Embed Size (px)
Citation preview

SOME OBSERVATIONS ON THE ECOLOGY
OF PHYTOSEIID MITES
(ACARINA : PHYTOSEIIDAE) IN CALIFORNIA
BY
D.A. CH.~.NT & C.A. FLZSCHNER
I n t r o d u c t i o n
Phytosei id mites were shown in 1906 (PARROTT, et. al) to feed on mites of other families and many references were made subsequently to their predacious habit. These references doubtless gave rise to the belief tha t these predators are valuable in controlling phytophagous mite pests on fruit trees and agricultural crops. This belief was later supported by a number of empirical field studies tha t have recently been criticized. One of us (CHANT, 1958, and in press) showed that, despite the contrary indications from empirical observations, in general Typhlodromus (T.) pyri SCHEUTEN is of little value in the control of Panonychus ulmi (KOCH) in orchards in southeastern England. On the other hand, FLESCHNER (1958) indicated tha t the phytoseiids on citrus and avocado in southern California sometimes significantly control Oligonychus punicae (HI,ST) and Panonychus cirri (MCG.). Some workers considered these findings to be contradictory, and in discussions on the value of predacious mites sometimes quoted them to support one or the other point of view and as a basis for sweeping generalities. The authors recently had the opportuni ty to work together at the Citrus Exper iment Station, Riverside, California, and we were able to resolve partially the apparent ly contrasting v i e w s and to contr ibute to the clarification of the status of phytoseiid mites in general as factors in the control of other mites on fruit trees.
Our joint studies had three aims: to obtain data on the feeding habits of the phytoseiids that are common on fruit trees in southern California; to obtain ecological da ta on the distribution of the Cali- fornian species; and to determine if the difference in effectiveness of the Californian and English species lies in different feeding habits or distributions.
~NTOMOPHAOA T O M ~ V~ N ~ 2~ 2 r T R I M E S T R E I ~ 6 0

132 D . A . C H A N T A N D C. A . F L E S C H N E R
Feeding Habits.
Phytosei ids feed on both the egg and active stages of other mite species. GILLIATT (1935) surmised tha t some species also feed on plants, and this was demonstrated with T. (T.) pyri in southeastern England (CHANT, 1959, and in press).
About 12 species of Phytoseiidae occur on citrus and avocado leaves in southern California. However, only two of these were common in orchards investigated in winter; T. (.4.) hibisci CHANT and T. (.4.) ZsTfto~b~8 (GARMAN et MCOREGOR), and these were the subject of the present investigation. Because no statistically significant differences could be demonstrated between their r espec t ive abilities to feed on the materials tested they are considered together in the following discussion.
The object of this s tudy was to determine if these Californian species are restricted to feeding on other mites or if they too can main- tain themselves on plant foods. Most rearings was done in a constant tempera ture (76oF. 4- 2 ~ and in two types of cages. One was a modified " HUFFAKER " cage, as further modified b y CHANT (1956), and was used when food was not living plant material. The second, described b y M.ATHYS (1954) and modified by CHANT (lot. cir.), was a small plastic cell used to confine mites to living leaves. When not under constant conditions, rearing was done out-of-doors so tha t laboratory results could be interpreted in relation to field conditions.
As t ime did not permit a complete study, the only criteria of food suitabil i ty recorded were longevity and fecundi ty of adul t females and, in a few instances, the effect of various foods on the abili ty of immature mites to develop. Eleven combinations of food were tested and a check was provided in which the mites received no food or water. Pollen was provided by removing the anthers and nectaries of the blossoms and using only these parts. Honeydew from aphids was tested where it occured natural ly on leaves from an infestation on greenhouse seedlings. Mildew of undetermined species occured in some of the cages tha t were moistened with honey and water. No a t t empt was made to ensure tha t these foods were free from associated microor- ganisms and these could in some way have been responsible for the suitabil i ty of certain foods. In some instances the number of specimens of mites tested was small because of morta l i ty in handling or because a particular food was difficult to obtain. The plants used were two or three-year old trees, three to six feet tall, growing in metal containers of soil. The results of this s tudy, shown in Table I, may be discussed as three groups.
