Embed Size (px)
Citation preview

Journal ofNeurochemistryLippincott—Raven Publishers, Philadelphia© 1998 International Society for Neurochemistry
Selective Increase of a2A-Adrenoceptor Agonist Binding Sitesin Brains of Depressed Suicide Victims
Luis F. Callado, J. Javier Meana, *Bernardo Grijalba, *Angel Pazos,~Magda1ena Sastre, and tJesüs A. GarcIa-Sevilla
Department of Pharmacology, University of the Basque Country, Leioa, Bizkaia; *Department of Physiology andPharmacology, University of Cantabria, Santander; and tLaboratory of Neuropharmacology,
Department of Biology, University of the Balearic Islands, Palma de Mallorca, Spain
Abstract: The a2A- and ct2~-adrenoceptorsubtypes wereevaluated in postmortem brains from suicides with de-pression (n = 22), suicides with other diagnoses (n= 12), and controls (n = 26). Membrane assays withthe antagonist [
3H]RX821 002 (2- [3H]methoxyidazoxan)suggested the presence of a
2A-adrenoceptors in the fron-tal cortex and both a2c-adrenoceptors and a2A-adreno-ceptors in the caudate. The proportions in caudate weresimilar in controls (a2A, 86%; a2c, 14%), depressed sui-cides (a2A, 91%; a2c, 9%), and suicides with other diag-noses (a2A, 88%; a2c, 12%). Autoradiography of [
3H]-RX821002 binding under a2B/C-adrenoceptor-maskingconditions confirmed the similar densities of a
2A-adreno-ceptors in the cortex, hippocampus, and striatum fromcontrols and suicides. In the frontal cortex of depressedsuicides, competition of [
3H]RX821002binding by (—)-
adrenaline revealed a greater proportion (61 ± 9%) ofa
2A-adrenoceptors in the high-affinity conformation foragonists than in controls (39 ±5%). Simultaneous analy-sis with the agonists [
3H]clonidineand [3H]UK14304 andthe antagonist [3H]RX821002in the same depressed sui-cides confirmed the enhanced a
2A-adrenoceptor densitywhen evaluated by agonist, but not by antagonist, radioli-gands. The results indicate that depression is associatedwith a selective increase in the high-affinity conformationof the brain a2A-adrenoceptors. Key Words: a2-Adreno-ceptor subtypes—Depression— Human brain—Suicide—Frontal cortex.J. Neurochem. 70, 1114—1123 (1998).
A variety of studies have focused on specific alter-ations of monoamine receptors as a mechanism in-volved in the pathogenesis of depression (Caldecott-Hazard et a!., 1991; Cheetham et al., 1991). Amongothers, evidence of changes in the sensitivity of a2-adrenoceptors has been accumulated (for review, seePiletz and Halaris, 1994). In the CNS, a2-adrenocep-tors play an importantphysiological role as autorecep-tors on noradrenergic neurons (Starke and Montel,1973; Svensson et a!., 1975) and as heteroreceptorson serotonergic terminals (Göthert and Huth, 1980).
Several studies performed in platelets of depressed pa-tients have shown an increased a2-adrenoceptor den-sity (GarcIa-Sevilla et al., 1981, 1987; Doyle et al.,1985; Piletz and Halaris, 1988; Piletz et al., 1990,1991; for review, see Piletz and Halaris, 1994) associ-ated with an enhanced receptor sensitivity (GarcIa-Sevilla et al., 1986, 1990; Karege et al., 1996). In thiscontext, it has been also demonstrated that a varietyof antidepressant drug treatments, as well as electro-convulsive therapy, decrease and/or reverse the en-hanced density and sensitivity of platelet a2-adreno-ceptors in depressed patients (GarcIa-Sevilla et al.,1981, 1986, 1987, 1990; Piletz and Halaris, 1988; Pan-dey et al., 1989; Karege et al., 1992; Piletz et al.,1996). In the rat CNS, several antidepressant treat-ments, when given chronically, reduce the density andactivity of a2-adrenoceptors (Smith et al., 1981; No-mura et al., 1987; Giralt and GarcIa-Sevilla, 1989;Menargues et al., 1990; Heal et al., 1991; Mongeau etal., 1994). Such findings, which have been demon-strated by several but not all the authors, are in keepingwith the hypothesis that postulates an enhanced a2-adrenoceptor activity in the pathogenesis of some typesof depression.
Postmortem studies in human brain have confirmedan enhancement of a2-adrenoceptor density in de-pressed suicide victims when labeled with agonist ra-dioligands (Meana and GarcIa-Sevilla, 1987; Meanaet al., 1992; Gonzalez et al., 1994; Ordway et al.,1994). In contrast, in studies conducted with antago-nist radioligands, the alteration has not been observed
Received May 19, 1997; revised manuscript received September22, 1997; accepted October 6, 1997.
Address correspondence and reprint requests to Dr. J. JavierMeana at Departamento de Farmacologla, Universidad del PalsVascoIEuskal Herriko Unibertsitatea, E-48940 Leioa, Bizkaia,Spain.
Abbreviations used: ARC239, 2- [2- [4- (o-methoxyphenyl)piper-azin- l-yl]ethyl] -4,4-dimethyl-1,3-(2H,4H) -isoquinolindione HC1;DSM-III-R, Diagnostic and Statistical Manual ofMental Disorders,third edition, Revised; 5-HT, 5-hydroxytryptamine.
1114

a2-ADRENOCEPTORS IN DEPRESSION 1115
(Crow et al., 1984; Ferrier et a!., 1986; Ordway et al.,1994). Thus, the findings are suggestive of an in-creased proportion of the high-affinity conformationof the receptors (Asakura et a!., 1985). However, thedirect involvement of the high-affinity state of the a2-adrenoceptor in depressionand/or suicide has notbeenclarified as yet.
Molecular and pharmacological evidence supportsthe existence of multiple a2-adrenoceptor subtypes(Bylund, 1992). The a2A-adrenoceptor is the exclusivesubtype in platelets (Bylund et a!., 1988) and predomi-nates in the human brain with a preferential locationincortex, hippocampus, and cerebellum (DeVos et al.,1992; Ordway et al., 1993; Sastre and Garcia-Sevilla,1994a; Grijalba et a!., 1996). The a2c-adrenoceptorsubtype appears to be expressed in human brain areassuch as the caudate nucleus and hippocampus (De Voset al., 1992; Ordway et al., 1993; Sastre and GarcIa-Sevilla, 1994a; Grijalba et a!., 1996). On the basis ofthe previous evidence in platelets and postmortembrain, depression might be associated with a selectiveenhanced activity of the a2A-adrenoceptor subtype.
In the present study, the density and anatomical dis-tribution of a2-adrenoceptor subtypes were assessed inselected brain areas from control subjects and suicidevictims withdepression or other psychiatric diagnoses.The biochemical status of the high-affinity conforma-tion of the a2A-adrenoceptor subtype in depressed sui-cides was also evaluated to determine whether its pos-tulated supersensitivity in depression could result froman increased density of the high-affinity state of thereceptor.
EXPERIMENTAL PROCEDURES
Brain samplesHuman brains were obtained from 60 subjects who had
died mainly as a result of suicide or other violent causes.Samples were collected at autopsy over the same periodof time (from January 1988 to July 1994) in the InstitutoAnatómico Forense, Bilbao and Palma de Mallorca, Spain.The collection was performed in accordance with an ap-proved protocol of the Instituto Anatómico Forense for post-mortem human studies. All the deaths were subjected toretrospective careful searching for previous medical diagno-sis and treatment using the medical examiner’s informationand records of general and psychiatric hospitals. Subjectswithout evidence of neurological or psychiatric disorder andwho also died by a nonsuicide mechanism were ascribed tothe control groups. Toxicological screening for psychotropicdrugs and alcohol was carried out on tissue samples. Controlsubjects with a positive test were excluded from the study.
