Embed Size (px)
Citation preview

S
Sa
b
c
d
a
ARA
KITTNIC
I
owaaif4
Nti
Ca
tK
0h
Immunobiology 218 (2013) 772– 779
Contents lists available at SciVerse ScienceDirect
Immunobiology
j o ur nal homep ag e: www.elsev ier .com/ locate / imbio
erial cytokine expressions in infants with incontinentia pigmenti�
ui-Ling Liaoa, Shen-Hao Laib, Jing-Long Huangc, Ni-Chung Leed, Wen-I Leec,∗
Department of Pediatrics, Chang Gung Memorial Hospital at Keelung, TaiwanDivision of Pulmonology, Department of Pediatrics, Chang Gung Memorial Hospital and Chang Gung University, College of Medicine, Taoyuan, TaiwanDivision of Allergy, Asthma, and Rheumatology, Department of Pediatric, Chang Gung Memorial Hospital and Chang Gung University, College of Medicine, Taoyuan, TaiwanDepartment of Pediatrics and Medical Genetics, National Taiwan University Hospital, Taipei, Taiwan
r t i c l e i n f o
rticle history:eceived 27 December 2011ccepted 26 August 2012
eywords:ncontinentia pigmentiumor necrosis factor alphaoll-like receptorEMO
nterleukin-6ytokine
a b s t r a c t
NF-�B dysfunction resulting from NEMO (NF-kappaB essential modulator) mutation can lead to sig-nificant alterations in cytokine production. However, little is known about changes in the expression ofdownstream molecules in patients with incontinentia pigmenti (IP). We aim to investigate serial cytokineexpressions during the first 2 years of life in young infants with IP, the period in which skin inflammationand morphological changes are most significant. Gene analysis and X-inactivation test were performedfor the two neonates with IP. Peripheral mononuclear cells were obtained after birth and successively at6-month interval up to the age of two years. Levels of TNF-� and IL-6 were analyzed with ELISA beforeand after stimulating with Toll-like receptor ligands. The result showed the male IP patient had normalNEMO allele. His cytokine level, although initially lower, had returned to a level comparable with thoseof controls at 12 months of age. The female infant had mutated NEMO gene. Her baseline TNF-� levelwas significantly higher than those of the control subjects at birth and remained high by 6 months of age.
All cytokine responses had decreased significantly by 2 years of age, the time in which all vesicular skinlesions had resolved. Both infants had normal serum immunoglobulin level and remained infection freeduring the follow up period. To our knowledge, this is the first report that demonstrates serial changes ofcytokine profiles in humans with IP. This study showed that in the presence of NEMO mutation, alterationof cytokine production was remarkable during the first year of life, which may account for the prominentinflammatory changes in skin morphology.ntroduction
Incontinentia pigmenti (IP) is a rare X-linked dominant gen-dermatosis. It is usually lethal in majority of affected males,hile the heterogenous female survive with characteristics of
bnormal skin pigmentation, dental abnormalities, nail dystrophy,nd opthalmologic or central nervous system deficits. Mutations
nvolving the NEMO (NF-�B essential modulator) gene accountsor 80% of cases of IP, mostly consisting of the deletion of exons–10. (Aradhya and Nelson 2001; Smahi et al. 2000) The skin defectAbbreviations: IP, incontinentia pigmenti; NF-�B, nuclear factor kappa B; NEMO,F-kappaB essential modulator; IKK, Ikappa kinase; TNF-�, tumor necrosis fac-
or alpha; IL-6, interleukin-6; IL-8, interleukin-8; IL-1B, interleukin-1 beta; iNOS,nducible nitric oxide synthases.� This work was financially supported by Chang-Gung Memorial Hospital (GrantsMRPG270162 and CMRPG270163), and NSC 98-2314-B-002-017 from Dr. N.C. Leend 99-2314-B-182-003-MY3 from Dr. W.I. Lee.∗ Corresponding author at: Division of Pediatric Allergy, Asthma, and Rheuma-
ology, Department of Pediatrics, Chang Gung Children’s Hospital, 5 Fu-Hsin Street,weishan 333, Taoyuan, Taiwan. Tel.: +886 3 3281200x8206; fax: +886 3 3288957.
E-mail address: [email protected] (W.-I. Lee).