First, phytoseiids tha t were s tarved of food and water lived only for a short t ime under the experimental conditions and death was probably due to desiccation rather than to starvation. They did not

ECOLOGY OF PHYTOSEIID MITES IN CALIFORNIA 183
TABLE I
Effects of various foods on the longevity and fecundity of adult females of T. (-4.) hibisci and T. (.4.) limtm@us in an inseetary and in the field.
Food Mean No. eggs No. eggs/ No. longevity laid Mite!Day specimens
{days)
Insectary Without food or water . . . . . . . . . . . . Avocado leaves . . . . . . . . . . . . . . . . . . . Orange leaves . . . . . . . . . . . . . . . . . . . . Water and Honey . . . . . . . . . . . . . . . . Lemon leaves . . . . . . . . . . . . . . . . . . . . .
2 .6 :1: 0.3" 3.9 ~- 0.t, ~.2 : t : 0.~ ~.z, : t : 0.3 ~.8 -I- O.2
Water only . . . . . . . . . . . . . . . . . . . . . . . . 5.7 -4- 0.3 Orange pollen and water . . . . . . . . . . 11.0 -4- 1.9 Honeydew on leaves . . . . . . . . . . . . . . 12.0 4- 0.0 Avocado pollen and water'* . . . . . . 15.2 ~- 1.3 Mildew and water . . . . . . . . . . . . . . . . 15.5 ~ 5.5
Field : Orange leaves . . . . . . . . . . . . . . . . . 9.1 ::1:0.3 Avocado leaves . . . . . . . . . . . . . . . . . . . 10.6 ::[: 2.3
(*) Standard error. - - (**)
0 0 1t 0 0 t 9 0 0 21 0 0 7 0 0 36 0 0 9
18 0.3 6 15 0.2 5
t 55 0.7 16 0 0 2
0 0 6 0 0 5
Not completed because of lack of time.
reproduce and lived but slightly longer on water alone, or when fed only living leaves. Some of the differences in this group were statis- tically significant but their biological significance is doubtful.
Second, orange and avocado pollen with water, aphid honeydew on living orange and lemon leaves, and mildew with water all enabled the mites to live significantly longer than the other foods tested, but oviposition occurred only with the pollen or honeydew. By comparison with four species in England, oviposition was at a slightly higher level as 0.3 eggs per mite per day was the highest rate recorded by CHANT (in press) with mites that were fed animal prey.
Third, mites which were placed in plastic cells on avocado and orange trees in a grove out-of-doors lived more than twice as long as
t h o s e on foliage alone in the laboratory. Further tests could not be done but, if the difference in longevity between laboratory and field with pollen or honeydew as food is of a similar order these mites would be able to live in groves for as long as four weeks in the absence of animal food. Because these foods allowed the mites to lay eggs in the laboratory, they would presumably also be able to reproduce on this food in the field.
The few immature mites tested developed normally on diets of pollen and nectar, and on aphid honeydew, but not on citrus or avocado leaves alone.
These experiments indicated that T. (A.) hibisci and T. (A.) limonicus are not more restricted to a predacious way of life than are the English species studied: they too are facultative predators. The species tested in both areas can live on plant foods alone, with animal food not essential for development or reproduction.

134 D. A. CHANT AND C. A. F L E S C H N E R
Distribution.
O N SURFACES OF LEAVES AND ALONG TWIGS AND BRANCHES.
