Embed Size (px)
Citation preview

PRECLINICAL STUDIES
Small-molecule inhibition of oncogenic eukaryotic proteintranslation in mesothelioma cells
Esther Z. Chen & Blake A. Jacobson & Manish R. Patel & Aniekan M. Okon &
Shui Li & Kerry Xiong & Abhishek J. Vaidya & Peter B. Bitterman &
Carston R. Wagner & Robert A. Kratzke
Received: 4 December 2013 /Accepted: 18 February 2014# Springer Science+Business Media New York 2014
Summary Deranged cap-mediated translation is implicatedin the genesis, maintenance and progression of many humancancers including mesothelioma. In this study, disrupting theeIF4F complex by antagonizing the eIF4E-mRNA-cap inter-action is assessed as a therapy for mesothelioma. Mesotheli-oma cells were treated with 4Ei-1, a membrane permeableprodrug that when converted to the active drug, 7-benzylguanosine monophosphate (7Bn-GMP) displaces cappedmRNAs from the eIF4F complex. Colony formation wasmeasured in mesothelioma treated with 4Ei-1 alone or com-bined with pemetrexed. Proliferation was examined in cellstreated with 4Ei-1. Binding to a synthetic cap-analogue wasused to study the strength of eIF4F complex activation inlysates exposed to 4Ei-1. 4Ei-1 treatment resulted in a dosedependent decrease in colony formation and cell viability.Combination therapy of 4Ei-1 with pemetrexed further re-duced colony number. Formation of eIF4F cap-complex de-creased in response to 4Ei-1 exposure. 4Ei-1 is a novelprodrug that reduces proliferation, represses colony forma-tion, diminishes association of eIF4F with the mRNA cap,and sensitizes mesothelioma cells to pemetrexed.
Keywords Mesothelioma . eIF4E . 4Ei-1 . 7-benzylguanosinemonophosphate . Cap-dependent translation
Introduction
Malignant mesothelioma, which in the United States is re-sponsible for the deaths of 3,000 people annually, arises fromthe serosal lining of the pleural or peritoneal cavity and is notcurable with existing therapies [1]. Cisplatin and pemetrexedin combination is the current standard of care for unresectablemesothelioma and leads to an overall response rate of 22 %, amedian time to progression of 7 months and overall survivalof 12 months [2]. There is a clear need for more effectivetherapeutic strategies for this fatal disease.
The eIF4F complex is comprised of the RNA helicaseeIF4A, the scaffolding protein eIF4G and eIF4E, the cap-binding protein. Each newly synthesized transcript is modifiedby the insertion of a 7-methylguanosine cap at their 5′ end.eIF4E binds to the cap at the 5′ end of the mRNA, eIF4Gstabilizes the eIF4E-cap interaction and RNA helicase eIF4Aunwinds the secondary structure of the mRNA allowing accessof the 43s preinitiation complex to the start codon. eIF4E isconsidered to be the rate-limiting component for the assemblyof the eIF4F complex [3, 4]. During normal physiologic con-ditions the eIF4E binding proteins (4E-BP) negatively regulatetranslation initiation by sequestering eIF4E from eIF4G. Whenthe 4E-BP1s are phosphorylated by the Ras/phosphoinositide3-kinase (PI3K)/AKT/mammalian target of rapamycin(mTOR) kinase cascade, they dissociate from eIF4E, enablingeIF4F assembly and cap-dependent translation under permis-sive growth conditions [5–7]. Throughmechanisms that remainto be fully elucidated eIF4F hyperactivation, leads to selectivetranslation of mRNAs involved in cell growth, cell survival,angiogenesis and metastasis [8, 9]. This enhanced eIF4F po-tency can result from an increase in eIF4E expression per se or
E. Z. Chen :B. A. Jacobson :M. R. Patel :K. Xiong :A. J. Vaidya :P. B. Bitterman :R. A. KratzkeDepartment of Medicine, University of Minnesota, Minneapolis,MN 55455, USA
A. M. Okon : S. Li : C. R. WagnerDepartment of Medicinal Chemistry, University of Minnesota,Minneapolis, MN 55455, USA
R. A. Kratzke (*)Division of Hematology, Oncology and Transplantation,University of Minnesota Medical School, MMC 480 420Delaware St, SE, Minneapolis,MN 55455, USAe-mail: [email protected]
Invest New DrugsDOI 10.1007/s10637-014-0076-7

by constitutive activation of upstream positive regulators (e.-g.the PI3K/AKT/mTOR pathway) or by both - a commonfinding in most cancers including mesothelioma.