171-2985/$ – see front matter © 2012 Elsevier GmbH. All rights reserved.ttp://dx.doi.org/10.1016/j.imbio.2012.08.280
© 2012 Elsevier GmbH. All rights reserved.
is characterized by four distinct dermatological stages, with thetransition of three stages during the first year of life. Lesions beginshortly after birth with inflammation and blister formation (vesicu-lobullous stage or stage (1) followed by verrucous hyperkeratoticlesions (verrucous stage or stage (2), with peak onset between 2and 6 weeks of life, which subsequently evolve and disappear overtime. Stage 3 (hyperpigmented stage) begins around 12–36 weeksof age with typical lesions of brownish pigmentations that followthe lines of Blaschko. The lesions may persist for several yearsand proceed to stage 4 by adolescence characterized by atrophichypopigmentation (hypopigmented/atrophic stage) (Berlin et al.2002; Makris et al. 2000).
NEMO and IKK-2 subunits of IKK (Ikappa kinase), although thelatter is of less importance, are the key components for the inflam-matory process that activates the expression of nuclear factor (NF)�B. NF-�B signaling pathway plays an essential role in the initi-ation, amplification, and resolution of inflammation and preventscell apoptosis against tumor necrosis factor � (TNF-�). By respon-
ding to several stress and immune stimuli, it controls the expressionof dozens of mediators of inflammation, such as IL-1�, IL-6, IL-8,TNF-�, or iNOS (Courtois and Israel 2000; Courtois and Smahi 2006;Sebban and Courtois 2006).
S.-L. Liao et al. / Immunobiology 218 (2013) 772– 779 773
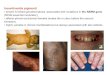
Fig. 1. Classical blistering and plaque like lesions in patient F at day 2, mostly at the extremities (a); persistence of scattered vesicular lesions by age 6 months, specially ont was
p wing
d
tdpaiiaHfo2wat
bpwi
he scalp (b). By age of 12 months, except for whorls of hyperpigmentation, the skineg-shaped teeth by the age of 2 years (d). Clinical features of the male patient shoay 2 (e and f).
Loss-of-function of NEMO gene causes complete inhibition ofhe transcriptor factor NF-�B, which results in prenatal lethalityue to TNF-induced liver apoptosis, as seen in males. Hypomor-hic mutations of the NEMO gene cause IP in heterogenous femalesnd hypohidrotic ectodermal dysplasia with immunodeficiencyn males. The defect was attributed to impaired NF-�B activ-ty affecting T and B cell function as well as the production ofnti-inflammatory cytokine production in response to infection.owever, albeit an impaired NF-�B activity, the heterozygous
emale mouse, model of IP, showed increased expression of numer-us downstream cytokine and chemokines (Jean-Baptiste et al.002; Mancini et al. 2008; Kenwrick et al. 2001). Nevertheless,hether similar findings are observed in human IP is not yet known,
nd the systemic expression of numerous mediators of inflamma-ion as part of the innate immunity has yet to be determined.
We hypothesized that similar TNF signaling disruption may also
e found in humans with IP during the early months of life. Thiseriod is characterized by severe skin inflammation and is alsohen most dermatological changes take place. In this study, wenvestigate serial TNF-� alternation in humans with IP.
free of any vesicle or verrucous formation (c). She had features of hypodontia withoverlapping of vesicular, verrucous and streaks of brownish hyperpigmentation by
Materials and methods
Case presentation
Case 1 (patient F)This full-term female neonate was born to a G2P2 mother via
Caesarean section (due to previous C/S). Prenatal examination wasnormal and perinatal course was uneventful. Family history wasnegative for IP. At birth, multiple erythematous blisters rangingfrom 1 mm to 1 cm (stage 1 IP) were scattered over the body sur-face, particularly on the four extremities, while sparing the face(Fig. 1a). Complete blood count showed eosinophilia of 1430/mm3.Both blood culture and viral survey yielded negative results. Skinbiopsy confirmed the diagnosis of IP and DNA analysis by PCRyielded the specific 1.8-kb PCR product diagnostic for a commonexon 4–10 deletion of the NEMO gene (Fig. 2). By the third day of
life, the blisters were oozing so severely that the patient remainedhospitalized for another week for intensive wound care. Her nailswere intact, but alopecia was found at the vertex of the scalp. By theage of 6 months, most of the skin lesions had advanced to verrucous
774 S.-L. Liao et al. / Immunobiology 218 (2013) 772– 779
