs (I-V) during the first postnatal days. This fact has been neglected in several
studies using thein vitro brainstem spinal cord preparation. (Murakoshi, 1985; Greer
et al., 1995). These findings will be discussed in the context of development of
respiratory control later on.
OPIOIDS AND RESPIRATORY DEPRESSION
Opioids inhibit respiration by µµµµ- including µµµµ-1 and κκκκ-opioid receptors
To study the effects of opioids on brainstem opioid receptors effects of theµ-
receptor agonist DAGO, theδ-receptor agonist DPDPE and theκ-receptor agonist
U50,488 were evaluated in the brainstem spinal cord preparation (II and III). The
opioid receptor antagonists naloxone (µ) and naloxanazine (µ-1) were also used.
DAGO reduced respiratory frequency and inspiratory time in a concentration-
dependent manner and at high concentrations (10µM) caused a reduction of peak
integrated C4 amplitude. Theµ-1 receptor antagonist naloxanazine shifted the
concentration- respiratory frequency response curve for DAGO to the right (p<0.05).
Thus, higher concentrations of DAGO were needed to give an equivalent respiratory
depression as that in animals not pre-treated with theµ-1 antagonist. DPDPE had no
effect on respiratory activities whereas U50,488, like DAGO, reduced respiratory
frequency and amplitude of integrated C4 in a concentration-dependent manner. Thus,
µ-opioid receptors, including theµ-1 receptors are involved in respiratory frequency
reduction whereasκ-opioid receptors are involved in reduction of both respiratory
frequency and respiratory amplitude.δ-opioid receptors do not seem to participate in
respiratory modulation in the neonatal rat.
The finding that aκ-opioid receptor agonist depresses respiration is in conflict
with the findings of Greeret al. (Greeret al., 1995). However, it should be noted that
these authors only conducted their measurements 5 min after drug application (Greer
et al., 1995) while our evaluations were performed after 20 min. This methodological
difference most certainly explains the discrepancy between the studies.
Our results are in agreement with previous findings in adult rats in which
microinjection ofκ-opioid receptor agonists into ventral medullary structures reduced
VT and respiratory frequency (Hassen, 1984). Nevertheless, most of the previousin
vivo studies (Leightonet al., 1987; Yeadon and Kitchen, 1990; Dosaka-Akitaet al.,

Results and Discussion__________________________________________________________________________
27
1993) have indicated thatκ-opioid receptors do not participate in opioid-induced
respiratory depression. This discrepancy between the results of thesein vivo studies
and the presentin vitro study suggest that suprabulbar structures may modify
medullary κ-receptors-mediated respiratory depressant effects. If=κ-receptors are
involved in the respiratory depression of opioids this is important knowledge in the
development of future opioidergic analgesia without respiratory depressanteffects.
Temperature and respiratory control
The temperature normally used in the different mammalianin vitro preparations
is 25-26°C. The low temperature compared toin vivo physiological conditions is used
to decrease oxygen consumption and to sustain regular respiratory activity for long
periods of time. Reduced temperature may have consequences beyond the desired
effects of reduced metabolic rate and prolonging the viability of the preparation. Low
temperatures may alter membrane fluidity, thereby altering membrane bound proteins
including receptors and ion channels. (Mitchell, 1993). In paper III we examined the
possible influences of temperature on respiratory depression induced by opioids. We
demonstrate that lower temperature markedly reduced the ability of morphine (Fig. 6)
and DAGO to decrease respiratory frequency. Changes in temperature did not affect
the dose-response curve forκ-receptor activation by U50,488. We conclude that the
respiratory effect of morphine and DAGO via activation of medullaryµ-opioid
receptors is temperature-dependent.

Brainstem perinatal respiratory activity___________________________________________________________________________
28
Figure 5 Temperature dependent respiratory depression by morphineThe influence of change in temperature on the C4 activity (upper trace)
integrated C4 (lower trace) and their response to morphine 50µM. A) the respiratory
activities recorded from thein vitro preparation in standard solution at 25.5°C. B) 20
minutes after increasing temperature to 28.5°C) 30 min after perfusion with morphine
50µM D) 30 min after decreasing chamber temperature to 25.5°C.

Results and Discussion__________________________________________________________________________
29
The reduced respiratory frequency, caused by morphine in our study, is in
agreement with what has been previously reported(Murakoshi, 1985). Hitherto, not
much has been described about effects of temperature on opioid receptor mechanisms.
It has been proposed that the affinity of theµ-receptor for opioids can be modified at
different temperatures (Garauletet al., 1992). Since receptor proteins can undergo
conformational changes due to temperature changes and, hence, alter their affinities
for the opioid molecule, this could explain our findings. However, the affinity of
naloxone for opioid receptors is not temperature dependent (Puiget al., 1987;
Kuemmerle and Makhlouf, 1992). This concords with previous studies of intestinal
muscle preparations, in which the potency of DAGO (µ-agonist), DPDPE (δ-agonist)
and morphine decreased with a lower temperature while in the range from 37-30°C
(Puiget al., 1987; Garauletet al., 1992).
The opioid receptor is a classic seven transmembrane receptor with three
extracellular loops deciding ligand specificity (Befort, 1997).µ- andδ-agonists require
the third extracellular loop for binding.κ-selectivity lies primarily within the 1st and 2nd
extracellular loop which may gain structural stability from the cysteine-cysteine bridge
connecting these extracellular receptor domains (Traynor, 1996; Befort, 1997). A
speculative explanation of the selectiveµ-receptor temperature dependency could be
that the third extracellular loop, which is not stabilised with a disulphide bridge, is
more susceptible to changes in the temperature. Thus, instability of the third
extracellular loop of the opioid receptor may lead to conformational changes with
temperature and alter the affinity forµ- andδ-agonists.
Our observations of temperature dependent opioid receptor function are of
clinical interest since opioids are frequently used in patients, not least during
hypothermia for cardiopulmonary bypass surgery. In the clinical setting analgesia and
not respiratory depression is the goal. With large doses of opioids administered during
hypothermia, significant side effects will appear at normothermia. Linget al. (Ling et
al., 1985) suggested that opioid-induced analgesia and respiratory depression are
mediated via activation of different subtypes ofµ-opioid receptors. According to their
in vivo study, opioid-induced analgesia was linked toµ1 opioid receptor activation and
respiratory depression was linked toµ2. However, using this brainstem spinal cord

Brainstem perinatal respiratory activity___________________________________________________________________________
30
preparation we demonstrate that the medullaryµ1 opioid receptors also participate in
respiratory frequency reduction (III).
This observation links analgesia with respiratory depression. Clinically it is well
known that the respiratory depressant effect of opioids does not appear until significant
pain relief is achieved. Such a clinical effect is straightforward and logical if the two
modalities, analgesia and respiratory depression, are mediated via the same receptor.
Our results are not necessarily in disagreement with the observation of Linget al., who
reported thatµ2-receptors have an impact on respiratory timing. However, it challenges
the assumption that pain relief viaµ-receptors can, one way or another, be separated
from respiratory depression (Linget al., 1985).
Do opioids and adenosine interact in respiratory depression?
Some of the pharmacological effects of morphine seem to be due to the
presynaptic modulation of transmitter release of substances such as acetylcholine,
substance P and adenosine (Jessell and Inversen, 1977; Sawynoket al., 1989; Taguchi
et al., 1993). In the spinal cord a component of the antinociceptive action of morphine
is due to the release of adenosine (Sollevi, 1997; Sawynok, 1998). Thus to test the
hypothesis that part of the effect of morphine on respiration is mediated by indirect
effect of other transmitters / modulators, we tried to reverse the morphine-induced
respiratory depression by adding adenosine antagonists, substance P and acetylcholine.
Acetylcholine (10µM) and substance P (50 nM) could partly, but not fully, reverse the
morphine-induced reduction of respiratory frequency via respiration-related structures
in the rostral medulla (Takita, Herleniuset al. unpublished observations). However,
the adenosine receptor antagonist theophylline (100µM) does not affect morphine-
induced respiratory depression at all. This would suggest that acetylcholine, substance
P and adenosine are not crucially involved in morphine-induced respiratory
depression.
ADENOSINE AND RESPIRATORY DEPRESSION
Adenosine levels increase during hypoxia and have been proposed to be involved
in hypoxia-induced depression of breathing due to its inhibitory action on the central