Hyperactivated eIF4F can enhance multiple oncogenic path-ways and it is for this reason that dysregulation of cap-dependent translation is commonly observed in tumors andparticipates in the genesis, progression and maintenance ofsome cancers. Indeed, forced expression of eIF4E in fibroblastsand primary epithelial cells is sufficient to induce transforma-tion [10, 11]. Additionally, eIF4E levels are frequently elevatedin malignancies of the prostate, lung, breast, stomach, colon,skin as well as cancers of the hematopoietic system [12–18].Further, tumor cells of different types commonly manifestincreased levels of eIF4G [19] and eIF4A [20]. These observa-tions support the notion that cancers need to sustain activationof eIF4F to drive translation of the subset of mRNAs requiredfor themalignant phenotype.With this inmind, eIF4F can be anappealing therapeutic target that functions at the intersection ofcellular pathways controlling malignancy.
Therapies that target oncogenic eukaryotic translation pro-duced by eIF4F hyperactivation have been investigated incancer. eIF4F suppression by forced overexpression ofhypophosphorylated 4E-BP1 led to decreased tumorigencityof xenograft models in breast [11] and lung cancer [13].Evidence suggests that constitutive activity of cap-mediatedtranslation exists in mesothelioma cell lines [21] and forcedexpression of a hypophosphorylated 4E-BP1 suppressedtumorigencity of mesothelioma xenografts [22]. These resultsdemonstrate that targeting cap-dependent translation has po-tential to be a potent therapy for mesothelioma.
Directly targeting eIF4F integrity and function in differentcancer types has provided evidence for therapeutic efficacy.For example, studies that employ small molecule inhibition ofthe eIF4E - eIF4G interaction [23, 24], peptides directed to theeIF4E - eIF4G interaction site [25, 26], and antisense oligo-nucleotide [27] or short-interfering RNA decreasing eIF4Eprotein levels [28, 29] have all led to attenuation of themalignant phenotype. Pharmacological inhibition of eIF4E -mRNA cap interaction is another way to antagonize oncogen-ic eIF4F hyperactivation and has been studied in lung andbreast cancer cells [30]. This approach uses a synthetic nucle-otide derivative, an analog to the cap (Me7GTP), to block thebinding of eIF4E to mRNA cap. To alleviate the difficultiesconnected with employing nucleotides as drugs, apronucleotide strategy has been developed. This work led tothe discovery of 4Ei-1, a prodrug that is converted into theactive compound, 7Bn-GMP, by the histidine triad nucleotidebinding protein-1 (Hint1) [30, 31]. In breast and lung cancer,4Ei-1 is converted to 7Bn-GMP and chemosensitizes cells tonon-toxic levels of the cytotoxic drug gemcitabine [30].
In this study, 4Ei-1 is assessed as a therapeutic agentagainst mesothelioma. 4Ei-1 potently diminished colony for-mation, repressed cap-dependent complex formation, reduced
mesothelioma cell viability, and sensitized mesothelioma cellsto pemetrexed.
Materials and methods
Cell lines and cell culture Mesothelioma cell lines H513,H2373, H2461 and H2596 were obtained from the AmericanType Culture Collection (ATCC). Cells were cultured in RPMI1640 (Life Technologies) containing 10 % calf serum(Biofluids) and 1 % antibiotic-antimycotic (Life Technologies)and maintained at 37 °C in 5 % CO2.
Cell lysate preparation Adherent cells were washed with 1 XPBS, scraped, collected and centrifuged. Cell pellets wereresuspended in five times the pellet volume of either freeze-thaw lysis buffer (50 mM Tris–HCl pH 7.5, 150 mM NaCl,50 mM NaF, 1 mM EDTA, 10 mM tetrasodium pyrophos-phate) or TNESV lysis buffer (50 mM Tris–HCl, pH 7.4; 1 %NP-40; 2 mM EDTA, pH 8.0; 0.1 M NaCl). Both lysis bufferswere supplemented with a protease inhibitor cocktail (Roche)and a phosphatase inhibitor cocktail (Sigma). The cell mixturein the freeze-thaw lysis buffer was subjected to three consec-utive freeze-thaw cycles (15 min at −80 °C followed by 2 minat 37 °C). Protein concentrations were determined by Brad-ford assay (BioRad) and stored at −80 °C.
