Embed Size (px)
Citation preview

297Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Važna svojstva Jadranskoga mora su: a) položaj rijeke Po koja je najveći
izvor slatke vode i hranjivih tvari u Sredozemnom moru; b) zimska
bura koja sudjeluje u stvaranju Sjevernojadranske guste vode i pokreće
cirkulaciju u istočnom dijelu Sredozemnog mora; c) Istočnojadranska
struja koja uz hrvatsku obalu donosi oligotrofnu vodu iz istočnog dijela
Sredozemnog mora; d) Istarska fronta; e) uzdužni i poprečni gradijent
raspodjele fizikalno-kemijskih i bioloških svojstava; f) cvjetanje
fitoplanktona s pojavom makroagregata organske tvari u sjevernom
Jadranu; g) mali raspon plime i oseke; h) krška hidrogeologija i postojanje
visokostratificiranih estuarija uz hrvatsku obalu; i) veći udio manjih
veličinskih frakcija fitoplanktona u ukupnoj primarnoj biomasi, kao i veći
udio biomase bakterija u ukupnoj mikrobnoj biomasi u južnom, nego u
sjevernom Jadranu; j) izolirana staništa koja su nastala potapanjem krša.
Najvažnije obilježje istočnog dijela Jadrana (po čemu se razlikuje od
zapadnog dijela) jest njegova oligotrofija, što je komparativna prednost
koju Hrvatska treba koristiti za razvoj prikladnih privrednih aktivnosti.
Ključne riječi: jadransko more, oceanografska svojstva, ekološka svojstva, pelagijal, gospodarenje, zaštita
specifična OceanOlOšKa svOjstva hrvatsKOg dijela jadrana
prof. dr. sc. damir viličić, dipl. ing. biol.Prirodoslovno-matematički fakultet
Sveučilišta u ZagrebuBiološki odsjek
Rooseveltov trg 6, [email protected]
pregledni članak Review paper UDk 551.46(497.5 Jadran)primljeno (Received): 23.11.2013.; prihvaćeno (Accepted): 17.9.2014.
1.UVODOvaj pregledni rad prikazuje oceanografska svojstva i
ekološka svojstva pelagijala - zajednice organizama koji naseljavaju vodeni stupac u otvorenim vodama Jadrana i u obalnim područjima, od kojih su među najzanimljivijima frontalno područje između Istre i Italije, Kvarner, estuariji malih krških rijeka, Malostonski zaljev, sjeverozapadni rub Južnojadranske kotline i objekti potopljenog krša.
2. OCEANOgRAfIJA I OSNOvNA SvOJSTvA PELAgIjALA OTVORENIh VODA jADRANA
Jadransko je more najsjeverniji izdanak Sredozemnog mora. Čini samo 4,6% od ukupne njegove površine. Odvojeno je od Jonskog mora Otrantskim vratima (70 km širokim) i Otrantskim pragom (789 m dubokim). Razvedenost hrvatske obale je osam puta veća nego talijanske obale. Veći je dio Jadrana (73,9%) plići od 200

298 Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Slika 1: Jadransko more – batimetrijska karta
Slika 2: Važna oceanografska svojstva Jadranskoga mora (Viličić 1994).
m (slika 1). Najveća je dubina izmjerena u Južnojadranskoj kotlini (1 228 m). Granicu između sjevernog i srednjeg Jadrana čini Jabučka (Srednjojadranska) kotlina. Južni i srednji Jadran graniče na Palagruškom pragu.
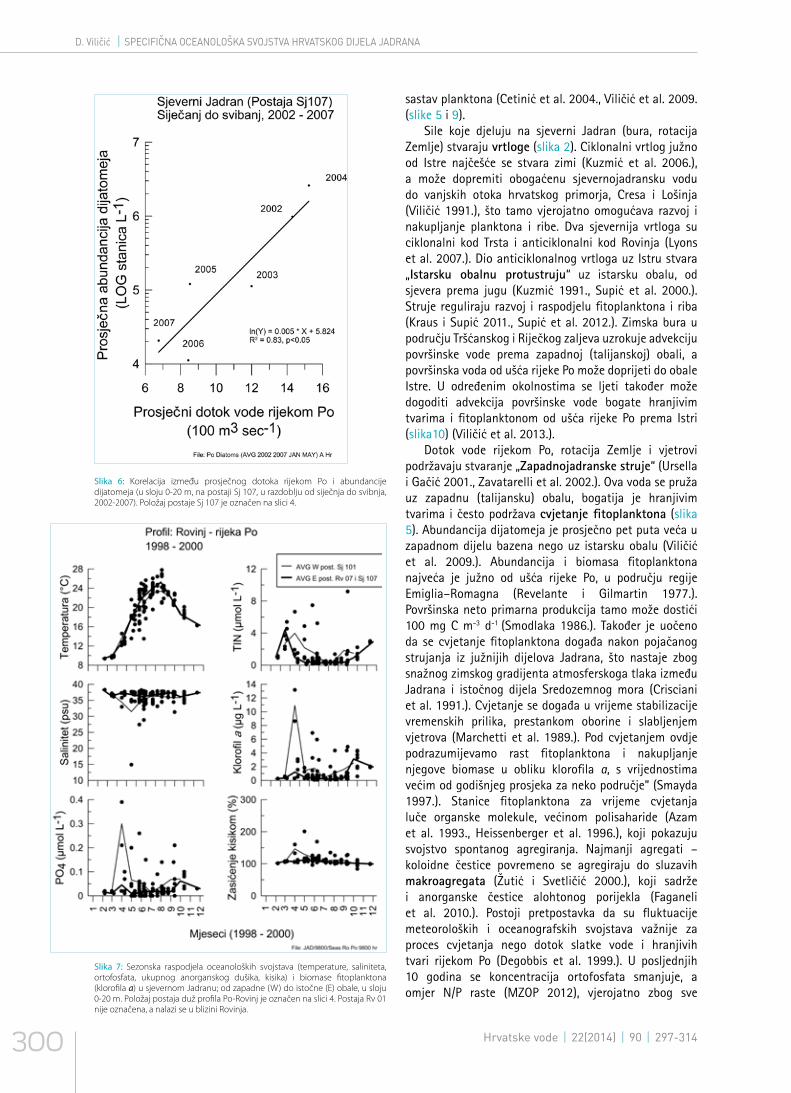
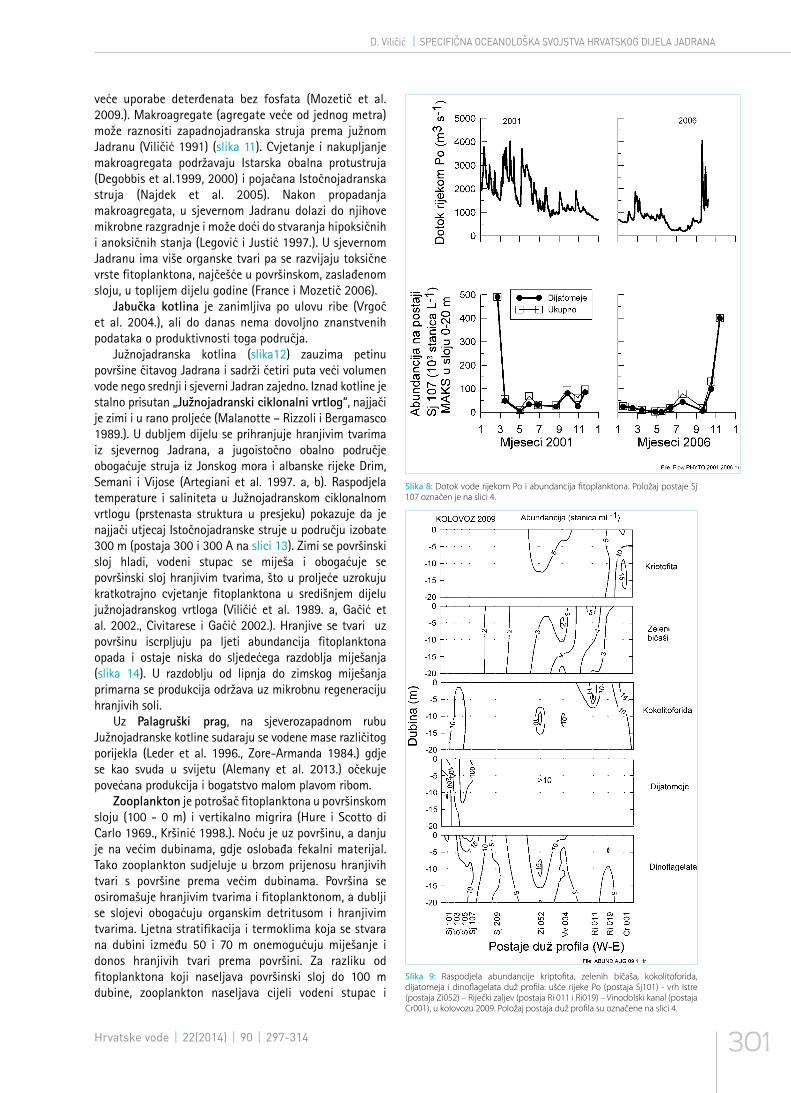
Najvažniji oceanografski čimbenici koji djeluju na raspodjelu živog svijeta u Jadranu su: salinitet i koncentracija hranjivih tvari, intenzitet svjetlosti, morske struje (horizontalne, vertikalne i kružne), frontalni sustavi, temperatura, odnosi među populacijama i dr. (slika 2). Istočnojadransko slivno područje je manje od zapadnojadranskog slivnog područja (slika 3). Važan su izvor hranjivih tvari rijeke u sjeverozapadnom dijelu Jadrana, od kojih je najveća rijeka Po (Chiaudani et al. 1980., Degobbis i Gilmartin 1990.) (slika 4). Od zapada prema istoku se stvara gradijent koncentracije hranjivih tvari, što prati raspodjela fitoplanktona (Smodlaka 1986., Viličić 1991., Polimene et al. 2006.) (slika 5), otopljene organske tvari (Gašparović i Ćosović 2001.), te zooplanktona (Hure i Kršinić 1998.). Razvoj fitoplanktona u sjevernom Jadranu ovisi o režimu oborine i topljenju snijega u Alpama, te o dotoku hranjivih tvari rijekom Po (slike 6 i 7). Proljetni je maksimalni dotok vode rijekom Po rezultat topljenja snijega u Alpama, a jesenski maksimum je posljedica oborinskog maksimuma (slika 8). Kada je u Alpama zima siromašna snijegom, manji je proljetni dotok hranjivih soli i proljetni razvoj fitoplanktona (Viličić et al. 2007., 2009.). Koncentracija otopljenog kisika u vodenom stupcu je najčešće visoka (slika7).