Results and Discussion__________________________________________________________________________
31
nervous system. The following results describe where and how adenosine can
modulate respiration through respiration-related neurones in the brainstem.
Adenosine modulates respiratory activity by acting directly on brainstemrespiration-related neuronal networks
In paper I we demonstrate that R-PIA and dipyridamole decrease the activity of
inspiratory neurones (I-neurones) and the C4 respiratory burst rate. Theophylline or
DPCPX reverse the effects of both R-PIA and dipyridamole on C4 respiratory rate and
intra-burst fq of I-neurones. Thus, adenosine depresses both the I-neurones in the RVL
and the respiratory motor output. This depression of I-neurones and respiratory rate
can be abolished by theophylline primarily through a blockade of medullary adenosine
A1 receptors. These results were confirmed and also shown to be valid in the foetal
period in papers IV and V.
The studies presented in this thesis, demonstrate that endogenous adenosine
modulates respiration by acting directly on the neuronal network for breathing
rhythmogenesis (I, IV and V). This does not exclude the possibility that adenosine also
may inhibit respiration via an indirect effect on body temperature and metabolism
(Lagercrantzet al., 1986). However, as indicated in a recent study, the decrease in
metabolism that occurs during hypoxia does not seem to be responsible for the
decrease of respiration induced by hypoxia (Rehanet al., 1996). Our findings confirm
and verify previous suggestions that adenosine modulates respiration through an action
on medullary respiratory networks (Eldridgeet al., 1985; Thomaset al., 1994).
Theophylline and caffeine are adenosine antagonists that cause ventilation to
increase when given systemically (Aranda and Turmen, 1979; Eldridgeet al., 1983).
Furthermore, theophylline can attenuate both the hypoxia-induced depression of
respiration in vivo (Darnall, 1985; Runoldet al., 1989) and the depression of C4
motoneuronal dischargein vitro (Kawai et al., 1995). This indicates that part of the
respiratory decrease induced by hypoxia (Cross and Warner, 1951; Haddad and
Mellins, 1984) can be explained through increased adenosine levels and A1-R
mediated inhibition of respiration-related neurones in the RVL. This finding is
important since two of the most widely used drugs to treat apnoea of prematurity are
theophylline and caffeine (Aranda and Turmen, 1979). This also implies that the
decreased incidence of neonatal apnoeas during theophylline treatment is mainly due
to the antagonistic effect of theophylline on A1-receptors in the medulla oblongata.

Brainstem perinatal respiratory activity___________________________________________________________________________
32
Adenosine modulates the synaptic activity in brainstem respiratory neurones
What are the mechanism(s) behind adenosinergic depression of respiration? After
having found that adenosine acts on respiratory neurones in the rostral ventrolateral
medulla to depress respiratory output we wanted to clarify the mechanisms behind this
depression. To be able to determine synaptic events and changes in membrane
properties of individual neurones intracellular recordings were necessary. To achieve
this goal, the modified blind whole cell patch-clamp technique was used on respiratory
neurones identified by their characteristic firing patterns and correlated in time to the
respiratory cycle of C4/C5-activity. This allowed us to investigate adenosinergic
effects on membrane potential and resistance of RVL neurones, simultaneously
recording changes in C4 respiratory output. Lucifer Yellow labelled neurones were
located 50-500µm below the closest ventral surface and ventral of the ambiguous
nucleus, in agreement with previous findings (Arataet al., 1990). Camera Lucida
reconstruction of the major respiratory neurone subtypes found in vitro are illustrated
in Fig. 6.
Figure 6. Camera Lucida reconstruction of respiratory neurones: biphasic expiratory (Pre-
I) and expiratory neurone in proximity of an inspiratory neurone.
In paper V, we demonstrate that R-PIA consistently causes a reduction of
spontaneous synaptic activities during the expiratory phase in all RVL neurones
examined. Figure 7 (modified from paper V) depicts the characteristics of major

Results and Discussion__________________________________________________________________________
33
groups of respiration-related neurones found. The A1-R agonist and antagonist induced
modulation of excitatory postsynaptic potentials did not correlate with changes in
membrane potential (Em) or input resistance (Rm). This indicates that adenosine
depresses respiration through presynaptic adenosine A1-receptor mediated inhibition of
synaptic transmission in the medulla oblongata. This is in agreement with the now
well established role of adenosine as a regulator of CNS synaptic transmission
(Snyder, 1985; Brundege and Dunwiddie, 1997; Dunwiddie and Fredholm, 1997).
Several possible mechanisms exist for by which presynaptic A1-receptor can decrease
transmitter release (Brundege and Dunwiddie, 1997). One of these is mediated via a
G-Protein dependent inhibition of N-type Ca++ channels (Mynlieff and Beam, 1994).
This will decrease Ca++-induced transmitter release. Other mechanisms not involving
Ca++ entry from the extracellular space exist (Brundege and Dunwiddie, 1997).
However, it has not been within the scope of the present studies to determine the exact
mechanisms following presynaptic A1-R activation. Nevertheless, we indicate here that
the decreased synaptic activity by adenosine observed in other parts of the brain
(Brundege and Dunwiddie, 1997) is also valid for the medullary central pattern
generator for breathing.

Brainstem perinatal respiratory activity___________________________________________________________________________
34
Figure 7 Respiration related neurones in the neonatal rat brainstemInspiratory (Insp) neurones receive excitatory synaptic input (EPSPs) and
discharge action potentials during inspiratory C4 ventral root activity (C4). Insp-I
neurones receive EPSPs prior to and after termination of C4 activity whereas Insp-
III neurones are inhibited during both these phases. Biphasic expiratory (Biphasic
E) are characterised by pre- and postinspiratory excitation and inspiratory related
inhibition. Expiratory (Exp) neurones are inhibited by hyperpolarising IPSPs
during the inspiratory phase and either discharge tonically in the expiratory phase
(Exp-i) or receive continued inhibition and discharge primarily in the late
expiratory phase.

Results and Discussion__________________________________________________________________________
35
Figure 8 Irregular C4 respiratory activity (bottom) and discharges of a biphasic
expiratory neurone, discharging before and after C4 and hyperpolarised during C4
inspiratory activity, preparation from new-born pup (2 hours after birth). Note the
regular distance between neuronal discharge burst and how the long postinspiratory
discharges

