Embed Size (px)
Citation preview

BIOCHIMIE, 1977, 59, 275-285.
RNAs viraux des cellules infectdes par un oncornavirus :
I . - Caract6risation des RNAs viraux associ6s h des particules cytoplasmiques.
M. L. MICHEL, M. ROUSSEL, E. POITEVIN, A. SAMSO, et C. J. LARSEN ~.
Laboratoire d'H~matologie Expdrimentale (Directeur : Prof. M. Boiron), 10, Place du Docteur Fournier, H~pilal Saint-Louis, 75010 Paris.
(15-11-1976).
Summary. - - Oncornaviral particles on the way of tireir synthesis have been isolated from cytoplasm of a Friend virus-producing murine cell line. By use of electron microscopy and retr~)transcriptase assay, it was found that tke majori'ty of the viral structures is pelleted by sedimentation at 90.000 g. Tke Poly (A) RNA of this laellet conta5ns principally 70S and 35S RNA spedes which, as judged by size determi.n'afion and hybridization with Friend-virus c-DNA are the species embedded into tire virions. ~',e verified that conditions which were used in tire course of isolation, effectively dissociate polysomes and prevent ttrem from co-sedimenting with viruses.
INTRODUCTION.
L 'observa t ion des par t icules de type C en micro- scopie 61ectronique a Iuontr6 qu'el les sont com- pos6es de deux par t ies dist inctes : une r ibonucl6o- prot6ine centrale, le core ou nucl6oide qui con- t ient le g6nome et une enveloppe externe [1]. Dans les cellules qui p rodu i sen t des oncornavi rus , 1'as- semblage des v i r ions a l ieu au vois inage de la membrane plasmique, car c 'est seulement h cette place qu 'on peut observer des s t ructures de forme tor ique bourgeonnant h la surface cel lula i re en m6me temps qu'el les acqui6rent une forme sph6- rique. L 'o rganisa t ion du g6nome vi ra l est assez bien connue. I1 s 'agit d 'un complexe 60-70S form6 par l ' associa t ion de deux sous-unit6s ident iques 30-40S et d 'un nombre var iable de mol6cules de peti ts RNAs qui pour ra i en t s tabi l iser le complexe [2, 3]. Darts les cellules product r ices , on a pu carac t6r iser deux esp6ces mol6cula i res exis tant dans les polysomes li6s (RNAs 35 et 20S) et l ibres (RNA 35S) [4, 5]. On a pu mon t r e r que ces espbces sont ac t ivement impliqu6es dans la synthbse des polypept ides v i raux [6, 7, 8].
A c e jour, peu de t ravaux ont 6t6 consacr6s h la caract6r isat ion des RNAs des par t icules virales en vote de synth6se dans les cellules [9, 10]. Panem et coll. ont pu isoler d 'un extra i t cy toplasmique de cellules p roduisan t le virus leuc6mog6ne de Kirs- ten des par t icules de densit6 1,14 et 1,22 qui con- tenaient deux RNAs 36 et 20-24S [10]. Toutefois,
Abr~viatio.ns : HEPES : (N-2-Hydroxyethylpip~,razine N'-2-6thane-
sulforrique acide) ; MEM: (Minimum Essential Me- dium) ; SDS : (Dod~cyl, sulfate de Sod~ium).
A qui route correspondanee doit 6tre adress6e.
le seul cr i t6re de la tail le et des arguments met tant en jeu la vitesse d ' appar i t ion de ces RNAs dans les par t icules avaient 6t6 retenus pour carac t6r iser ces esp6ces. Par ail leurs, les quantit6s extr6me- ment faibles de RNA renda ien t difficile une exp6- r imen ta t ion accrue. Dans le pr6sent travail , nous avons cherch6 h carac t6r iser les RNAs des part i - cules virales des cellules en met tant h prof i t l 'exis- tence d 'un segment de polyad6nylate (Poly (A)) h leur extr6mit6 3'OH pour les s6parer des autres RNAs cel lulaires par une ch romatograph ie sur colonne d 'ol igo dT-cel lulose [11, 12], puts en les h y b r i d a n t ~ du DNA synth6t ique comp16mentaire du g6nome viral. Nous avons observ6 que les par- t icules virales en vote de synth6se peuvent 6tre concentr6es darts une f rac t ion du cytoplasme s6di- Inentant h 90.000 g. Le Poly (A) RNA de cette f rac t ion cont ient p r inc ipa l emen t du RNA 35S et
un degr6 m o in d re du RNA 70S. Ces deux RNAs hybr iden t avec le DNA compl6menta i re du g6nome viral.
A) MATERIELS ET METHODES.
1) Cellules : culture et marquage.
La souche Fr iend-Evel ine , obtenue h par t i r de fibroblastes de souris STU infect6s par le virus de Fr iend, nous a 06 grac ieusement fournie par ]e Prof. Sch~fer (Tubingen) [131. Elle est cult iv6e en suspension dans du mi l ieu de Eagle modif i6 (MEM) addi t ionn6 de 10 p. cent de s6rum de veau embryonna i r e et sans ant ibiot iques. Le temps moyen de g6n6ration est de 2,0 h 22 heures. Les cellules ne cont iennent pas de mycoplasmes visi- bles au mic roscope 61ectronique.
Pour m a rq u e r les cellules par le 32p, on ense- mence une cul ture h une concen t ra t ion ini t ia le de

276 M. L. M i c h e l et coll.
400.000 cell /nd, dans un mil ieu MEM priv6 de phosphate et t amponn6 par de I'H'EPES (2 raM). Six heures plus tard, le ~2p-orthophosphate (CEA, Saelay, France) est ajout6 h ra ison de 10 h 20 ~e/ nil. La cul ture est poursuiv ie pendan t 16 heures ou 40 heures. Dans le cas du marquage pendan t 40 heures, on dilue la culture avec 1/3 de MEM normal aprGs 24 heures. La vitesse de croissance cellulaire est notablement ralent ie par le phos- phore 32, alors qu'elle n 'est pus modifi~.e par le mil ieu de marquage sans phosphore radio-act if pendan t au moins 72 heures.
La souche D.55 utilis~e pour l 'expfirience de re- cons t ruct ion (V. I/Gsultats), provient de fibro- blastes embryonna i res de souris Swiss. Les eel- lules sont cultiw:~es en nmnocouche dans du MEM addi t ionn6 de 10 p. cent de s@um de veau em- b ryonna i re . Pour le marquage par l ' u r id ine 8H, le contenu de 10 rollers a 6t6 rbcolt6 par t ryps ina- tion, lay6 avec du mil ieu frais et suspendu dans un <<spinner>> contenan t 50 ml de MEN. AprGs 20 ran, 1 mc d 'Ur id ine [3H] (CEA, Saclay) a 6t6 ajoutG. Le lnarquage a 6t~ poursuivi pendan t 40 minutes.
2) Obtenlion des extraits cgtoplasmiques :
Les cellules sont recueil l ies par centr i fugat ion des mil ieux de cul ture A 3.000 rpm et rincGes rapt- dement dans du t ampon hypotonique glac6 : Tris- HC1, 0.01 M, pH = 8,2, EDTA 0.001 M, 5 p. cent de saccharose. Apr~s centr i fugat ion, elles sont resuspendues dans le m~,me tampon hypotonique. Dans certains cas, du mercapto-Gthanol (10-2 M) est ajout~ ~ la suspension. Apr~s 1,0 minutes dans la glace, la suspension est IransfGr~e dans un homogGnGiseur type Potter dont le pis ton est entra ln6 par un moteur t ou rnan t approximat ive- ment ~ 600 rpm. On donne dix coups de pis ton aller et retour. L 'homog~nat cellulaire est centri- lug6 h 1.500 rpm pendan t 5 minutes dans une cen- tr ifugeuse Jonan. Le culot nuclGaire est resus- pendu dans le t ampon hypotonique soumis au mSme t ra i tement et centrifug~ & nouveau. Les sur- nageants sont mGlangGs et centrifugGs 10 minutes
1.500 rpnl ; puts 10 minutes dans une centr i fu- geuse Sorvall & 10.000 rpm. Cette deuxibme cen- t r i fugation, destinGe 5 61trainer les mi tochondr ies a ~t6 supprim6e par ta suite.