Struje ovise o vjetrovima, rotaciji Zemlje, morfologiji dna i izmjeni vode kroz Otrantska vrata (Orlić et al. 1992., 1994.; Penzar et al. 2001.). Dominantni vjetrovi su bura i jugo (Poulain i Raicich 2001.). Bura je hladan i suh vjetar koji puše iz sjeveroistočnog kvadranta; miješa vodeni stupac, povećava isparavanje, gustoću mora i gubitak topline (Dorman et al. 2006.). Najjača je od prosinca do ožujka, u Tršćanskom zaljevu, Bakarskom zaljevu, oko Senja i Šibenika, u zaljevu Vrulja kod Makarske, te oko Dubrovnika. Jugo ili široko je jugoistočni vjetar, najjači je u kanalima između obale i otoka, otvorenima prema jugoistoku. Veća je učestalost juga u hladnijem dijelu godine, od studenoga do travnja, a učestalost se povećava od sjevernog prema južnom Jadranu.
Kretanje pojedinih tipova vodenih masa može se prepoznati po njihovim termohalinim svojstvima. U Jadranu se prepoznaje pet glavnih tipova vode (Cushman-Roisin et al. 2001.), koji se razlikuju prema temperaturi, salinitetu i gustoći: 1. „Jadranska površinska voda“ se zimi znatnije hladi, ali nije prepoznatljiva po karakterističnim vrijednostima temperature i saliniteta. 2. „Jonska površinska voda“ je zimi toplija i slanija voda. 3. „Levantinska intermedijarna voda“ definira se salinitetom većim od 38.75 i T = 14 ˚C, a u južni se Jadran pruža iz Jonskog mora u sloju između 40 i nekoliko stotina metara (Vilibić i Orlić 2002.). Brzina strujanja u spomenutom se sloju mijenja od godine do godine, a ovisi o gradijentu atmosferskoga tlaka i gradijentu gustoće vode u istočnom Sredozemlju te o atmosferskim i oceanskim oscilacijama; npr. sjevernoatlantskoj oscilaciji (Hurrell 1995.) ili jonskoj oscilaciji (Civitarese et al. 2010.). 4. „Sjevernojadranska gusta voda“ ima temperaturu 12.3 – 12.5 ̊ C i salinitet 38.3 – 38.4 (Manca i Giorgetti 1999.), a

299Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Slika 3: Površina slivova jadranskih rijeka.
Slika 4: Karta sjevernog Jadrana s postajama koje su analizirane u rezultatima.
Slika 5: Horizontalna raspodjela biomase mikrofitoplanktona (biovolumen, 107 µm3 L-1) uz površinu Jadranskoga mora.
nastaje u čitavom sjevernom Jadranu za vrijeme zimskog djelovanja bure (Vilibić et al. 2004., Vilibić i Supić 2005., Mihanović et al. 2013.). Ta voda najbrže tone u području konvergentne istarske fronte (Peters et al. 2007.), širi se u pridnenom sloju i popunjava Jabučku i Južnojadransku kotlinu. 5. U dubokom dijelu Južnojadranske kotline ta se voda transformira u „južnojadransku duboku vodu“ s gustoćom većom od 29.18 kg/m3, temperaturom < 13 ˚C i salinitetom@ 38.60 (Manca i Giorgetti 1999.).
Bitne komponente cirkulacije Jadrana jesu „Istočnojadranska struja“, „Zapadnojadranska struja“ i „Zapadnojadranska pridnena struja“(Zore-Armanda 1969., Orlić et al. 1992., Gačić et al. 1999., Poulain i Raicich 2001., Civitarese i Gačić 2002.). Površinske ulazne struje su intenzivnije duž istočne obale zimi, a površinske izlazne struje su intenzivnije duž zapadne obale ljeti. Postoje i transverzalne struje koje su usmjerene pretežno od istočne prema zapadnoj obali (Borzelli et al. 1999.). Istočnojadranska struja prenosi topliju i slaniju vodu uz istočnu obalu Jadrana iz istočnog dijela Sredozemnog mora i Jonskog mora (Zore-Armanda 1969.). U razdoblju pojačanih zimskih vjetrova u Egejskom moru slabi dotok Levantinske intermedijarne vode u Jadran i pada salinitet u južnom Jadranu (Samuel et al. 1999.). Struja je najjača u razdoblju kada je oscilacija u Jonskom moru u fazi ciklonalnog vrtloga (godine «ingresije») i tada je intenzivnija izmjena vode u sjevernom. To je dekadalna oscilacija, kao i druge oscilacije na široj prostornoj skali, (npr. sjevernoatlantska i jonska oscilacija) djeluju na biološku produkciju u Jadranskom moru ( Pucher–Petković i Zore – Armanda 1973., Civitarese i Gačić 2001., Vilibić et al. 2012.), kao i pasivni transport planktona u južni Jadran (Batistić et al. 2012.). Uočeno je da se cvjetanje fitopanktona i razvoj zooplanktona u sjevernom Jadranu događa u vrijeme pojačanog prijenosa vode iz južnog Jadrana (Crisciani et al. 1991., Najdek et al. 2005.). Važan je razvoj herbivora u vrijeme cvjetanja fitoplanktona pa se pretpostavlja da su uzroci cvjetanja fitoplanktona najprije meteorološki, a u kasnijoj fazi i biološki (Kršinić et al. 2007.). Kada jonska oscilacija poprimi suprotni predznak, uz prevladavanje anticiklonalnog vrtloga Jadransko more dolazi pod utjecaj zapadnog dijela Sredozemnog mora i manjeg saliniteta (Civitarese i Gačić 2002., Civitarese et al. 2010.). Klimatske promjene odrazile su se u promjeni saliniteta, količini hranjivih tvari, sastavu fitoplanktona (Marić et al. 2012.), trofičkog stupnja i cirkulacije (Đakovac et al. 2012.) u sjeveroistočnom dijelu Jadrana tijekom posljednje četiri dekade.