Fig. 2. True NEMO gene deletion of exons 4–10 was selectively amplified by the primer pairs of In2 and JF3 producing a 2.6 kb band. Pseudogene deletion of exons 4–10 wasselectively amplified by the primer of Rev2 and JF3 resulting in a 2.5 kb product. The female IP patient was identified as having true NEMO gene deletion of exons 4–10, notthe pseudogene, compatible with the 2.6 kb, but not the 2.5 kb amplification product as shown in the left panel. The right panel shows the product from the primers of Rev3Sand L2Rev as the internal control (A). X chromosome inactivation pattern in the whole blood was performed by PCR amplification using the highly polymorphic exon-1 ofthe Humara gene obtained from females. It is comprised of two major products that can be separated on gel electrophoresis. After digestion with HpaII, an enzyme whichcuts active, nonmethylated DNA, its PCR products would represent only the inactivated, nondigested, X-chromosome. When preferential X inactivation is present, one oft and do e IP pt l X-ch
supmtofhima
C
upItspbbadsol
he two X-chromosome is activated in all the cells and the corresponding Humara bther X-chromosome, which is always inactive, is unaffected. The gel on the femalhe healthy control showed the persistence of both bands indicating no preferentia
tage (stage 2 IP), with persistent scattered eruption of vesiculob-llous lesion on the scalp and trunk (Fig. 1b). Streaks of brownishigmentation (stage 3 IP) had also appeared, and were distributedostly on the lower limbs. By the age of 12 months, except for
he whorls of hyperpigmentation, the skin was free of any vesicler verrucous formation (Fig. 1c). By the age of 24 months, besidesrom features of persistent hyperpigmention and alopecia, she alsoad hypodontia with peg-shaped teeth (Fig. 1d). Serial brain imag-
ng study and ophthalmologic examinations were normal by 24onths of age. Her developmental milestone was adequate for her
ge.
ase 2 (patient M)This full-term male neonate was born to a G2P2 mother by nat-
ral spontaneous delivery. Perinatal examination was normal anderinatal course was uneventful. Family history was negative for
P, neurological disorders, or miscarriages. Soon after birth, scat-ered vesicular skin lesions were noted on all four extremities andome along the abdomen. Streaks of brownish pigmentation wereresent following the lines of Blaschko (Fig. 1e and f). The survey foracterial and viral studies were negative. Skin biopsy, as describedy Fraitag et al. (2009), showed moderate acanthosis with dysker-totic cells, and massive infiltration of eosinophils, confirming the
iagnosis of IP. DNA analysis showed normal NEMO allele with nopecific mutation sites (Fig. 2). Except for the hyperpigmentationsn the skin, all vesicles had resolved rapidly within the first week ofife. Serial brain imaging study and ophthalmologic examinationsisappears after digestion with HpaII. It is noted that the band corresponding to theatient showed Humara bands skewed to one X chromosome allele (B). In contrast,romosome inactivation.
were normal by 18 months of age and the patient did not sufferfrom frequent or serious infections.
NEMO gene analysis
Protocols were approved by the Chang Gung ethics commit-tee, and informed consents were obtained from the parents/legalguardians of the infants. Genomic DNA was prepared by phenol-chloroform extraction. RNA was prepared by means of Trizolreagent and converted to cDNA by Superscript reverse transcriptasePCR system (invitrogen) according to the manufacturer’s recom-mendations. The following NEMO specific primers were used toevaluate the full cDNA of both patients: CCC TTG CCC TGT TGG ATGand ACC CTC CAG AGC CTG GCA TTC (covering exon 2–4), and GACAAG GCC TCT GTG AAA GC and GGA GAG GAA AGC GCA GAC T (cov-ering exon 4–10). To confirm that identified NEMO mutation wasnot the NEMO pseudogene, genomic DNA was prepared by usingthe primers of GAG GAC CAA TAC CGA GCA TC (In2) and CTC GGAGAC ACA GGA ACC AGC A (JF3) to selectively amplify true NEMOdeletion as the 2.6 kb product, and by GCC ATC TGT TTT TGC GTG TG(REV-2) and JF3 for identifying pseudogene deletion as previouslydescribed(Bardaro et al. 2003) Internal control was also amplifiedby CTC TTT TGA CAA GAA CAC CGG A (Rev3S) and TCG GAG ACACAG GAA CCA GCA (L2Rev).
X-chromosome inactivation
Three hundred nanograms of purified genomic DNA weredigested in 100 ml volume of buffered solution with 6U RsaI (New

S.-L. Liao et al. / Immunobiology 218 (2013) 772– 779 775
Table 1Serum immunoglobulin and IgG subclass level.