L 'extrai t par t icula i re du cytoplasme est pr6par6 en d6posant le surnageant pr6c~dent (25 ml) sur un coussin de glycfrol ~ 20 p. cent dans le m6me tampon, dans un tube de rotor Beckman SW-27. Apr6s 90 nm de eentr i fngat ion ~ 24.000 rpm, on obtient un cnlot t rans lucide appel6 P-90 et un sur- nageant S-90, qui est d~cant6. Dans les expOfiences
BIOCHIMIE, 1977, 59, n o 3.
de sonicat ion, le matGriel du culot P-90 est repris duns 1 lnl de Tampon Tris-H,C1 0.01 M, pH 7.4, NaC1 0.01 M, EDTA 0.001 M (NTE) et trait6 pen- dant 2 fois 20 secondes dans un sonicateur MSE, avec la petite sonde h la posi t ion M~dium - - 5 - - . Le tube est anaintenu dans la glace.
3) Obtention des RNA polgaddnglOs (Polg (A) RNA) par chromatographic snr eolonne d'oligo dT-eellulose :
Les colonnes sont prGpar~es avec 2.0.0 mg d'oligo dY-cellulose (PL-grade 7) mblang~s h une 6gale quantit~ de cellulose pure (Macherey et Nagel). Elles sont lavGes et 6quilibrGes dans une solut ion de Na('l 0.5 M e t SDS 0.5 p. cent. Le matGriel du culot P-90 est d i rectement suspendu dans cette solution et incub6 pendan t 15 '~ 20 minutes h 20- 22 ° avant d'Gtre charg6 sur la colonne. Pour le surnageant S-90, on l 'amGne aux concent ra t ions requises de Nag1 et de SDS. AprGs un lavage suivi par l 'Gvolution de la densit6 optique h 260 nm, on 61ue les Poly (A) RNAs par de l 'eau bidistillGe et on les prGcipite par 2 volumes d'Gthanol absolu en prGsence de NaC1 0.1 M, pendan t une nu i t h - - 3 0 ° C . Les poly (A) RNAs sont p ra t iquement purs, ainsi qu 'en tGmoignent les rapports spectro- photom6tr iques 260 nm/280 mn et 260 ran/230 nm. Pour les expbrienees d 'hybr ida t ion , on les trai te par le ph6nol-SDS, afin d'61iminer route trace de protGines rGsiduelles [141.
4) Gels de polgaerglamide el gradients de sae- c h t l r o s e :
• Les gels mous h 1,7 p. cent de po lyacry lamide - 0,5 p. cent d'agarose sont prGpar~s selon la m6- thode de Tiollais et coll. [15]. Des fract ions de 1,5 mm sont d@oup@s et d i rectement compt~es grfice h l'effet Cerenkov dans un compteur h scin- t i l lat ion Packard 3,390, ou bien 61u~es par 0,5 ml d 'eau pendan t une nui t h 37 ° et mGlang~s avec 2,5 ml d ' Instagel (Packard).
Les Poly (A) RNAs sont analysGs par centr i fu- gation duns des gradients de saccharose 10- 30 p. cent duns du tampon NTE. Les gradients sont centrifuges h 4°C pendan t 2 h 30 h 44.000 rpm duns un rotor Spinco SW-50. Ils sont collectGs h l 'a ide d 'un apparei l Isco.
5) Preparat ion de DNA sgnthdt ique compI~.men- taire du RNA de virus de Fr iend :
Le virus de F r i end produi t par les cellules Fr iend-Eve l ine est isol6 h par t i r de cultures de 24 heures. Les nl i l ieux sont clarifiGs par centr i fu- gation h 10.000 rpm pendan t 2.0 minutes dans un rotor JA-10 de centrifugeuse Beckman J-21. On ajoute 8 p. cent de Polyethylene glycol 6.000 (Baker) et on agite la suspension h 4°C pe nda n t
R N A s d e s o n c o r n a v i r u s a s s o c i d s a u x c e l l u l e s . 2 7 7
3 h e u r e s , a v a n t de la c e n t r i f u g e r p e n d a n t 20 mi - n u t e s h 10.000 r p m . Les c u l o t s c o n t e n a n t ]e v i r u s s o n t r e d i s s o u s d a n s d u N T E et d6pos~s s u r u n g r a d i e n t de s a c c h a r o s e 10-70 p. c e n t d a n s u n t u b e de r o t o r SW-27. A p r b s 2 h e u r e s de c e n t r i f u g a t i o n
26.000' r p m , la b a n d e v i r a l e t r~s ~pa i s se es t re - cue i l l i e h l ' a i d e de l ' a p p a r e i l I sco , p r 6 c i p i t 6 e p a r s 6 d i m e n t a t i o n p e n d a n t 50 m i n u t e s h 10.000 r p m
d a n s u n r o t o r 50 T i t a n e , et p u r i f i 6 d a n s u n n o u - v e a u g r a d i e n t de s a c c h a r o s e 10-70 p. c e n t p e n - d a n t u n e n u i t h 30.000 r p m . Le v i r u s est c o n c e n t r b p a r c e n t r i f u g a t i o n et c o n s e r v ~ h - - 8 , 0 ° C .
Le c-DNA est p r 6 p a r ~ d a n s u n e r ~ a c t i o n e n d o - gbne m e t t a n t e n j eu le v i r u s p u r i f i ~ , s e l o n les c o n - d i t i o n s e x p 6 r i m e n t a l e s de Gal lo et col l . ~16]. Les m ~ l a n g e s r 6 a c t i o n n e l s c o n t i e n n e n t 3 m g / m l de
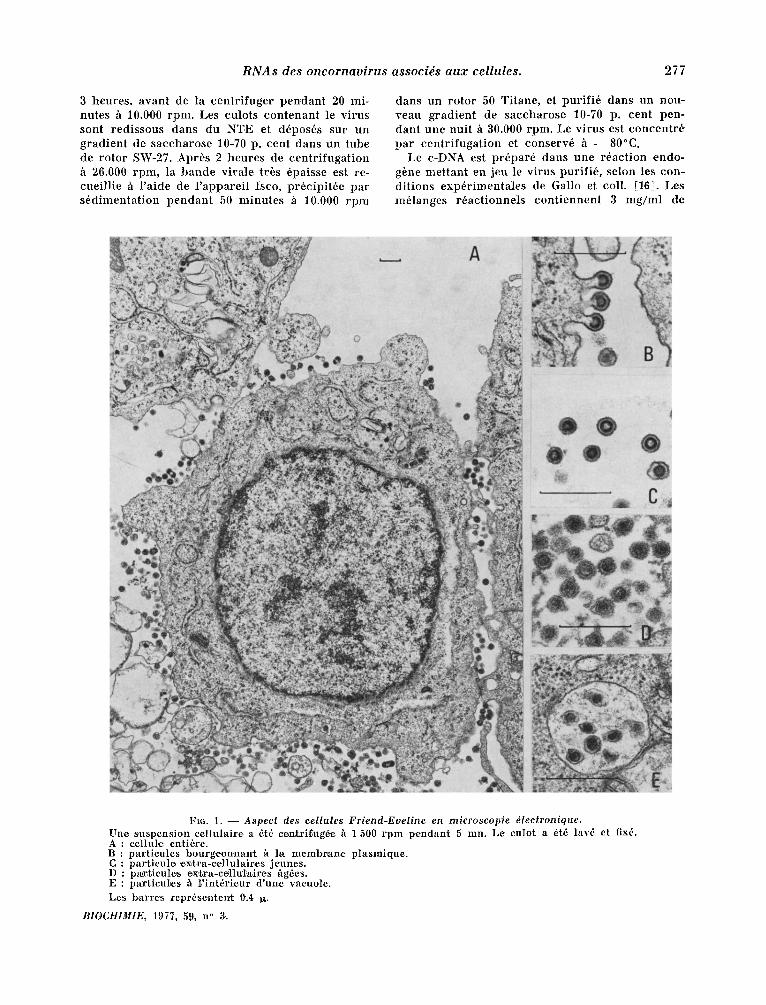
Fro. 1. - - Aspect des cellules Friend-Eueline en microscopie ~lectronique. Une suspens ion eel lu la i re a ~t6 ee,ntrifug6e h 1 500 r p m p en d an t 5 ran. Le eu]ot a ~t~ lay6 et fix6. A : eellule er~tibre. B : pa r t i eu les bourgeon 'nant h la membrane plasmique . C : par t ieul~ emtra-cel lulaires jeunes. D : p~rtieul~s ,ex~hra-ee~llula.ires ~g~es. E : par t ieu les h l ' in t~r ieur d 'une vacuole. Les ba r r e s repr~senter~t 0.4 ~.
BIOCH1MIE, 1977, 59, n ° 3.