Glavnina ulazne Istočnojadranske struje zakreće od vrha istarskog poluotoka prema zapadu. Stvara se frontalna zona na mjestu razgraničenja s vodom sjevernog Jadrana. Fronta je konvektivna i može biti široka samo stotinjak metara (Jeffries i Lee 2007.). U literaturi se spominje kao „Istarska fronta“ (Lee et al. 2005). Istarska fronta je područje gdje se površinska, zgušćena voda prenosi u dubinu. Sa svake strane Istarske fronte nalazimo različita termohalina svojstva mora i
300 Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Slika 6: Korelacija između prosječnog dotoka rijekom Po i abundancije dijatomeja (u sloju 0-20 m, na postaji Sj 107, u razdoblju od siječnja do svibnja, 2002-2007). Položaj postaje Sj 107 je označen na slici 4.
Slika 7: Sezonska raspodjela oceanoloških svojstava (temperature, saliniteta, ortofosfata, ukupnog anorganskog dušika, kisika) i biomase fitoplanktona (klorofila a) u sjevernom Jadranu; od zapadne (W) do istočne (E) obale, u sloju 0-20 m. Položaj postaja duž profila Po-Rovinj je označen na slici 4. Postaja Rv 01 nije označena, a nalazi se u blizini Rovinja.
sastav planktona (Cetinić et al. 2004., Viličić et al. 2009. (slike 5 i 9).
Sile koje djeluju na sjeverni Jadran (bura, rotacija Zemlje) stvaraju vrtloge (slika 2). Ciklonalni vrtlog južno od Istre najčešće se stvara zimi (Kuzmić et al. 2006.), a može dopremiti obogaćenu sjevernojadransku vodu do vanjskih otoka hrvatskog primorja, Cresa i Lošinja (Viličić 1991.), što tamo vjerojatno omogućava razvoj i nakupljanje planktona i ribe. Dva sjevernija vrtloga su ciklonalni kod Trsta i anticiklonalni kod Rovinja (Lyons et al. 2007.). Dio anticiklonalnog vrtloga uz Istru stvara „Istarsku obalnu protustruju“ uz istarsku obalu, od sjevera prema jugu (Kuzmić 1991., Supić et al. 2000.). Struje reguliraju razvoj i raspodjelu fitoplanktona i riba (Kraus i Supić 2011., Supić et al. 2012.). Zimska bura u području Tršćanskog i Riječkog zaljeva uzrokuje advekciju površinske vode prema zapadnoj (talijanskoj) obali, a površinska voda od ušća rijeke Po može doprijeti do obale Istre. U određenim okolnostima se ljeti također može dogoditi advekcija površinske vode bogate hranjivim tvarima i fitoplanktonom od ušća rijeke Po prema Istri (slika10) (Viličić et al. 2013.).
Dotok vode rijekom Po, rotacija Zemlje i vjetrovi podržavaju stvaranje „Zapadnojadranske struje“ (Ursella i Gačić 2001., Zavatarelli et al. 2002.). Ova voda se pruža uz zapadnu (talijansku) obalu, bogatija je hranjivim tvarima i često podržava cvjetanje fitoplanktona (slika 5). Abundancija dijatomeja je prosječno pet puta veća u zapadnom dijelu bazena nego uz istarsku obalu (Viličić et al. 2009.). Abundancija i biomasa fitoplanktona najveća je južno od ušća rijeke Po, u području regije Emiglia–Romagna (Revelante i Gilmartin 1977.). Površinska neto primarna produkcija tamo može dostići 100 mg C m-3 d-1 (Smodlaka 1986.). Također je uočeno da se cvjetanje fitoplanktona događa nakon pojačanog strujanja iz južnijih dijelova Jadrana, što nastaje zbog snažnog zimskog gradijenta atmosferskoga tlaka između Jadrana i istočnog dijela Sredozemnog mora (Crisciani et al. 1991.). Cvjetanje se događa u vrijeme stabilizacije vremenskih prilika, prestankom oborine i slabljenjem vjetrova (Marchetti et al. 1989.). Pod cvjetanjem ovdje podrazumijevamo rast fitoplanktona i nakupljanje njegove biomase u obliku klorofila a, s vrijednostima većim od godišnjeg prosjeka za neko područje” (Smayda 1997.). Stanice fitoplanktona za vrijeme cvjetanja luče organske molekule, većinom polisaharide (Azam et al. 1993., Heissenberger et al. 1996.), koji pokazuju svojstvo spontanog agregiranja. Najmanji agregati – koloidne čestice povremeno se agregiraju do sluzavih makroagregata (Žutić i Svetličić 2000.), koji sadrže i anorganske čestice alohtonog porijekla (Faganeli et al. 2010.). Postoji pretpostavka da su fluktuacije meteoroloških i oceanografskih svojstava važnije za proces cvjetanja nego dotok slatke vode i hranjivih tvari rijekom Po (Degobbis et al. 1999.). U posljednjih 10 godina se koncentracija ortofosfata smanjuje, a omjer N/P raste (MZOP 2012), vjerojatno zbog sve
301Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Slika 8: Dotok vode rijekom Po i abundancija fitoplanktona. Položaj postaje Sj 107 označen je na slici 4.
Slika 9: Raspodjela abundancije kriptofita, zelenih bičaša, kokolitoforida, dijatomeja i dinoflagelata duž profila: ušće rijeke Po (postaja Sj101) - vrh Istre (postaja Zi052) – Riječki zaljev (postaja Ri 011 i Ri019) – Vinodolski kanal (postaja Cr001), u kolovozu 2009. Položaj postaja duž profila su označene na slici 4.
veće uporabe deterđenata bez fosfata (Mozetič et al. 2009.). Makroagregate (agregate veće od jednog metra) može raznositi zapadnojadranska struja prema južnom Jadranu (Viličić 1991) (slika 11). Cvjetanje i nakupljanje makroagregata podržavaju Istarska obalna protustruja (Degobbis et al.1999, 2000) i pojačana Istočnojadranska struja (Najdek et al. 2005). Nakon propadanja makroagregata, u sjevernom Jadranu dolazi do njihove mikrobne razgradnje i može doći do stvaranja hipoksičnih i anoksičnih stanja (Legović i Justić 1997.). U sjevernom Jadranu ima više organske tvari pa se razvijaju toksične vrste fitoplanktona, najčešće u površinskom, zaslađenom sloju, u toplijem dijelu godine (France i Mozetič 2006).
Jabučka kotlina je zanimljiva po ulovu ribe (Vrgoč et al. 2004.), ali do danas nema dovoljno znanstvenih podataka o produktivnosti toga područja.