Patient M Patient F Unit
1 month 12 months 1 month 6 months 12 months
IgG 1130 793 415 (251–906) 242 (215–704) 361 (294–1069) mg/dlIgA <24.9 40.4 <24.9 (1.3–53) <24.9 (8.1–68) 13.8 (16–84) mg/dlIgM 63.4 172 34.6 (20–87) 54.5 (35–102) 68.1 (41–149) mg/dlIgG1 771 681 336 (190–620) 201 (190–620) 270 (190–620) mg/dlIgG2 346 154 78.3(30–140) 58.9 (30–140) 63.1 (30–140) mg/dlIgG3 15 48.9 11.3 (9–62) 9.64 (9–62) 13.6 (9–62) mg/dlIgG 29.1 13.9 19.5 (6–63) 33.6 (6–63) 45.7 (6–63) mg/dl
P
EiwTfpTf6Atat
S
1fsia(
T
h(wg((G(slaekM
wwrh
mso
4
IgE1 <18 <18 <18
atient M: male IP patient; patient F: female IP patient; month: age by months.
ngland Biolabs, Beverly, MA, USA) at 37 ◦C overnight and dividednto 2 fractions of 50 ml each. One aliquot was further digested
ith HpaII (New England Biolabs) and incubated overnight at 37 ◦C.he exon 1 of the polymorphic locus ‘Humara’ was amplified bothrom RsaI and RsaI–HpaII digested DNA samples, using the forwardrimer TGC GCG AAG TGA TCC AGA ACC and the backward primerGG GCT TGG GGA GAA CCA TCC. The amplification was performedor 38 cycles with a Perkin Elmer 480 thermocycler (95 ◦C 1 min,0 ◦C 30 min, and 74 ◦C 30 min) and Pfu polymerase (Stratagene,msterdam, NL), as previously described(Wengler et al. 1997) For
he analysis, 15 �l of each amplified sample was run on a 10% poly-crylamide gel with 10% urea. The gel was stained with silver nitrateo evaluate skewed X-chromosome inactivation.
tudy designs
Blood test from the female patient was withdrawn serially after8 days, 6 month, 12 month, 18 month, and 24 months of age, androm the male patient after the 8 days and 12 months of age. Controlamples were obtained from the cord blood of 20 healthy full termnfants and 20 adults volunteers (ages 20–35 years). Healthy infantsged 5–20 days (indicated as <1 month), 6, 12, 18, and 24 months8–10 in each age group) were also enrolled as control subjects.
LR1-TLR2, TLR3 and TLR4 ligand stimulation
Peripheral blood mononuclear cells (PBMC) were isolated fromeparinized venous blood by centrifugation of Ficoll-HypaquePharmacia Biotech, Piscataway, NJ). RPMI 1640 supplementedith 10% fetal calf serum (FCS; Hyclone, Logan, UT), 2 mmol/l
lutamine, 100 U/ml penicilline, and 100 �g/ml of streptomycincomplete media) was used for experiments. Toll-like receptorTLR) ligands used for cell stimulation were obtained from Invivo-en (San Diego, CA), which included synthetic bacterial lipoprotein
PAM3csk4) for TLR1-2 at 10 �g/ml; a synthetic analog of double-tranded RNA for TLR3 (poly I:C) at 20 ng/ml; and ultrapureipopolysaccharide (LPS) for TLR4 at 10 �g/ml. Medium withoutny added ligand is used to determine any baseline production ofach cytokine. As a positive control, cells were treated with the NF-B activator, phytohemagglutinin (PHA; Murex Pharmaceuticals,issouri) at 4 �g/ml in R10-FBS.To determine TLR responses, 3 × 105 PBMCs in 100 �l R10-FBS
ere added to each of the duplicate ligand- or medium-containingells and incubated at 37 ◦C for 20 h with 5% CO2. All assay prepa-
ations were performed using sterile technique in a laminar flowood.
TNF-� and IL-6 protein levels in culture supernatant wereeasured by enzyme-linked immunosorbent assay (ELISA; R&D
ystems, MN) according to the manufacturer’s directions. The limitf detection was 15.6 pg/ml for TNF-� and 3.12 pg/ml for IL-6.
<18 <18 IU/ml
Results
Analysis of exon 4–10 deletion in true NEMO gene andX-chromosome inactivation
The female IP infant carried a 2.6 kb band product, indicativeof a deletion involving exons 4–10 of the true NEMO gene. Itwas not a pseudogene because of the absence of a 2.5 kb product(Fig. 2A). The skewed X-inactivation assay revealed the presenceof heterozygous alleles from X chromosome in each nondigestedsample from the female subject. After the DNAs were digested witha methylation-sensitive restriction enzyme, HpaII, only one of theheterozygous alleles was detectable at around 270-bp, as shown inFig. 2B. The result indicated a highly skewed X-inactivation patternin favor of the mutant chromosome. The male IP patient did nothave detectable NEMO mutation or X-inactivation pattern.