In vitro cap-affinity assay Three hundred microlitre (1 μg/μL)of cell lysate was mixed with 50 μL of suspended (50 %mixture) m7GTP-Sepharose resin (Amersham Pharmacia Bio-tech) with and without 4Ei-1 (600 μM) for 2 h at 4 °C tocapture eIF4E and eIF4G. Captured proteins were eluted with35 μL elution buffer [25 mM Tris–HCl (pH 7.5), 150 mMKCl] containing 100 μM 7-methylguanosine 5-triphophate(Sigma-Aldrich) and prepared for immunoblotting.
Immuoblot analysis Protein samples were separated utilizing(8–15 % gradient) SDS-polyacrylamide gel electrophoresis(PAGE) and transferred to a PVDF membrane. Membraneswere blocked in 5% non-fat dry milk in TBS-Tween 20 (TBS-T) (0.15 M NaCl; 0.01 M Tris–HCl, pH 7.6; 0.05 % Tween20) for 20 h at 4 °C, rinsed, and incubated for 1 h at roomtemperature with the appropriate primary antibody. Blots wereprobed for eIF4E (Cell Signaling) and eIF4GI (Cell Signaling)both at 1:1000 dilution. Blots were washed three times for5 min in TBS-T prior to 1 h incubation with horseradishperoxidase labeled secondary antibody (SouthernBiotech)followed by three more TBS-Twashes. Detection was carriedout using ECL Plus Western Blotting System (AmershamBiosciences) to visualize the bands of interest. ImageJ, apublic domain Java processing program, was employed todetermine the density of protein bands. The density of proteinbands was normalized to the untreated condition.
Invest New Drugs
In vitro colony-forming assays H513, H2373 and H2596 cellswere seeded into 6 well plates at 500 cells per well. After 4 hcells were either left untreated or treated with 4Ei-1,pemetrexed, or 4Ei-1 and pemetrexed in combination. Whencolonies were of appropriate size (~9 d) colonies were washedwith 1 X PBS, fixed for 10 min in formalin, and stained withcoomassie blue. Images were collected, and colonies manual-ly counted. Colony numbers were expressed as the mean +/−SD normalized to untreated cells.
Cell proliferation assay Five thousand cells were seeded intowells of 96 well plates for H2373, H2461 and H2596 celllines. Following overnight incubation cells were then treatedwith varying doses of 4Ei-1 for 72 h at 37 °C. Cell viabilitywas determined by Cell Counting Kit-8 (Dojindo MolecularTechnologies). Tetrazolium substrate was added to each well,plates incubated for 2 h at 37 °C, and absorbance measured at450 nm. Cell viability values were normalized to untreatedcells. Experiments were performed in triplicate.