Brainstem perinatal respiratory activity___________________________________________________________________________
36
Adenosine and postsynaptic depression of respiration-related neurones
Except for reduced synaptic noise, evident in all neurones, the effects of
adenosinergic agents were different between the major groups of respiration-related
neurones. Expiratory neurones demonstrated a reversible decrease in input resistance,
a depression of action potential discharges and a hyperpolarisation of the membrane
potential during application of R-PIA. Similar responses of input resistance andEm to
R-PIA were apperent after synaptic activity had been blocked by 0.5µM tetrodotoxin
(TTX). As a group, biphasic expiratory (biphasic E) and inspiratory neurones
demonstrated no changes in input resistance or membrane potential during R-PIA
application with our without TTX present. However, whereas the response to
adenosinergic agents was uniform among inspiratory neurones, the response of
biphasic E neurones varied. In some biphasic E neurones the synchronisation between
neurone burst discharges and C4/C5 activity was reversibly disturbed by R-PIA. R-
PIA induced a reversible hyperpolarisation in a third of Biphasic E neurones
examined. Some of these neurones are characterised by a high level of excitatory
(subtreshold) synaptic input and a reduction of ongoing excitatory postsynaptic
potentials could lead to an apparent hyperpolarisation of the membrane potential. The
fact that only one out of 11 Biphasic E neurones demonstrated a change in input
resistance supports this hypothesis. The effects of R-PIA on respiratory activity, input
resistance and membrane potential could be reversed by the A1-receptor antagonist
DPCPX (200 nM).
Our data suggest that the modulation of respiratory output induced by
adenosinergic agents can be explained by: 1) a general decrease in synaptic
transmission between medullary respiration-related neurones mediated by presynaptic
A1-receptors; and 2) an inactivation, via membrane hyperpolarisation, of medullary
expiratory neurones mediated by postsynaptic A1-receptors.
Expiratory neurones are hyperpolarised and inactivated through postsynaptic A1-
receptors. This inactivation of expiratory neurones is similar to what has been
observed in vivo with intracellularly injected adenosine inducing a postsynaptic
membrane hyperpolarisation and inactivation of expiratory neurones (Schmidtet al.,
1995). Alsoin vitro, during hypoxic conditions, the expiratory neurones are reversibly

Results and Discussion__________________________________________________________________________
37
inactivated as demonstrated in the isolated neonatal mice respiratory network (Ramirez
et al., 1998b). Thus, it is plausible that ourin vitro findings could partially explainin
vivo response to hypoxia. Adenosine has been proposed as a link between cellular
energy metabolism and the excitability of neurones (Lloydet al., 1993), since
breakdown of ATP increases substantially during hypoxia and more adenosine is
formed. Adenosine is thus considered as a retaliatory metabolite by which cells
communicate their energy status to surrounding tissues (Newby, 1984).
Inhibition of expiratory neurones does not abolish the respiratory rhythm
As stated above, adenosine causes: 1) a general decrease in synaptic transmission
between medullary respiration-related neurones mediated by presynaptic A1-receptors;
and 2) an inactivation, via membrane hyperpolarisation, of medullary expiratory
neurones mediated by postsynaptic A1-receptors. Inactivation of tonic and late
expiratory neurones only modulates but does not abolish the generation of respiratory
rhythm in vitro. These findings suggests that these neuronesper seare not necessary
for respiratory rhythm generation. This is consistent with the view that respiratory
rhythm is generated primarily by inspiratory-related neurones (Bradleyet al., 1975;
Wyman, 1977; Richter, 1982) and that medullary expiratory neurones are less
important for the rhythmogenesis of respiration. It is contradictory to network models
for the generation of respiratory rhythm in adult mammals, which have phasic and
reciprocal inhibitory interactions between inspiratory and expiratory neurones as a key
feature (Ezure, 1990; Bianchiet al., 1995). However, the respiratory generation
matures during the first two postnatal weeks (Paton and Richter, 1995) and the
neurones in the reduced neonatalin vitro preparation may respond differently than in
adult in vivo systems. Nevertheless, as demonstrated by Richteret al. (Richter et al.,
1993), most expiratory type 2 neurones in the adult cat are inactivated by membrane
hyperpolarisation and stop discharging actionpotentials during hypoxia. As adenosine
levels increase during hypoxia (Winnet al., 1981) a possible explanation would be
that expiratory neuronesin vivo as well asin vitro are not essential for respiratory
rhythm and can be inactivated by hypoxia-induced increased adenosine levels.

Brainstem perinatal respiratory activity___________________________________________________________________________
38
K+
AdenosineBiphasic
E
Insp
Insp
Type II
Type III
PPPEEERRRIII ---III
Expl
Spinal cord
BiphasicE
InspType I
excitatory
inhibitory
Expt
EEEXXXPPP
IIINNNSSSPPP
Adenosine++Ca
1.
2.
Figure 9 Proposed model for the actions of adenosine A1-receptors (A1-R) in the
respiratory network of neonatal rats. Respiratory neurones are categorised into three
subgroups: inspiratory (INSP), peri-inspiratory (PERI-I) and expiratory (EXP).
1. Presynaptic A1-R decrease neurotransmitter release through inhibiting Ca++-channels
(N-type) (Mynlieff and Beam, 1994).2. Postsynaptic A1-R hyperpolarises and
inactivates expiratory neurones via a decreased K+-channel conductance (Thompsonet
al., 1992). This decreases the overall synaptic transmission in the network without
stopping the respiratory rhythm generation. Identified components of the CPG for
respiration in neonatal ratsin vitro and their connectivity. (Inspiratory neurone (Insp),
late expiratory neurone (Expl), tonic expiratory neurone (Expt), biphasic expiratory
(Biphasic E). The Biphasic E neurone is also characterised as Pre-inspiratory neurone
(Onimaruet al., 1987). (Modified, with kind permission of the authors, after (Onimaru
et al., 1997).

Results and Discussion__________________________________________________________________________
39
DEVELOPMENT OF RESPIRATORY CONTROL
The neuronal network generating respiratory rhythm does not undergo majorchanges in the early perinatal period.
In paper IV we show that an irregular rhythmic respiratory activity is
established in the foetal rat at embryonic day 18. The regularity of respiratory activity,
measured as a decrease in the coefficient of variation (CV) of the interval between C4
burst discharges, increased with postnatal age (P<0.01). Also time of inspiratory
activity in each C4 burst (Ti) increased with postnatal age (ANOVA P<0.05). On E21
a respiratory pattern resembling that after birth had been established with regard to
pontine inhibition, control frequency and Ti. Mean frequency of respiratory activity
did not differ between age groups when pons remained, but was significantly slower
on E18 after ponto-medullary transection. Removal of pons resulted in a significant
increase in frequency (from 2.3+1.4 to 9.1+3.1 burst/min, P<0.0001). The central
pattern generator for respiratory rhythm in medulla oblongata and its inhibition from
pontine structures is thus active on E18. Respiratory rhythmic activity in medulla
oblongata matures pre- and postnatally with regard to both frequency and regularity of
respiratory activity. Pontine inhibition of this respiratory rhythm do not seem to
change from E21 to P3. Our results confirm previous results concerning the
development of respiratory rhythm generation in the foetal and neonatal rat (Di
Pasqualeet al., 1992; Greer , 1992). Previously the inhibition by pons on respiratory
activity in the medulla oblongata has been demonstrated to exist on E20 (Di Pasquale
et al., 1992) but we find it already on E18.
In addition, the depressant effects of an adenosine A1-receptor agonist decrease
between E18 to P3. This parallels the increased regularity of respiratory rhythm and a
possibility could be that a weaker adenosinergic influence on respiration increase its
regularity. However, the increased maturation of dendrites and synaptic connections in
the developing neuronal networks is a more plausible explanation (Bryan and Bryan,
1978). Thus, the decreased role of adenosine in modulating respiration could either be
due to changes in receptor affinity with development (Runoldet al., 1986), or that
more mature neuronal network are less susceptible to the influences of an inhibitory
modulator such as adenosine.