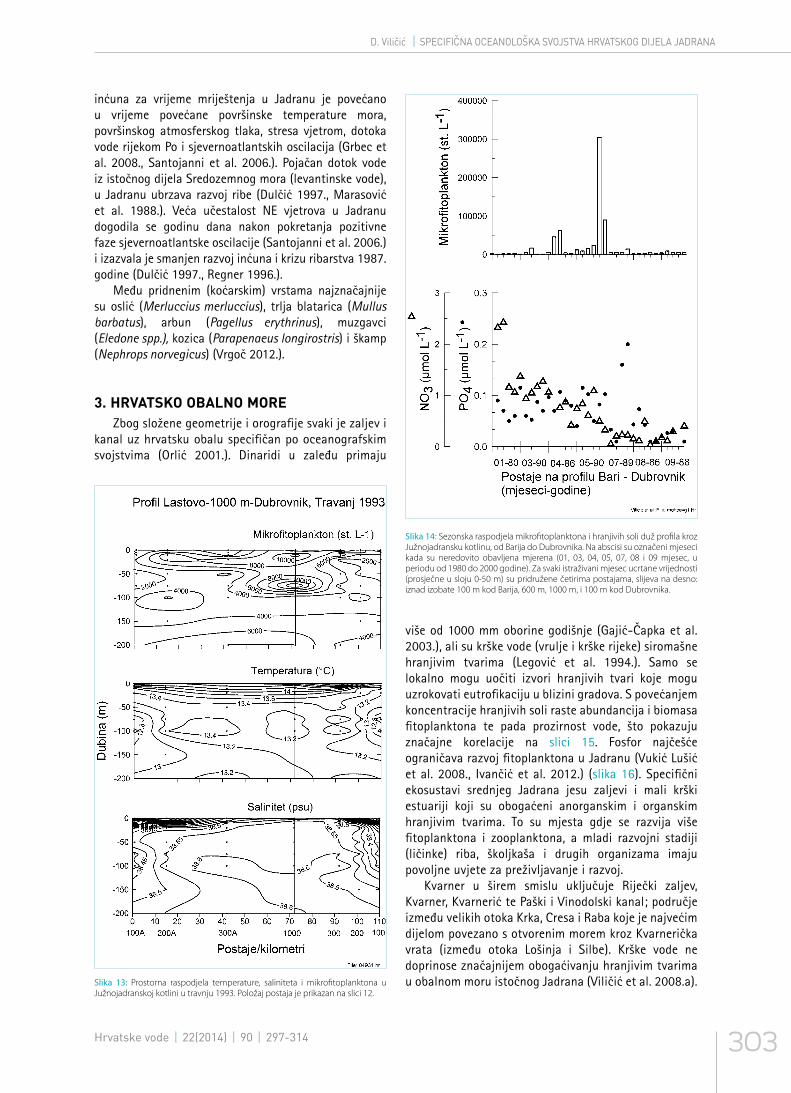
Južnojadranska kotlina (slika12) zauzima petinu površine čitavog Jadrana i sadrži četiri puta veći volumen vode nego srednji i sjeverni Jadran zajedno. Iznad kotline je stalno prisutan „Južnojadranski ciklonalni vrtlog“, najjači je zimi i u rano proljeće (Malanotte – Rizzoli i Bergamasco 1989.). U dubljem dijelu se prihranjuje hranjivim tvarima iz sjevernog Jadrana, a jugoistočno obalno područje obogaćuje struja iz Jonskog mora i albanske rijeke Drim, Semani i Vijose (Artegiani et al. 1997. a, b). Raspodjela temperature i saliniteta u Južnojadranskom ciklonalnom vrtlogu (prstenasta struktura u presjeku) pokazuje da je najjači utjecaj Istočnojadranske struje u području izobate 300 m (postaja 300 i 300 A na slici 13). Zimi se površinski sloj hladi, vodeni stupac se miješa i obogaćuje se površinski sloj hranjivim tvarima, što u proljeće uzrokuju kratkotrajno cvjetanje fitoplanktona u središnjem dijelu južnojadranskog vrtloga (Viličić et al. 1989. a, Gačić et al. 2002., Civitarese i Gačić 2002.). Hranjive se tvari uz površinu iscrpljuju pa ljeti abundancija fitoplanktona opada i ostaje niska do sljedećega razdoblja miješanja (slika 14). U razdoblju od lipnja do zimskog miješanja primarna se produkcija održava uz mikrobnu regeneraciju hranjivih soli.
Uz Palagruški prag, na sjeverozapadnom rubu Južnojadranske kotline sudaraju se vodene mase različitog porijekla (Leder et al. 1996., Zore-Armanda 1984.) gdje se kao svuda u svijetu (Alemany et al. 2013.) očekuje povećana produkcija i bogatstvo malom plavom ribom.
Zooplankton je potrošač fitoplanktona u površinskom sloju (100 - 0 m) i vertikalno migrira (Hure i Scotto di Carlo 1969., Kršinić 1998.). Noću je uz površinu, a danju je na većim dubinama, gdje oslobađa fekalni materijal. Tako zooplankton sudjeluje u brzom prijenosu hranjivih tvari s površine prema većim dubinama. Površina se osiromašuje hranjivim tvarima i fitoplanktonom, a dublji se slojevi obogaćuju organskim detritusom i hranjivim tvarima. Ljetna stratifikacija i termoklima koja se stvara na dubini između 50 i 70 m onemogućuju miješanje i donos hranjivih tvari prema površini. Za razliku od fitoplanktona koji naseljava površinski sloj do 100 m dubine, zooplankton naseljava cijeli vodeni stupac i

302 Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Slika 10: Advekcija površinske vode nižeg saliniteta s povećanom biomasom fitoplanktona od ušća rijeke Po prema istarskoj obali, nakon kratkotrajnog porasta dotoka vode rijekom Po, sredinom kolovoza 2010 (sliku pripremio dr. Milivoj Kuzmić, Institut Ruđer Bošković, Zagreb). Koncentracije klorofila a su u rasponu od 0,01 µg m-3 u Kvarneru do 5 µg m-3 uz ušće rijeke Po.
Slika 11: Makroagregati organske tvari .
Slika 12: Batimetrija Južnojadranske kotline i istraživački profil Lastovo (Glavat)– Južnojadranska kotlina (izobata 1000 m) – Dubrovnik.
razlikuju se površinska i potpovršinska zajednica (do 100 m), mezopelagička (100 do 600 m) i dubokomorska zajednica (ispod 600 m dubine). Rakovi i želatinozni plankton u dubokomorskoj zajednici tijekom noći migriraju do mezopelagičnog sloja (Benović 1973., Batistić et al. 2004., Lučić et al. 2005.).
Prema svim pokazateljima istočni dio južnog, srednjeg i sjevernog Jadrana je oligotrofno područje (Viličić 1989.; Viličić et al. 2008.a, 2009.; Precali et al. 2003.). U razdoblju od 1970-ih do 1990-ih godina postoje pokazatelji porasta eutrofikacije u srednjem (Šolić et al. 1997.) i u južnom Jadranu (Marasović et al. 1995.). U
sjevernom Jadranu se od 1970 godine bilježi opadanje donosa hranjivih tvari rijekom Po, jer se u Italiji prestaju rabiti deterđenti s fosforom (Mozetičet al. 2009.).
Uz istočnu obalu Jadrana krupniji se fitoplankton uglavnom sastoji od dijatomeja (Viličić et al. 2009., Jasprica et al. 2001.). Udio manjih veličinskih frakcija fitoplanktona u ukupnoj primarnoj biomasi, kao i udio biomase bakterija u ukupnoj mikrobnoj biomasi je veći u južnom nego u sjevernom Jadranu (Polimene et al. 2006.).
Dekadalno povećanje utjecaja levantinske vode povećava produkciju fitoplanktona i ribe u Jadranu (Pucher Petković i Zore-Armanda 1973., Civitarese i Gačić 2002.).