An important factor tha t prevents T. pyri from effectively control- ling Panonychus ulmi on apple in southeastern England seems to be the discrepancy between the distribution of the predator and tha t of its prey on apple leaves and twigs (CHANT, 1958 and in press). T. pyri, as is often the case with members of the subgenus Typhlodromus, is normally found only along the midribs and larger veins of the lower surfaces of apple leaves and apparent ly it prefers the younger leaves at the tips of shoots and spurs. P. ulmi, on the other hand, prefers the older leaves early in summer, and is fairly uniformly distributed on the lower surfaces and a significant par t of the population is on the upper surfaces of leaves, where T. pyri rarely occurs. Because of the importance tha t we a t t r ibute to this aspect of the ecology of predacious mites, citrus and avocado were investigated from the same viewpoint.
I t was found that in general the phytoseiids on these plants in southern California are distributed differently from T. pyri on apple in England. Both T. (A.) hibisci and T. (A.) limonicus, as is frequently the case with members of the subgenus AmbIyseius, roam over the entire surfaces, both upper and lower, of citrus and avocado leaves. Field observation (C.A.F.) indicated that in summer, on days when tempe- ratures are high, these phytoseiids retreat toward the shaded centres of the trees during the day and move back to the peripheri at night. Thus, there may be a frequent redistribution, and no one type of leaf seems to be constantly preferred as a habi tat . Nearly all areas inhabited by the phytophagous species are also inhabited b y the phytoseiids at one t ime or another.
In greenhouses in England T. pyri changes its habits and it too roams widely over both surfaces of the leaves (Cm~NT, in press). This may explain why in greenhouse and insectary experiments this species seems to control phytophagous species while it does not do so in the field.
V A R I A T I O N B E T W E E N LEAVES.
The between leaf, or interleaf, distributions of T. pyri and P. ulmi on apple in Engalnd early in summer are dissimilar (CHANT, 1958) and suggest tha t the two are independent and, therefore, tha t one does not influence the other. Later in summer the distribution of the two tend to become more similar but, b y tha t time, the pest has often increased to a point where the predator cannot control it regardless of how favourable conditions may be. Distribution can only be an impor tant factor at nearly equal densities of predator and prey or when

ECO LO G Y OF P H Y T O S E I I D MITES IN C A L I F O R N I A 1 8 5
the latter is outnumbered: when there are major differences in density the importance of distr ibution is superceded by the numerical faetor.
The interleaf distribution of the phytoseiid and phytophagous mites on citrus and avocado was investigated during the present study. Records obtained at Riverside (C.A.F. and staff) f rom 1951 to 1957 of the numbers of mites on individual leaves were examined. Specific identifications for the phytoseiids were not available bu t the Riverside slide collection showed only T. (A.) hibisci, T. (A.) limonicus, T. (T.) arboreus CI~ANT, T. (T.) mcgregori CHANT, a n d T. (T.) conspicuus (GARMAN) to occur commonly in the orchards investigated. As no data are available on the relative abundance of these, the populations sam- pled in 1951-1957 may have been composed of any, or all, of them.
Fig. 1 shows the interleaf distributions of predacious and phyto- phagous mites in England and California. The phytoseiids on avocado in California are distr ibuted as is T. pyri on apple in early summer in England. However, whereas in England P. ulmi, the prey species, was distr ibuted differently from the predator, in California the dis- tr ibution of the prey, Oligonychus punicae, closely resembles tha t of the phytoseiids. In England the predator seems unable to signifi- cantly influence the phytophagous mites bu t in California, where the distributions of the two are similar, the converse may be true. The picture on avocado is similar to that on citrus with P. cirri in California.
In addition to investigating the distribution between leaves of some phytophagous and predacious mites in California, an a t t empt was made to determine whether the predators tended to congregate on leaves inhabited by the phytophagous mites, and thus to determine if the predator is a t t rac ted by the prey (or possibly b y the same factors tha t a t t rac t the prey). Da ta from the leaf samples mentioned above were used and they cover several years. Only the samples with both preda- cious and phytophagous mites were considered and, further, only for the months of each year in which the mites were actively reproducing and feeding. Though such composite da ta are not as conclusive as individual counts from a large number of leaves taken at one time no be t te r method could be devised because the sample size of 10 leaves was by itself too small to be significant. Moreover, the number of mites present inflenced the results: for example, in a leaf sample in which every leaf had phytophagous mites, all predators present obvious- ly will be on infested leaves.