.978 M. L. M i c h e l el coll.
prol6ines virales ; 50 mM de Tris-HCl, pH 7,5; 20 mM de KC1, 2 mM de di thiothrei tol ; 1 mM de MnC12, 1 mM de dATP dGTP dG.TP froids, 0.1 mM de TI'P[ZH] (activit6 sp6cifique : 0,5 C/raM), 50 ~xg/ml d ' ac t inomyc ine ,D (.Sigma) et 0,05 p. cent de Tr i ton X-100 (Packard). Apr6s 2 heures d ' incu- bat ion, le c-DNA est isol6 selon le protocole d6crit par Tavi t ian et coll. [17]. La majeure par t ie du produi t migre h 5-6,S dans un gradient de centr i - fugation de saccharose. Mats 5-20 p. cent du pro- duit nfigrent /~ la posi t ion du RNA r ibosomal 28S, ce qui ind ique qu 'une petite p ropor t ion des mol6- cules de DNA a une taille presque 6quivalente /~ celle du g6nome viral. Ce r6sultat est couforme /~ ceux d 'autres groupes qui out montr~ qu 'une aug- menta t ion de la concent ra t ion en pr6curseurs accroit la taille du DNA [18, 19]. Le c-DNA syn- th6tis6 en presence d ' ac t inomyc ine D est prot6g6 h plus de 85 p. cent par du RN~A viral (0,5 ~g), alors que le p rodui t p rovenan t d 'une r6act ion ne con tenan t pas d ' ac t inomyc ine D n'est prot6g6 qu'h 30 p. cent par le m~me RNA. Ce r6sultat conf i rme ceux d 'autres groupes [20].
6) Tests d 'hybr idat ion entre c-DNA viral el RNAs :
a) Pour d6terminer les p ropor t ions de RNA viral dans les cellules productr ices . Mille h 2.0.00 cpm de c-DNA sont incub6s dans uu volmne final de 60 ~1 con tenan t 40 p. cent de Formamide , 0,015 M Tris-HC1 pH 7,4, Na,CI 0,2:5 M e t des quan- tit6s croissantes de RNA [21]. Les $chant i l lons sont plac6s duns des micropipet tes de 100 ~1 pr6a- lablement silicon6es et incub6s pendan t 48 h 72 h b 41°C. Les hybr ides soul d6tect6s par leur r6sis- tance ~ la nucl6ase S~ [22]. Les blanes ddterminds h par t i r d '6chant i l lons sans I~NA (5 /~ 7 p. cent du DNA initial) sont soustraits des volumes obtenus pour les hybr ides .
b) Pour 6tudier la d is t r ibut ion du RNA viral duns les gradients de centr i fugat ion. Les lqNAs de chaque fract ion des gradients sont co-pr~cipit6s avec 50 ~g de RNA 4S de levure et redissous darts un volume 6gal de tampon d 'hybr ida t ion . Un ali- quot est hybr id6 /~ une quant i t6 fixe de c-DNA (500/~ 1.00'0 cpm) selon les condi t ions d6crites pr6- c~demment.
7) Microscopie ~lectronique :
Les cellules et les culots r6sul tant de la centr i - fugation du cytoplasme h 90.0.00 g sont fixes par du glutarald6hyde h 2,5 p. cent duns u n tampon de S6rensen et postfix6s par l 'ac ide osmique, 1 p. cent b ichromate de K 2 p. cent. Ils sont ensuite, lav6s, d~shydrat~s et inclus. Les coupes sont examin6es • h l 'aide d 'un microscope Phi l ips EM 300.
RESULTATS.
1) Iso lement de part icules virales darts la trac- t ion cy topIasmique des cellules Friend-Eoel ine :
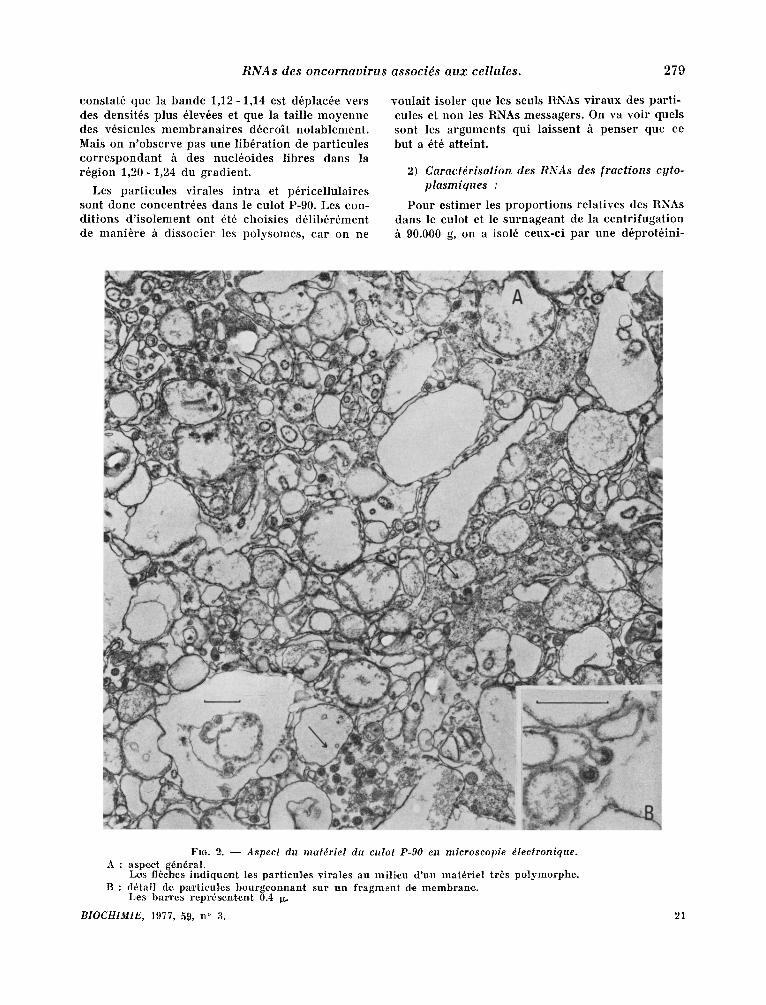
l .orsqu 'on examine en microscopie 61ectronique des coupes de cellules, qui ont ~t6 r6cott6es en phase exponent iel le de croissance, on peut obser- ver, p lus ieurs types de s t ructures virales (fig. 1). Ce sont tout d 'abord parrot les par t icules extra- cellulaires, les virus mOrs ou fg6s, qui poss~dent un core dense aux ~lectrons et d 'autres plus jeunes dont le core est plus ou moins t ransparent . A la surface cellulaire et dans le cytoplasme, on dist in- gue deux lypes de s t ructures virales : des par t i - cules bourgeonnantes en divers ~tats, depuis le simple 6paississement sous la membrane plasmi- que jusqu 'au v i r ion complet pr~t h se d6tacher ; des part icules "it l 'aspect mal d6fini, regroup6es dans des vacuoles et ressemblant h des v i r ions hgbs qui aura ient pe rdu leur enveloppe externe. Pour isoler les part icules virales des cellules, le cytoplasme a 6t6 pr~par6 selon ]e proloco]e dbcrit danz les mat6riels et m~thodes. Apr~s ~clatement des cellules duns un tampon hypotonique, le cyto- plasme a 6t6 d~barrass6 des noyaux et des mito- chondr ies par des centr i fugat ions diff6rentielles h 1.500 et 10.000 rpm (par la suite, la centr i fugat ion h 10.0.00 rpm a 6t6 omise). I1 a 6t6 ensuite centri- lug6 clans des condi t ions choisies pour s6dimenter les part icules virales completes ou en vole en for- marion. Les coupes pr6par6es ~ par t i r d 'un tel culot (culot P-90) mon t ren t la presence d 'un ma- t6riel tr6s po lymorphe off l 'on pent r econna i t re de tr~s nombreuses membranes plasmiques et ergasto- plasnfiques (fig. 2). La grande majorit6 des mem- branes est tisse, on pent en observer quelques unes revalues de structures h l 'aspect r ibosomique. On reconnai t ais6ment des part icules virales h divers stades de leur formation. Afin de s~.parer ce mat6- riel viral des const i tuants cellulaires, le culot P-90 a 6t6 resuspendu darts du tampon NTE, pH 8,2, puts centrifug~ clans un gradient de saccharose 10-70 p. cent. Deux bandes s ' ind iv idua l i sen t , l ' une darts la zone de densit6s 1,12 - 1,14 g /ml et l 'autre au sommet du tube (densit6 inf~rieure h 1,10 g/ ml). Des coupes du mat6riel venan t de ces deux bandes r~v~lent la presence de v i r ions dans la bande la plus lourde. Cette observat ion est confir- m6e par la caractdrisat ion d 'une activit6 t ranscr ip- tase inverse en pr6sence d 'une matr ice artificieile oligo rA-dT. Un pic un ique et homog6ne de Poly dT est centr6 sur la bande 1,12 - 1,14.