Three separate groups of experiments were performed us-ing different human brain specimens. In the first group,which was directed to evaluate the a2-adrenoceptor subtypepopulations in brain membranes, 10 control subjects and 10suicide victims with DSM-III-R criteria for recurrent depres-sion (n = 6) or bipolar disorder (n = 4) (American Psychiat-ric Association, 1987) were included. Ten suicide victimswith other diagnoses (five schizophrenia, two paranoid disor-der, one anxiety disorder, one neurotic disorder, and one
TABLE 1. Demographic characteristics and postmortdelays of control subjects and suicide victims
em
PostmortemGender Age delay(FJM) (years) (hours) n
Group IControls 5F/5M 48 ±6 34 ±6 10Major affective disorders 7F/3M 54 ±6 39 ±6 10Other diagnoses 3F/7M 51 ±6 39 ±5 10
Group IIControls 5F/2M 55 ±7 21 ±5 7Suicide victims 8F/1M 52 ±6 36 ±6 9
Group IIIControls 4F/5M 48 ±7 32 ±4 9Major affective disorders 4F/5M 50 ±6 37 ±5 9
Causes of death were as follows: jumping from a height (n =
17); motor-vehicle accident (n = 10); hanging (n = 8); accidentalfall (n = 5); neoplasia (n = 4); cardiac arrest (n = 4); drowning (n= 3); labor accident (n = 2); motor-vehicle suicide (n = 1); causticintoxication (n = 1); gunshot wound (n = 1); brain hemorrhage (n= 1); aortic rupture (n = 1); homicide (n = U; and psychotropicintoxication (n = I). Group values are means ±SEM.
reactive depression) were also added to the group (Table 1,group I). Medical records revealed that four subjects wereon antidepressant treatment (clomipramine, amitriptyline,imipramine, maprotiline, and electroconvulsive therapy).Other psychotropic medications in thegroup includedanxio-lytics (n = 10), neuroleptics (n = 9), anticholinergic drugs(n = 4), and lithium (n = 2). All the controls were freeof medical diagnoses and treatments. The second group ofexperiments was aimed at evaluating the autoradiographicaldistribution of a2-adrenoceptor subtypes in depression. Thisgroup included seven control subjects and nine suicide vic-tims, seven of them with firm criteria for “major affectivedisorder” (five recurrent depression, two bipolar disorder)and two other suicide victims who did not meet full criteriafor a diagnosis of depression or bipolar disorder (Table 1,group II). The medical records of this group II showed thepresence of antidepressants (n = 3), neuroleptics (n = 3),anxiolytics (n = 3), anticholinergic drugs (n = 1), andl4thium (n = 1) among the suicide victims. This group sharedthree suicide victims with group I. In a third experimentalapproach to evaluate the high-affinity conformation of thea2A-adrenoceptorsubtype, nine control subjects and nine sui-cide victims with firm diagnosis of “major affective disor-der” (seven recurrent depressive disorder and two bipolardisorder) were included (Table 1, group III). Seven casesof the suicide sample in this group were on antidepressanttreatment (clomipramine, amitriptyline, fluoxetine, fiuvox-amine, mianserine). Other treatments were benzodiazepines(n = 2), neuroleptics (n = 2), and lithium (n = 1). Onesubject in the suicide sample of this group III was sharedwith group I. The demographic characteristics (age, gender,postmortem delay, andcause of death) of the various controland suicide groups are shown in Table 1 and did not differwithin groups.
Brain areas were dissected at the time of autopsy andimmediately stored at —70°C.Freezing and storage periods(20 ±4 months, range 1—54 months) were similar in controland suicide groups. For the binding studies in homogenates,P2 fractions of membranes were isolated from prefrontal cor-
i. Neurochem., vol. 70, No. 3, 1998

1116 L. F. CALLADO ET AL.
tex (Brodmann’s area 9) and caudate head and were resus-pended in incubation buffer (50 mM Tris-HC1, pH 7.5) aspreviously described (Meana et al., 1989). Protein contentswere quantified by the method of Lowry et al. (1951) andwere similar in the brains of suicide victims (range 510—1,020 ~ig/ml) and control subjects (range 560—1,070 ,uglml). For autoradiography, l0-~.tmsections from frontal cor-tex (Brodmann’ s areas 8 and 9), hippocampus, and striatumwere cut at —22°C using a cryostat, mounted on gelatin-coated slides, and stored at —25°C(Gonzalez et al., 1994).
Radioligand binding assays in homogenatesBinding experiments were performed as described pre-
viously (Sastre and GarcIa-Sevi!!a, 1994b; Grijalba et a!.,1996). Radioligand binding assays were done by incubating,for 30 mm ([3H]RX821002 and [3Hlclonidine) or for 60mm ([3H]UK14304) at 25°Cwith shaking, 500 p3 of mem-brane preparation, 10 bel of radioligand, and 40 p3 of eitherincubation buffer (50 mM Tris-HC!, pH 7.5) or drugs. Theassay buffer for the 3H-labeled agonists included 10 mMMgC1
2 (for [3H]clonidine) or 0.1 mM MnC1
2 (for [3H]-
UK14304) to express the high-affinity conformation of thereceptors fully (Sastreand GarcIa-Sevilla, 1 994b). In satura-tion and competition studies against the mixed a
2A,B/C-adre-noceptor antagonist [
3HI RX82 1002, brain membranes werealso incubated in the presence of 0.3
1tM 5-hydroxytrypta-mine (5-HT) to block the binding of the radioligand to 5-HT-sensitive binding sites (Vauquelin et al., 1990; Grijalbaet a!., 1996). In saturation studies, the final concentration ofradioligand (nine points) ranged from 0.03 nM to 8 nM([
3H] RX821002 and [3H] UK14304 binding assays) or from0.06 nM to 16 nM ([3Hlclonidine binding assays). Drugcompetition studies were performed with I nM [3H]-RX82 1002 in the absence or presence of various concentra-tions of competing drugs (1012_ l0~ M; 19 concentra-tions). Nonspecific binding, as estimated in the presence of10 pM (—)-adrenaline, ranged from 5% to 30% and wassimilar in the various control and suicide groups.
Incubations were terminated by dilution and rapid filtra-tion under vacuum through Whatman GF/C glass-fiber fil-ters that had been presoaked with 0.5% polyethylenimine.Then the filters were rinsed twice with 10 ml of ice-coldincubation buffer, air-dried, transferred to minivials con-taining 5 ml of OptiPhase HiSafe II cocktail (Wallac, U.K.),and counted for radioactivity by liquid scintillation spec-trometry.
Radioligand binding assays in tissue sectionsThe autoradiographic labeling was performed as described
previously (Grijalba et al., 1996). After a30-mm preincuba-tion at roomtemperature, tissue sections were incubatedwith5 nM [3H]RX821002 for 30 mm in the 50 mM Tris-HC1buffer containing 1 mM MgCl
2 and 1 g/L ascorbic acid, pH7.4, also at room temperature. After the incubation, sectionswere washed twice for 20 s in ice-cold buffer, dipped in ice-cold water, and then dried under acold air stream. Typically,two consecutive sections from each brain and level wereincubated: one in the presence of ARC239 (20 nM) fordetermination of the total binding and one in the presenceof ARC239 (20 nM) plus 10 ,uM (—) -adrenaline for thenonspecific binding. The drug ARC239, a selectivea2~,~-adrenoceptorantagonist with 5-HTIA receptor agonistproperties (Esteban et al., 1996; Meana et al., 1996), andthe concentration used (20 nM) were chosen to evaluate thespecific binding of li
3H] RX82 1002 to the a2A-adrenoceptor
subtype (Grijalba et al., 1996). Nonspecific binding rangedfrom 5% to 10% and was similar in the control and suicidegroups. Sections were exposed at 4°Cfor 75 days by apposi-tion against tritium-sensitive film (Hyperfilm-
3H, AmershamInternational, U.K.), and thefilms were developedand quan-tified in a blind manner using a computerized image analysissystem (Microm, Barcelona, Spain). Optical densities wereconverted into amount of radioligand bound by comparisonwith the optical densities of the standards ([3H] Microscales,Amersham International).
