Embed Size (px)
Citation preview

Brain Research Bulletin, Vol. 1, pp. 489-492, i976. Copyright 0 ANKHO international Inc. Ail rights of reproduction in any form reserved. Printed in the U.S.A.
BRIEFCOMMUNICATION
Responses of Neurons in Rostra1 and Caudal
Trigeminal Nuclei to Tooth Pulp Stimulation’
Department of Neurology, Upstate Medical Center, Syracuse NY 13210
SAMUEL G. NORD
(Received 24 August 1976)
NORD, S. G. Responses of neurons in rostra1 ond caudul trigeminal nuclei to tooth pulp sti~u~tio~. BRAIN RES. BULL. l(5) 489-492, 1976. - Using ~mobil~ed, lightly anesthetized cats, the responses of neurons in the nucleus principals-subnucleus oralis and subnucleus caudalis regions of the sensory trigem~al complex were studied following electricat stimulation of the canine tooth pulp. Recording loci were verified histologically. pulpal stimulation activated 122 cells in the rostral nuclei and 44 in the caudal one. Neurons in the two, spatially segregated, regions exhibited different, though overlapping distributions of response and receptive field properties. More specifically, the rostra1 region cells tended to have lower thresholds and to reach peak firing rates at lower stimulus intensities. Their peripheral fields were generally more restricted and more frequently homolateral. Following supra-maximal stimulation, they ordinarily had briefer initial spike latencies and their response bursts typically contained a greater number of spikes. These findings are consistent with the view that each of the regions operates in a different manner in the mediation of ore-facial pain.
Nociception Pain mechanisms Trigeminal nuclei Dental pulp Somatosensory systems Brain stem
THE sensory trigeminal complex of the brain stem contains an abundance of neurons which respond to electrical stimulation of the tooth pulp. Although the neurons have an extensive longitudinal distribution, two particularly dense concentrations have been identified. One of these is approximately at the level of the obex in the subnucleus caudalis [3, 7, 8, I1 I. The other is located rostrally in contiguous portions of the subnucleus oralis and the nucleus principalis [ 1, 3, 8). Using the cat as an ex- perimental animal, Sessle et al. [81 have studied sub- stantial numbers of neurons in each group and have provided evidence that the two populations have appre- ciably different functional properties. Compared to rostra1 cells, for example, caudal neurons have longer latencies and more widespread peripheral fields; they are less frequently activated by contralateral thalamic stimulation, and they are influenced less by cortical conditioning stimuli [ 81. The results of other electrophysiological investigations in which either rostra1 [1, 3, 4, 9, 10, 131 or caudal [2, 3, 5-7, 11, 12, 141 neurons were studied in the cat indicate that additional differences may exist. Unfortunately, however, variations in experimental design and methodology pre- clude systematic, quantitative comparisons of their data. The present ~vestigation addresses this problem.
METHOD
Data were obtained from 16 immobilized (gallamine triethiodide), lightly anesthetized (sodium pentobarbital) cats. The animals were artificially ventilated and Pco, , arterial blood pressure and body temperature were main- tained at adequate physiological levels. Concentric, bipolar, stimulating electrodes were inserted into the pulpal canals of the ipsilateral mandibular, the ipsilateral maxillary, and the contralateral maxillary canine teeth and were cemented in place. Stimuli consisted of single, square pulses which typically were less than 100 rA in intensity and 2.5 msec in duration. In previous experiments we demonstrated that similar stimuli applied through identical electrode con- figurations did not excite receptors in adjacent, nondental tissue [ 5 1 .
The subnucleus caudalis and the subnucleus oralis- nucleus principalis regions noted above were explored with tungsten microelectrodes for neurons which responded to our pulpal stimuli. When a responsive cell was encountered, stimulus intensity was varied from below threshold to maximally effective values, and concomitant changes in response patterns were recorded. The responsiveness of each neuron to mechanical stimulation of dental and non-dental tissue was also noted.
’ This work was supported by NINCDS Grant NS 10814 and a grant from the Dr. Merrill H. Ross Medical Fund, Inc. The author is grateful to M. Agi, C. Bessmer, and D. Rolince for technical assistance.