Des essais de sonica t ion du materiel du culot P-90 destin6s "h facil i ter une 6ventuelle l ib@ation des part icules virales ne nous out pus donn~ de r6sultats sup@ieurs aux pr~cddents. Nous avons
BIOCHIMIE, 1977, 59, n ° 3.
R N A s des o n c o r n a v i r u s associds a u x cel lules. 2 7 9
c o n s t a t 6 que la b a n d e 1 , 1 2 - 1,1.4 es t d6plac6.e v e r s des d e n s i t 6 s p l u s 61ev6es et que la t a i l l e m o y e n n e des v 6 s i c u l e s m e m b r a n a i r e s d 6 c r o l t n o t a b l e m e n t . Mais o n n ' o b s e r v e p a s u n e l i b 6 r a t i o n de p a r t i c u l e s c o r r e s p o n d a n t h des nuc16o ides l i b r e s d a n s la r 6 g i o n 1,2 '0- 1,24 d u g r a d i e n t .
Les p a r t i c u l e s v i r a l e s i n t r a et p 6 r i c e l l u l a i r e s s o n t d o n c c o n c e n t r 6 e s d a n s le c u l o t P-90. Les c o n - d i t i o n s d ' i s o l e m e n t on t 6t6 c h o i s i e s d 6 1 i b 6 r 6 m e n t de m a n i ~ r e h d i s s o c i e r les p o l y s o m e s , c a r o n n e
v o u l a i t i s o l e r q u e les seu ls RN,As v i r a u x des p a r t i - eu les et n o n les RNAs m e s s a g e r s . O n v a v o i r q u e l s s o n t les a r g u m e n t s q u i l a i s s e n t h p e n s e r q u e ce b u t a 6t6 a t t e in t .
2) Caractdrisation des RNAs des [ractions cyto- plasmiques :
P o u r e s t i m e r les p r o p o r t i o n s r e l a t i v e s des RNAs d a n s le cu lo t et le s u r n a g e a n t de l a c e n t r i f u g a t i o n h 90.000 g, o n a isol6 c e u x - e i p a r u n e d ~ p r o t 6 i n i -
FIG. 2. - - Aspect du matdriel du culot P-90 en microscopie dlectronique. A : aspect g6n~ral.
L~s fl~ehtes indique~t les particul.es viral,es au mi l ieu d 'un mater ie l tr~s polymorph,e. B : d6tail d:e par t i cu les bourgeon, nan.t sur un f r agmen t de membrane .
Les bar res repr~sent.enl 0.4 Ix.
BIOCHLVllE, 1977, 59, n ° 3. 21

280 M. L. Miche l el coll.
s a t i o n b. l ' a i d e de S D S - p h 6 n o l . Les r~su l t a t s de la t a b l e I m o n t r e n t que l a t r b s g r a n d e m a j o r i t 6 des RNAs c y t o p l a s m i q u e s t o t a u x r e s t e d a n s le s u r n a - gean t . Le cu lo t P-90 c o n t i e n t m o i n s de 10 p. c e n t d ' e n t r e eux. P o u r d ~ t e r m i n e r l e u r t e n e u r e n P o l y (A), les RNAs d u s u r n a g e a n t et d u cu lo t o n t 6t6 c h r o m a t o g r a p h i 6 s s u r u n e c o l o n n e d ' o l i g o dT- ce l lu lose . Les r ~ s u l t a t s e x p r i m ~ s en f o n c t i o n d u R N A c y t o p l a s m i q u e t o t a l i n d i q u e n t que le c n l o t P-9O c o n t i e n t u n p e u p l u s d u 1 / 5 des RNAs p o l y - a d ~ n y l ~ s du c y t o p l a s m e .
TABLE I.
R~partit ion des RNAs cytoplasmiques dans le sur- nageant et le culot de centrifugation & 90.0.00 g.
RNA total
Poly (A) RNA
S-90
93 p. cent
1,1 p. cent
P-90
7 p. eent
0 , 2 - 0 , 3 p. eent
Les va leurs ment ionn~cs , qui regr~senten~t l~a moyen- ne de 3 exp6rienees r6alis6es avee des celhfles n o n ma.rqn6es, ,ont fit6 obSen,ues pa r speet rophotom~tr ie (1 UD.0m, = 40 lag de RNA). T o u s l e s po*u.reentages sont expr im~s pa r r appor t au RNA cy top lasmique total .
Le f a i b l e p o u r c e n t a g e de R N A d a n s le cu lo t sug- gbre que la t r b s g r a n d e m a j o r i t 6 des p o l y s o m e s es t r e s t6e d a n s le s u r n a g e a n t p u i s q u e les RNAs r i b o - s o m i q u e s c o n s t i t u e n t j u s q u ' h 80 p. c e n t des RNAs c y t o p l a s m i q u e s . Ce r 6 s u l t a t l a i s s e h p e n s e r que ]es RNAs m e s s a g e r s des p o l y s o m e s d o i v e n t 6 t re p a s s 6 s d a n s le s u r n a g e a n t . P o u r v 6 r i f i e r t r b s p r 6 c i s 6 m e n t ce p o i n t , n o u s a v o n s m a r q u 6 a v e c de l ' U r i - d i n e [UH], (20 g c / m l ) p e n d a n t 40 n f i n u t e s , u n e cul - t u r e de ce l lu l e s e m b r y o n n a i r e s n o n i n f e c t 6 e s de s o u r i s Swiss . A p r ~ s ce t e m p s , la g r a n d e m a j o r i t 6 de la r a d i o - a c t i v i t 6 i n c o r p o r 6 e d a n s les RNAs cy to - p l a s m i q u e s se r 6 p a r t i t en u n e l a r g e b a n d e e n t r e 4 et 20S d a n s u n g r a d i e n t de s a c c h a r o s e e n v61o- c i t6 ( r6 su l t a t n o n p r 6 s e n t 6 ) . Ce r 6 s u l t a t i n d i q u e que les seu ls RNAs m e s s a g e r s s o n t m a r q u 6 s p e n - d a n t ce d61ai. Ces ce l lu l e s o n t 6t6 e n s u i t e r 6co l t 6e s et a jou t6es h des ce l l u l e s F r i e n d - E v e l i n e n o n m a r - qu~es . Le m61ange a 6t6 divis6, e n d e u x p a r t s ~gales et l ' o n a isol6 la f r a c t i o n c y t o p l a s m i q u e de cha - c u n e d ' e n t r e e l les e n p r 6 s e n c e d ' i o n s Mg +÷ (10 -z M) p o u r la p r e m i b r e et d ' E D T A , h la c o n c e n t r a t i o n h a b i t u e l l e p o u r la s e c o n d e . Les c u l o t s et les s u r n a - g e a n t s de c e n t r i f u g a t i o n o n t 6t6 a n a l y s ~ s p o u r l e u r t e n e u r en p r 6 c u r s e u r r a d i o - a c t i f i n c o r p o r ~ d a n s la f r a c t i o n TCA p r 6 c i p i t a b l e . E n p r e s e n c e de Mge÷, la m a j e u r e p a r t i e de la r a d i o - a c t i v i t 6 a c i d o - i n s o - l u b l e d o l t s 6 d i m e n t e r a v e c le cu lo t , p u i s q u e les p o l y s o m e s n e d o i v e n t p a s ~t re d i s s o c i 6 s d a n s ces
BIOCHIMIE, 1977, 59, n ° 3.
c o n d i t i o n s . P a r c o n t r e , d a n s le t a m p o n c o n t e n a n t de I 'EDTA, o n do l t s ' a t t e n d r e h la t r o u v e r d a n s le s u r n a g e a n t . Le t a b l e a u I I m o n t r e q u ' i l e n e s t b i e n a ins i . On p e u t d o n c e s t i m e r que la p l u s g r a n d e p o r t i o n du P o l y (A) RNA d u cu lo t P-90 p r o v i e n t des p a r t i c u l e s v i r a l e s d a n s les ce l lu les . U n e d e u x i ~ m e e x p 6 r i e n c e a c o n s i s t 6 h d ~ t e r m i n e r di- r e c t e m e n t le c o n t e n u en RNA d u cu lo t P-90 isol~ clans des c o n d i t i o n s s t a n d a r d , de ce l lu l e s D55. 0.7 p. c e n t du RNA c y t o p l a s m i q u e est l oca l i s6 d a n s P-90 et n ' e s t p a s r e t e n u s u r u n e c o l o n n e d ' o l i go dT-ce l l u lo se . Ce r ~ s u l t a t i n d i q u e que la f r a c t i o n P-90 de ce l lu l e s n o n i n f e c t ~ e s n e c o n t i e n t p a s de P o l y (A) RNA d a n s les s t r u c t u r e s qu i s ~ d i m e n t e n t d a n s ces c o n d i t i o n s .