Data analysisAnalyses of saturation andcompetition isotherms to deter-
mine the equilibrium dissociation constants (K0, K,) and
maximal number of sites (B~,ax)and the fitting of data to theappropriate binding models were performed by computer-assisted nonlinear regression using the EBDA-LIGAND pro-grams (Munson andRodbard, 1980; McPherson, 1985). Theselection between binding models was made statistically bythe extra sum of squares principle (F test), as outlined byMunson andRodbard (1980). The more complex model wasaccepted if the p value resulting from the F test was <0.05.
The competition curves by the a2-adrenoceptor antagonistARC239 against the antagonist [
3H]RX821002 were alwaysanalyzedsimultaneously with the saturation curves for [3H] -
RX821002 binding performed in the same subjects to esti-mate individual K
0, K,, and B~,axparameters more exactly.Furthermore and for each subject, the saturation experimentswith the agonists [
3Hjclonidine and [3H]UK14304werealso coanalyzed with the saturation experiments with theantagonist [3H]RX821002 and with the competition experi-ments by the agonist (— ) -adrenaline (Sastre and GarcIa-Sevilla, 1994b). The two-site binding model was chosen bythe LIGAND program (F test) as the most appropriate todescribe theexperimental data (for further details, see Sastreand Garcia-Sevilla, l994b).
Results are expressed as means ±SEM. Two-way ANOVAand Student’s t test were used for the statistical evaluations.The level of significance was chosen at p = 0.05.
Materials[3H]RX82 1002 (2-methoxyidazoxan) (53—60 Ci/mmol)
was purchased from Amersham International and stored at4°C. [3H]Clonidine (66—68 Ci/mmol) and [3H]UK14304(5-bromo-N-(4,5-[3H] dihydro- 1H-imidazol-2-yl)-6-quin-oxalinamine } (60—88 Ci/mmol) were obtained from DuPont/New England Nuclear (Germany) and stored at—30°C.Other drugs (and their sources) included the follow-ing: ARC239 {2-[2-[4-(o-methoxyphenyl)piperazin- 1-yllethyl] -4,4- dimethyl - 1,3 - (2H, 4H) - isoquinolindioneHC1} (Thomae GmbH, Germany), (—)-adrenaline bitar-trate (Sigma Chemical Co., St. Louis, MO, U.S.A.). Allother reagents were purchased from either Sigma or Merck(Germany).
RESULTS
a2-Adrenoceptor subtypes in brain membranes of
control subjects and suicide victimsThe specific binding of the antagonist [
3H] -
RX821002 to human brain membranes from the pre-frontal cortex and caudate resulted in saturable pro-cesses of high affinity (K
0 values in the nanomolarrange) that met the pharmacological criteria for the
J. Neurochem., vol. 70, No. 3, 1998

a2-ADRENOCEPTORS IN DEPRESSION 1117
TABLE 2. Parameters of [3H]RX821002 specific binding and affinity of ARC239 to a
2-adrenoceptors in brainof control subjects and suicide victims with major depression or other diagnoses
membranes
Frontal cortex Caudate head
Bmax Bmaxi(fmol/mg (fmol/mg
K0 (nM) K, (nM) of protein) K0 (nM) K11 (nM) K12 (nM) of protein)
Bmax2
(fmol/mgof protein)
Controls (n = 10)Suicide victims with
depression (n = 10)Suicide victims with
other diagnoses(n = 10)
0.69 ±0.24 699 ±96 107 ±5 0.45 ±0.13 2.7 ±0.9 942 ±266 3 ±1
0.78 ±0.23 904 ±119 85 ±11 0.28 ±0.07 2.2 ±0.9 1,750 ±470 2 ±1
0.78 ±0.01 854 ±116 119 ±11 0.23 ±0.08 2.1 ±0.7 775 ±772 4 ± 1
18 ±3
20 ±2
29 ±5
Neural membranes were incubated for 30 mm at 25°Cwith [3H]RX821002(0.03—8 nM) for saturation studies. Simultaneous competition
studies were performed with [3H]RX821002(1 nM) in the absence or presence of ARC239 (10_12_ l0—~M; 19 concentrations). Assay bufferincluded 0.3 pM 5-HT to prevent binding of [3H]RX821002to nonadrenoceptor binding sites. K
0 values are the equilibrium dissociationconstants of [
3H]RX821002binding. K~,K~,and K
12 are the inhibition constants of ARC239 competing for [3H]RX821002binding sites. B~,
Bmaxi, and Bm
2 are the maximal number of [3HIRX821002binding sites. The sites numbered I and 2 correspond to the a2c- and £52A-
adrenoceptor subtypes, respectively. For each subject and each brain region, the saturation curves and the competition curves by ARC239were simultaneously fitted using the EBDA-LIGAND programs to a one-site or two-site binding model. The more appropriate model waschosen by an F test using the extra sum of squares principle. In the frontal cortex, a one-site model (a
2A-adrenoceptor) gave a statisticallysignificant better fit than a two-site model. In contrast, in caudate nucleus, a two-site binding model (a2A- and a2c-adrenoceptors) improvedthe goodness of fit. Values are the means ±SEM of n experiments performed in different subjects.
identification of a2-adrenoceptors (Grijalba et al.,1996).
In the frontal cortex, the coanalysis of the saturationexperiments with [
3HI RX821002 and the competitionexperiments with ARC239/[3H]RX821002 resulted inmonophasic curves with K, values in the high nanomo-lar range, suggesting the presence of a single subtypepopulation of a
2A-adrenoceptors (Table 2). No sig-nificant differences in K, values for ARC239 were ob-served between the control and suicide groups (Table2). The KD values for {
3H]RX821002 were similar indepressed suicides, suicides with other diagnoses, andcontrols (Table 2). The density (Bmax values) of thesea2A-adrenoceptor antagonist binding sites in the frontal
cortex did not differ significantly between the threegroups (Table 2).
In the caudate, the coanalysis of similar saturationand competition experiments unraveled the presenceof two binding sites (Table 2). The minority site(Bmaxi), which represented 4—27% of the specificbinding, was recognized with high affinity byARC239 (K, values in the low nanomolar range). Incontrast, the predominant site (Bmax2) displayed lowaffinity for ARC239 (K1 values in the high nanomolarrange) (Table 2). Thus, in the head of caudate, amixed population of a2c- and a2A-adrenoceptors wasfound. The proportions of both subtypes were similarin the control group (a2A, 14%; a2c, 86%) and thetwo suicide groups (a2A, 9—12%; a2c, 88—91%)(Table 2). Binding affinities (K0 values) for ~
3H]-RX821002 labeling a
2A/C-adrenoceptors were simi-lar in controls, depressed suicides, and suicides withother diagnoses (Table 2). As the antagonist [
3H]-RX82 1002 does not discriminate between a
2-adreno-ceptor subtypes (see Sastre and GarcIa-Sevilla,
1994a; Tunstall et al., 1996), differences of the bind-ing affinity between both a2-adrenoceptor subtypescould not be discriminated (Table 2).
a2A-Adrenoceptors in brain sections of controlsubjects and suicide victims
In the frontal cortex, incubation of tissue sectionswith ARC239 (20 nM) selectively masked the pop-ulation of a2c-adrenoceptors in layer III and the5-HT-sensitive component identified by [
3H] -
RX821002 (Grijalba eta!., 1996). This masking pro-cedure allowed the selective evaluation of a
2A-adre-noceptors with the antagonist [
3H]RX821002 (Fig.1). A two-way ANOVA revealed no significant dif-ferences in the density of a
2A-adrenoceptors betweencontrols and suicides along the different layers ana-lyzed in the frontal cortex (Table 3 and Fig. 2).Similar analyses revealed significant differences be-tween cortical layers (p < 0.01) without any signifi-cant influence of the interaction of diagnoses andanatomical location.