Brainstem perinatal respiratory activity___________________________________________________________________________
40
The development of respiratory activity and chronic maternal intake of caffeine.
The caffeine treated group differed significantly from control in their basal
respiratory frequency, which was higher than control. Furthermore, the pontine
inhibition of brainstem generated C4 activity was more pronounced in the caffeine
group (MANOVA, repeated measures design; P<0.05) (Paper IV Fig. 2C).
Furthermore, chronic exposure to a low dose of caffeine increased the pontine
inhibition of breathing activity and changed the response to an anoxic challenge
without detectable changes in A1-receptor ontogeny. Furthermore, this pontine
inhibition is dependent on age and is influenced by chronic caffeine intake.
The in vitro findings are strengthened by thein vivo findings of a reduced
gasping frequency during anoxia in the P7 caffeine group. Pons is involved in the
respiratory depression during hypoxia (Okadaet al., 1998). Thus, it is possible that
this reduced gasping frequency reflects an increased hypoxic respiratory depression
mediated by structures in the pons (Okadaet al., 1998).
Even if the levels of adenosine A1-receptors increase with age and high levels are
encountered in pons-medulla in postnatal life (Fig 8) the effects of adenosine on
respiration decrease. It is difficult to quantify the effects of adenosine. The modulation
of synaptic transmission and neuronal membrane properties involve a complex cascade
of mechanisms mediated by several neuromodulators (Lagercrantz, 1987; Feldman,
1990). Changes may be direct by A receptors or indirect effects on other transmitter
systems. Our data indicate that receptor expression is not equivalent to functional
importance. This may be due to that the expression of receptors with increasing
postnatal age are relatively lower in areas and neurones involved in respiratory
regulation compared to surrounding brainstem regions. Post-translatory changes or
changes in network properties may also be involved.
Fig. 10A1-mRNA expression
In a P21 rat

Results and Discussion__________________________________________________________________________
41
GENERAL DISCUSSION
Before birth the isolated respiration-related neuronal networks in the brainstem
generate a similar output as that during the first postnatal days (IV). Thus, changing
respiratory activity at birth from intermittent to continuous breathing does not seem to
depend on major changes in the network.
Birth brings the foetus into contact with a totally different environment than that
in utero. New tactile stimuli, light, lower temperature, removal of the umbilical
circulation, arousal, gas with oxygen and carbon dioxide in the upper and lower
airways, increased pulmonary blood flow, increased oxygen consumption, vagal input
from mechanoreceptors are all involved in the initiation and maintenance of breathing
(Blanco, 1991). The decreased inhibition from neuromodulators such as adenosine and
prostaglandin and increased excitatory afferent input after birth has given rise to the
expression “the inhibited state of the foetus”.
As is apparent from the present studies, the modulatory effects of adenosine on
central respiratory rhythm generation decrease within the first two postnatal days (I,
IV and V). A decreased sensitivity to adenosinergic agents in the neonatal rat after
birth could explain our observations. This concords with earlier findings in neonatal
rabbits (Runoldet al., 1986) and piglets (Elnaziret al., 1996), in which a considerably
more pronounced effect of adenosine agonists on respiration was evident in newborns
(1-3 days) compared with older animals (8 days and 3 weeks). Furthermore, the
present studies indicate that the decreasing sensitivity for adenosine may contribute to
the maintenance of continuous breathing during the first postnatal days. This can be
concluded based on the facts that: 1) adenosine levels ought to decrease as partial
pressure of oxygen in arterial blood rapidly increases after birth; 2), as presented in our
studies (I, IV and V), the effect of A1-R activation on central respiratory activity
decreases during the first two postnatal days.
The instability of breathing in the new-born period has been attributed to the
immaturity of neuronal networks (Bryan and Bryan, 1978). Thus, according to this
view the increased regularity of the respiratory rhythm can be partly explained by the
increased organisation of central neurones as well as the afferent input. The increased
regularity of control respiratory rhythm activities during the perinatal period
investigated in our studies (E18-P7) may be explained by a maturation of the central
neuronal networks. This maturation seems in part to be independent of birth since the

Brainstem perinatal respiratory activity___________________________________________________________________________
42
increased regularity of central respiratory output in postnatal life has already started in
foetal life (IV). The CV
During pre- and postnatal brain development the brain becomes bigger because
neurones grow in size, and the number of axons and dendrites as well as the extent of
their connections increases (Shatz, 1992). To achieve the precision of the adult pattern,
neuronal function is necessary.
There is a general requirement for neuronal activity to transform immature
neuronal circuits into the organised connections that subserve adequate brain function
(Katz and Shatz., 1996). A key function for neurones is to communicate with other
neurones. “Neurones that fire together wire together while the others are left in the
sink”. Changes in the tonic adenosinergic modulation of synaptic transmission by
caffeine may interfere in the normal development of neuronal circuits. Rat pups whose
mothers drank caffeine had a slightly changed development of respiratory activity and
its control. A speculative explanation to these findings may thus be that the changes
are not due to changes in receptor development but in the development of neuronal
networks.
It may be of functional significance that vital brainstem neurones and their
resulting respiratory outputs remain active, while overall synaptic transmission in the
network is decreased. This will attenuate local neuronal damage induced by hypoxia
and at the same time increase the time for respiration to possibly restore oxygen
saturation to normal levels. Overall, adenosine decreases oxygen consumption and has
neuroprotective effects (Fredholm, 1997) without itself stopping respiratory rhythm
generation. The extensive use of xanthines may thus have unwanted side-effects and
should, may be first be used as a secondary choice of treatment when equivalent
alternatives exists.

Results and Discussion__________________________________________________________________________
43
CONCLUSIONS
• The central respiratory rhythm generation does not undergo major changes in the
perinatal period around birth.
• Adenosine and opioids depress respiration by acting directly on the neuronal
networks in Medulla Oblongata generating respiratory activity. Opioids depress
respiration throughµ- and κ-opioid receptors in the ventral Medulla Oblongata.
The potency ofµ-, but notκ-opioid receptors to depress respiration is temperature
dependent and increases between 22.5-30.5oC.
• The modulatory effect of adenosine and opioids on respiration changes during the
first postnatal days. Whereas adenosinergic depression of respiratory activity is
inversely correlated to perinatal age, the effects of morphine increases with
postnatal age.
• Chronic maternal intake of caffeine during gestation does not affect A1-receptor
development but the development of central respiratory activity and its modulation
by pontine structuresin vitro is altered. Most likely this applies alsoin vivo since
caffeine treated pups, at a postnatal age of 7 days, have a reduced anoxia-induced-
gasping. This indicates that changes in pontine inhibition of respiratory neurones
may have long-term functional consequences. This raises new questions of the
extensive use of caffeine during pregnancy and in treatment of neonates.
• Adenosine depresses respiration through presynaptic adenosine A1-receptor-
mediated inhibition of synaptic transmission in the medulla oblongata.
Furthermore, postsynaptic A1-receptors hyperpolarises and inactivates tonic and
late expiratory neurones but only modulates the generation of respiratory rhythmin
vitro, thus indicating that these neuronesper seare not necessary for respiratory
rhythm generation.

Brainstem perinatal respiratory activity___________________________________________________________________________
44
ACKNOWLEDGEMENTS
Professor Hugo Lagercrantz, for introducing me to the fascinating field of
developmental physiology and neuroscience, for his great enthusiasm for research and
new ideas, providing laboratory facilities, help with manuscripts and giving me the
possibilities to develop as an independent researcher.
Yuji Yamamoto for introducing me to the isolated respiratory network and Japan
and for his knowledge in neurophysiology.
Professor Sten Lindahl, for never-ending support, advice, bananas and encouragement
as well as crucial help with making some manuscripts readable.
Professor Hiroshi Onimaru, Ikuo Homma and members at their laboratory for giving
me the ability to come to Tokyo and Showa university and hereafter for a welcome
supervision and warmth which made my stay both valuable and memorable.
Koichi Takita, co-author and collaborator for long nights at the laboratory during
experiments and analysis. The smiles and ironic remarks we share
Lie-Qi Tang, Ulrika Åden coauthors and collaborators
Shi-Zeng Yuan and Ronny Wikström for help or dart when needed
Professor Bertil Fredholm for valuable discussions
Dr. Mikael Runold for your helpful advice on life and adenosine, aid with the barometric
method, constructive reading of this thesis and your reflections of life and being such a nice
guy
Professor John Wahren for constructive reading of this thesis.
Jurij Shvarev, Henning Johanssen, Shinhiro T for help in unravelling the mysterious
Torbjörn Hertzberg, Josef Milerad and Hans Holgert for valuable advice
Thomas Ringstedt, Per Uhlen and Ingrid Dahlin for knowing what to do
All colleagues working at the research and neonatal department at Astrid Lindgren
Children’s Hospital