TABLE IL
Dissociation des polgsomes dans les conditions exp~rimentales d'isolement da cytoplasme.
Mg~-+ (10-:~M)
EDTA (10-3M)
S-90
6490 cpm 37 p. cent
24090 cpm 88 p. cent
P-90
17050 cpm 63 p. eent
3285 epm 12 p. eent
200 million,s de cellules 4e sourls D55 Swiss ont ~t6 marqu6es pend, an t 40' mixmte.s pa r 20 ~ e / m l d 'u r id ine [3HI. Les cellul:es on.t 6t6 ,ensuite a jout6es h 200 mil- l ions de ceHules STU-2; luts ~g,aux ont 6t6 pr6pa~6s. Le premiea" lo t a 6t6 suspendu dans u n t a m p o ~ hypo- ' tonique Tris 0.01 M HC1, pH 8.2, Ae6tate &e Mg, 0.0,01 M, saeeharase', 5 p. cent. Le d,e.uxibm,e lot a 6t6 rcmis en suspension darts 1.e t a m p o n habi tueL Ponr parfai~re l'~elaLe,ment des ecllules du p remie r lot, on~ a a jout6 apr6s l 'homog~n6isa t ion dans le po t te r 1 p. een.t de N, onidet P40. Le evtopla, sme. des deux lots a 6t6 ensui fe h'aetion'n~ en S-fr0 et P-90. Les eulots ont ~td resus- pendu.s dans du t a m p o n NTE et d,es a l iquots de ehaque fraetion, ont 6t6 d6prot~ini~s~s pa r un t ra i t e rnen t SDS- Phenol . Les RNAs ont 6t6 ensu i t e p.r6eipit6s pa r l 'aeide t r ieRlorae6tique 10 p. cent ,ct filtr~s sur des mem- bra~nes Millipo~re 0..22 I~. Aprbs s~ehoge, on a immerg6 les memba'anes darts 3 nfl ~e solut ion scintill~a~ate e¢ on los a e ompt6es darts un eompteu~r Packard 3,390~. Les epm ~epr~sentent la to ta l i t6 d;e la radio-aet ivi t~ de ehaque fract ion.
3) D~termination de la taille des BNAs :
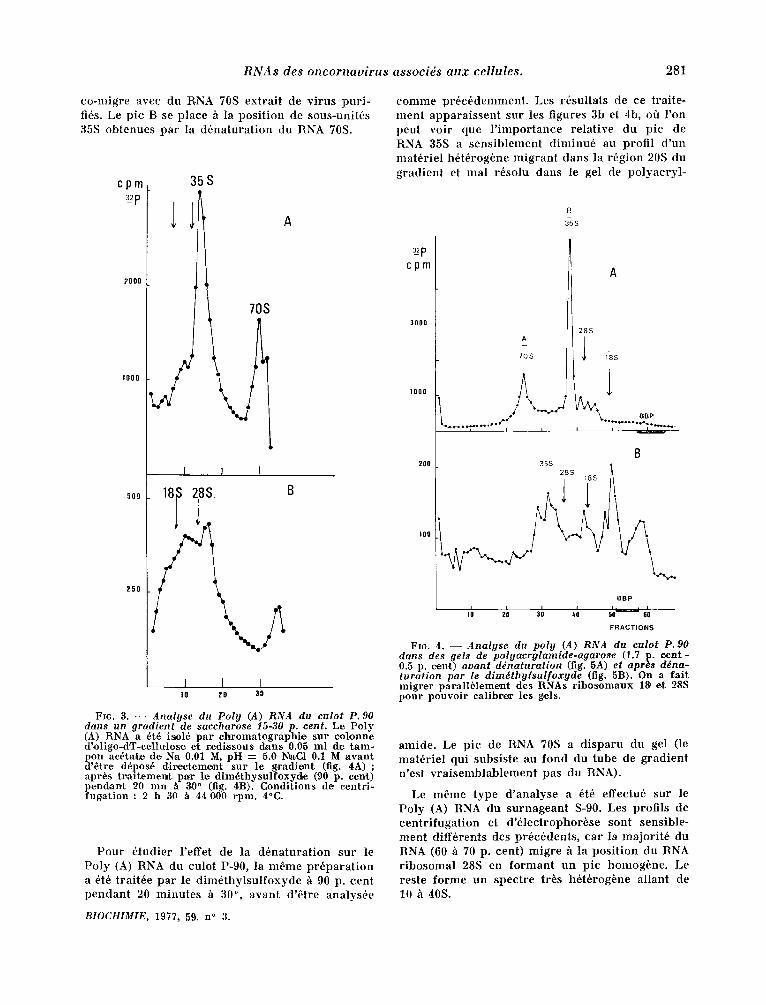
Les RNAs p o l y a d ~ n y l ~ s des f r a c t i o n s P-90 et S-90 on t ~t6 a n a l y s ~ s s u c c e s s i v e m e n t p a r u l t r a c e n - t r i f u g a t i o n d a n s u n g r a d i e n t de dens i t~ et p a r 61ec- t r o p h o r ~ s e en gel de p o l y a c r y l a m i d e . L a f igure 3a m o n t r e le p r o f i l d u g r a d i e n t de v~locitt~ d u P o l y (A) RNA e x t r a i t d u c u l o t P-90. On n o t e la p r e s e n c e de d e u x p i c s p r i n c i p a u x , d o n t les coeff i - c i e n t s de s ~ d i m e n t a t i o n e s t im~s p a r r a p p o r t h c e u x des RNAs 18 et 28:S a v o i s i n e n t 35 et 70S. Ce p r o f i l es t c o n f i r m ~ p a r les r ~ s u l t a t s de l ' ~ l e c t r o p h o r ~ s e en gel de p o l y a c r y l a m i d e - a g a r o s e (fig. 4a). Le p i c 1
R N A s d e s o n c o r n a v i r u s a s s o c i E s a u x c e l l u I e s . 281
c o - m i g r e avec du RNA 70S ex t r a i t de v i ru s pu r i - fi6s. Le p i c B se p l a c e h la p o s i t i o n de sous-uni t6s 35S ob t enues p a r la d 6 n a t u r a t i o n du RNA 70S.
cpm a2p
?O00
tO00
500
250
35S
1
70S
185 288 B
? I I I
10 20 30
FIG. 3. - - Analyse du Poly (.4) RNA du culot P. 90 dans un gradient de saccharose 15-30 p. cent. L~ Poly (A) RNA a 6t6 isol6 par ch~omatographie sur colonne d'oligo-dT-ee~lulos~ et r~Mi,ssous clans 0.05 ml d~ tam- pon ae6¢~te de Na 0.01 M, pH = 5.0 NraC1 0.1 M avant d 'etre d6pos6 d irec~ememt sur le 9radienL (fig. 4 A ) ; a pr6s tvaitemen¢~ par l~e dim6thysuJfoxy~e (90 p. cent) pendan.t 20 mn /t 9() ° (fig. 4B). Conditions d~ eentri- fug~atLon : 2~ h 30 ~ 44 0,0# rpm. 4°C.
P o u r 6,tudier l ' e f fe t de la d 6 n a t u r a t i o n su r le P o l y (A) RNA du culo t P-90, la m 6 m e p r 6 p a r a t i o n a 6t6 t ra i t6e p a r le d i m 6 t h y l s u l f o x y d e /~ 90 p. c e n t p e n d a n t 20 m i n u t e s h 3,0 °, avan t d '6 t re ana lys6e
c o m m e p r 6 c 6 d e m m e n t . Les r6su l ta t s de ce t ra i t e - m e n t a p p a r a i s s e n t su r les f igures 3b et 4b, off l ' o n peu t v o i r que l ' i m p o r t a n c e r e l a t i v e d u p i c de RNA 35S a s e n s i b l e m e n t d i m i n u 6 au prof i l d ' u n m a t 6 r i e l b6 t6rog6ne m i g r a n t dans la r6g ion 20S du g r a d i e n t et real r6so lu dans le gel de p o l y a c r y l -
B
£s
~P cpm
3000
1000
200
100
A
28S
7 0 S 18S
b
B
BBP
FRACTIONS
Fro. 4. - - Analyse du polg (A) RNA du culot P. 90 dans des gels de polyacry[amide-agarose (1.7 p. c en t - 0.5 p. cevt) aoant ddnaturat ion (fig. 5A) et apr~s d~na- turat ion par le d i m d t h y l s u l f o x y d e (fig. 5B). On a fai t migrer paral161ement ties RNAs r ibosomaux l& et 28S poux pouvoir c~alib~or les gels.
amide . Le p i c de RNA 70S a d i s p a r u du gel (le m a t 6 r i e l qu i subs is te au f o n d du tube de g r a d i e n t n ' e s t v r a i s e m b l a b l e m e n t pas du RNA).