In hippocampal sections, ARC239 (20 nM) was alsoable to block selectively the population of cr~~-adreno-ceptors and the 5-HT-sensitive component of [
3H1-RX821002 binding (data not shown; see Grijalba et
a!., 1996). Under these experimental conditions, theautoradiographic density of a
2A-adrenoceptors in thehippocampus of suicides was not different from thatin controls (Table 3 and Fig. 2). A two-way ANOVAof [
3H]RX821002 binding values in the dentate gymsrevealed significant differences attributable to anatomi-cal layers (p < 0.01) without significant differencesattributable to the diagnoses or to the interaction be-tween diagnoses and anatomical location. The stratumlacunosum moleculare of the CA 1 (cornu Ammonis)
J. Neurochem., vol. 70, No. 3, 1998
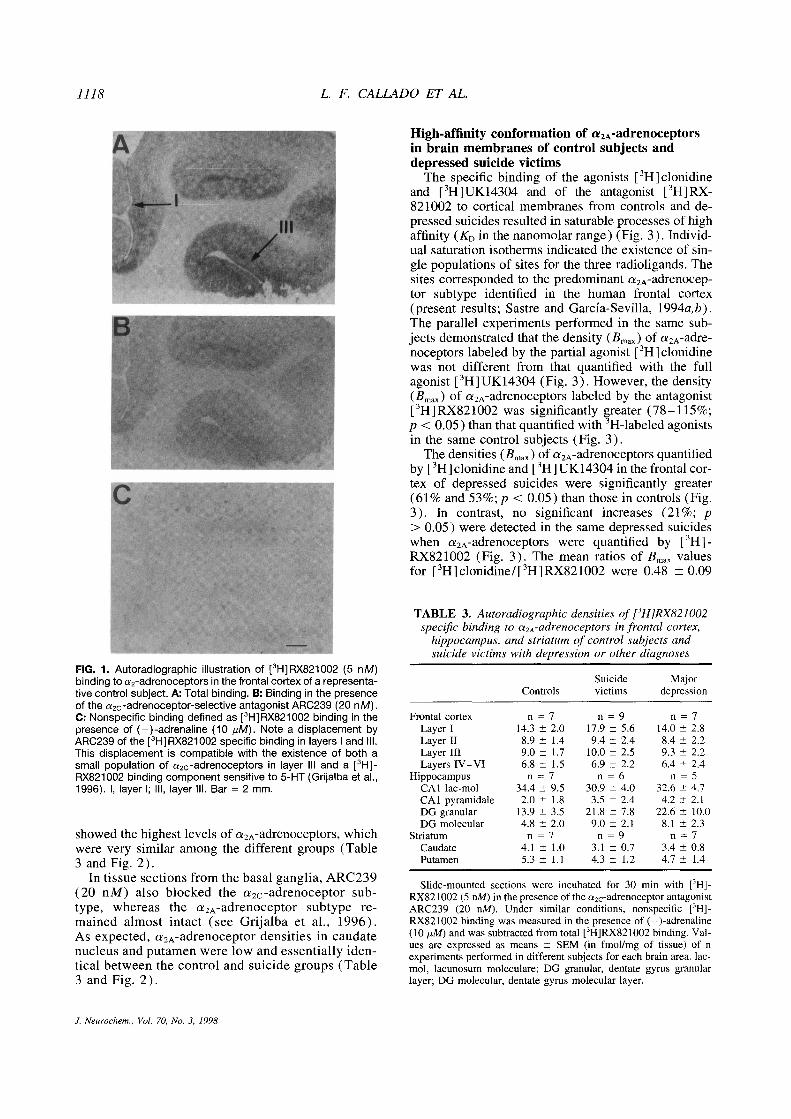
1118 L. F. CALLADO ET AL.
High-affinity conformation of E2A-adrenoceptorsin brain membranes of control subjects anddepressed suicide victims
The specific binding of the agonists [3Hjc!onidine
and [3HjUK14304 and of the antagonist [3H]RX-82 1002 to cortical membranes from controls and de-pressed suicides resulted in saturable processes of highaffinity (KD in the nanomolarrange) (Fig. 3). Individ-ual saturation isotherms indicated the existence of sin-gle populations of sites for the three radioligands. Thesites corresponded to the predominant a
2A-adrenocep-tor subtype identified in the human frontal cortex(present results; Sastre and Garcia-Sevilla, 1994a,b).The parallel experiments performed in the same sub-jects demonstrated that the density (Bmax) of a2A-adre-noceptors labeled by the partial agonist [
3H]clonidinewas not different from that quantified with the fullagonist [3H]UK14304 (Fig. 3). However, the density(Bmax) of a
2A-adrenoceptors labeled by the antagonist[3H]RX821002 was significantly greater (78—115%;
p < 0.05) than that quantified with 3H-labeled agonistsin the same control subjects (Fig. 3).
The densities (Bmax) of a2A-adrenoceptors quantified
by3H I clonidine and [3HI UK14304 in the frontalcor-
tex of depressed suicides were significantly greater(61% and 53%; p < 0.05) than those in controls (Fig.3). In contrast, no significant increases (21%; p> 0.05) were detected in the same depressed suicideswhen a
2A-adrenoceptors were quantified by [3HI -
RX821002 (Fig. 3). The mean ratios of Bmax valuesfor [3HIclonidine/~3H]RX821002 were 0.48 ±0.09
FIG. 1. Autoradiographic illustration of [3H]RX821002(5 nM)binding to a
2-adrenoceptors in the frontal cortex of a representa-tive control subject. A: Total binding. B: Binding in the presenceof the a2c-adrenoceptor-selective antagonist ARC239 (20 nM).C: Nonspecific binding defined as [
3H]RX821002binding in thepresence of (—)-adrenaline (10 ~sM).Note a displacement byARC239 of the [3H]RX821002specific binding in layers I and Ill.This displacement is compatible with the existence of both asmall population of a
2c-adrenoceptors in layer Ill and a [3Hj-
RX821 002 binding component sensitive to 5-HT (Grijalba et al.,1996). I, layer I; Ill, layer Ill. Bar = 2 mm.
showed the highest levels of a2A-adrenoceptors, which
were very similar among the different groups (Table3 and Fig. 2).
In tissue sections from the basal ganglia, ARC239(20 nM) also blocked the a2~-adrenoceptorsub-type, whereas the a2A-adrenoceptor subtype re-mained almost intact (see Grijalba et al., 1996).As expected, a2A-adrenoceptor densities in caudatenucleus and putamen were low and essentially iden-tical between the control and suicide groups (Table3 and Fig. 2).
TABLE 3. Autoradiographic densities of [3H]RX821002
specific binding to a2A-adrenoceptors in frontal cortex,
hippocampus, and striatum of control subjects andsuicide victims with depression or other diagnoses
Suicide MajorControls victims depression
Frontal cortex n = 7 n = 9 n = 7Layer I 14.3 ±2.0 17.9 ±5.6 14.0 ±2.8Layer II 8.9 ±1.4 9.4 ±2.4 8.4 ±2.2Layer III 9.0 ±1.7 10.0 ±2.5 9.3 ±2.2Layers IV—VI 6.8 ±1.5 6.9 ±2.2 6.4 ±2.4
Hippocampus n = 7 n = 6 n = 5
CAl lac-mol 34.4 ±9.5 30.9 ±4.0 32.6 ±4.7CAl pyramidale 2.0 ±1.8 3.5 ±2.4 4.2 ±2.1DG granular 13.9 ±3.5 21.8 ±7.8 22.6 ±10.0DG molecular 4.8 ±2.0 9.0 ±2.1 8.1 ±2.3
Striatum n = 7 n = 9 n = 7Caudate 4.1 ±1.0 3.1 ±0.7 3.4 ±0.8Putamen 5.3 ±1.1 4.3 ±1.2 4.7 ±1.4
Slide-mounted sections were incubated for 30 mm with [3HJ-
RX821002 (5 nM) in the presence of thea2c-adrenoceptor antagonist
ARC239 (20 nM). Under similar conditions, nonspecific [3HI-
RX821002 binding was measured in the presence of (—)-adrenaline(10 jsM) and was subtracted from total [3H]RX821002binding. Val-ues are expressed as means ±SEM (in fmol/mg of tissue) of nexperiments performed in different subjects for each brain area. lac-mol, lacunosum moleculare; DG granular, dentate gyms granularlayer; DG molecular, dentate gyms molecular layer.