Results
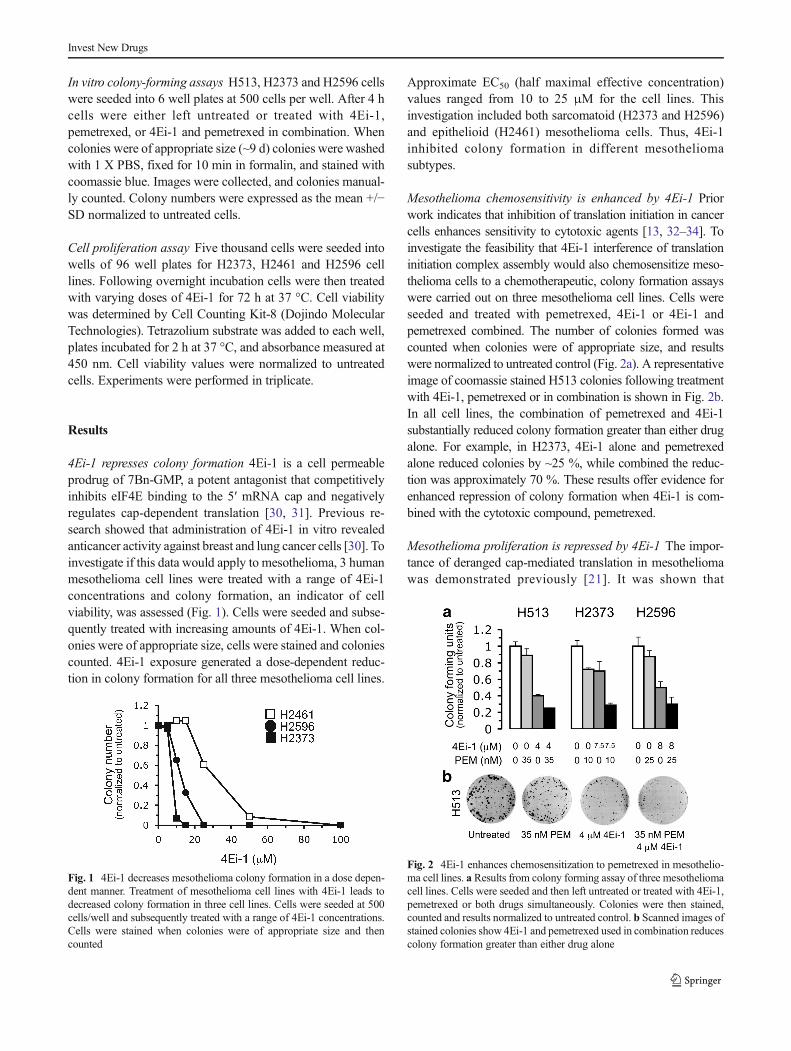
4Ei-1 represses colony formation 4Ei-1 is a cell permeableprodrug of 7Bn-GMP, a potent antagonist that competitivelyinhibits eIF4E binding to the 5′ mRNA cap and negativelyregulates cap-dependent translation [30, 31]. Previous re-search showed that administration of 4Ei-1 in vitro revealedanticancer activity against breast and lung cancer cells [30]. Toinvestigate if this data would apply to mesothelioma, 3 humanmesothelioma cell lines were treated with a range of 4Ei-1concentrations and colony formation, an indicator of cellviability, was assessed (Fig. 1). Cells were seeded and subse-quently treated with increasing amounts of 4Ei-1. When col-onies were of appropriate size, cells were stained and coloniescounted. 4Ei-1 exposure generated a dose-dependent reduc-tion in colony formation for all three mesothelioma cell lines.
Approximate EC50 (half maximal effective concentration)values ranged from 10 to 25 μM for the cell lines. Thisinvestigation included both sarcomatoid (H2373 and H2596)and epithelioid (H2461) mesothelioma cells. Thus, 4Ei-1inhibited colony formation in different mesotheliomasubtypes.
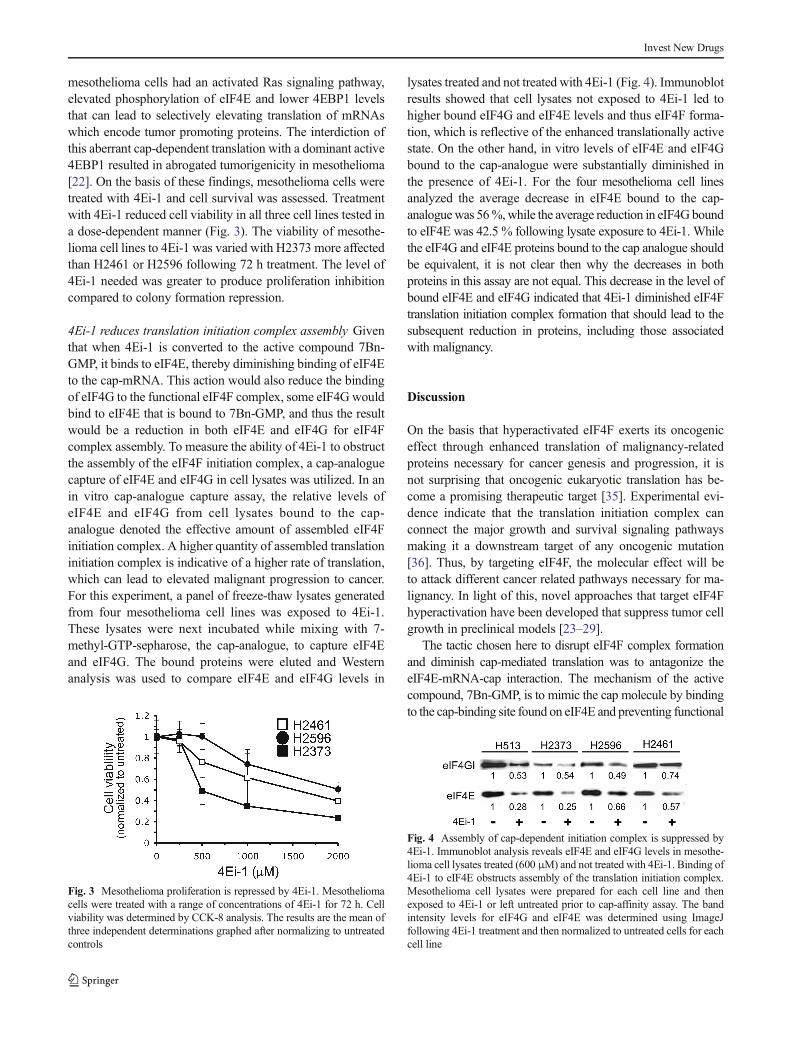
Mesothelioma chemosensitivity is enhanced by 4Ei-1 Priorwork indicates that inhibition of translation initiation in cancercells enhances sensitivity to cytotoxic agents [13, 32–34]. Toinvestigate the feasibility that 4Ei-1 interference of translationinitiation complex assembly would also chemosensitize meso-thelioma cells to a chemotherapeutic, colony formation assayswere carried out on three mesothelioma cell lines. Cells wereseeded and treated with pemetrexed, 4Ei-1 or 4Ei-1 andpemetrexed combined. The number of colonies formed wascounted when colonies were of appropriate size, and resultswere normalized to untreated control (Fig. 2a). A representativeimage of coomassie stained H513 colonies following treatmentwith 4Ei-1, pemetrexed or in combination is shown in Fig. 2b.In all cell