Results and Discussion__________________________________________________________________________
45
Anna Hårdemark for your curiosity, taste for fantasy and bringing life into the
laboratory
Professor Curt von Euler for starting it all
Charin Carlsson and Viveca Karlsson for expert secretarial help, smiles, laughter and
keeping tracks of foundations, grants etc˜.
Professor Sten Grillner and the members at the Department of Neuroscience and
Neurophysiology, for introducing me to the fascinating field of neuroscience and
neuronal networks.
All the staff at the Spinalis unit and members of the Swedish and International
Foundation for Disabled Sailing for giving me the possibility to enjoy sailing and
working in a exciting clinical and sport environment.
Colleagues and friends made during my years in the Doktorandföreningen
Britta and Patrik for being full of laughter, curiosity and being ultimate parents in law
Mom and Dad for bringing me here and opening the world, Nils, Anne and Carl for
help and joys in it.
Joakim, Per, Martin, Niklas for never speaking science
Vivianne, Gittan, Emilie, Christina for always speaking about life
And all of you out there..
Marie the love of my life
Saga Rebecka for being such a wonder
The present study was generously supported by:Swedish Medical Research Council (SMFR 19X-5234, B96-04R-11693 and 14X-
0907), the Swedish Society for Medical Research, Samariten, the Fraenkel and Laerdal
Fund, Wera-Ekstráms Fund for Pediatric Research, Erik and Edith Fernstrám Fund for
Medical Research, Erik Callergrens Memory Foundation, Mia och Erik Myrins
Memory Foundation, Golja and Anna Cederbergs Foundation and Karolinska Institutet
“NO BREATHE, NO LIVE” – The KARATE KID II

Brainstem perinatal respiratory activity___________________________________________________________________________
46
REFERENCES
Aranda JV and Turmen T (1979), Methylxanthines in apnea of prematurity.Clin Perinatology,
6, 87-108.
Arata A, Onimaru H and Homma I (1990), Respiration-related neurons in the ventral medulla of
newborn rats in vitro.Brain Res Bull, 24, 599-604.
Ball KT, Gunn TR, Gluckmann PD and Power GG (1996), Suppressive action of endogenous
adenosine on ovine fetal nonshivering thermogenesis.Journal of Applied Physiology, 81,2393-2398.
Befort K and Kieffer BL (1997), Structure-activity relationships in the delta opioid receptor.
Pain Reviews, 4, 100-121.
Berne RM (1986), Adenosine: an important physiological regulator.Trends Neuroscience, 1,163-167.
Bianchi AL, Denavit-Saubié M and Champagnat (1995), Central control of breathing in
mammals: neuronal circuitry, membrane properties, and neurotransmitters.Physiological
Rev, 75, 1-45.
Bissonette JM, Hohimer R, Chao CR, Knopp SJ and Notaroberto NF (1990), Theophylline
stimulates fetal breathing movements during hypoxia.Pediatr Res, 28, 83-86.
Blanco C (1991) Role of the brainstem in the changes at birth; initiation of continous breathing
and its maintenance. In:The Fetal and Neonatal Brainstem; developmental and clinical
issues. Edited by M. A. Hanson. Cambridge: University Press, Cambridge, pp. 106-126.
Blanton MG, Lo Turco JJ and Kriegstein, AJ(1989), Whole-cell recordings from neurons in
slices of reptilian and mammalian cerebral cortex.J. Neurosci. Methods, 30, 203-210.
Boddy K, Dawes GS, Fisher RL, Pinter S and Robinson, JS (1974), Foetal respiratory
movements, electrocortical and cardiovascular responses to hypoxaemia.J. Physiol., 243,599-618.
Borday V, Fortin, G and Champagnat J (1997), Early Ontogeny Of Rhythm Generation and
Control Of Breathing.Respiration Physiology, 110,245-249.
Bradley GW, von Euler C, Martilla I and Roos B (1975), A model of the central and reflex
inhibition of inspiration in the cat.Biol. Cybernetics, 19, 105-116.
Brismar H, Asghar M, Carey RM Greengard P and Aperia A (1998), Dopamine-induced
recruitment of dopamine D1 receptors to the plasma membrane.PNAS, 95, 5573-5578.
Brockhaus J, Ballanyi K, Smith JC and Richter DW (1993), Microenvironment of respiratory
neurons in the in vitro brainstem-spinal cord of neonatal rats.J Physiol, 462,421-445.
Brundege JM and Dunwiddie TV (1997), Role of adenosine as a modulator of synaptic activity
in the central nervous system.Advances in Pharmacology, 39, 353-91.
Bryan AC and Bryan MH (1978), Control of respiration in the newborn.Clin Perinatology, 5,269-281.

Results and Discussion__________________________________________________________________________
47
Chernick V and Craig RJ (1982), Naloxone reverses neonatal depression caused by fetal
asphyxia.Science, 216,1252-1253.
Ciruela F, Saura C, Canela EI, Mallol J, Lluis C and Franco R (1997), Ligand-induced
phosphorylation, clustering, and desensitization of A1 adenosine receptors.Molecular
Pharmacology, 52, 788-97.
Cross K and Warner P (1951), The effect of inhalation of high and low oxygen concentration on
the respiration of the newborn infant.J Physiol, 114,283-295.
Darnall RJ (1985), Aminophylline reduces hypoxic ventilatory depression: possible role of
adenosine.Pediatr Res, 19, 706-710.
de Vries JIP, Visser GHA and Prechtl HFR (1982), The emergence of behaviour I. Qualitative
aspects.Early Hum. Dev., 7, 301-322.
Devoe LD, Murray C, Youssif A and Arnaud M (1993), Maternal caffeine consumption and
fetal behavior in normal third-trimester pregnancy.American Journal of Obstetrics &
Gynecology, 168,1105-11.
Di Pasquale E, Monteau R and Hilaire G (1992), In vitro study of central respiratory-like
activity in the fetal rat.Exp Brain res, 89, 459-464.
Dosaka-Akita K Tortella, FC, Holaday JW and Long JB (1993), The kappa opioid agonist U-
50,488H antagonizes respiratory effects of mu opioid receptor agonists in conscious rats.J
Pharmacol Exp Ther, 264,631-637.
Drorbaugh JE and Fenn WD (1955), A barometric method for measuring ventilation in newborn
infants.J Pediatrics,16, 81-87
Drury AN and Szent-Gyrörgyi A (1929), The physiological activity of adenine compounds with
especial reference to their action upon the mammalian heart.J. Physiol, 68, 213-237.
Dunwiddie TV and Fredholm BB (1997), Adenosine regulation of neurotransmitter release. ,kk,kk.
Eldridge FL (1981), Input-output relationships of central neural circuits involved in respiration
in cats.Journal of Physiology, 311,81-95.
Eldridge FL, Millhorn DE and Kiley, JP (1985), Antagonism by theophylline of respiratory
inhibition induced by adenosine.Journal of Applied Physiology, 59 (5),1428-1433.
Eldridge FL, Millhorn DE, Waldrop, TG and Kiley, JP (1983), Mechanism of respiratory effects
of methylxanthines.Respiration Physiology, 53, 239-61.
Elnazir B, Marshall JM and Kumar P (1996), Postnatal development of the pattern of respiratory
and cardiovascular response to systemic hypoxia in the piglet: the roles of adenosine.J
Physiol, 492,573-585.
England SJ (1993) Central effects of hypoxia in the neonate. In:Respiratory control-Central and
Peripheral mechanisms. Edited by Speck DF, Dekin MS, Revelette WR and Frazier . T.
Lexington: The University press of Kentucky, pp. 163-166.
Euler C and Lagercrantz H (1987),Neurobiology of the control of breathing: Raven Press.