J. Neurochem., v
0l. 70, No. 3, 1998

a2-ADRENOCEPTORS IN DEPRESSION 1119
FIG. 2. Autoradiographic illustrations ofthe density ofcs2A-adrenoceptors in the frontal cortex (A and B), me-dial hippocampus (C and D), and ventral striatum (Eand F) in control subjects (A, C, and E) and suicidevictims with major depression (B, D, and F). Brain sec-tions were incubated with [
3H]RX821002(5 nM) in thepresence ofARC239 (20 n M) to mask aac -adrenocep-tors and 5-HT-sensitive receptors. Note a similarauto-radiographical density in depressed suicides and con-trols along the different structures, Ill, cortical layer Ill;Im, hippocampal lacunosum moleculare; p, hippocam-pal pyramidale; DG, hippocampal dentate gyrus; C,caudate; P, putamen. Bar = 2 mm.
in controls and 0.60 ±0.06 in suicides (25% greater).The corresponding mean ratios of Bmax values for [3HI -
UK14304/[3HIRX821002 were 0.57 ± 0.07 in con-trols and 0.69 ± 0.09 in suicides (21% greater). Theaffinities (K
0 values) of the3H-labeled agonists and the
3H-labeled antagonist for the receptors did not differsignificantly between groups (Fig. 3).
In frontal cortex of controls and depressed suicides,
the competition curves of [3H]RX821002 (1 nM)binding by (— )-adrenaline were biphasic, which indi-cated the existence of two conformational states forthe a
2A-adrenoceptor (Table 4). In the presence ofguanine nucleotides, (— )-adrenaline displayed mono-phasic competition curves of low affinity against [
3H11-RX821002 in frontal cortex (data not shown; see Sas-tre and GarcIa-Sevilla, 1994b). The simultaneousanal-
.1. Neurochem., vol. 70, No. 3, 1998
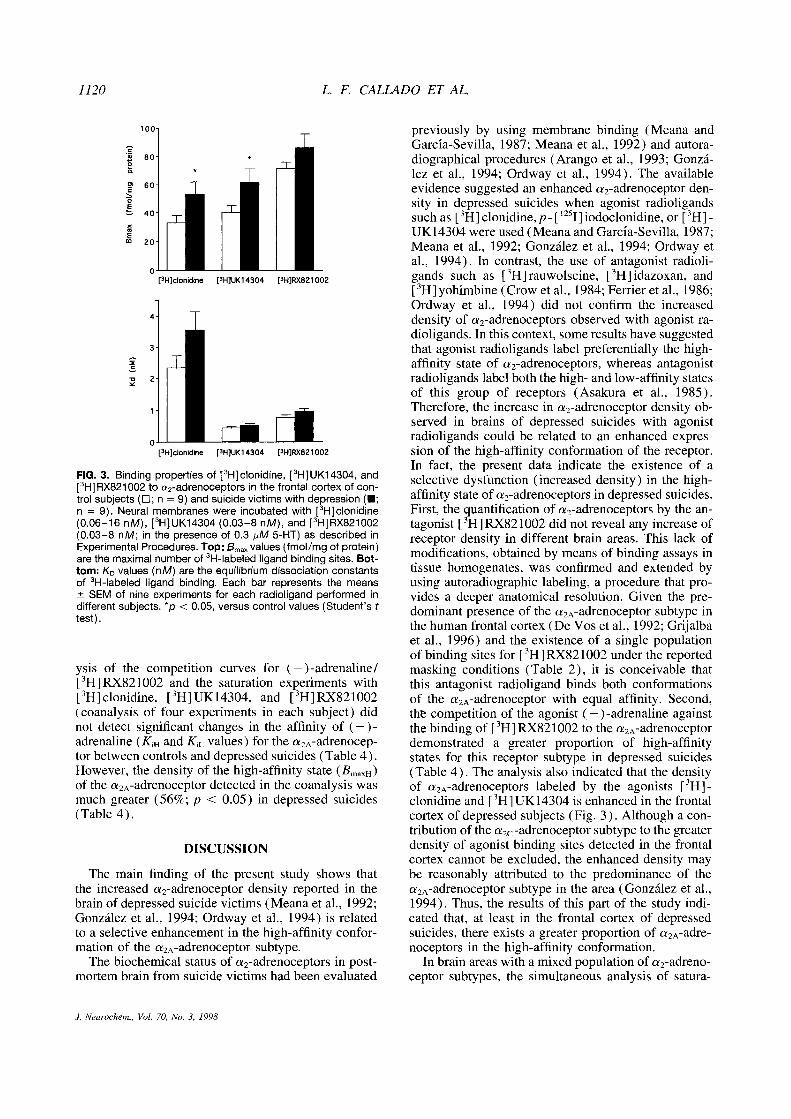
1120 L F. CALLADO ET AL
FIG. 3. Binding properties of [3H]clonidirie,[3H]UK14304, and[3H1RX821002 to a
2-adrenoceptors in the frontal cortex of con-trol subjects (~n = 9) and suicide victims with depression (U;n = 9). Neural membranes were incubated with [
3Hjclonidine(0.06—16 nM), [3H1UK14304(0.03—8 nM), and [3H1RX821002(0.03—8 nM; in the presence of 0.3 p~M5-HT) as described inExperimental Procedures. Top: B~values (fmol/mg of protein)are the maximal number of 3H-labeled ligand binding sites. Bot-tom: K
0 values (nM) are the equilibrium dissociation constantsof
3H-labeled ligand binding. Each bar represents the means± SEM of nine experiments for each radioligand performed indifferent subjects. *p < 0.05, versus control values (Student’stest).
ysis of the competition curves for (—)-adrenaline/[3HI RX821002 and the saturation experiments with[3HI clonidine, 3H I UK14304, and t~3H] RX821002(coanalysis of four experiments in each subject) didnot detect significant changes in the affinity of (— ) -
adrenaline (KIH and K11 values) for the a2A-adrenocep-
tor between controls and depressed suicides (Table 4).However, the density of the high-affinity state (Bm~H)of the a2A-adrenoceptor detected in the coanalysis wasmuch greater (56%; p < 0.05) in depressed suicides(Table 4).
DISCUSSION
The main finding of the present study shows thatthe increased a2-adrenoceptor density reported in thebrain of depressed suicide victims (Meana et al., 1992;Gonzalez et a!., 1994; Ordway et al., 1994) is relatedto a selective enhancement in the high-affinity confor-mation of the a2A-adrenoceptor subtype.