lines, the combination of pemetrexed and 4Ei-1substantially reduced colony formation greater than either drugalone. For example, in H2373, 4Ei-1 alone and pemetrexedalone reduced colonies by ~25 %, while combined the reduc-tion was approximately 70 %. These results offer evidence forenhanced repression of colony formation when 4Ei-1 is com-bined with the cytotoxic compound, pemetrexed.
Mesothelioma proliferation is repressed by 4Ei-1 The impor-tance of deranged cap-mediated translation in mesotheliomawas demonstrated previously [21]. It was shown that
Fig. 1 4Ei-1 decreases mesothelioma colony formation in a dose depen-dent manner. Treatment of mesothelioma cell lines with 4Ei-1 leads todecreased colony formation in three cell lines. Cells were seeded at 500cells/well and subsequently treated with a range of 4Ei-1 concentrations.Cells were stained when colonies were of appropriate size and thencounted
Fig. 2 4Ei-1 enhances chemosensitization to pemetrexed in mesothelio-ma cell lines. a Results from colony forming assay of three mesotheliomacell lines. Cells were seeded and then left untreated or treated with 4Ei-1,pemetrexed or both drugs simultaneously. Colonies were then stained,counted and results normalized to untreated control. b Scanned images ofstained colonies show 4Ei-1 and pemetrexed used in combination reducescolony formation greater than either drug alone
Invest New Drugs
mesothelioma cells had an activated Ras signaling pathway,elevated phosphorylation of eIF4E and lower 4EBP1 levelsthat can lead to selectively elevating translation of mRNAswhich encode tumor promoting proteins. The interdiction ofthis aberrant cap-dependent translation with a dominant active4EBP1 resulted in abrogated tumorigenicity in mesothelioma[22]. On the basis of these findings, mesothelioma cells weretreated with 4Ei-1 and cell survival was assessed. Treatmentwith 4Ei-1 reduced cell viability in all three cell lines tested ina dose-dependent manner (Fig. 3). The viability of mesothe-lioma cell lines to 4Ei-1 was varied with H2373 more affectedthan H2461 or H2596 following 72 h treatment. The level of4Ei-1 needed was greater to produce proliferation inhibitioncompared to colony formation repression.