Najvažniji riblji resursi Jadranskoga mora jest sitna plava riba; inćun (lat. Engraulis encrasicolus L.) i srdela (lat. Sardina pilchardus Walbaum). Razvoj ličinki srdele i inćuna ovisi o temperaturi, donosu hranjivih tvari i razvoju planktona (Regner 1996.). Klimatske oscilacije mijenjaju režim vjetrova, oborine i temperature, dotok slatkih voda i hranjivih tvari, cirkulaciju mora, a time i trofičke odnose. U Jadranskom moru inćun se mrijesti u razdoblju kada je temperatura mora između 12 do 27 °C, a salinitet između 9 i 39 (Regner 1996.). Jaja inćuna nalaze se u planktonu u periodu od travnja do listopada, ali se mogu pojaviti ranije ako je temperatura povišena u siječnju i veljači. Produkcija jaja inćuna i količina ličinki prati produkciju zooplanktona s vremenskim pomakom od oko dva mjeseca. Atmosferski procesi na široj prostornoj skali značajno utječu na razvoj ribe i glavonožaca. Nakupljanje
303Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Slika 13: Prostorna raspodjela temperature, saliniteta i mikrofitoplanktona u Južnojadranskoj kotlini u travnju 1993. Položaj postaja je prikazan na slici 12.
Slika 14: Sezonska raspodjela mikrofitoplanktona i hranjivih soli duž profila kroz Južnojadransku kotlinu, od Barija do Dubrovnika. Na abscisi su označeni mjeseci kada su neredovito obavljena mjerena (01, 03, 04, 05, 07, 08 i 09 mjesec, u periodu od 1980 do 2000 godine). Za svaki istraživani mjesec ucrtane vrijednosti (prosječne u sloju 0-50 m) su pridružene četirima postajama, slijeva na desno: iznad izobate 100 m kod Barija, 600 m, 1000 m, i 100 m kod Dubrovnika.
inćuna za vrijeme mriještenja u Jadranu je povećano u vrijeme povećane površinske temperature mora, površinskog atmosferskog tlaka, stresa vjetrom, dotoka vode rijekom Po i sjevernoatlantskih oscilacija (Grbec et al. 2008., Santojanni et al. 2006.). Pojačan dotok vode iz istočnog dijela Sredozemnog mora (levantinske vode), u Jadranu ubrzava razvoj ribe (Dulčić 1997., Marasović et al. 1988.). Veća učestalost NE vjetrova u Jadranu dogodila se godinu dana nakon pokretanja pozitivne faze sjevernoatlantske oscilacije (Santojanni et al. 2006.) i izazvala je smanjen razvoj inćuna i krizu ribarstva 1987. godine (Dulčić 1997., Regner 1996.).
Među pridnenim (koćarskim) vrstama najznačajnije su oslić (Merluccius merluccius), trlja blatarica (Mullus barbatus), arbun (Pagellus erythrinus), muzgavci (Eledone spp.), kozica (Parapenaeus longirostris) i škamp (Nephrops norvegicus) (Vrgoč 2012.).
3. hRVATSKO ObALNO MOREZbog složene geometrije i orografije svaki je zaljev i
kanal uz hrvatsku obalu specifičan po oceanografskim svojstvima (Orlić 2001.). Dinaridi u zaleđu primaju
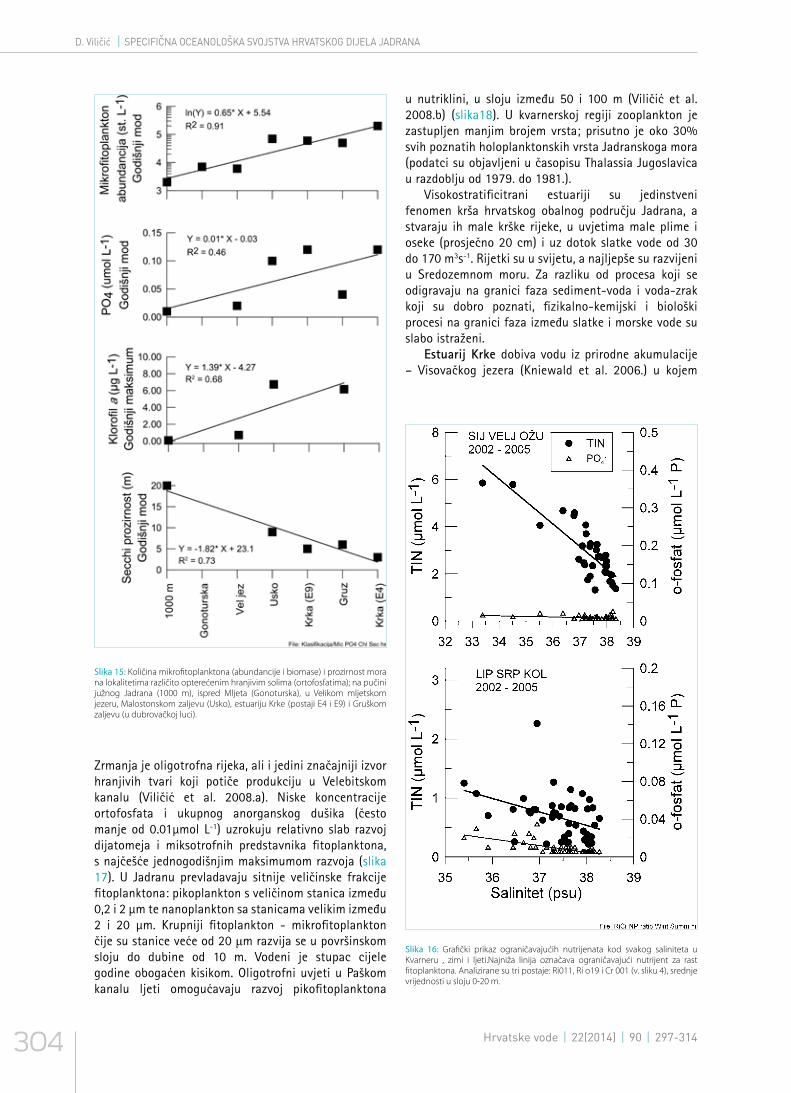
više od 1000 mm oborine godišnje (Gajić-Čapka et al. 2003.), ali su krške vode (vrulje i krške rijeke) siromašne hranjivim tvarima (Legović et al. 1994.). Samo se lokalno mogu uočiti izvori hranjivih tvari koje mogu uzrokovati eutrofikaciju u blizini gradova. S povećanjem koncentracije hranjivih soli raste abundancija i biomasa fitoplanktona te pada prozirnost vode, što pokazuju značajne korelacije na slici 15. Fosfor najčešće ograničava razvoj fitoplanktona u Jadranu (Vukić Lušić et al. 2008., Ivančić et al. 2012.) (slika 16). Specifični ekosustavi srednjeg Jadrana jesu zaljevi i mali krški estuariji koji su obogaćeni anorganskim i organskim hranjivim tvarima. To su mjesta gdje se razvija više fitoplanktona i zooplanktona, a mladi razvojni stadiji (ličinke) riba, školjkaša i drugih organizama imaju povoljne uvjete za preživljavanje i razvoj.
Kvarner u širem smislu uključuje Riječki zaljev, Kvarner, Kvarnerić te Paški i Vinodolski kanal; područje između velikih otoka Krka, Cresa i Raba koje je najvećim dijelom povezano s otvorenim morem kroz Kvarnerička vrata (između otoka Lošinja i Silbe). Krške vode ne doprinose značajnijem obogaćivanju hranjivim tvarima u obalnom moru istočnog Jadrana (Viličić et al. 2008.a).
304 Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Slika 15: Količina mikrofitoplanktona (abundancije i biomase) i prozirnost mora na lokalitetima različito opterećenim hranjivim solima (ortofosfatima); na pučini južnog Jadrana (1000 m), ispred Mljeta (Gonoturska), u Velikom mljetskom jezeru, Malostonskom zaljevu (Usko), estuariju Krke (postaji E4 i E9) i Gruškom zaljevu (u dubrovačkoj luci).