Brainstem perinatal respiratory activity___________________________________________________________________________
48
Ezure K (1990) Synaptic connections between respiratory neurons and considerations on the
genesis of respiratory rhtythm. In:Progress in Neurobiology, Volume 35, pp. 429-450.
Feldman JL (1987) Emergent properties of neuronal mechanisms controling breathing in
mammals. In:Respiratory muscles and their neuromotor control: Alan R. Liss Inc., pp. 3-8.
Feldman JL, Smith JC and Liu G (1991), Respiratory pattern generation in mammals: in vitro en
bloc analyses.Curr Opin Neurobiol, 1, 590-4.
Feldman JL, Smith JC, Ellenberger HH Connelly CA, Liu G, Greer, JJ, Lindsay AD and Otto
MR (1990), Neurogenesis of respiratory rhythm and pattern: emerging concepts.American
Journal of Physiology, 259,879-886.
Flórez J and Hurlé MA (1993) Opioids in respiration and vomiting. In:Hand book of
Experimental pharmacology, Opioids II. Edited by H. A. New York: Springer-Verlag, pp.
263-292.
Fredholm BB (1995), Purinoreceptors in the nervous system.Pharmacology & Toxicology, 76,228-239.
Fredholm BB (1997), Adenosine and neuroprotection. [Review] [130 refs].International Review
of Neurobiology, 40, 259-80.
Garaulet JV, Laorden ML and Milanes MV(1992), Influence of temperature on the effects ofµ-,
δ- andκ-opioid receptor agonists in the guinea-pig ileum myenteric plexus.Eur. J. Pharmac.,
223,19-23.
Greer JJ, Carter JJ and AL-Zubaidy Z (1995), Opioid depression of respiration in neonatal rats.
Journal of Physiology, 485,845-855.
Greer JJ, Smith JC and Feldman JL (1992), Respiratory and locomotor patterns generated in the
fetal rat brain stem-spinal cord in vitro.Journal of Neurophysiology, 67, 996-9.
Grunstein MM, Hazinski, TA and Schlueter MA (1981), Respiratory control during hypoxia in
newborn rabbits: Implied action of endorphins.Journal of Applied Physiology: Respiratory,
Environmental & Exercise Physiology, 51, 122-130.
Guillet R (1990), Neonatal caffeine exposure alters adenosine receptor control of locomotor
activity in the developing rat.Dev. pharmmacol. ther, 15, 94-100.
Guillet R and Dunham L (1995), Neonatal caffeine exposure and seizure susceptibility in adult
rats.Epilepsia, 36, 743-9.
Guntheroth WG and Kawabori I (1975), Hypoxic apnea and gasping.Journal of Clinical
Investigation, 56, 1371-7.
Haddad GG and Mellins RB (1984), Hypoxia and respiratory control in early life. [Review].Ann
Rev Physiol, 46, 629-643.
Hanson M (1996) Neonatal, perinatal and developmental aspects of respiratory control. In:
Pediatrics & Perinatology: The scientific basis. Edited by P. D. Gluckman and M. A.
Heymann. London: Arnold, pp. 836-844.
Hassen AH, Feuerstein G and Faden AI (1984), Kappa opioid receptors modulate
cardiorespiratory function in hindbrain nuclei of rat.J. Neurosci., 4, 2213-2221.

Results and Discussion__________________________________________________________________________
49
Hedner J, Hedner T, Wessberg P, Jonason J and Mueller RA (1985), Central respiratory effects
of adenosine analogues, theophylline and enprofylline. In:Anti-asthma xanthines and
adenosine. Edited by K. E. Andersson and C. G. A. Persson. Copenhagen: Excerpta medica,
pp. 467-471.
Hedner T, Hedner J, Jonason and Wessberg P (1984), Effects of theophylline on adenosine-
induced respiratory depression in the preterm rabbit.Eur J Respir Dis, 65, 153-156.
Hille B (1984),Ionic channels of excitable membranes. Sinauer: Sunderland, M.A.
Holloway Jr WR (1982), Caffeine:Effects of acute and chronic exposure on the behaviour of
neonatal rats.Neurobehv. Toxicol. Teratol., 4, 21-32.
Holloway Jr WRandThor DH (1982), *Caffeine sensitivity in the neonatal rat.Neurobehav.
Toxicol. Teratol, 4, 331-333.
Holloway WRJandThor DH (1982), Caffeine sensitivity in the neonatal rat.Neurobehavioral
Toxicology & Teratology, 4, 331-3.
Irestedt L et al. (1989), Adenosine concentration in umbilical cord blood of newborn infants
after vaginal delivery and cesarean section.Pediatric Research, 26, 106-8.
Isom GE and Elshowihy RM (1982), Naloxone-induced enhancement of carbon dioxide
stimulated respiration,.Life Sci.., 31, 113-118.
Jessell TM and Inversen LL (1977), Opiate analgesics inhibit substance P release from rat
trigeminal nucleus.Nature, 268,549-551.
Johnston BM (1991) Brainstem inhibitory mechanisms in the control of fetal breathing
movements. In:The fetal and neonatal brainstem. Edited by M. Hanson. Cambridge:
Cambridge Uiversity Press, pp. 21-47.
Katz LC and Shatz, JC (1996), Synaptic activity and the construction of cortical circuits.
[Review] [86 refs].Science, 274,1133-8.
Kawai A, Ballantyne D, Muckenhoff K and Scheid P (1996), Chemosensitive medullary
neurones in the brainstem--spinal cord preparation of the neonatal rat.Journal of Physiology,
492,277-292.
Kawai A Okada Y, Muckenhoff K and Scheid P (1995), Theophylline and hypoxic ventilatory
response in the rat isolated brainstem-spinal cord.Resp Physiol, 100,25-32.
Kitterman JA (1996), The effects of mechanical forces on fetal lung growth.Clinics in
Perinatology, 23, 727-40.
Koos BJ and Matsuda K (1990), Fetal breathing, sleep state, and cardiovascular responses to
adenosine in sheep.J Appl Physiol, 68, 489-495.
Kuemmerle JF and Makhlouf GM (1992), Characterization of opioid receptors in intestinal
muscle cells by selective radioligands and receptor protection.Am. J. Physiol., 263,269-276.
Lagercrantz H (1987), Neuromodulators and respiratory control during development.TINS, 10,368-372.