4Ei-1 reduces translation initiation complex assembly Giventhat when 4Ei-1 is converted to the active compound 7Bn-GMP, it binds to eIF4E, thereby diminishing binding of eIF4Eto the cap-mRNA. This action would also reduce the bindingof eIF4G to the functional eIF4F complex, some eIF4G wouldbind to eIF4E that is bound to 7Bn-GMP, and thus the resultwould be a reduction in both eIF4E and eIF4G for eIF4Fcomplex assembly. To measure the ability of 4Ei-1 to obstructthe assembly of the eIF4F initiation complex, a cap-analoguecapture of eIF4E and eIF4G in cell lysates was utilized. In anin vitro cap-analogue capture assay, the relative levels ofeIF4E and eIF4G from cell lysates bound to the cap-analogue denoted the effective amount of assembled eIF4Finitiation complex. A higher quantity of assembled translationinitiation complex is indicative of a higher rate of translation,which can lead to elevated malignant progression to cancer.For this experiment, a panel of freeze-thaw lysates generatedfrom four mesothelioma cell lines was exposed to 4Ei-1.These lysates were next incubated while mixing with 7-methyl-GTP-sepharose, the cap-analogue, to capture eIF4Eand eIF4G. The bound proteins were eluted and Westernanalysis was used to compare eIF4E and eIF4G levels in
lysates treated and not treated with 4Ei-1 (Fig. 4). Immunoblotresults showed that cell lysates not exposed to 4Ei-1 led tohigher bound eIF4G and eIF4E levels and thus eIF4F forma-tion, which is reflective of the enhanced translationally activestate. On the other hand, in vitro levels of eIF4E and eIF4Gbound to the cap-analogue were substantially diminished inthe presence of 4Ei-1. For the four mesothelioma cell linesanalyzed the average decrease in eIF4E bound to the cap-analoguewas 56%,while the average reduction in eIF4G boundto eIF4E was 42.5 % following lysate exposure to 4Ei-1. Whilethe eIF4G and eIF4E proteins bound to the cap analogue shouldbe equivalent, it is not clear then why the decreases in bothproteins in this assay are not equal. This decrease in the level ofbound eIF4E and eIF4G indicated that 4Ei-1 diminished eIF4Ftranslation initiation complex formation that should lead to thesubsequent reduction in proteins, including those associatedwith malignancy.
Discussion
On the basis that hyperactivated eIF4F exerts its oncogeniceffect through enhanced translation of malignancy-relatedproteins necessary for cancer genesis and progression, it isnot surprising that oncogenic eukaryotic translation has be-come a promising therapeutic target [35]. Experimental evi-dence indicate that the translation initiation complex canconnect the major growth and survival signaling pathwaysmaking it a downstream target of any oncogenic mutation[36]. Thus, by targeting eIF4F, the molecular effect will beto attack different cancer related pathways necessary for ma-lignancy. In light of this, novel approaches that target eIF4Fhyperactivation have been developed that suppress tumor cellgrowth in preclinical models [23–29].
The tactic chosen here to disrupt eIF4F complex formationand diminish cap-mediated translation was to antagonize theeIF4E-mRNA-cap interaction. The mechanism of the activecompound, 7Bn-GMP, is to mimic the cap molecule by bindingto the cap-binding site found on eIF4E and preventing functional
Fig. 3 Mesothelioma proliferation is repressed by 4Ei-1. Mesotheliomacells were treated with a range of concentrations of 4Ei-1 for 72 h. Cellviability was determined by CCK-8 analysis. The results are the mean ofthree independent determinations graphed after normalizing to untreatedcontrols
Fig. 4 Assembly of cap-dependent initiation complex is suppressed by4Ei-1. Immunoblot analysis reveals eIF4E and eIF4G levels in mesothe-lioma cell lysates treated (600 μM) and not treated with 4Ei-1. Binding of4Ei-1 to eIF4E obstructs assembly of the translation initiation complex.Mesothelioma cell lysates were prepared for each cell line and thenexposed to 4Ei-1 or left untreated prior to cap-affinity assay. The bandintensity levels for eIF4G and eIF4E was determined using ImageJfollowing 4Ei-1 treatment and then normalized to untreated cells for eachcell line
Invest New Drugs

eIF4F from forming. In previous work 4Ei-1 was shown to becell permeable, converted to 7Bn-GMP and to chemosensitizelung and breast cancer cells. Further, 4Ei-1 induced eIF4Eproteasomal degradation [30] in both cancer cell types.