Slika 16: Grafički prikaz ograničavajućih nutrijenata kod svakog saliniteta u Kvarneru , zimi i ljeti.Najniža linija označava ograničavajući nutrijent za rast fitoplanktona. Analizirane su tri postaje: Ri011, Ri o19 i Cr 001 (v. sliku 4), srednje vrijednosti u sloju 0-20 m.
Zrmanja je oligotrofna rijeka, ali i jedini značajniji izvor hranjivih tvari koji potiče produkciju u Velebitskom kanalu (Viličić et al. 2008.a). Niske koncentracije ortofosfata i ukupnog anorganskog dušika (često manje od 0.01µmol L-1) uzrokuju relativno slab razvoj dijatomeja i miksotrofnih predstavnika fitoplanktona, s najčešće jednogodišnjim maksimumom razvoja (slika 17). U Jadranu prevladavaju sitnije veličinske frakcije fitoplanktona: pikoplankton s veličinom stanica između 0,2 i 2 µm te nanoplankton sa stanicama velikim između 2 i 20 µm. Krupniji fitoplankton - mikrofitoplankton čije su stanice veće od 20 μm razvija se u površinskom sloju do dubine od 10 m. Vodeni je stupac cijele godine obogaćen kisikom. Oligotrofni uvjeti u Paškom kanalu ljeti omogućavaju razvoj pikofitoplanktona
u nutriklini, u sloju između 50 i 100 m (Viličić et al. 2008.b) (slika18). U kvarnerskoj regiji zooplankton je zastupljen manjim brojem vrsta; prisutno je oko 30% svih poznatih holoplanktonskih vrsta Jadranskoga mora (podatci su objavljeni u časopisu Thalassia Jugoslavica u razdoblju od 1979. do 1981.).
Visokostratificitrani estuariji su jedinstveni fenomen krša hrvatskog obalnog području Jadrana, a stvaraju ih male krške rijeke, u uvjetima male plime i oseke (prosječno 20 cm) i uz dotok slatke vode od 30 do 170 m3s-1. Rijetki su u svijetu, a najljepše su razvijeni u Sredozemnom moru. Za razliku od procesa koji se odigravaju na granici faza sediment-voda i voda-zrak koji su dobro poznati, fizikalno-kemijski i biološki procesi na granici faza između slatke i morske vode su slabo istraženi.
Estuarij Krke dobiva vodu iz prirodne akumulacije – Visovačkog jezera (Kniewald et al. 2006.) u kojem
305Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
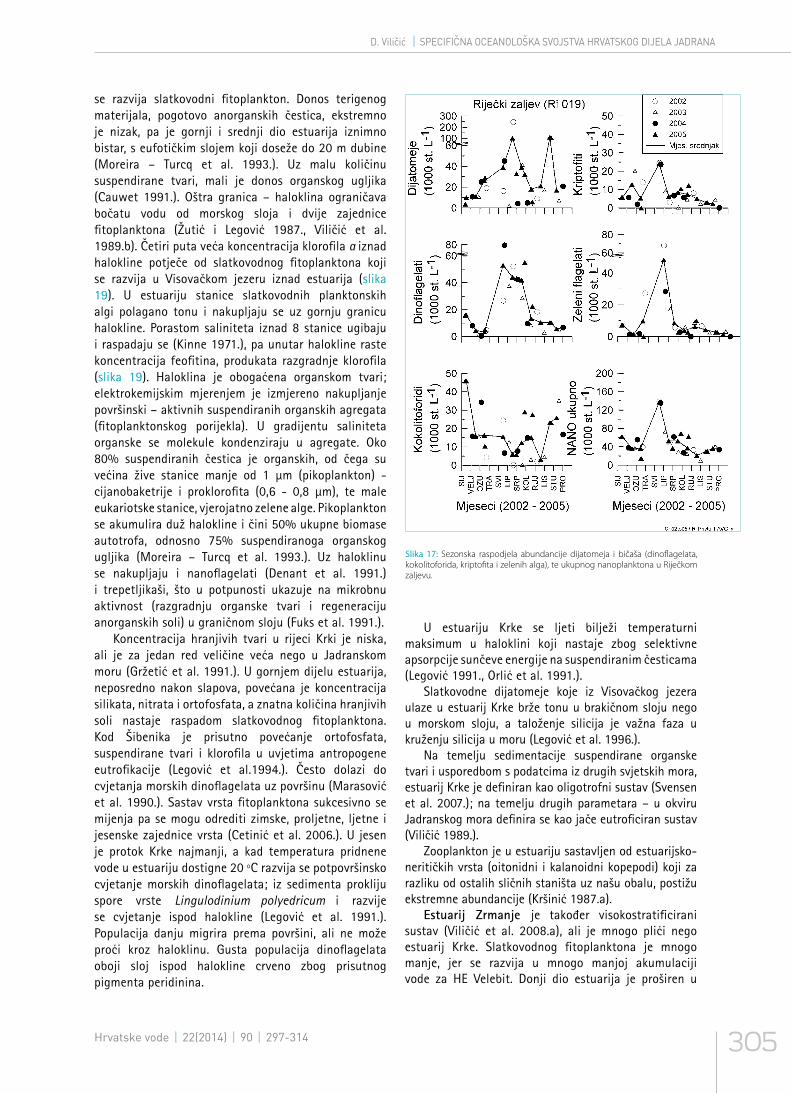
Slika 17: Sezonska raspodjela abundancije dijatomeja i bičaša (dinoflagelata, kokolitoforida, kriptofita i zelenih alga), te ukupnog nanoplanktona u Riječkom zaljevu.
se razvija slatkovodni fitoplankton. Donos terigenog materijala, pogotovo anorganskih čestica, ekstremno je nizak, pa je gornji i srednji dio estuarija iznimno bistar, s eufotičkim slojem koji doseže do 20 m dubine (Moreira – Turcq et al. 1993.). Uz malu količinu suspendirane tvari, mali je donos organskog ugljika (Cauwet 1991.). Oštra granica – haloklina ograničava bočatu vodu od morskog sloja i dvije zajednice fitoplanktona (Žutić i Legović 1987., Viličić et al. 1989.b). Četiri puta veća koncentracija klorofila a iznad halokline potječe od slatkovodnog fitoplanktona koji se razvija u Visovačkom jezeru iznad estuarija (slika 19). U estuariju stanice slatkovodnih planktonskih algi polagano tonu i nakupljaju se uz gornju granicu halokline. Porastom saliniteta iznad 8 stanice ugibaju i raspadaju se (Kinne 1971.), pa unutar halokline raste koncentracija feofitina, produkata razgradnje klorofila (slika 19). Haloklina je obogaćena organskom tvari; elektrokemijskim mjerenjem je izmjereno nakupljanje površinski – aktivnih suspendiranih organskih agregata (fitoplanktonskog porijekla). U gradijentu saliniteta organske se molekule kondenziraju u agregate. Oko 80% suspendiranih čestica je organskih, od čega su većina žive stanice manje od 1 µm (pikoplankton) - cijanobaketrije i proklorofita (0,6 - 0,8 µm), te male eukariotske stanice, vjerojatno zelene alge. Pikoplankton se akumulira duž halokline i čini 50% ukupne biomase autotrofa, odnosno 75% suspendiranoga organskog ugljika (Moreira – Turcq et al. 1993.). Uz haloklinu se nakupljaju i nanoflagelati (Denant et al. 1991.) i trepetljikaši, što u potpunosti ukazuje na mikrobnu aktivnost (razgradnju organske tvari i regeneraciju anorganskih soli) u graničnom sloju (Fuks et al. 1991.).