Le m 6 m e t y p e d ' a n a l y s e a 6t6 effectu6 sur ]e P o l y (A) RNA du s u r n a g e a n t S-90. Les prof i l s de c e n t r i f u g a t i o n et d '61ec t rophorbse son t sens ib le - m e n t d i f f6 ren t s des p r6c6den t s , c a r la m a j o r i t 6 du RNA (60 h 70 p. cent) m i g r e h la p o s i t i o n du R N A r i b o s o m a l 28S en f o r m a n t un p i c h o m o g b n e . Le res te f o r m e un spec t r e tr6s h6 t6rog6ne a l lant de 10 /l 40S.
BIOCHIMIE, 1977, 59, n ° 3.
,982 M. L . M i c h e l et col l .
4) H g b r i d a t i o n des R N A s c y l o p l a s m i q u e s avec du c-DNA h o m o l o g u e du R N A du virus de F r i e n d :
La pr6sence de s6quences virales dans le RNA cytoplasmique peut 6tre sp6cif iquement recher - ch6e en hybr idan t le RNA contre du c-DNA repr6- sentant une copie cor rec te du g6nome viral .
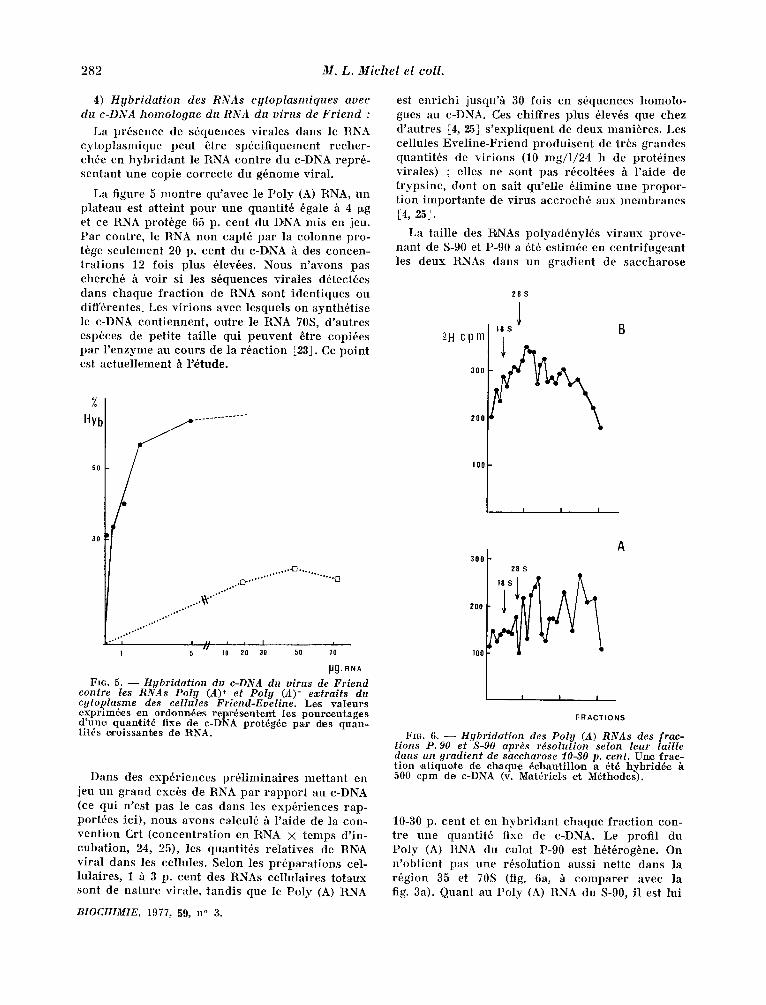
La figure 5 mont re qu 'avec le Poly (A) RNA, un plateau est atteint pour une quantit6 6gale /t 4 ~g et ce RNA prot6ge 65 p. cent du D'NA mis en jeu. Par contre, le R'NA non capt6 par la colonne pro- t6ge seulement 20 p. cent du c-DNA h des concen- trat ions 12 fois plus 61ev6es. Nous n 'avons pas cherch6 h voi r si les s6quences virales d6tect6es dans chaque f rac t ion de R, NA sont ident iques ou diff6rentes. Les v i r ions avec lesquels on synth6tise le c-DNA cont iennent , outre le RNA 70S, d 'autres espbces de peti te tai l le qui peuvent ~tre copi6es par l ' enzyme au cours de la r6act ion [23]. Ce point est ac tuel lement h r6tude.
7~
ttyb~o ~ ..............
30
o..,E].o. .o0.O, " ' '° ' '°° . . . . " ° ' " 0
• "; ........................ i'"// . . . . I 5 1O 20 310 50 70 pg..NA
FIG. 5. - - Hybridation du c-DNA du virus de Friend contre les RNAs Polg (A) + ef Polg (A)- extraits du cytoplasme des eellules Friend-Eveline. Les vale~rs exprim&es vn ord.onn&es rel~r~sente~t les p,ouroentages d'une quan*tit~ fixe de c-DNA prat~g~e pa,r des quan- titds c~oissantes de I~NA.
Dans des exp6riences pr61iminaires mettant en jeu un grand exc6s de RNA par r appor t au c-DNA (ce qui n 'est pas le cas dans les exp6mences rap- port6es ici), nous avous calcul6 ~ l 'a ide de ]a con- vent ion Crt (concent ra t ion en RNA × temps d ' in- cubation, 24, 25), les quantit6s relat ives de RNA vira l darts les cellules. Selon les pr6para t ions cel- lulaires, 1 h 3 p. cent des BNAs cel lulaires totaux sont de nature virale, tandis que le Poly (A) I~NA
BIOCHIMIE, 1977, 59, n ° 3.
est enr ich i jusqu'h 30 fois en s6quences homolo- gues au c~DNA. Ces chiffres plus 61ev6s que chez d 'autres [4, 25] s ' exp l iquen t de deux mani6res. Les cellules Eve l ine -Fr i end produisen t de tr6s grandes quantit~s de v i r ions (10 rag / l /24 h de prot6ines virales) ; elles ne sont pas r6colt6es ~ l 'a ide de t rypsine, dont on sait qu 'el le 61imine une p ropor - t ion impor tan te de virus accroch6 aux membranes N, 25].
La taille des I~NAs polyad6nyl6s v i raux prove- nant de S-90 et P-90 a 6t6 estim6e en cent r i fugeant les deux BNAs dans un gradient de saccharose
_3H cpm 300
2 0 0
I 0 0
28S
1 8 S
I I
300
200
I00
28 S
A
t I I
FRACTIONS
FIG. 6. - - Hybridation des Poly (A) RNAs des frac- tions P. 90 et S-90 apr~s r~solution selon leur faille dans un gradient de saccharose 10-30 p. cent. Une frac- tion .aliquote de chaque ~cha~till.en a ~t~ hybrid~e & 500 cpm de c-DNA (v. M~at~r~els et M~thodes).
10-30 p. cent et en hyb r idan t chaque f rac t ion con- tre une quantit6 fixe de c-DNA. Le profi l du Poly (A) RNA du culot P-90 est h6t6rogbne. On n 'obt ient pas une r~solution aussi nette dans la r6gion 35 et 70S (fig. 6a, ~ compare r avec la fig. 3a). Quant au Poly (A) RNA du S-90, il est lui

R N A s des o n c o r n a v i r u s associds a u x celIules. 283
aussi tr~s h6t6rog~ne avec un pic large centr6 sur la r6gion 28-30S (fig. 6b). Ce mat6riel correspond vra i semblablement aux RNAs messagers v i raux d6tach6s des polysomes au cours de la pr6para t ion de l 'extrai t cytoplasmique.