Screening of humoral immune system
Due to the fact that defective NF-�B activation can influencehumoral immune system, we had examine whether B cell functionwas affected in our patients with IP. The results showed that bothpatients had normal immunoglobulin level (adjusted for age), witha normal capacity for class switching as well (Table 1).
Abnormal TLR function in patients with incontinentia pigmenti
Derangement in multiple cytokine production would beexpected during defective NF-�B signaling, but how they respondto external stimuli in the presence of heterozygote NEMO muta-tion has yet to be determined. Cytokine production was assessedfor the female patient several days after birth during which skinlesions were most severe. Analysis for the male patient and cordblood from 20 healthy neonates, and 20 adults were also per-formed for comparison. The baseline TNF-� level of the male patientwas below detection limit. And his cytokine responses, thoughdecreased compared to healthy control subjects, were capable torespond to several TLR ligands. In contrast, the baseline levels ofTNF-� and IL-6 production were profoundly increased in the femalepatient, probably reflecting a pre-existing increased expression ofthese inflammatory cytokines even before stimulation. Except forTLR4 stimulated response, cytokine reaction to several other TLRligands was also greatly increased, demonstrating a hyperrespon-sive state in the female patient with IP (Fig. 3).
Transition of cytokine production during the first two years of life
Cytokine levels of the male patient were similar to those of con-
trolled subjects when he reached the age of 12 months (data notshown). By the time when the female patient reached 6 monthsof age, there were still scattered vesiculobullous lesions over herscalp and trunk. As for the cytokine response, baseline level of
776 S.-L. Liao et al. / Immunobiology 218 (2013) 772– 779
Baseline
M F CB Adult0
600
1200
1800
2400
3000PHA
M F CB Adult0
5000
10000
15000
20000
25000
TLR1-2
M F CB Adult0
6000
12000
18000
24000
30000
TLR3
M F CB Adult0
600
1200
1800
2400
3000
TLR4
M F CB Adult0
6000
12000
18000
24000
30000
B
A
TLR1-2
M F CB Adult0
300
600
900
1200
1500TLR3
M F CB Adult0
200
400
600
800
1000
TLR4
M F CB Adult0
500
1000
1500
2000
2500
M F CB Adult0
200
400
600
800
1000Baseline PHA
M F CB Adult0
1000
2000
3000
4000
5000T
NF
– α (p
g/m
l)IL
-6 (
pg
/ml)
Fig. 3. Comparison of cytokine responses between neonates with IP, and healthy newborns or adults. Peripheral mononuclear cell were stimulated with medium for baselinecytokine level, or stimulated with TLR1-2, TLR3, TLR4 ligands, and PHA as described in the methods. TNF-� (A) and IL-6 (B) level are shown for the male patient (bar M), thef ult dop
Tcysbr
Tfi
D
stai
emale patient (bar F), and cord blood from 20 healthy newborns (bar CB), and 20 adhytohemagglutinin.
NF-� had remained high, while other TLR and PHA stimulatedytokine production had decreased prominently. By the age of 1ear, when all necrotic skin lesions had resolved, both baseline andtimulated cytokine production had decreased to levels compara-le to those of age-matched controls. These cytokine responses hademained commensurate until the age of 2 years (Fig. 4).
Collectively, these findings suggest that increased baselineNF-� level may be correlated with severity of skin lesions, buturther studies are required to determine whether this increments responsible for the onset or initiation of dermal inflammation.
iscussion
The NF-�B transcription factor is central to the regulation of
everal expression of genes related to immunity and inflamma-ion, which includes cytokines, chemokines, adhesion molecules,nd regulators of apoptosis and cellular proliferation. It is also crit-cal in the regulation of dermal homeostasis as demonstrated bynors. Values represent means ± SD for healthy donors TLR: toll-like receptor; PHA:
complex skin pathology caused by the disruption of the NEMO genein patients with IP. This may have resulted from an up-regulation ofmultiple pro-inflammatory cytokines and recruitment of immunecells into the skin (Pasparakis et al. 2002; Stratis et al. 2006; Sebbanand Courtois 2006). Moreover, among all these cytokines, a TNF-mediated inflammatory response seemed indispensable for thedisease pathogenesis seen in mice. We hypothesized that similarTNF signaling disruption may also be found in humans with IP dur-ing the early months of life. This period is characterized by severeskin inflammation and is also when most dermatological changestake place. We first investigated whether serial TNF-� expressionsthrough TLR signaling would parallel with the dermatologic mani-festations in humans with IP.