489
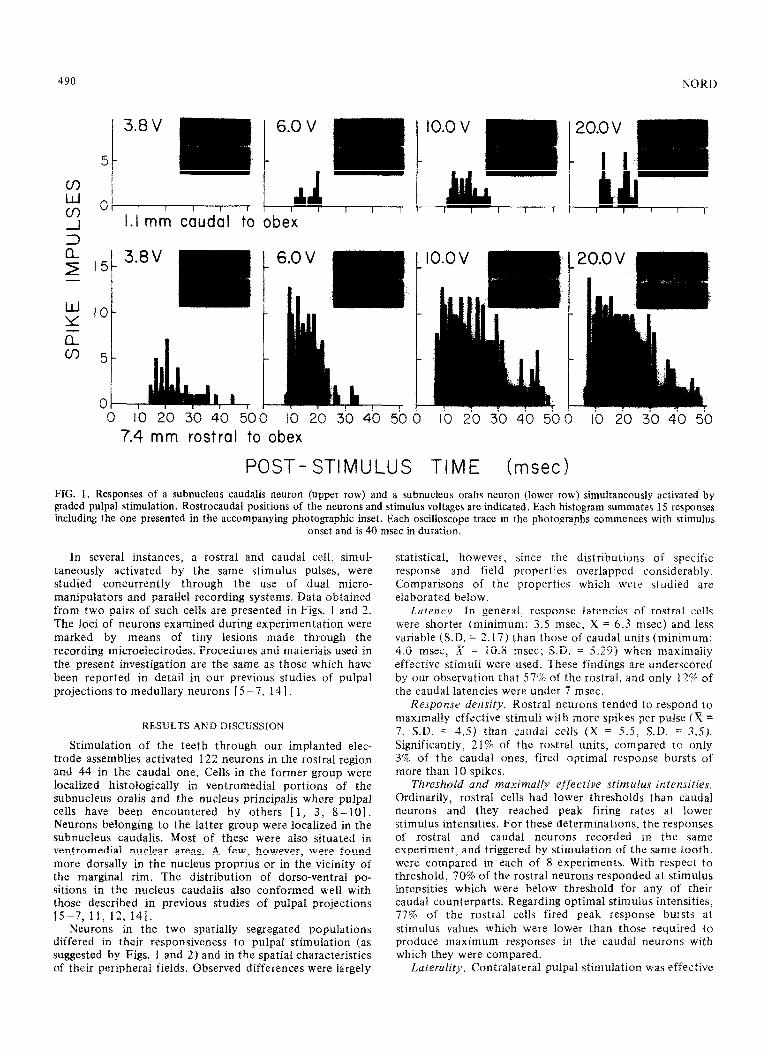
1.1 mm caudal to obex
7.4 mm rostra1 to obex
POST- STIMULUS TIME hsec> FIG. 1. Responses of a subnucleus caudalis neuron (upper row) and a subnucleus oralis neuron (lower row) simultaneously activated by graded pulpal stimulation. Rostrocaudal positions of the neurons and stimulus voltages are indicated. Each histogram summates 15 responses including the one presented in the accompanying photographic inset. Each oscilloscope trace in the photographs commences with stimulus
onset and is 40 msec in duration.
In several instances, a rostra1 and caudal cell, simul- taneously activated by the same stimulus pulses, were studied concurrently through the use of dual micro- manipulators and parallel recording systems. Data obtained from two pairs of such cells are presented in Figs. 1 and 2. The loci of neurons examined during experimentation were marked by means of tiny lesions made through the recording microelectrodes. Procedures and materials used in the present investigation are the same as those which have been reported in detail in our previous studies of pulpal projections to medullary neurons [ 5 -7, 14 1.
RESULTS AND DISCUSSION
Stimulation of the teeth through our implanted elec- trode assemblies activated 122 neurons in the rostra1 region and 44 in the caudal one. Cells in the former group were localized histologically in ventromedial portions of the subnucleus oralis and the nucleus principalis where pulpal cells have been encountered by others [ 1, 3, 8-101. Neurons belonging to the latter group were localized in the subnucleus caudalis. Most of these were also situated in ventromedial nuclear areas. A few, however, were found more dorsally in the nucleus proprius or in the vicinity of the marginal rim. The distribution of dorso-ventral po- sitions in the nucleus caudalis also conformed well with those described in previous studies of pulpal projections 15-7, 11, 12, 141.
Neurons in the two spatially segregated populations differed in their responsiveness to pulpal stimulation (as suggested by Figs. 1 and 2) and in the spatial characteristics of their peripheral fields. Observed differences were largely
statistical, however, since the distributions of specific response and field properties overlapped considerably. Comparisons of the properties which were studied are elaborated below.