Koncentracija hranjivih tvari u rijeci Krki je niska, ali je za jedan red veličine veća nego u Jadranskom moru (Gržetić et al. 1991.). U gornjem dijelu estuarija, neposredno nakon slapova, povećana je koncentracija silikata, nitrata i ortofosfata, a znatna količina hranjivih soli nastaje raspadom slatkovodnog fitoplanktona. Kod Šibenika je prisutno povećanje ortofosfata, suspendirane tvari i klorofila u uvjetima antropogene eutrofikacije (Legović et al.1994.). Često dolazi do cvjetanja morskih dinoflagelata uz površinu (Marasović et al. 1990.). Sastav vrsta fitoplanktona sukcesivno se mijenja pa se mogu odrediti zimske, proljetne, ljetne i jesenske zajednice vrsta (Cetinić et al. 2006.). U jesen je protok Krke najmanji, a kad temperatura pridnene vode u estuariju dostigne 20 ºC razvija se potpovršinsko cvjetanje morskih dinoflagelata; iz sedimenta prokliju spore vrste Lingulodinium polyedricum i razvije se cvjetanje ispod halokline (Legović et al. 1991.). Populacija danju migrira prema površini, ali ne može proći kroz haloklinu. Gusta populacija dinoflagelata oboji sloj ispod halokline crveno zbog prisutnog pigmenta peridinina.
U estuariju Krke se ljeti bilježi temperaturni maksimum u haloklini koji nastaje zbog selektivne apsorpcije sunčeve energije na suspendiranim česticama (Legović 1991., Orlić et al. 1991.).
Slatkovodne dijatomeje koje iz Visovačkog jezera ulaze u estuarij Krke brže tonu u brakičnom sloju nego u morskom sloju, a taloženje silicija je važna faza u kruženju silicija u moru (Legović et al. 1996.).
Na temelju sedimentacije suspendirane organske tvari i usporedbom s podatcima iz drugih svjetskih mora, estuarij Krke je definiran kao oligotrofni sustav (Svensen et al. 2007.); na temelju drugih parametara – u okviru Jadranskog mora definira se kao jače eutroficiran sustav (Viličić 1989.).
Zooplankton je u estuariju sastavljen od estuarijsko-neritičkih vrsta (oitonidni i kalanoidni kopepodi) koji za razliku od ostalih sličnih staništa uz našu obalu, postižu ekstremne abundancije (Kršinić 1987.a).
Estuarij Zrmanje je također visokostratificirani sustav (Viličić et al. 2008.a), ali je mnogo plići nego estuarij Krke. Slatkovodnog fitoplanktona je mnogo manje, jer se razvija u mnogo manjoj akumulaciji vode za HE Velebit. Donji dio estuarija je proširen u
306 Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
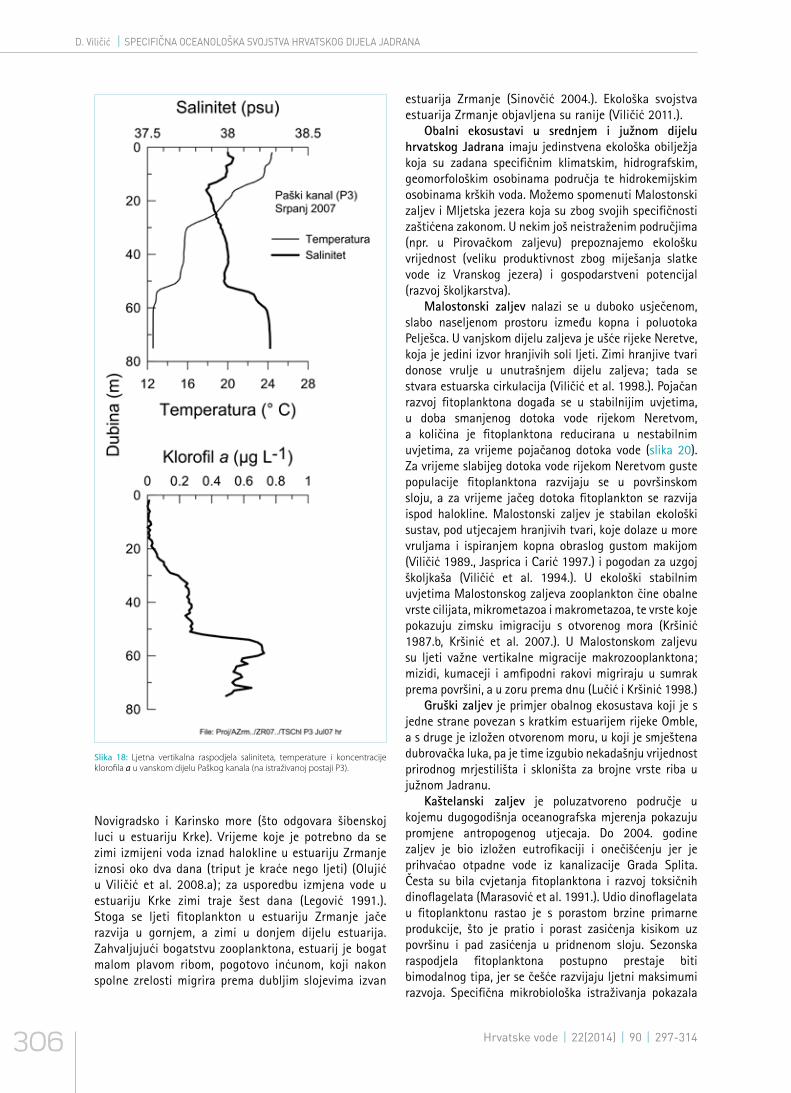
Slika 18: Ljetna vertikalna raspodjela saliniteta, temperature i koncentracije klorofila a u vanskom dijelu Paškog kanala (na istraživanoj postaji P3).
Novigradsko i Karinsko more (što odgovara šibenskoj luci u estuariju Krke). Vrijeme koje je potrebno da se zimi izmijeni voda iznad halokline u estuariju Zrmanje iznosi oko dva dana (triput je kraće nego ljeti) (Olujić u Viličić et al. 2008.a); za usporedbu izmjena vode u estuariju Krke zimi traje šest dana (Legović 1991.). Stoga se ljeti fitoplankton u estuariju Zrmanje jače razvija u gornjem, a zimi u donjem dijelu estuarija. Zahvaljujući bogatstvu zooplanktona, estuarij je bogat malom plavom ribom, pogotovo inćunom, koji nakon spolne zrelosti migrira prema dubljim slojevima izvan
estuarija Zrmanje (Sinovčić 2004.). Ekološka svojstva estuarija Zrmanje objavljena su ranije (Viličić 2011.).
Obalni ekosustavi u srednjem i južnom dijelu hrvatskog Jadrana imaju jedinstvena ekološka obilježja koja su zadana specifičnim klimatskim, hidrografskim, geomorfološkim osobinama područja te hidrokemijskim osobinama krških voda. Možemo spomenuti Malostonski zaljev i Mljetska jezera koja su zbog svojih specifičnosti zaštićena zakonom. U nekim još neistraženim područjima (npr. u Pirovačkom zaljevu) prepoznajemo ekološku vrijednost (veliku produktivnost zbog miješanja slatke vode iz Vranskog jezera) i gospodarstveni potencijal (razvoj školjkarstva).