5) Discussion :
Ce travai l a 6t6 entrepr is dans le but d ' isoler dans des cellules infect6es par un oncornav i rus et le p roduisan t en permanence , les seuls RNAs des part icules virales en vote de format ion dans les cellules. En outre, nous d6sirions obtenir des quanti t6s pond6rales de ces RNAs afin de pouvoir les ut i l iser dans des exp6riences de t raduc t ion in oitro. Pour at te indre ces buts, il 6tait n6cessaire de rempl i r au moins deux condi t ions :
a) isoler une fract ion cellulaire dans laquelle seraient concentr6es toutes les part icules h divers moments de leur synthbse ;
b) 6tablir des condi t ions exp6rimentales aptes h 61trainer les RNAs viraux impliqu6s dans d'au- tres 6tapes du m6tabolisme viral et en par t icu l ie r les RNAs messagers li6s aux ribosomes.
Pour satisfaire h la premibre condi t ion, nous avons centrifug6 le cytoplasme h des vitesses telles que les cores puissent s6dimenter au fond du tube au cas oh ils existeraient h l '6tat l ibre dans les cel- lules. Dans les premi6res exp6.riences, r ex t ra i t cytoplasmique 6tail d6barrass6 des mi tochondr ies par une centr i fugat ion in term6dia i re ~t 10.00~0 rpm. Nous l 'avons abandonn6e par la suite car le rende- ment en vi r ions 6tail sens ib lement moindre en rai- son de l '61imination concommit tan te de fragments de membranes et de part icules qui leur sont atta- ch6es. L 'observat ion microscopique des coupes pr6par6es "h par t i r des culots de centr i fugat ion 90.0.00 g mont re b ien l 'existence de structures ayant les caract~res de v i r ions h diff6rents stades de leur formation. Le fail qu 'un grand nombre d 'entre elles demeurent attach6es aux membranes explique que nos essais de puri,fication suppl6men- taire n 'on t pas abouti. La bande de mat6riel de densit6 1,12- 1,14 obtenue par centr i fugat ion dans un gradient de saccharose 10-70 p. cent cont ient la quasi totalit6 des part icules virales de la cel- lule, ainsi qu 'en t6moigne le profil d 'activit6 trans- criptase inverse, mats la microscopie 61ectronique mont re la pers is tance d 'une propor t ion tr~s 6levee d'616ments cellulaires. La sonicat ion du cytoplas- me, qui, dans les mains de Panem et coll., semblai t l ib6rer les cores ne nous a pas donn6 de r6sultats satisfaisants car nous n 'avons pas retrouv6 dans les zones de densit6s 1,20 et 1,24 de bande conte- nan t une activit6 enzymat ique notable.
Le culot P-90 cont ient moths de 10 p. cent des RNAs cytoplasmiques, mats un peu plus de 20 p.
cent des RNAs polyad6nyl6s totaux. Les analyses dans des gels de po lyacry lamide et en gradient de saccharose ind iquen t que ce Poly (A) RNA con- t ient essentiel lement deux espbces homogbnes 70 et 35S. Puisque le culot P-90 est trbs h6t6rogbne, ces espbces pour ra ien t th6or iquement p roven i r des part icules vira]es des mi tochondr ies et des polysomes. S'il est facile d'61iminer ies mi tochon- dries, qui ne con t i ennen t pas de Poly (A) RNA de cette taille, il 6tail impor tan t de mont re r que les polysomes avaient 6t6 dissoci6s par nos condi t ions exp6rimentales. Une exp6rience de recons t ruc t ion , off nous avons suivi la rbpar t i t ion d 'u r id ine [aH] incorpor6e pendan t une courte p6riode de temps dans les mRNAs de cellules non infect6es nous a montr6 que le pourcentage de radio-activit6 acido- pr6cipi table n 'exc~de pas 12 p. cent dans le culot P-90, alors qu'elle at teint 03 p. cent dans le culot pr6par6 en pr6sence d ' ions Mg ++. Ce r6sultat ne signifie pas que toute la radioactivi t6 du culot est incorpor6e dans du Poly (A) PrimA. En tout 6tat de cause, il ind ique que la f ract ion du Poly (A) RNA qui pour ra i t p roven i r des polysomes n'exc~de pas 12 p. cent.
La nature virale du Poly (A) 1LNA du culot P-90 6tait d6jh fortement sugg6r6e par la taille des espb- ces qu' i l contient. L 'hybr ida t ion avec du c-DNA homologue du g6nome vira l confirme l 'exis tence de mat6riel s6dimentant dans la r6gion 35 et 70S. Toutefois, le profil d 'hybr ides est loin d'6tre mo- nodisperse. I1 en va de m6me pour le profil obtenu avec le Poly (A) RNA de S-90..Ces r6sultats qui ont 6t6 rapport6s par d 'autres auteurs [4, 26], sont en cont rad ic t ion avec ceux des gels de polyacryl- amide et des gradients de saccharose, oh / 'on ob- serve ~ l ' inverse une grande homog6n6it6. Les con- centra t ions raises en jeu dans les exp6riences d 'hy- b r ida t ion 6talent plus de 20 fois sup6.rieures h celles des gels et des gradients pour lesquels on avail utilis6 le marquage par le 3~p. I1 est possible que des ph6nombnes d 'agr6gation se produisent , qui alt6rent la d is t r ibut ion r6elle des nlol6cules dans les gradients. La pr6sence de tels agr6gats dans les RNAs viraux du noyau a 6t6 signal6e dans un trbs r6cent t ravai l [27].
L'espbce virale majeure pr6sente dans ]es part i- cules naissantes ou en vote de format ion est un RNA 35,S qui a la mC, lne taille apparente que les sous-unit6s r6sul tant de ]a d6natura t ion du g6nome des part icules extracellulaires. Il est tr~s probable que ce RNA est la sous-unit6 elle-m6me, ce qui im- pl ique que le g6nome des virus naissants n 'est pas encore sous la forme d 'un complexe 70S. Chez ]es vi rus aviaires, le g6nome de virus r6colt6s tr~s pr6cocement apr~s leur l ib6rat ion dans le mil ieu
BIOCHIMIE, 1977, 59, n ° 3.

2 8 4 M. L . M i c h e l e t co l l .
de c u l t u r e es t c o m p o s 6 de RN A 30-40S q u i se c o n - v e r t i t e n s u i t e en RN A 70S [28]. D a n s le v i r u s leu- c 6 m o g 6 n e du cha t , u n e p r o p o r t i o n n o t a b l e du g6- n o m e v i r a l est u n e e sp6ce 30-35S [29]. Des p h 6 n o - m 6 n e s de r 6 a r r a n g e m e n t on t 6t& d 6 c r i t s p o u r le v i r u s d u s a r c o m e m u r i n [30]. Nos r 6 s u l t a t s s ' a c - c o r d e n t p a r f a i t e m e n t a v e c ces t r a v a u x , s i ] ' on c o n - s i d 6 r e que les p a r t i c u l e s v i r a l e s c o n t e n n e s d a n s P-90 c o r r e s p o n d e n t h des v i r i o n s c o m p l e t s et na i s - s a n t s et des p r 6 c u r s e u r s de ces v i r i o n s , te ls q u ' i l s a p p a r a i s s e n t sous la f o r m e de s t r u c t u r e s i d e n t i f i a - b l e s e n m i c r o s c o p i e 61ec t ron ique . Nos r 6 s u l t a t s ne n o u s p e r m e t t e n t p a s de d i r e s i d ' a u t r e s s t r u c t u r e s c o n t e n a n t d u R N A v i r a l e x i s t e n t d a n s la ce l lu le , p u i s q u e n o u s n ' a v o n s p a s les m o y e n s de c a r a c t 6 - r i s e r . E n p a r t i c u l i e r , i l se p o u r r a i t q u e le S-90 c o n - t i e n n e e n p l u s des ]~N,A m e s s a g e r s v i r a u x des r i b o - n u c l 6 o p r o t 6 i n e s de f a i b l e t a i l l e qu i p o u r r a i e n t 6 t r e so i l des f o r m e s de t r a n s f e r t d u RNA v i r a l d u n o y a u au c y t o p l a s m e , so i l des p r 6 c u r s e u r s t r6s p r 6 c o c e s des v i r i o n s .
P o u r ce qu i es t d u R N A 70S c o e x i s t a n t a v e c le RNA 35S, d a n s le cu lo t P-90, i l p r o v i e n t t r6s v r a i - s e m b l a b l e m e n t de p a r t i c u l e s v i r a l e s p lu s 5g6es de- m e u r 6 e s fix&es a u x ce ] lu les ou r e t r o u v 6 e s d a n s les v a c u o l e s (Vo i r la f igu re 1).