The leaves from these samples were placed in several categories, as shown in Table II . The data for each of three orchards were arranged in a 2 • 2 contingency table and tested with the X 2 test. The results, also shown in Table II, are not conclusive bu t indicate tha t sometimes more predacious mites are found on leaves inhabited by phytophagous mites than are to be expected if they are randomly distributed, though the differences be tween the observed and calculated values are not
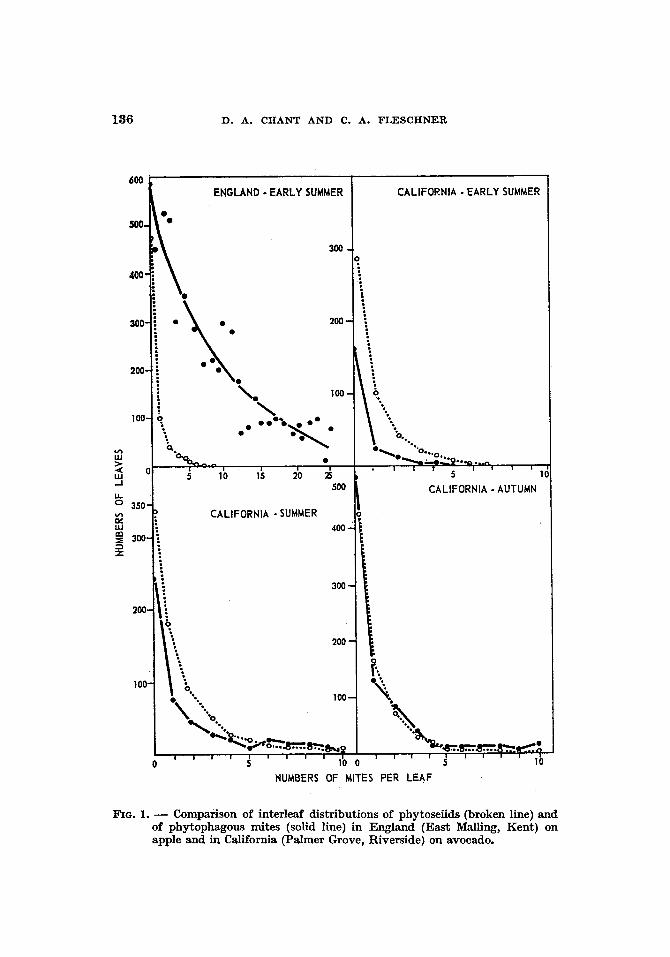
186 D. A. CHANT AND C. A. FLESCHNER
J 600 p t ENGLAND - EARLY SUMMER
�9 ~0- �9
400- ' e l ~ e 300 -,
300- �9 ~ o ~ 200-
\ 2(NI-
ILl ,..1 u_ o 350-
:~ 31111- Z :
. ' i00-
CALIFORNIA - EARLY SUMMER
)
Io \ _ _ 1oo-
100- o �9 ooo,.e,,~ oe �9 ~, �9 �9 O"~,~,,,,~ 0000
O ~ o ""
0~ o ~ _ 5 ,}, 1; 2o "~ " ~ ~ ......... 5 500 CALIFORNIA - AUTUMN
CALIFORNIA - SUMMER 400
I
300 2
200 - -
100-- e~.,
i ; I0
\ " "
: ~ = ~ , ~ ~ . . ~ " , ' . . . . . I 1~ . . . . . " " ; ' ' ; ' ' " ' " ' 'WI [ " ' " ! .5 0 5 10
NUMBERS OF MITES PER LEAF
FzG. 1. - - Comparison of interleaf distributions of phytoseiids (broken line) and of phytophagous mites (solid line) in England (East Malling, Kent) on apple and in California (Palmer Grove, Riverside) on avocado.