Phenotypic consequence of a defective NF-�B activity is more
severe and prominent in hypomorphic gene mutation (such as inanhidrotic ectodermal dysplasia and immunodeficiency: EAD-IP).While in patients with IP, the complete loss of function of NF-�Bactivity results in prenatal male demise and skewed X-inactivation
S.-L. Liao et al. / Immunobiology 218 (2013) 772– 779 777
0
6000
12000
18000
24000
30000
36000
24 m/o18 m/o12 m/o6 m/o<1 m/o0
500
1000
1500
2000
2500
3000
24 m/o18 m/o12 m/o6 m/o<1 m/o
0
6000
12000
18000
24000
30000
36000
<1 m/o 6 m/o 12 m/o 18 m/o 24 m/o
Baseline
TLR4
TLR1-2
0
5000
10000
15000
20000
25000
24 m/o18 m/o12 m/o6 m/o<1 m/o
PHA
B
0
250
500
750
1000
1250
1500
24 m/o18 m/o12 m/o6 m/o<1 m/o0
200
400
600
800
1000
1200
24 m/o18 m/o12 m/o6 m/o<1 m/o
0
500
1000
1500
2000
2500
3000
24 m/o18 m/o12 m/o6 m/o<1 m/o
0
1000
2000
3000
4000
5000
24 m/o18 m/o12 m/o6 m/o<1 m/o
TLR1-2Baseline
TLR4 PHA
TN
F–α
(p
g/m
l)
A
IL-6
(p
g/m
l)
Fig. 4. Serial changes in cytokine response profiles during the first two years of life. TNF-� (A) and IL-6 (B) production for the female patient at ages 18 days, 6, 12, 18, and 24months of age. The gray regions represent ± 5 SD from the mean for 8–10 control infant donors (aged 5–20 days (indicated as <1 month), 6 months (6 months), 12 months(12 months), 18 months (18 months), and 24 months (24 months) of age.

7 obiolo
in(dpptfsiett2we3tsIi1ANm
riaiMdoIiInIahTorsckabicW�wpswhsep
aOsot
78 S.-L. Liao et al. / Immun
n females. Our male patient exhibited classical sequential cuta-eous linear eruptions of IP. The finding of a normal NEMO alleleor probably a very subtle mutation rather than NEMO that was notetected by current methodology) was not surprising, since the boyresented with very mild clinical disease and unaltered cytokinerofile. Several studies had shown that in order for male patientso survive, most had to have a normal or at least a mosaicismor the NEMO alleles, or the existence of an extra set of chromo-ome such in Klinefelter’s Syndrome, although the later is rarelynvolved in IP (Fusco et al. 2007; Pacheco et al. 2006; Kenwrickt al. 2001). Our male patient had XY genotype, so this was nothe case. Due to the fact that some studies have reported coexis-ence of IP and EDA-ID in male patients with IP (Mansour et al.001; Chang et al. 2008), and a non-classical IP female patientho also suffered transient immunodeficiency (Martinez-Pomar
t al. 2005), we surveyed both patients’ immune status through-year-period observation for repeated infections, and laboratoryests such as cytokine response to Toll-like receptor ligands anderum immunoglobulin level. Although TNF-� response in the maleP patient was initially lower at birth, the cytokine production hadncreased to levels comparable to those of age-matched controls by2 months of age, and he did not suffer from any serious infections.ltogether, these findings further suggested a normal functioningF-�B pathway in the male IP patient without detectable NEMOutation.Theoretically, increased cytokine production is most often the
esult of an augmented NF-�B activity. But, in fact, several studiesn the mice model have shown that defective NF-�B activation isctually the primary trigger of enhanced cytokine production andnflammation in the NEMO knockout mice (Pasparakis et al. 2002;
akris et al. 2000). Moreover, the NEMO-knockout mice modelemonstrated an improved skin lesion after generalized ablationf tumor necrosis factor receptor 1 (TNFR1) (Nenci et al. 2006).n accordance to the findings from the mouse model, the femalenfant with NEMO mutation showed abnormally high TNF-� andL-6 levels along with severe cutaneous inflammation during theeonatal period. We suggested that the initially increased level of
L-6 might be the result of severe skin inflammation, because by thege of 6 months, IL-6 level had decreased prominently, as most ofer blister lesions had resolved. The mechanism of persistently highNF-� level could be explained by the proposed theory that necrosisf IKK�− cells occurred after birth, while triggering inflammatoryesponse and secreting cytokines such as TNF-�. Since NF-�B weretill expressed by IKK�+ cells in the female heterozygote, the sameytokine appeared to work in an autocrine manner in attempt toill the neighbor IKK�− cells that were susceptible to apoptosis. Theutocrine loop was further amplified by the cell contents releasedy IKK�− cells after death. The whole process resulted in an
ncreased TNF-� level. As a consequence, the elimination of IKK�−ells and the disappearance of necrotic skin lesions occur over time.