The biochemical status of a2-adrenoceptors in post-mortem brain from suicide victims had been evaluated
previously by using membrane binding (Meana andGarcia-Sevilla, 1987; Meana et a!., 1992) and autora-diographical procedures (Arango et a!., 1993; Gonzá-lez et al., 1994; Ordway et a!., 1994). The availableevidence suggested an enhanced a2-adrenoceptor den-sity in depressed suicides when agonist radioligandssuch as [
3H]clonidine, p- [12511iodoclonidine, or [3HI -
UK14304 were used (Meana and GarcIa-Sevilla, 1987;Meana et a!., 1992; Gonzalez et a!., 1994; Ordway eta!., 1994). In contrast, the use of antagonist radioli-gands such as [3HI rauwolscine, [3Hj idazoxan, and[3Hjyohimbine (Crow eta!., 1984; Ferrier eta!., 1986;Ordway et a!., 1994) did not confirm the increaseddensity of a
2-adrenoceptors observed with agonist ra-dio!igands. In this context, some results have suggestedthat agonist radioligands label preferentially the high-affinity state of a2-adrenoceptors, whereas antagonistradioligands label both the high- and low-affinity statesof this group of receptors (Asakura et al., 1985).Therefore, the increase in a2-adrenoceptor density ob-served in brains of depressed suicides with agonistradioligands could be related to an enhanced expres-sion of the high-affinity conformation of the receptor.In fact, the present data indicate the existence of aselective dysfunction (increased density) in the high-affinity state of a2-adrenoceptors in depressed suicides.First, the quantification of cr2-adrenoceptors by the an-tagonist II
3HI RX821002 did not reveal any increase ofreceptor density in different brain areas. This lack ofmodifications, obtained by means of binding assays intissue homogenates, was confirmed and extended byusing autoradiographic labeling, a procedure that pro-vides a deeper anatomical resolution. Given the pre-dominant presence of the a
2A-adrenoceptor subtype inthe human frontal cortex (De Vos et a!., 1992; Grijalbaet al., 1996) and the existence of a single populationof binding sites for [
3H]RX821002 under the reportedmasking conditions (Table 2), it is conceivable thatthis antagonist radioligand binds both conformationsof the a
2A-adrenoceptor with equal affinity. Second,th~competition of the agonist (— ) -adrenaline againstthe binding of [
3H]RX821002 to the a2A-adrenoceptor
demonstrated a greater proportion of high-affinitystates for this receptor subtype in depressed suicides(Table 4). The analysis also indicated that the densityof a2A-adrenoceptors labeled by the agonists [
3H]-
clonidine and [3H]UK 14304 is enhanced in the frontalcortex of depressed subjects (Fig. 3). Although a con-tribution of the a2c -adrenoceptor subtype to the greaterdensity of agonist binding sites detected in the frontalcortex cannot be excluded, the enhanced density maybe reasonably attributed to the predominance of thea2A-adrenoceptor subtype in the area (Gonzalez et a!.,
1994). Thus, the results of this part of the study indi-cated that, at least in the frontal cortex of depressedsuicides, there exists a greater proportion of a2A-adre-noceptors in the high-affinity conformation.
In brain areas with a mixed population of a2-adreno-ceptor subtypes, the simultaneous analysis of satura-
J. Neurochem., vol. 70, No. 3, 1998

a2-ADRENOCEPTORS IN DEPRESSION 1121
TABLE 4. Parameters of the high- and low-affinity states of a2A-adrenoceptors in corticalmembranes from control subjects and suicide victims with major depression
Slope(n0)
KIH
(nM)KIL
(nM)Bm,,xH
(fmol/mg of protein)
Controls (n = 9) 0.51 ±0.03” 4.8 ±1.6 1,921 ±998 39 ±5Suicide victims with major
depression (n = 9) 0.49 ±0.03” 5.5 ±1.7 4,238 ±3,607 61 ±9l~
Neural membranes were incubated with [3H]clonidine (0.06—16 nM), [3H]UK14304 (0.03—8 nM), or
[3H]RX821002 (0.03—8 nM; in the presence of 0.3 pM-5-HT) for saturation experiments. Competitionexperiments of [3H]RX821002 binding (1 nM; in the presence of 0.3
1iM 5-HT) by the agonist (H-adrenaline (10_12_ l0~M; 19 concentrations) were also performed. Saturation and competition experimentswere coanalyzed (four experiments in each subject) using the EBDA-LIGAND programs on the assumptionthat the a2A-adrenoceptor expresses two conformational states [high- (H) and low- (L) affinity states foragonists] that are recognized with similar and nondiscriminating affinity by antagonists. Slope values (flH)
for the competition curves of the agonist (H-adrenaline against the antagonist [3H]RX821002 were different
from unity, suggesting the existence of a two-state binding model with high- and low-affinity inhibitionconstants (KH, K,~)for the agonist (F test). Bm=H values are the maximal number of high-affinity bindingsites obtained from the coanalysis of the four experiments. Values are the means ±SEM of the experimentsperformed in n different subjects.
flf
4 values <1 (p < 0.001).1’p < 0.05, versus control value (Student’s t test).
tion curves for [3HI RX821002 together with competi-tion curves of II3H1RX821002 binding by the selectivea213 IC-adrenoceptor antagonist ARC239 provided a
methodological approach for the delineation of a2A-
and a2~-adrenoceptorsubtypes in the same subject(Sastre and GarcIa-Sevilla, 1994a; Grijalba et a!.,1996). By using this approach, the proportions of braina2A- and a2c -adrenoceptor subtypes evaluated by an-tagonist drugs (high- and low-affinity conformationstogether) were very similar for depressed suicides, sui-cides with other diagnoses, and control subjects (Table2). In agreement with previous studies, the a2c -adre-noceptor subtype is expressed mainly in the humanstriaturn (Petrash and Bylund, 1986; De Vos et al.,1992; Perhlh et al., 1992; Ordway et al., 1993; Sastreand GarcIa-Sevilla, 1994a; Grijalba et a!., 1996).However, the density of a2c-adrenoceptors in the pres-ent study was lower than that reported in previousstudies with the same radioligand (De Vos et al., 1992;Sastre and GarcIa-Sevi!!a, 1994a; Grijalba et a!.,1996). This finding is probably due to the absence ofguanine nucleotides in the medium (Sastre and Garcia-Sevilla, 1994a; Callado and Meana, 1997). This verylow density of a2c-adrenoceptors in the human stria-turn did not allow us to evaluate the affinity conforma-tions of this adrenoceptor subtype after selective mask-ing of the a2A-adrenoceptor subtype. Thus, the exis-tence of hypothetical dysfunctions in the high-affinitystate of thea2c -adrenoceptor subtype related to depres-sion could not be ruled out.
The present findings are consistent with those fromother studies in human platelets that contain exclu-sively the a2A-adrenoceptor subtype (By!und, 1985;Bylund et a!., 1988). In platelets from depressed pa-tients, a higher a2-adrenoceptor density has been re-ported with the use of agonist radioligands (Garcia-Sevi!!a et al., 1981, 1986, 1987; Doyle et al., 1985;
Piletz and Halaris, 1988; Piletz et a!., 1990, 1991).However, the attempts to confirm these findings withthe use of antagonist radioligands have been unsuc-cessful, leading to the hypothesis that only the high-affinity conformation of the platelet a2-adrenoceptoris elevated in depression (GarcIa-Sevil!a et al., 1986;Piletz et a!., 1986).
It is of note that most of the suicides in the presentstudy were receiving antidepressant or other psy-chotropic drugs immediately prior to death. In this con-text, it is well accepted that antidepressant treatmentsinduce down-regulation and desensitization of a2-adre-noceptors in the platelets of depressed patients (a2A
subtype) (Garcia-Sevi!la et a!., 1981, 1986, 1987,1990; Piletz and Halaris, 1988; Pandey et a!., 1989;Karege et al., 1992; Piletz et al., 1996) and in the ratCNS (Smith et al., 1981; Nomura et al., 1987; Giraltand GarcIa-Sevilla, 1989; Menargues et a!., 1990; Heal~t a!., 1991; Mongeau et al., 1994). Therefore, apossi-ble down-regulation of brain a2-adrenoceptors bychronic antidepressant treatment could be expected inthe suicide group (De Paermentier et a!., 1997). How-ever, it should be pointed out that in the present studythe density of a2A-adrenoceptors in depressed suicideswas enhanced in spite of the reported negative effectof antidepressant drug treatment. The control groupsincluded herein showed various causes of death, andapparently the subjects lacked a previous diagnosis ofneuropsychiatric disorder. However, the presence ofundiagnosed subjects with depression or another disor-der cannot be completely excluded.