ECOLOGY OF PHYTOSEIID MITES IN CALIFORNIA 137
large. At other times this does not seem to be the case; this can per- haps be explained on the grounds tha t the complex of species of pre- dators varied in composition between seasons and years.
TABLE I I The incidence of Typhlodromus spp. on avocado and citrus leaves
infested with phytophagous mites in southern California.
, . $~ ~
o
Orchard Fruit "r. ~, '~ -- ~ o --~ ~ Z a, - . ~ ~ -~.~ "~ o
Palmcr's Avocado July-Sept. 630 1~6 253 107 124 No Oct.-Mar. 810 217 t52 158 283 No Apr.-June ~20 37 27 ~9 t07 No
Bucklew's Avocado Apr.-Oct. 1,072 174 523 182 t93 Yes
Limoncria Orange Apr.-Oct. rt97 236 119 35 107 Yes
The need for fundamental studies along this and other lines of ecology is apparent if we are to understand more fully the nature of predacious mites and to be able to assess their importance to horti- culture and agriculture.
D i s c u s s i o n .
As indicated above, this s tudy was to contribute to our understand- ing of why phytoseiids are more effective in controlling phytophagous mites on citrus and avocado in southern California than on apple in southeastern England. A full explanation was impossible to obtain in the time available but sufficient data were obtained to indicate some possible reasons for this difference.
Major differences in feeding habits apparently do not exist bet- ween t h e species t ha t have been studied and tha t are abundant in each country; none is an obligate predator and all are able to live, develop, and reproduce on foods such as pollen, mildews, and aphid
honeydew (or on the microorganisms these contain), as well as on animal prey. They can also live for some time on leaf juices or moisture alone, though they cannot reproduce or develop on these.
Our studies indicate tha t a t least some of the reasons for the difference probably lie in distribution. The phytoseiids tha t were examined on citrus and avocado in southern California are more or less uniformly distributed over the leaf surfaces, both upper and lower. They are, therefore, a controlling factor of potential importance on all parts of the leaf inhabited by phytophagous mites, and not only in a small fraction of this area as is the case with T. pyri on commercial apple in England.

138 D.A. CHANT AND C. A. :FLESCHNER
Further , in England the interleaf distributions of the predacious and phytophagous mites are dissimilar whereas in California they are alike. Because the phytoseiids in both England and California are similarly distr ibuted it seems that the phytophagous mites on citrus and avocado, 0. punicae and P. cirri, are more suitable than P . ulmi on apple for control by these predators. When the prey distribution on the leaves on a tree is different from tha t of the predator the relation- ships between the two species probably are not close.
The phytoseiids on apple in England tend to inhabit young leaves whereas the phytophagous mites inhabit the older leaves early in the year and may not become generally distr ibuted over all types of leaves until midsummer or later. Conversely, in California the common phytoseiid species seem to be influenced b y temperature and light to move freely between the peripheri and the centres of the trees. Because of this, leaves that are inhabited b y the phytophagous mites are more likely to be visited by the roaming phytoseiids of California than by the more sedentary ones of England.
Winter mortali ty, which may be as high as 90 per cent each year, is impor tant in limiting the effectiveness of T. pyr i in England (CHANT, in press). Few predators are present each spring and before these can increase to an effective level the prey frequent ly increases to the point where it cannot be normally controlled by biotic factors. In California, however, the phytoseiids studied overwinter on the citrus and avocado foliage, which is evergreen, and winter mortal i ty is probably light, though some reduction in numbers occurs in the fall when the humidi ty is very low and in winter when reproduction is curtailed by cooler temperatures. Because the mites overwinter on the leaves, they are able to exert an immediate effect on the prey when the lat ter begins to increase in the spring.