e presume that this might be the reason for the decreased TNF- level in our female patient when she reached 12 months of age,hen most of her skin lesions had resolved. In addition, these twoatients did not present the same stage of skin eruption at birth,uch that the female patient had stage 1 skin lesion (the time athich she had high serum TNF-� level), while the male patientad already presented with stages 2 and 3 skin lesion with a lowerum TNF-� level. These differences in skin morphology might alsoxplain the distinct cytokines expression found between the twoatients.
Although the importance of TNF has been documented in thenimal model of IP, never has it been investigated in human IP.
ur finding point to the significant role of TNF-� during the earlytage of IP in a female infant with NEMO gene mutation. Although,nly limited conclusions can be drawn from a case regardinghe disease mechanism, but we propose a longitudinal TNF-�
gy 218 (2013) 772– 779
alternation through TLR signaling, which might be helpful in for-mulating hypotheses on the pathogenesis of human IP in thefuture.
Conflicts of interest
None declared.
Acknowledgments
The authors want to acknowledge the families and volunteersfor their participation in the study.
References
Aradhya, S., Nelson, D.L., 2001. NF-kappaB signaling and human disease. Curr. Opin.Genet. Dev. 11, 300–306.
Bardaro, T., Falco, G., Sparago, A., Mercadante, V., Gean, M.E., Tarantino, E.,Ursini, M.V., D’Urso, M., 2003. Two cases of misinterpretation of molecularresults in incontinentia pigmenti, and a PCR-based method to discriminateNEMO/IKKgamma gene deletion. Hum. Mutat. 21, 8–11.
Berlin, A.L., Paller, A.S., Chan, L.S., 2002. Incontinentia pigmenti: a review andupdate on the molecular basis of pathophysiology. J. Am. Acad. Dermatol. 47,169–187.
Chang, T.T., Behshad, R., Brodell, R.T., Gilliam, A.C., 2008. A male infant withanhidrotic ectodermal dysplasia/immunodeficiency accompanied by inconti-nentia pigmenti and a mutation in the NEMO pathway. J. Am. Acad. Dermatol.58, 316–320.
Courtois, G., Israel, A., 2000. NF-kappa B defects in humans: the NEMO/incontinentiapigmenti connection. Sci. STKE 2000, e1.
Courtois, G., Smahi, A., 2006. NF-kappaB-related genetic diseases. Cell Death Differ.13, 843–851.
Fraitag, S., Rimella, A., de, P.Y., Brousse, N., Hadj-Rabia, S., Bodemer, C., 2009. Skinbiopsy is helpful for the diagnosis of incontinentia pigmenti at late stage (IV): aseries of 26 cutaneous biopsies. J. Cutan. Pathol. 36, 966–971.
Fusco, F., Fimiani, G., Tadini, G., Michele, D., Ursini, M.V., 2007. Clinical diagnosis ofincontinentia pigmenti in a cohort of male patients. J. Am. Acad. Dermatol. 56,264–267.
Jean-Baptiste, S., O’Toole, E.A., Chen, M., Guitart, J., Paller, A., Chan, L.S., 2002.Expression of eotaxin, an eosinophil-selective chemokine, parallels eosinophilaccumulation in the vesiculobullous stage of incontinentia pigmenti. Clin. Exp.Immunol. 127, 470–478.
Kenwrick, S., Woffendin, H., Jakins, T., Shuttleworth, S.G., Mayer, E., Greenhalgh, L.,Whittaker, J., Rugolotto, S., Bardaro, T., Esposito, T., D’Urso, M., Soli, F., Turco, A.,Smahi, A., Hamel-Teillac, D., Lyonnet, S., Bonnefont, J.P., Munnich, A., Aradhya,S., Kashork, C.D., Shaffer, L.G., Nelson, D.L., Levy, M., Lewis, R.A., 2001. Survivalof male patients with incontinentia pigmenti carrying a lethal mutation can beexplained by somatic mosaicism or Klinefelter syndrome. Am. J. Hum. Genet. 69,1210–1217.