Laiency. In general, response latencies of rostra1 cells were shorter (minimum: 3.5 msec, X = 6.3 msec) and less variable (SD. = 2.17) than those of caudal units (minimum: 4.0 msec, x = 10.8 msec; S.D. = 5.29) when maximally effective stimuli were used. These findings are underscored by our observation that 57% of the rostral, and only 12% of the caudal latencies were under 7 msec.
Response density. Rostra1 neurons tended to respond to lnaxinlally effective stimuli with more spikes per pulse (x = 7, SD. = 4.5) than caudal cells (x = 5.5. SD. = 3.5). Significantly, 21% of the rostra1 units, compared to only 3% of the caudal ones, fired optimal response bursts of more than 10 spikes.
T~r~s~oid and ~ax~~al~y efj‘ective st~~l~l~s il~tensities. Ordinarily, rostra1 cells had lower thresholds than caudal neurons and they reached peak firing rates at lower stimulus intensities. For these determinations, the responses of rostra1 and caudal neurons recorded in the same experiment, and triggered by stimulation of the same tooth, were compared in each of 8 experiments. With respect to threshold, 70% of the rostra1 neurons responded at stimulus intensities which were below threshold for any of their caudal counterparts. Regarding optimal stimulus intensities, 77% of the rostra1 cells fired peak response bursts at stimulus values which were lower than those required to produce maximum responses in the caudal neurons with which they were compared.
Laterfflity. Contralateral pulpal stimulation was effective
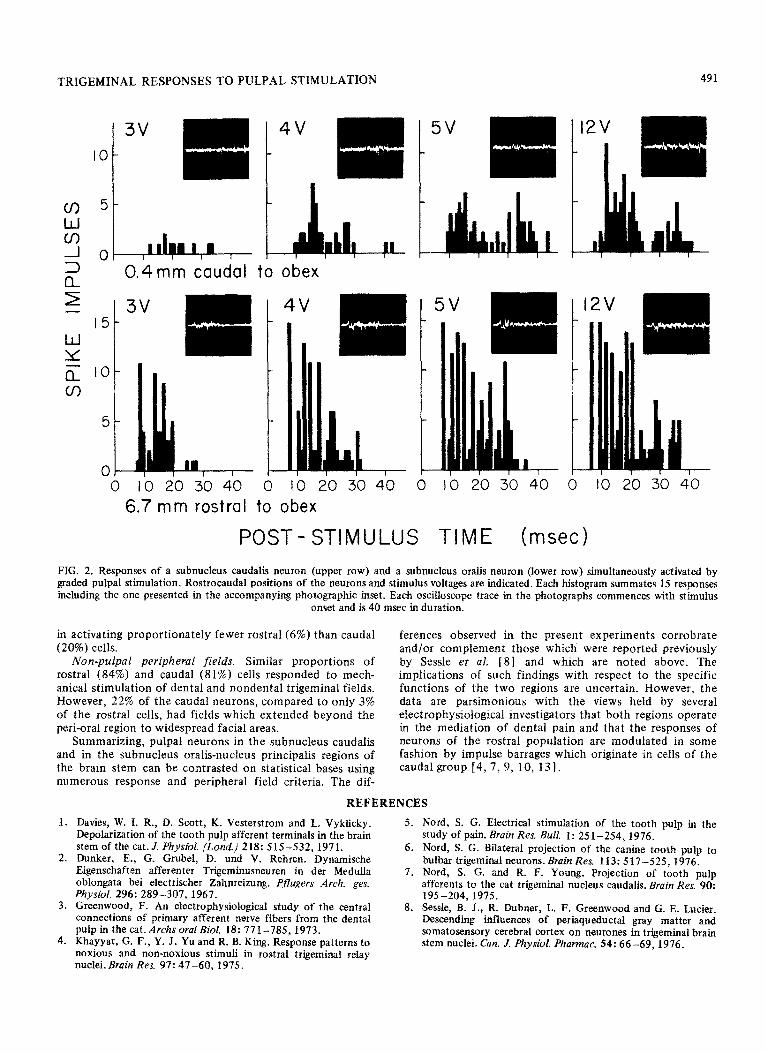
TRIGEMINAL RESPONSES TO PULPAL STIMULATION
0.4mm caudal to obex
6.7 mm rostra1 to obex
POST- STIMULUS TIME (msed
FIG. 2. Responses of a subnucleus caudalis neuron (upper row) and a subnucleus oralis neuron (lower row) simultaneously activated by graded pulpal stimulation. Rostrocaudal positions of the neurons and stimulus voltages are indicated. Each histogram summates 15 responses in&ding the one presented in the accompanying photographic inset. Each oscilloscope trace in the photog~p~s commences with stimulus
onset and is 40 msec in duration.
in activating proportionately fewer rostra1 (6%) than caudal (20%) cells.