The current findings further support the hypothesisthat an enhanced a2A-adrenoceptor autoreceptor activ-ity could be involved in the etiology of depression. Inthe therapeutic context of this hypothesis, the develop-ment of non — subtype-selective a2-adrenoceptor antag-onists, such as mirtazapine, appears to be a new and
J. Neurochem., vol. 70, No. 3, 1998

1122 L F. CALLADO ET AL
effective pharmacological approach to the treatment ofdepressive disorders (Bremner, 1995; Claghorn andLesen, 1995).
Acknowledgment: This study was supported by grantsfrom the FIS (95/1731), Basque Government (P1 93/75and Department of Health) (J.J.M.), Salud 2000 Foundation,CICYT (SAF 95/0362) (A.P.), and DGICYT (PB 94/0002-Mod C) (J.A.G.-S.). L.F.C., B.G., and MS. were supportedby fellowships from the Basque Government, the Universityof Cantabria, and MEC, respectively. The authors wish tothank the staffmembers of the Instituto Anatómico Forense,Bilbao and Palma de Mallorca, for their cooperation. Thetechnical assistance of Ms. Lourdes Lanza andMs. M. JosefaCastillo is kindly acknowledged.
REFERENCES
American Psychiatric Association (1987) Diagnostic and StatisticalManual of Mental Disorders, 3rd edit., revised. American Psy-chiatric Association, Washington, D.C.
Arango V., Ernsberger P., Sved A. F., and Mann J. J. (1993) Quanti-tative autoradiography of a
1- and a2-adrenergic receptors in thecerebral cortex of controls and suicide victims. Brain Res. 630,271—282.
Asakura M., Tsukamoto T., Imafuku J., Matsui H., Ino M., andHasegawa K. (1985) Quantitative analysis of rat brain a2-recep-tors discriminated by [
3H]clonidine and [3H]rauwolscine. Eur.J. Pharmacol. 106, 141—147.
BremnerJ. D. (1995) A double-blind comparison of Org 3770, ami-triptyline, and placebo in major depression. J. Clin. Psychiatry56, 5 19—525.
Bylund D. B. (1985) Heterogeneity of alpha-2 adrenergic receptors.Pharmacol. Biochem. Behav. 22, 835—843.
Bylund D. B. (1992) Subtypes of a1- and a2-adrenergic receptors.
FASEB .1. 6, 832—839.Bylund D. B., Ray-Prenger C., and Murphy T. J. (1988) Alpha-2A
and alpha-2B adrenergic receptor subtypes: antagonist bindingin tissues and cell lines containing only one subtype. J. Pharma-cot. Exp. Ther. 245, 600—607.
Caldecott-Hazard S., Morgan D. G., DeLeon-Jones F., OverstreetD. H., and Janowsky D. (1991) Clinical and biochemical as-pects of depressive disorders. II. Transmitter/receptor theories.Synapse 9, 251—301.
Callado L. F. and Meana J. J. (1997) Low-affinity conditions foragonists increase the binding of the antagonist [
3H]RX82 1002to the a
2B,C-adrenoceptor subtypes in human brain and rat kid-ney. Eur. J. Pharmacol. 332, 109—112.
Cheetham S. C., Katona C. L. E., and Horton R. W. (1991) Postmor-tem studies of neurotransmitter biochemistry in depression andsuicide, in Biological Aspects of Affective Disorders (HortonR. and Katona C., eds), pp. 191 —221. Academic Press, London.
Claghorn J. L. and Lesen M. D. (1995) A double-blind placebo-controlled study of Org 3770 in depressed outpatients. J. Affect.Disord. 34, 165—171.
Crow T. J., Cross A. J., Cooper S. J., Deakin J. F. W., Ferrier I. N.,Johnson J. A., Joseph M. H., Owen F., Poulter M., LofthouseR., Corsellis J. A. N., Chambers D. R., Blessed G., Perry E. K.,Perry R. H., and Tomlinson B. E. (1984) Neurotransmitter re-ceptors and monoamine metabolites in the brain of patientswith Alzheimer-type dementia and depression, and suicides.Neuropharmacology 23, 1561—1569.
De Paermentier F., Mauger J. M., Lowther S., Crompton M. R.,Katona C. L. E., and Horton R. W. (1997) Brain a-adrenocep-tors in depressed suicides. Brain Res. 757, 60—68.
De Vos H., Vauquelin G., De Keyser J., De Backer J.-P., and VanLiefde I. (1992) Regional distribution of a2A- and a25-adreno-ceptor subtypes in postmortem human brain. J. Neurochem. 58,1555— 1560.
Doyle M. C., George A. J., Ravindran A. V., and Philpott R. (1985)Platelet a2-adrenoceptor binding in elderly depressed patients.Am. J. Psychiatry 142, 1489—1490.
Esteban S., Lladó J., and GarcIa-Sevilla J. A. (1996) a2-Autorecep-tors and a2-heteroreceptors modulating tyrosine and tryptophanhydroxylase activity in the rat brain in vivo: an investigationinto the a2-adrenoceptor subtypes. Naunyn SchmiedebergsArch. Pharmacol. 353, 391—399.
Ferrier I. N., McKeith I. G., Cross A. J., Perry E. K., Candy J. M.,and Perry R. H. (1986) Postmortem neurochemical studies indepression. Ann. NYAcad. Sd. 487, 128—142.
GarcIa-Sevilla J. A., Zis A. P., Hollingsworth P. J., Greden J. F., andSmith C. B. (1981) Platelet a2-adrenergic receptors in majordepressive disorder. Binding of tritiated clonidine before andafter tricyclic antidepressant drug treatment. Arch. Gen. Psychi-atry 38, 1327—1333.
GarcIa-Sevilla J. A., Guimón J., Garcia-Vallejo P., and Fuster M. J.(1986) Biochemical and functional evidence of supersensitiveplatelet a2-adrenoceptors in major affectivedisorder. Arch. Gen.Psychiatry 43, 5 1—57.
GarcIa-Sevilla J. A., Udina C., Fuster M. J., Alvarez E., and CasasM. (1987) Enhanced binding of [
3H] (— )adrenaline to plateletsof depressed patients with melancholia: effect of long-termclomipramine treatment. Acta Psychiatr. Scand. 75, 150— 157.
GarcIa-Sevilla J. A., Padró D., Giralt M. T., Guimón J., and AresoP. (1990) a
2-Adrenoceptor-mediated inhibition of platelet ade-nylate cyclase and induction of aggregation in major depression.Effect of long-term cyclic antidepressant drug treatment. Arch.Gen. Psychiatry 47, 125—132.
Giralt M. T. and Garcia-Sevilla J. A. (1989) Acute and long-termregulation of brain a2-adrenoceptors after manipulation of nor-adrenergic transmission in the rat. Eur. J. Pharmacol. 164,455—466.
Gonzalez A. M., Pascual J., Meana J. J., Barturen F., del Arco C.,Pazos A., and GarcIa-Sevilla J. A. (1994) Autoradiographicdemonstration of increased a2-adrenoceptor agonist bindingsites in the hippocampus and frontal cortexof depressed suicidevictims. J. Neurochem. 63, 256—265.
Gothert M. and Huth H. (1980) Alpha-adrenoceptor-mediated mod-ulation of 5-hydroxytryptamine release from rat brain cortexslices. Naunyn Schmiedebergs Arch. Pharmacol. 313, 21—26.
Grijalba B., Callado L. F., Meana J. J., GarcIa-Sevi!la J. A., andPazos A. (1996) a2-Adrenoceptor subtypes in the human brain:a pharmacological delineation of [
3H]RX82 1002 binding tomembranes and tissue sections. Eur. .J. Pharmacol. 310, 83—93.