Brainstem perinatal respiratory activity___________________________________________________________________________
50
Lagercrantz H, Runold M, Yamamoto Y, and Fredholm BB (1986) Adenosine: A putative
mediator of the hypoxic ventilatory response of the neonate. In:Neurobiology of the Control
of Breathing. Edited by H. Euler and H. Lagercrantz. New York: Raven Press, pp. 133-139.
Lagercrantz H, Yamamoto Y, and Fredholm BB, Prabhakar NR and von Euler C (1984),
Adenosine analogues depress ventilation in rabbit neonates. Theophylline stimulation of
respiration via adenosine receptors?Pediatr Res, 18, 387-390.
Lawson EE and Long WA (1983), Central origin of biphasic breathing patterns during hypoxia
in newborns.J. Appl. Physiol., 55, 1299-1304.
Le Gallois (1812),Expériences sur le principe de la vie. Paris: D´Hautel.
Leighton GE, Johnson MA, Meecham KG, Hill RG and Hughes J (1987), Pharmacological
profile of PD117302, a selectiveκ-opioid agonist.Br J Pharmacol, 92, 915-922.
Ling GSF, Simantov R, Clark JA and Pasternak GW (1986), Naloxonazine actions in vivo.Eur.
J. Pharmacol., 129,33-38.
Ling GSF, Spiegel K, Lockhart SH and Pasternak GW (1985), Separation of opioid analgesia
from respiratory depression: evidence for different receptor mechanisms.J. Pharmacol. Exp.
Ther., 232,149-155.
Lloyd HG, Lindström K and Fredholm BB (1993), Intracellular formation and release of
adenosine from rat hippocampal slices evoked by electrical stimulation or energy depletion.
Neurochem Int, 23, 173-185.
Lopes JM, Davis GM, Mullahoo K and Aranda JV (1994), Role of adenosine in the hypoxic
ventilatory response of the newborn piglet.Pediatr Pulmonol, 17, 50-55.
Maloney JE, Adamson TM, Brodecky V, Dowling MH and Ritchie BC (1975), Modification of
respiratory center output in the unanesthetised fetal sheep "in utero"J Appl Physiol,39, 552-
558.
Mansour A, Khachaturian H, Lewis ME Akil H and Watson SJ (1988), Anatomy of CNS opioid
receptors.Trends in Neurosci, 11, 308-314.
Miller MJ and Martin RJ (1992) Pathophysiology of apnea of prematurity. In:Fetal and
Neonatal Physiology, Volume 1. Edited by R. F. Polin, WW. Philadelphia: W.B. Saunders
Company, pp. 872-885.
Mitchell GS (1993) Overview: In vitro studies. In:Respiratory control-Cenral and peripheral
mechanisms. Edited by D.F. Speck, M.S. Dekin, W.R. Revelette and D.T. Frazier. Lexington:
Kentucky University press, pp. 30-33.
Mitchell GS and Scheid P (1997), Special Issue - Neural Control Of Breathing - Molecular to
Organismal Perspectives - Foreword.Respiration Physiology, 110,69-70.
Morin SM, Boudinot E, Fournie, Zaluski MC, Champagnat J Roques BP and Denavit-Saubié M
(1992), Control of breathing by endogenous opioid peptides: possible involvement in sudden
infant death syndrome.Neurochem Int, 20, 103-7.
Morton and Garrison (1991),Morton´s Medical Bibliography, Edited by J. M. Norman: Scolar
Press.

Results and Discussion__________________________________________________________________________
51
Moss IR, Denavit-Saubié M, Eldridge FL, Gillis RA, Herkenham M and Lahiri S (1986),
Neuromodulators and transmitters in respiratory control.Federation Proceedings, 45, 2133-
47.
Moss IR, Runold M, Dahlin I, Fredholm BB, Nyberg F and Lagercrantz H (1987), Respiratory
and neuroendocrine responses of piglets to hypoxia during postnatal development.Acta
Physiolog Scandinav, 131,533-541.
Murakoshi T, Suzue T, Tamai S. (1985), A pharmacological study on respiratory rhythm
generation in the in isolated brainstem-spinal cord preparation of the newborn rat.Br. J.
Pharmacology, 86, 95-104.
Mynlieff M and Beam KG (1994), Adenosine acting at an A1 receptor decreases N-type calcium
current in mouse motoneurons.J Neurosci, 14, 3628-3634.
Nehlig A and Debry G (1994), Potential teratogenic and neurodevelopmental consequences of
coffe and caffeine exposure: a review on human and animal data.Neurotoxicology and
teratology, 16, 531-543.
Neubauer JA, Melton JE and Edelman NH (1990), Modulation of respiration during hypoxia.J
Appl Physiol, 68, 441-451.
Newby AC (1984), Adenosine and the concept of "retaliatory metabolites".Trends Biochem Sci,
9, 42-49.
Neylon M and Marshall JM (1991), The role of adenosine in the respiratory and cardiovascular
response to systemic hypoxia in the rat.J Physiol, 440,529-545.
Okada Y, Kawai A, Muckenhoff K and Scheid P (1998), Role of the pons in hypoxic respiratory
depression in the neonatal rat.Respiration Physiology, 111,55-63.
Okada Y, Muckenhoff K, Holterman G, Acker H and Scheid P (1993), Depth profiles of pH and
and PO2 in the isolated brainstem spinal cord of the neonatal rat.Respir Physiol, 93, 315-326.
Olson EB Jr (1987), Naloxone accelerates the rate of ventilatory acclimatization to hypoxia in
awake rats.Life Sci., 41, 161-167.
Onimaru H (1995a), Studies of the respiratory center using isolated brainstem-spinal cord
preparations. [Review] [79 refs].Neuroscience Research, 21, 183-90.
Onimaru H, Arata A and Homma I (1987), Localization of respiratory rhythm-generating
neurons in the medulla of brainstem-spinal cord preparations from newborn rats.
Neuroscience Letters, 78, 151-5.
Onimaru H, Arata A and Homma I (1988), Primary respiratory rhythm generator in the medulla
of brainstem-spinal cord preparation from newborn rat.Brain Research, 445,314-24.
Onimaru H, Arata A and Homma I (1990), Inhibitory synaptic inputs to the respiratory rhythm
generator in the medulla isolated from newborn rats.Pflugers Arch, 417,425-432.
Onimaru H, Arata A and Homma I (1997), Neuronal mechanisms of respiratory rhythm
generation: an approach using in vitro preparation.Japanese Journal of Physiology, 47, 385-
403.

Brainstem perinatal respiratory activity___________________________________________________________________________
52
Onimaru H, Ballanyi K and Richter DW (1996), Calcium-dependent responses in neurons of the
isolated respiratory network of newborn rats.J Physiol (London), 491,677-95.
Onimaru H and Homma I (1992), Whole cell recordings from respiratory neurons in the medulla
of brainstem-spinal cord preparations isolated from newborn rats.European journal of
physiol., 420,399-406.
Onimaru H, Arata A and Homma I (1995b), Intrinsic burst generation of preinspiratory neurons
in the medulla of brainstem-spinal cord preparations isolated from newborn rats.
Experimental Brain Research, 106,57-68.
Orlowski JP (1986), Cerebrospinal fluid endorphins and the infant apnea syndrome.Pediatrics,
78, 233-237.
Pasternak GW, Childers SR and Snyder SH (1980), Opiate analgesia: evidence for mediation by
a subpopulation of opiate receptors.Science, 208,514-516.
Paton JF and Richter DW (1995), Maturational changes in the respiratory rhythm generator of
the mouse.Pflugers Archiv European Journal of Physiology, 430,115-24.
Pierrefiche O, Schwarzacher SW Bischoff AM and Richter DW (1998), Blockade of synaptic
inhibition within the pre-Botzinger complex in the cat suppresses respiratory rhythm
generation in vivo.Journal of Physiology, 509,245-54.
Puig MM, Warner W Tang CK, Laorden ML and Turndorf H (1987), Effects of temperature on
the interaction of morphine with opioid receptors,.Br. J. Anaesth., 59, 1459-1464.
Ramirez JM, Quellmalz UJA, Wilken B and Richter DW (1998a), Developmental changes in the
hypoxic response of hypoglossus respiratory motor output in vitro.Journal of
Neurophysiology, 78, 383-392.
Ramirez JM, Quellmalz UJA, Wilken B and Richter DW (1998b), The hypoxic response of
neurones within the in vitro mammalian respiratory network.Journal of Physiology, 507,571-582.
Ramirez JM, Schwarzacher SW, Pierrefiche O, Schwarzacher SW and Richter DW (1998c),
Selective lesioning of the cat pre-Bötzinger complex in vivo eliminates breathing but not
gasping.Journal of Physiology, 507,895-907.
Ramirez JM, , Telgkamp P Elsen FP, Quellmalz UJA, and Richter DW (1997), Respiratory
Rhythm Generation In Mammals - Synaptic and Membrane Properties.Respiration
Physiology, 110,71-85.
Rehan V, Haider, AZ, Alvaro RE, Nowaczyk B, Cates DB, Kwiatkowski K and Rigatto H
(1996), The biphasic ventilatory response to hypoxia in preterm infants is not due to a
decrease in metabolism.Pediatric Pulmonology, 22, 287-294.
Rekling J, Champagnat J and Denavit-Saubié M (1996), Thyrotropin-releasing hormone (TRH)
depolarizes a subset of inspiratory neurons in the newborn mouse brain stem in vitro.Journal
of Neurophysiology, 75, 811-9.