Makris, C., Godfrey, V.L., Krahn-Senftleben, G., Takahashi, T., Roberts, J.L., Schwarz,T., Feng, L., Johnson, R.S., Karin, M., 2000. Female mice heterozygous for IKKgamma/NEMO deficiencies develop a dermatopathy similar to the human X-linked disorder incontinentia pigmenti. Mol. Cell 5, 969–979.
Mancini, A.J., Lawley, L.P., Uzel, G., 2008. X-linked ectodermal dysplasia with immu-nodeficiency caused by NEMO mutation: early recognition and diagnosis. Arch.Dermatol. 144, 342–346.
Mansour, S., Woffendin, H., Mitton, S., Jeffery, I., Jakins, T., Kenwrick, S., Murday,V.A., 2001. Incontinentia pigmenti in a surviving male is accompanied by hypo-hidrotic ectodermal dysplasia and recurrent infection. Am. J. Med. Genet. 99,172–177.
Martinez-Pomar, N., Munoz-Saa, I., Heine-Suner, D., Martin, A., Smahi, A., Mata-moros, N., 2005. A new mutation in exon 7 of NEMO gene: late skewedX-chromosome inactivation in an incontinentia pigmenti female patient withimmunodeficiency. Hum. Genet. 118, 458–465.
Nenci, A., Huth, M., Funteh, A., Schmidt-Supprian, M., Bloch, W., Metzger, D., Cham-bon, P., Rajewsky, K., Krieg, T., Haase, I., Pasparakis, M., 2006. Skin lesiondevelopment in a mouse model of incontinentia pigmenti is triggered by NEMOdeficiency in epidermal keratinocytes and requires TNF signaling. Hum. Mol.Genet. 15, 531–542.
Pacheco, T.R., Levy, M., Collyer, J.C., de Parra, N.P., Parra, C.A., Garay, M., Aprea,G., Moreno, S., Mancini, A.J., Paller, A.S., 2006. Incontinentia pigmenti in malepatients. J. Am. Acad. Dermatol. 55, 251–255.
Pasparakis, M., Courtois, G., Hafner, M., Schmidt-Supprian, M., Nenci, A., Toksoy, A.,Krampert, M., Goebeler, M., Gillitzer, R., Israel, A., Krieg, T., Rajewsky, K., Haase, I.,2002. TNF-mediated inflammatory skin disease in mice with epidermis-specificdeletion of IKK2. Nature 417, 861–866.
Sebban, H., Courtois, G., 2006. NF-kappaB and inflammation in genetic disease.Biochem. Pharmacol. 72, 1153–1160.
Smahi, A., Courtois, G., Vabres, P., Yamaoka, S., Heuertz, S., Munnich, A., Israel, A.,Heiss, N.S., Klauck, S.M., Kioschis, P., Wiemann, S., Poustka, A., Esposito, T., Bar-daro, T., Gianfrancesco, F., Ciccodicola, A., D’Urso, M., Woffendin, H., Jakins, T.,

obiolo
S
S.-L. Liao et al. / Immun
Donnai, D., Stewart, H., Kenwrick, S.J., Aradhya, S., Yamagata, T., Levy, M., Lewis,
R.A., Nelson, D.L., 2000. Genomic rearrangement in NEMO impairs NF-kappaBactivation and is a cause of incontinentia pigmenti. The International Inconti-nentia Pigmenti (IP) Consortium. Nature 405, 466–472.tratis, A., Pasparakis, M., Markur, D., Knaup, R., Pofahl, R., Metzger, D., Chambon, P.,Krieg, T., Haase, I., 2006. Localized inflammatory skin disease following inducible
gy 218 (2013) 772– 779 779
ablation of I kappa B kinase 2 in murine epidermis. J. Invest. Dermatol. 126,
614–620.Wengler, G.S., Parolini, O., Fiorini, M., Mella, P., Smith, H., Ugazio, A.G., Notarangelo,L.D., 1997. A PCR-based non-radioactive X-chromosome inactivation assayfor genetic counseling in X-linked primary immunodeficiencies. Life Sci. 61,1405–1411.


















![First IKBKG Gene Mutation Study in Serbian Incontinentia ... · Incontinentia pigmenti (IP; Bloch-Sulzberg-er syndrome; MIM 308300) is a rare X-linked dominant genodermatosis [5]](https://img.pdfslide.net/doc/110x75/5f3bedf5651a4c1377610355/first-ikbkg-gene-mutation-study-in-serbian-incontinentia-incontinentia-pigmenti.jpg)