4Ei-1 was investigated in the present study as a therapy formesothelioma. With the reliance of mesothelioma uponhyperactivated eIF4F induced by activation of eIF4E coupledwith reduced 4E-BP1 expression and activated Ras signalingpathway it is anticipated that mesothelioma cells would besensitive to 4Ei-1 [21]. Here, results demonstrate that followinginternalization of 4Ei-1 into cells that the prodrug is thenconverted to the active compound, 7Bn-GMP, where it thencan target eIF4F integrity. In mesothelioma, 4Ei-1 effectivelyreduced colony formation, suppressed cap-mediated complexformation in vitro, diminished cell proliferation, and sensitizedcells to pemetrexed. Driving the oncogenic force ofhyperactivated translation initiation in cancer cells to levels ofnormal cellular homeostasis through pharmacological interven-tion will result in reducing the abundance of oncogenic proteinsthereby reverting tumorigenesis [8]. The ability to reduce mul-tiple malignancy-related proteins should have a clear benefitover molecular targeted therapy whereby only a single onco-gene is targeted. While different cancer types will depend uponhyperactivated eIF4F to different degrees for their neoplasticproperties it is clear that, instead of targeting individual alter-ations singularly, the small-molecule interdiction at the assem-bly of eIF4F complex will target regulatory pathways in thecancer protein network and has the potential to elicit greatersustained responses. The development of small-molecule inhib-itors of oncogenic eukarotic translation, like 4Ei-1, has a greatpotential for therapeutic response in the clinical setting.
Acknowledgments We would like to thank Sabine Darling, NateRietvold and Ryan McDonald for their technical assistance.
Disclosure of Potential Conflicts of Interest All other authors declareno potential conflicts of interest.
References
1. Britton M (2002) The epidemiology of mesothelioma. Semin Oncol29:18–25
2. Carbone M, Albelda SM, Broaddus VC et al (2007) Eighth interna-tional mesothelioma interest group. Oncogene 26:6959–6967
3. Hiremath LS, Webb NR, Rhoads RE (1985) Immunological detec-tion of the messenger RNA cap-binding protein. J Biol Chem 260:7843–7849
4. Duncan R, Milburn SC, Hershey JW (1987) Regulated phosphory-lation and low abundance of HeLa cell initiation factor eIF-4Fsuggest a role in translational control. Heat shock effects on eIF-4F.J Biol Chem 262:380–388
5. Silvera D, Formenti SC, Schneider RJ (2010) Translational control incancer. Nat Rev Cancer 10:254–266
6. Thumma SC, Kratzke RA (2007) Translational control: a target forcancer therapy. Cancer Lett 258:1–8
7. Sonenberg N, Hinnebusch AG (2009) Regulation of translationinitiation in eukaryotes: mechanisms and biological targets. Cell136:731–745
8. Hsieh AC, Ruggero D (2010) Targeting eukaryotic translation initi-ation factor 4E (eIF4E) in cancer. Clin Cancer Res 16:4914–4920
9. Polunovsky VA, Bitterman PB (2006) The cap-dependent trans-lation apparatus integrates and amplifies cancer pathways. RNA Biol3:10–17
10. Lazaris-Karatzas A, Montine KS, Sonenberg N (1990) Malignanttransformation by a eukaryotic initiation factor subunit that binds tomRNA 5′ cap. Nature 345:544–547
11. Avdulov S, Li S, Michalek V et al (2004) Activation of translationcomplex eIF4F is essential for the genesis and maintenance of themalignant phenotype in human mammary epithelial cells. CancerCell 5:553–563
12. Graff JR, Konicek BW, Lynch RL et al (2009) eIF4E activation iscommonly elevated in advanced human prostate cancers and sig-nificantly related to reduced patient survival. Cancer Res 69:3866–3873
13. Jacobson BA, AlterMD, KratzkeMG et al (2006) Repression of cap-dependent translation attenuates the transformed phenotype in non-small cell lung cancer both in vitro and in vivo. Cancer Res 66:4256–4262
14. Coleman LJ, Peter MB, Teall TJ et al (2009) Combined analysisof eIF4E and 4E-binding protein expression predicts breastcancer survival and estimates eIF4E activity. Br J Cancer 100:1393–1399
15. Chen CN, Hsieh FJ, ChengYM, Lee PH, ChangKJ (2004) Expressionof eukaryotic initiation factor 4E in gastric adenocarcinoma and itsassociation with clinical outcome. J Surg Oncol 86:22–27