Results and Discussion__________________________________________________________________________
53
Rekling JC and Feldman JL (1998), Prebötzinger complex and pacemaker neurons -
hypothesized site and kernel for respiratory rhythm generation [Review].Annual Review of
Physiology, 60, 385-405.
Richter DW (1982), Generation and maintenance of the respiratory rhythm.Journal of
Experimental Biology, 100,93-107.
Richter DW (1992), Mechanisms of respiratory rhythm generation. [Review].Current Opinion
in Neurobiology, 2, 788-93.
Richter DW, Bischoff A, Anders K, Bellingham M and Windhorst U (1993) Modulation of
respiratory patterns during hypoxia. In:Respiratory Control Central and Peripheral
mechanisms. Edited by D. F. Speck, M. S. Dekin, W. R. Revelette and D. T. Frazier.
Lexington: The university Press of Kentucky, pp. 21-28.
Rigatto H, Brady JP and Chir B (1972), Periodic breathing and apnea in preterm infants. II.
Hypoxia as a primary event.Pediatrics, 50, 219-28.
Runold M, Lagercrantz H and Fredholm BB (1986), Ventilatory effect of an adenosine analogue
in unanesthetized rabbits during development.J Appl Physiol, 61, 255-259.
Runold M, Lagercrantz H, Prabhakar NR and Fredholm BB (1989), Role of adenosine in
hypoxic ventilatory depression.J Appl Physiol, 67, 541-546.
Sawynok J (1998), Adenosine receptor activation and nociception.Eur J Pharm, 317,1-11.
Sawynok J, Sweeney MI and White TD (1989), Adenosine release may mediate spinal analgesia
by morphine [see comments]. [Review].Trends in Pharmacological Sciences, 10, 186-9.
Schmidt C et al. (1995), Adenosinergic modulation of respiratory neurones and hypoxic
responses in the anaesthetized cat.J Physiol, 483,769-781.
Schwieler (1968),Acta Phys Scand Suppl, 304,1-123.
Shao XM and Feldman JL (1997), Respiratory Rhythm Generation and Synaptic Inhibition Of
Expiratory Neurons In Pre-Bötzinger Complex - Differential Roles Of Glycinergic and
Gabaergic Neural Transmission.Journal of Neurophysiology, 77, 1853-1860.
Shatz C (1992), The developing Brain.Scientific American, 267,35-41.
Shook JE, Watkins WD and Camporesi EM (1990), Differential roles of opioid receptors in
respiration, respiratory disease, and opiate-induced respiratory depression.Am. Rev. Respir.
Dis., 142,895-909.
Sinton CM, Valatay JL and Jouvet M (1981), Gestational caffiene modifies offspring behaviour
in mice.Psycopharmacology, 75, 69-74.
Smith JC, Ellenberger HH, Ballanyi K, Richter DW and Feldman, JL (1991), Pre-Bötzinger
complex: a brainstem region that may generate respiratory rhythm in mammals.Science, 254,726-729.
Smith JC, Greer JJ, Liu GS and Feldman, JL (1990), Neural mechanisms generating respiratory
pattern in mammalian brain stem-spinal cord in vitro. I. Spatiotemporal patterns of motor and
medullary neuron activity.J Neurophysiol, 64, 1149-1169.

Brainstem perinatal respiratory activity___________________________________________________________________________
54
Snyder SH (1985), Adenosine as a neuromodulator. [Review].Annual Review of Neuroscience,
8, 103-24.
Sobotka TJ, Spaid SJ and Brodie RE (1979), Neurobehaviuoral teratology of caffeine exposure
in rats.Neurotoxicology, 1, 403-416.
Sollevi A (1997), Adenosine for pain control.Acta Anaesthesiologica Scandinavica.
Supplementum., 110,135-6.
Speck DF, Dekin DF, Revelette WR and Frazier DT (1993), Respiratory Control: Central and
Peripheral Mechanisms. Lexington: The University Press of Kentucky, pp. 238.
Steinbrook RA, Feldman HA, Fencl V, Forte VA Jr, Gobel RA, Leith, DE and Weinberger SE
(1984), Naloxone does not affect ventilatory responses to hypoxia and hypercapnia in rats.
Life Science, 34, 881-887.
Suzue T (1984), Respiratory rhythm generation in the in vitro brainstem-spinal cord preparation
of the neonatal rat.J Physiol, 354,173-183.
Taguchi Ket al. (1993), Effects of morphine on release of acetylcholine in the rat striatum: an in
vivo microdialysis study.Naunyn-Schmiedebergs Arch Pharmacol, 347, 9-13.
Thomas T, Elnazir, BK and Marshall JM 1994), Differentiation of the peripherally mediated
from the centrally mediated influences of adenosine in the rat during systemic hypoxia.Exper
Physiol, 79, 809-822.
Thomas T and Soyer KM (1996), The role of adenosine receptors in the rostral entrolateal
medulainthe cardiovascular response to defence area stimulaton in the rat.Exp Physiol, 81,67-77.
Thompson SM, Haas HL and Gahwiler BH (1992), Comparison of the actions of adenosine at
pre- and postsynaptic receptors in the rat hippocampus in vitro.J Physiol Lond, 451,347-63.
Traynor JR (1996), Theµ-opioid receptor.Pain Reviews, 3, 221-248.
Vitez TS, White TS and Eger El il (1974), Effects of hypothermia on halothane MAC and
isoflurane MAC in the rat.Anesthesiology, 41, 80-1.
Voipio J and Ballanyi K (1997), Interstitial P-Co2 and Ph, and Their Role As Chemostimulants
In the Isolated Respiratory Network Of Neonatal Rats.Journal of Physiology London, 499,527-542.
von Euler C (1980), Central pattern generation during breathing.TINS, 3, November, 275-277.
von Euler C (1991) Neuronal organization and rhythm generation. In:The Lung. Edited by R. G.
Crystal and J. B. West. New York: Raven Press, Ltd., pp. 1-12.
Waites BA, Ackland GL, Noble R and Hanson MA (1996), Red nucleus lesions abolish the
biphasic respiratory response to isocapnic hypoxia in decerebrate young rabbits.Journal of
Physiology, 495,217-25.
Winn HR, Rubio R and Berne RM (1981), Brain adenosine concentration during hypoxia in rats.
Am J Physiol, 241,235-242.
Wyman RJ (1977), Neuronal generation of the breathing rhythm.Ann. Rev. Phys., 39, 417-448.

Results and Discussion__________________________________________________________________________
55
Xia Y and Haddad GG (1991), Ontogeny and distribution of opioid receptors in the rat
brainstem.Brain Research, 549,181-193.
Yeadon M and Kitchen I (1989), Opioids and respiratory depression.Prog. Neurobiol., 33, 1-16.
Yeadon M and Kitchen I (1990), Multiple opioid receptors mediate the respiratory depressant of
fentanyl-like drugs in the rat.Gen. Pharmacol., 21, 655-664.