I t seems, therefore, tha t the factors tha t cause the difference between the effectiveness of phytoseiids in England and California are both inherent and environmental. I n h e r e n t factors such as the tendency to be uniformly distributed within the tree or on the surface of a leaf are undoubtedly important bu t it is impossible to conclude a t present whether they are more impor tant than environmental fac- tors such as prey distribution and winter temperatures.
In conclusion, we should like to point out tha t most valid examples of phytoseiids controlling other mites have involved members of the subgenus Amblyseius and it may be tha t these will prove more useful in this regard than species of Typhlodromus s. str.
A C K N O W L E D G M E N T S
The senior author expresses his appreciation to Dr. C. P. C~USEN, formerly Head, Biological Control Department, Citrus Experiment Station, Riverside, California, and his staff for their kindness and interest during this investigation.

ECOLOGY OF PHYTOSEIID MITES IN CALIFORNIA 139
S U M M A R Y
A n a t t e m p t is made to show w h y p h y t o s e i i d mi tes on ci t rus and avocado in sou the rn California seem to be more effective p r eda to r s of p h y t o p h a g o u s mi tes t h a n those on apple in sou theas t e rn Eng land . I t is sugges ted t h a t th is difference is caused b o t h by factors inhe ren t in the p reda to r , such as i ts d is t r ibut ion, a n d b y fac tors in i ts env i ronment , such as t he d i s t r ibu t ion of t h e p rey a n d win te r t empe- ra tures .
S O M M A I R E
Une t e n t a t i v e faite af in de d~terminer les causes pour lesquelles les acariens phy tose i ides s emblen t 6tre des p rdda teurs p lus efficaces des acarieus phy tophages sur les Citrus e t les Avoca t s du sud de la Californie, qu ' i ls le sont sur les pommiers dans le sud de l 'Angle ter re , sugg~re que ce t te difference es t a t t r ibuab le /L la fois
des fae teurs inh~rents aux pr~dateurs , telle leur d is t r ibut ion , e t ~ des fac teurs du milieu, telles la d i s t r ibu t ion des proies e t les t empdra tu r e s hivernales .
R E F E R E N C E S
CHANT, D. A. - - 1956. On the ecology and morphology of some predacious lealaptid mites; with observations on other orchard-inhabiting arachnids. - - Ph. D. thesis (unpublished), University of London, pp. IL.6. - - t958. On the ecology of typhlodromids in southeastern England. - - Proc. 10th Int. (:ongr. Ent., 4, 6t,9-658. - - t959. Observations sur la Famille des Phytoseiidue. - - Acarologia, 1, 11-22. - - In press. Phytoseiid mites (Acarina : Phytoseiidae). Part I. Bionomics of seven species in southeastern England. Part. II. A taxonomic review of the family Phytoseiidae, wittl descriptions o~ 37 new species. - - Canad. Ent. Suppl.
FL~SCnNEn, C.A. - - 1958. Natural enemies of Tetranychid mites on citrus and avocado in southern California. - - Proc. lOth Int. Congr. Ent., 4, 627-631.
GILLXATT, F. C. - - 1935. Some predators of the European red mite, Paratetranychus pilosus c. & F., in Nova Scotia. - - Canad. J. Res., D, 13, 19-38.
MATnYS, G. - - 195t~.. Contribution 6thologique ~ la r6solution du complexe Bryobia praetiosa KOCH (Acar., Tetranych.). - - Mitt. Schweiz. Entomol. Ges., 27, t37-It~6.
PAnnOTT, P . J . , H . E . HODCKISS and W . J . SCUOENE. - - 1906. The Eriophyidae Part I. The apple and pear mites. ~ Ne~v York Agr. Expt. Sta. Bull., 283, 302- 303.
(Entomology Research Inst i tute /or Biological Control, Research Branch, Canada Department o] Agriculture, Belleville, Ontario ;
Department o/ Biological Control, Citrus Experi- ment Station, Riverside, Cali /ornia.)





