Non-pulpal peripheral fields. Similar proportions of rostra1 (84%) and caudal (81%) cells responded to mech- anical stimulation of dental and nondental trigeminal fields. However, 22% of the caudal neurons, compared to only 3% of the rostra1 cells, had fields which extended beyond the peri-oral region to widespread facial areas.
Summarizing, pulpal neurons in the subnucleus caudalis and in the subnucleus oralis-nucleus principalis regions of the brain stem can be contrasted on statistical bases using numerous response and peripheral field criteria. The dif-
ferences observed in the present experiments corrobrate and/or complement those which were reported previously by Sessle et al. [8] and which are noted above. The implications of such findings with respect to the specific functions of the two regions are uncertain. However, the data are parsimonious with the views held by several electrophysiological investigators that both regions operate in the mediation of dental pain and that the responses of neurons of the rostra1 population are modulated in some fashion by impulse barrages which originate in cells of the caudal group [4,7,9, 10, 131.
REFERENCES
Davies, W. I. R., D. Scott, K. Vesterstrom and I,. Vyklicky. Depolarization of the tooth pulp afferent terminals in the brain stem of the cat.J. Physiol. (Lwzd.] 218: 515-532,197l. Dunker, E., G. Grubel, D. und V. Rehren. Dynamische Eigenschaften afferenter Trigeminusneuren in der Medulla oblongata bei electrischer Zahnreizung. Pflugers Arch. ges. Physiol. 296: 289-307,1967. Greenwood, F. An electrophysiological study of the central connections of primary afferent nerve fibers from the dental pulp in the cat. Archs oral Biol. 18: 771-785, 1973. Khayyat, G. F., Y. J. Yu and R. B. King. Response patterns to noxious and non-noxious stimuli in rostra1 trigeminal relay nuclei. Brain Rex 97: 47-60, 1975.
5.
6.
7.
8.
Nord, S. G. Electrical stimulation of the tooth pulp in the study of pain. Brain Res. Bull. 1: 25 l-254,1976. Nord, S. G. Bilateral projection of the canine tooth pulp to bulbar trigeminal neurons. Brain Res. 113: 517-X25,1976. Nord, S. G. and R. F. Young. Proiection of tooth uulu afferents to the cat trigeminal nucleus caudalis. Brain Res.~9d: 195-204, 1975. Sessle, B.. J., R. Dubner, L. F. Greenwood and G. E. Lucier. Descending influences of periaqueductat gray matter and somatosensory cerebral cortex on neurones irt trjgeminal brain stem nuclei. Can. J. Physiol. Pharmac. 54: 66-69, 1976.

492 NORD
9. Sessle, B. J. and F. Greenwood. Influence of trigeminal nucleus caudalis on the resuonses of cat triaeminal brain stem neurons
12. Yokota, T. Two types of tooth pulp units in the bulbar lateral reticular formation. Brain Res. 104: 325-329. 1976.
with orofacial mechanoreceptive- fields. Brain Res. 67: 330-333,1974.
10. Sessle, B. J. and F. Greenwood. Effects of trigeminal tractotomy and of carbamazepine therapy on single trigeminal sensory neurones in cats. J. dent. Res. 54B: 3201 -B206. 1975.
11. Yokota, T. Excitation of units in marginal rim of subnucleus caudalis elicited by tooth pulp stimulation. Brain Rex 95: 154-I 58,1975.
13. Young, R. F. and R. B. King. Excitability changes in trigeminal primary afferent fibers in response to noxious and nonnoxious stimuli.,L Neurophysiol. 3.5: 87-95, 1972.
14. Young, R. F. and S. G. Nord. Experimental modulation of medullary dental pulp units by mechanical stimulation of oro-facial fields. E’xpl. A’euvol. 49: 8 I3 -82 1, 1975.