Heal D. J., Prow M. R., and Buckett W. R. (1991) Determinationof the role of noradrenergic and 5-hydroxytryptaminergic neu-rones in postsynaptic a
2-adrenoceptor desensitization by desi-pramine and ECS. Br. J. Pharmacol. 103, 1865—1870.
KaregeF.,BovierP.,WidmerJ., GaillardJ. M., and Tissot R.(l992)Platelet membrane alpha2-adrenergic receptors in depression.Psychiatry Res. 43, 243—252.
Karege F., Bovier P., Rudolph W., and Gaillard J-M. (1996) Plateletphosphoinositide signaling system: an overstimulated pathwayin depression. Biol. Psychiatry 39, 697—702.
Lowry0. H., Rosebrough N. J., Fan A. L., and Randall R. J. (1951)Protein measurement with the Folin phenol reagent. J. Biol.Chem. 193, 265—275.
McPherson G. A. (1985) Analysis of radioligand binding experi-ments: a collection of computer programs for IBM PC. J. Phar-macol. Methods 14, 213—228.
Meana J. J. and GarcIa-Sevilla J. A. (1987) Increased a2-adrenocep-tor density in the frontal cortex of depressed suicide victims.J. Neural Transm. 70, 377—381.
Meana J. J., Barturen F., and Garcia-Sevilla J. A. (1989) Character-ization and regional distribution of a2-adrenoceptors in post-mortem human brain using the full agonist [
3H]UK 14304. J.Neurochem. 52, 1210—1217.
Meana J. J., Barturen F., and GarcIa-Sevilla J. A. (1992) a2-Adreno-
ceptors in the brain of suicide victims: increased receptor den-
.1. Neurochem., vol. 70, No. 3, 1998

a2-ADRENOCEPTORS IN DEPRESSION 1123
sity associated with major depression. Biol. Psychiatry 31,471—490.
Meana J. J., Callado L. F., Pazos A., Grijalba B., and GarcIa-SevillaJ. A. (1996) The subtype-selective a2-adrenoceptor antagonistsBRL 44408 and ARC 239 also recognize 5-HTIA receptors inthe rat brain. Eur. J. Pharmacol. 312, 385—388.
Menargues A., Obach R., and GarcIa-Sevilla J. A. (1990) Modula-tion by antidepressant drugs of CNS postsynaptic a2-adrenocep-tors mediating mydriasis in the rat. Naunyn SchmiedebergsArch. Pharmacol. 341, 101—107.
Mongeau R., De Montigny C., and Blier P. (1994) Electrophysio-logic evidence for desensitization of a2-adrenoceptors on seroto-nm terminals following long-term treatment with drugs increas-ing norepinephrine synaptic concentration. Neuropsychophar-macology 10, 41—51.
Munson P. J. and Rodbard D. (1980) LIGAND: a versatile compu-terized approach for characterization of ligand binding system.Anal. Biochem. 107, 220—239.
Nomura S., Duman R. S., and Enna S. J. (1987) In vivo or in vitroexposure to imipramine reduces a2-adrenoceptor-mediated inhi-bition of cyclic AMP production in rat brain cerebral corticalslices. Brain Res. 410, 195—198.
Ordway G. A., Jaconetta S. M., and Halaris A. (1993) Characteriza-tion of subtypes of alpha-2 adrenoceptors in the human brain.J. Pharmacol. Exp. Ther. 264, 967—976.
Ordway G. A., Widdowson P. S., Smith K. S., and Halaris A. (1994)Agonist binding to a2-adrenoceptors is elevated in the locuscoeruleus from victims of suicide. J. Neurochem. 63, 617—624.
Pandey G. N., Janicak P. G., Javaid J. I., and Davis J. M. (1989)Increased
3H-clonidine binding in the platelets of patients withdepressive and schizophrenic disorders. Psychiatry Res. 28, 73—88.
PerSlä M., Hirvonen H., Kalimo H., Ala-Uotila S., Regan J. W.,Akerman K. E. 0., and Scheinin M. (1992) Differential expres-sion of two a
2-adrenergic receptor subtype mRNAs in humantissues. Mol. Brain Res. 16, 57—63.
Petrash A. C. and Bylund D. B. (1986) Alpha-2 adrenergic receptorsubtypes indicated by [
3HI yohimbine binding in human brain.Life Sci. 38, 2129—2137.
Piletz J. E. and Halaris A. (1988) Super high affinity [3H]-para-aminoclonidine binding to platelet adrenoceptor in depression.Prog. Neuropsychopharmacol. Biol. Psychiatry 12, 541—553.
Piletz J. E. and Halaris A. (1994) Psychopharmacology of imidazo-line and a
2-adrenergic receptors: implications for depression.Crit. Rev. Neurobiol. 9, 29—66.
Piletz J. E., Schubert D. S. P., and Halaris A. (1986) Evaluation ofstudies of platelet a2-adrenoceptors in depressive illness. L(feSci. 39, 1589—1616.
Piletz J. E., Halaris A., Saran A., and Marler M. (1990) Elevated3H-para-amino-clonidine binding to platelet purified plasmamembranes from depressed patients. Neuropsychopharmacol-ogy 3,201—210.
Piletz J. E., Halaris A., Saran A., and Marler M. R. (1991) Desipra-mine lowers tritiated para-aminoclonidine binding in plateletsof depressed patients. Arch. Gen. Psychiatry 48, 813—820.
Piletz J. B., Halaris A., Nelson J., Qu Y., and Ban M. (1996) PlateletI~-imidazolinebinding sites are elevated in depression but notgeneralized anxiety disorder. J. Psychiatry Res. 30, 147—168.
Sastre M. and Garcia-Sevilla J. A. (l994a) a2-Adrenoceptor sub-
types identified by [3H]RX82 1002 binding in the human brain:
the agonist guanoxabenz does not discriminate different formsofthe predominant a2A subtype. J. Neurochem. 63, 1077—1085.
Sastre M. and GarcIa-Sevilla J. A. (1994b) Density of alpha-2Aadrenoceptors and G~proteins in the human brain: ratio of high-affinity agonist sites to antagonist sites and effect of age. J.Pharmacol. Exp. Ther. 269, 1062—1072.
Smith C. B., GarcIa-Sevilla J. A., and Hollingsworth P. J. (1981)a2-Adrenoreceptors in rat brain are decreased after long-term
tricyclic antidepressant drug treatment. Brain Res. 210, 413—418.
Starke K. and Montel H. (1973) Alpha-receptor-mediated modula-tion of transmitter release from central noradrenergic neurons.Naunyn Schmiedebergs Arch. Pharmacol. 279, 53—60.
Svensson T. H., Bunney B. S., and Aghajanian G. K. (1975) Inhibi-tion of both noradrenergic and serotonergic neurones in thebrain by the a-adrenergic agonist clonidine. Brain Res. 92,291—306.
Tunstall S., Shaw J. S., McPheat J. C., and Tatum C. (1996) Bindingto the high- and low-affinity states of rat recombinant a2-adre-noceptor subtypes. Br. J. Pharmacol. 119, 358P.
Vauquelin G., De Vos H., De Backer J.-P., and Ebinger G. (1990)Identification of a2 adrenergic receptors in human frontal cortexmembranes by binding of [
3H]RX82 1002, the 2-methoxy ana-log of [3Hlidazoxan. Neurochem. mt. 17, 537—546.
.1. Neurochem., vol. 70, No. 3, 1998












![Adrenoceptor Agents [Compatibility Mode]](https://img.pdfslide.net/doc/110x75/577d26cc1a28ab4e1ea236f9/adrenoceptor-agents-compatibility-mode.jpg)
