16. Rosenwald IB, Chen JJ, Wang S, Savas L, London IM, Pullman J(1999) Upregulation of protein synthesis initiation factor eIF-4E isan early event during colon carcinogenesis. Oncogene 18:2507–2517
17. Salehi Z, Mashayekhi F, Shahosseini F (2007) Significance of eIF4Eexpression in skin squamous cell carcinoma. Cell Biol Int 31:1400–1404
18. Wang S, Rosenwald IB, Hutzler MJ et al (1999) Expression of theeukaryotic translation initiation factors 4E and 2alpha in non-Hodgkin’s lymphomas. Am J Pathol 155:247–255
19. Bauer C, Brass N, Diesinger I, Kayser K, Grasser FA, MeeseE (2002) Overexpression of the eukaryotic translation initiationfactor 4G (eIF4G-1) in squamous cell lung carcinoma. Int J Cancer98:181–185
20. Shuda M, Kondoh N, Tanaka K et al (2000) Enhanced expression oftranslation factor mRNAs in hepatocellular carcinoma. AnticancerRes 20:2489–2494
21. Patel MR, Jacobson BA, De A et al (2007) Ras pathway activation inmalignant mesothelioma. J Thorac Oncol 2:789–795
22. Jacobson BA, De A, Kratzke MG et al (2009) Activated 4E-BP1represses tumourigenesis and IGF-I-mediated activation of the eIF4Fcomplex in mesothelioma. Br J Cancer 101:424–431
23. Moerke NJ, Aktas H, Chen H et al (2007) Small-molecule inhibitionof the interaction between the translation initiation factors eIF4E andeIF4G. Cell 128:257–267
24. Cencic R, Hall DR, Robert F et al (2010) Reversing chemoresistanceby small molecule inhibition of the translation initiation complexeIF4F. Proc Natl Acad Sci U S A 108:1046–1051
25. Herbert TP, Fahraeus R, Prescott A, Lane DP, Proud CG (2000)Rapid induction of apoptosis mediated by peptides that bind initiationfactor eIF4E. Curr Biol 10:793–796
26. Ko SY, Guo H, Barengo N, Naora H (2009) Inhibition of ovariancancer growth by a tumor-targeting peptide that binds eukaryotictranslation initiation factor 4E. Clin Cancer Res 15:4336–4347
27. Graff JR, Konicek BW, Vincent TM et al (2007) Therapeuticsuppression of translation initiation factor eIF4E expression
Invest New Drugs

reduces tumor growth without toxicity. J Clin Invest 117:2638–2648
28. DeFatta RJ, Nathan CA, De Benedetti A (2000) Antisense RNA toeIF4E suppresses oncogenic properties of a head and neck squamouscell carcinoma cell line. Laryngoscope 110:928–933
29. Soni A, Akcakanat A, Singh G et al (2008) eIF4E knockdowndecreases breast cancer cell growth without activating Akt signaling.Mol Cancer Ther 7:1782–1788
30. Li S, Jia Y, Jacobson B et al (2013) Treatment of breast and lungcancer cells with a N-7 benzyl guanosine monophosphate tryptaminephosphoramidate pronucleotide (4Ei-1) results in chemosensitizationto gemcitabine and induced eIF4E proteasomal degradation. MolPharm 10:523–531
31. Jia Y, Chiu TL, Amin EA, Polunovsky V, Bitterman PB, Wagner CR(2010) Design, synthesis and evaluation of analogs of initiation factor4E (eIF4E) cap-binding antagonist Bn7-GMP. Eur J Med Chem 45:1304–1313
32. Cencic R, Carrier M, Trnkus A, Porco JA Jr, Minden M, Pelletier J(2009) Synergistic effect of inhibiting translation initiation in combi-nation with cytotoxic agents in acute myelogenous leukemia cells.Leuk Res 34:535–541
33. Dong K, Wang R, Wang X et al (2009) Tumor-specific RNAitargeting eIF4E suppresses tumor growth, induces apoptosis andenhances cisplatin cytotoxicity in human breast carcinoma cells.Breast Cancer Res Treat 113:443–456
34. Jacobson BA, Thumma SC, Jay-Dixon J et al (2013) Targetingeukaryotic translation in mesothelioma cells with an eIF4E-specificantisense oligonucleotide. PLoS One 8:e81669
35. Bitterman PB, Polunovsky VA (2012) Attacking a nexus of theoncogenic circuitry by reversing aberrant eIF4F-mediated translation.Mol Cancer Ther 11:1051–1061
36. Bitterman PB, Polunovsky VA (2012) Translational control of cellfate: from integration of environmental signals to breaching antican-cer defense. Cell Cycle 11:1097–1107
Invest New Drugs

























![Mesothelioma lawyers ] mesothelioma attorneys](https://img.pdfslide.net/doc/110x75/5497f892ac795959288b5644/mesothelioma-lawyers-mesothelioma-attorneys.jpg)