U n e p o r t i o n i m p o r t a n t e d u P o l y (A) I~NA d u cu lo t P-90 es t c o n v e r t i e p a r le d i m 6 t h y l s u l f o x y d e en molGcu les hGtGrogbnes de t a i l l e p lu s pe t i t e , n i l - g r a n t d a n s la zone 2~)-22:S d u g r a d i e n t de s a c c h a - rose . Ces rGsul ta t s n e p e r m e t t e n t p a s de d i r e s ' i l s ' ag i t de molGcu les p r G e x i s t a n t e s s e m b l a b l e s "~ ce l l es qu i e x i s t e n t s u r les p o l y s o m e s liGs [4, 6, 7] , o u de d G g r a d a t i o n s a p p a r a i s s a n t au c o u r s de l ' i so - l e m e n t des p a r t i c u l e s c e l l u l a i r e s . C e r t a i n s a u t e u r s o n t s i gna l6 la p r G s e n c e de molGcu les de ce t t e t a i l l e d a n s le RNA de p a r t i c u l e s e x t r a - c e l l u l a i r e s a p r b s d & n a t u r a t i o n [31]. S e l o n d ' a u t r e s , la d G n a t u r a t i o n n ' e n g e n d r e que les sous -un i tGs 35S [5]. I1 s e r a i t i n t G r e s s a n t de c o m p a r e r ces RNAs a v e c l ' espGce 20S t rouvGe s u r ]es p o l y s o m e s liGs.
Remerc iements .
No~.s sommes trGs re~o.nnaissants au Profe,sseur Sch~ifer (Tubingen) qui no,us a donn6 la lignGe cel lu]aire F,riend-Evclin.e. Ce tra~a~il a bGnGfiei6 d.e l'aide, f inan- eibre d'e I'IN'SERM (Coatra t libce 76.5.128.2.) ef d 'une subven t ion d,u Commissari.a~t h l'En.ergie Atomiqt~e pour les radio-61~m, ents.
Nous teno.ns h remeroi,er M mo Mauchauff6 pour sa eol- ]aborati ,on teehniqu,e.
R~SVM~.
Le cytoplasme, de c,ellu~es de souris STU produisan,t e~ pern~an,eaaee 1~ v i rus d,c F r i end con.fluent dt~s par t i - cul,~s viral,es h diff6rents statics de l'e.ur d6veloppement . I1 e s t possible de l ib6rer ees particul~es en f a i s an t
BIOCHIMIE, 1977, 59, n ° 3.
6claimer 1.es eell ules dans un t ampon hypotonique . Apr6s s6dimcIrtabion h 90000 g, la tr6s grantee major i t6 des pa r t i cu les viral.~s ,est concen~tr6e dans 1,e culot, a ins i que ]~e: montren¢ ]a microscopie 61ectronique et la ca,ract6ris'a¢ion de. l 'act ivi t4 t ranscr ip tase inv,erse. Les Poly (A) RNAs de ee cul,ot conti:ennent pr incipa~ement 2 esp6ctes mol6cula i res 35 S e:t 70 S. L 'hybr ida t ion dc ee Poly (A) RNA h du c-DNA homologtre du g6nome vira l rdv61,e 6~ale.~r~nt ]a prise,nee de 2 RNAs 35 S et 70 S ain,si qwe d'a.utres ,e;sp6ees qui sorer pe,ut-~tre des agr6gats. Pour 6viter l',iaaterfdreaaee des RNAs messa- gers vir~nx, nons avons uti l is6 des condi.tions exp6- r ime n~a~es qui p e,rmet~en~ ]a dissociat ion des poly- somes ,el empdchent leur passage dar~s le eulot de cen t r i fuga t ion h 90 O00 g.
REFERENCES,
1. B.~rnhardt, W. (1958) Cancer Res., 18, 491-501, 2. Duesberg, P. H. (196.8) Proc. Nat. Ac. Sei. USA,
60, 151:1. 3. King, A. M. Q. (1976) J. Biol. Chem., 251, 141-149. 4. Fan, H. a B~ltimove, D. (1973) J. MoL Biol., 80,
93-117. 5. Tsuchida, N. ,~ Gree.n, M. (1974) Virology, 59, 258-
265. 6. Shanmugan , G., B h,~duri, M. ~ Green, M. (1974)
Biophem. Biophys . Bes. Commun., 56, 697-702. 7. Gi,ell~ens, L., Salde~l, M. H. ~ Blo,em'endal, H. (1974)
Proe. Nat. Ac. Sci. USA, 71, 1093-1097. 8. Gi.elk,en,s, A. L. J., \5~n Za:an.e, D., Bloem~rs~ H. P. I.
Bloemendal , H. (1976) Proc. Nat. Ac. Sci. USA, 73, 356-360.
9. Watson, J., Ralph, P., Sarka~-, S. a Cohn., M. (1970) Proe. Nat. Ac. Sci. USA, 66, 344-351.
10. Pan~em, S. a Kirsten, W. H. (1973) d. Nat. Cancer Inst., 51, 866-874.
11. L at a Duesb~rg, P. H. (1972) Nature, 235, 38~3-386. 12. Seol,niek, E. M., Benvenis.~e, R. a Parks, E. W. (1973)
d. Virol., 11, 600-603. 13. Seifert, E., Claviez, M., Frank, H., H~lnsmann, G.,
Sehw.arz, H. ~ Sel~af, e.r, W. (1975) Z. Na lur forsch., 30 c, 64~8-700.
14. Gal,ibevt, F., Larsen, C. J., Lelong, J. C. a Bolron, M. (1965) Nature, 207, 1939'-1941.
15. Tiollais, P., Galibert , F. Leoetit , A. a Auger, M. A. (1972), Biochimie, 54, '339"-354.
16. GaIlo, P~. C., Mil, l,e,r, R. N,, Saxi,n~er & GiIl,~spie~ D. (1973) Proc. Nat. Ac. Sei. USA, 70, 3219-3223.
17. Taviti.an A,, Hamelin., R., Tehen, P., Olofsson, B. Boiron, M. (197~4), Proc. Nat. Ae. Sei. USA, 71, 75'5-754.
18. Rothenbrerg, E. a BaLtimore, D. (1976)J . Virol., 17, 16,8-174,
19. Junghans , R, P., Dlaesb~rg, P. H. s~ Krfight, C. A. (1975) Proe. Nat. Ae. Sei. USA, 72, 4805-4899.
20. G arapin , A. G., Vaemus; ~. E., Far~s, A. J., Levin- son, W. E. a Bishop, J. M. (1973,) Virology, 52, 26:4-274.
21, La~sen, C. J., Marry, M., ~amc l in , R., I~eri~es, J., Boiro~n, M. a Tavi t ian , A. (1975) Proc. NaL Ae. Set. USA, 72, 49~)0-4904.
22. Sutton, W. D. (1971) Bioehim. Biophys . Aeta, 940, 522-531.
23. G a,~avin, A. C. (19'73,) Bull . Ins t . Pasteur, 71, 185- ~3,2.
24. Birnsti.e], M. L., Sells, B. H. ~ Purd.om, I. F. (1972) J. Mol. Biol., 63, 2,1-39.
26,. Le,ong, J. A., C~arapi,n, A. C., Jackson, N., Fansh ie r , L., Levinson, W. a Bishop, N. (1972) J. Virol., 9, 891-902.

R N A s des o n c o r n a v i r u s a s s o c i d s a u x c e h u l e s . 285
26. Okab.e, H., Gilden, R. V. & H~atanaka, M. (1973) J. Virol., 12, 984-995.
27. Has,elti~ne, W. A. & Baltimore, D. (1976) J. Virol., 19, 331-337.
28. Canaani, E., Van d~er H.elm, K. V. S. & Du.~sberg, P. H. (1973) Proc. Nat. Ac. Sci. USA, 72, 401-405.
29. Jal-retts, O., P itts, J. D., Whalley, J. M., Cla.son, A. E. & M,a~y, J. (1971) Virology, 43, 317-320.
3~}. East, J. L., KrLesek, J. E., Chan, J. C., Bowen, J. M. & Dmochowski, L. (1973) J. Virol., 11, 709-720.
31. Riggin, C. H., Bondurant , M. C. a Mitetrell, W. (1973/74) Intervirolooy, 2, 209-~21.
BIOCHIMIE, 1977, 59, n ° 3.





