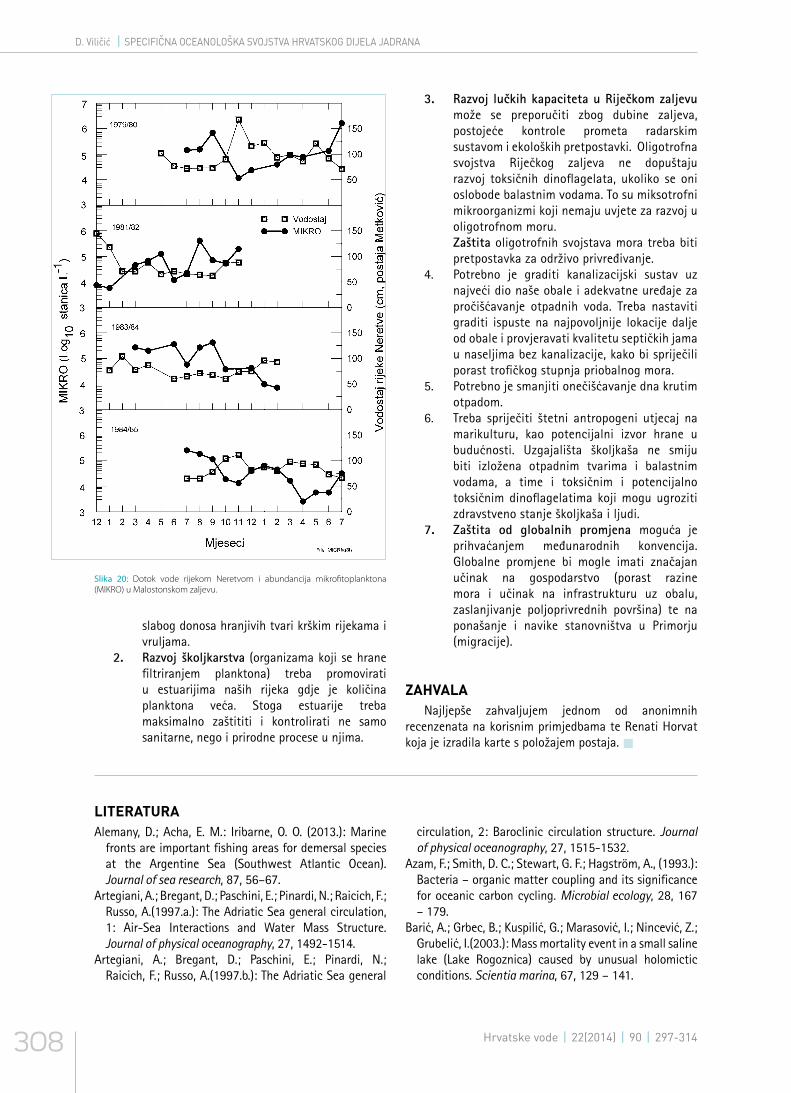
Malostonski zaljev nalazi se u duboko usječenom, slabo naseljenom prostoru između kopna i poluotoka Pelješca. U vanjskom dijelu zaljeva je ušće rijeke Neretve, koja je jedini izvor hranjivih soli ljeti. Zimi hranjive tvari donose vrulje u unutrašnjem dijelu zaljeva; tada se stvara estuarska cirkulacija (Viličić et al. 1998.). Pojačan razvoj fitoplanktona događa se u stabilnijim uvjetima, u doba smanjenog dotoka vode rijekom Neretvom, a količina je fitoplanktona reducirana u nestabilnim uvjetima, za vrijeme pojačanog dotoka vode (slika 20). Za vrijeme slabijeg dotoka vode rijekom Neretvom guste populacije fitoplanktona razvijaju se u površinskom sloju, a za vrijeme jačeg dotoka fitoplankton se razvija ispod halokline. Malostonski zaljev je stabilan ekološki sustav, pod utjecajem hranjivih tvari, koje dolaze u more vruljama i ispiranjem kopna obraslog gustom makijom (Viličić 1989., Jasprica i Carić 1997.) i pogodan za uzgoj školjkaša (Viličić et al. 1994.). U ekološki stabilnim uvjetima Malostonskog zaljeva zooplankton čine obalne vrste cilijata, mikrometazoa i makrometazoa, te vrste koje pokazuju zimsku imigraciju s otvorenog mora (Kršinić 1987.b, Kršinić et al. 2007.). U Malostonskom zaljevu su ljeti važne vertikalne migracije makrozooplanktona; mizidi, kumaceji i amfipodni rakovi migriraju u sumrak prema površini, a u zoru prema dnu (Lučić i Kršinić 1998.)
Gruški zaljev je primjer obalnog ekosustava koji je s jedne strane povezan s kratkim estuarijem rijeke Omble, a s druge je izložen otvorenom moru, u koji je smještena dubrovačka luka, pa je time izgubio nekadašnju vrijednost prirodnog mrjestilišta i skloništa za brojne vrste riba u južnom Jadranu.
Kaštelanski zaljev je poluzatvoreno područje u kojemu dugogodišnja oceanografska mjerenja pokazuju promjene antropogenog utjecaja. Do 2004. godine zaljev je bio izložen eutrofikaciji i onečišćenju jer je prihvaćao otpadne vode iz kanalizacije Grada Splita. Česta su bila cvjetanja fitoplanktona i razvoj toksičnih dinoflagelata (Marasović et al. 1991.). Udio dinoflagelata u fitoplanktonu rastao je s porastom brzine primarne produkcije, što je pratio i porast zasićenja kisikom uz površinu i pad zasićenja u pridnenom sloju. Sezonska raspodjela fitoplanktona postupno prestaje biti bimodalnog tipa, jer se češće razvijaju ljetni maksimumi razvoja. Specifična mikrobiološka istraživanja pokazala
307Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
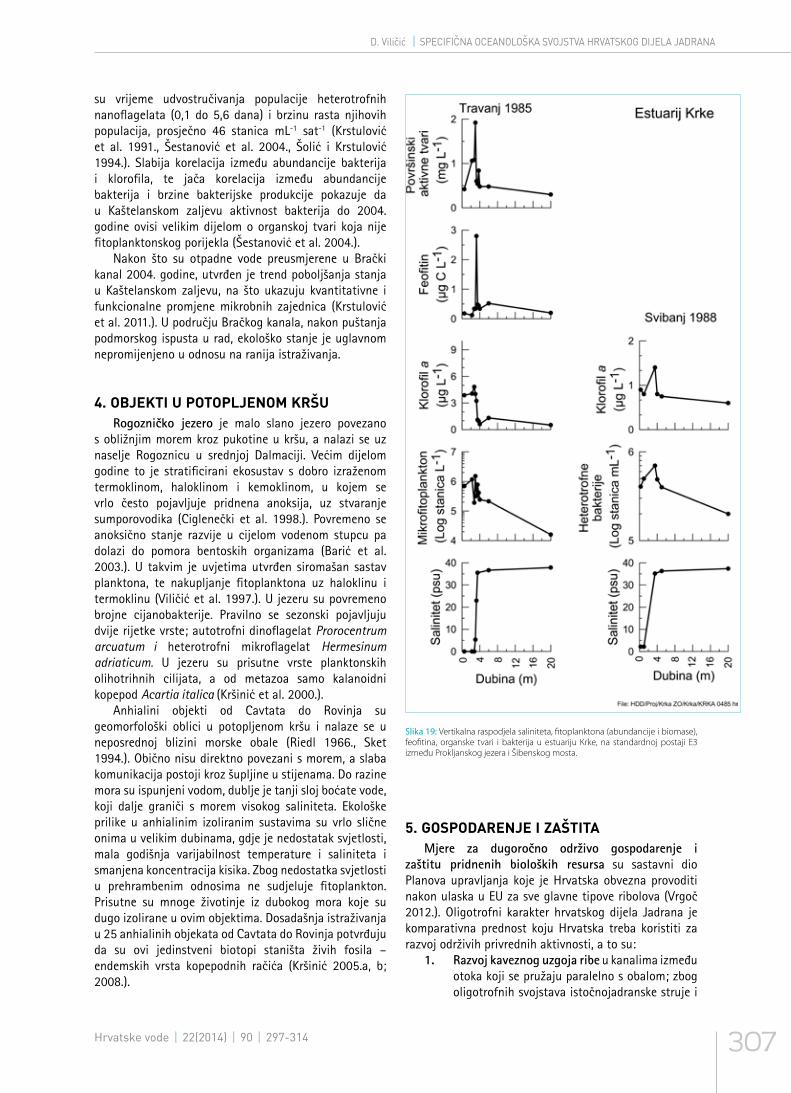
Slika 19: Vertikalna raspodjela saliniteta, fitoplanktona (abundancije i biomase), feofitina, organske tvari i bakterija u estuariju Krke, na standardnoj postaji E3 između Prokljanskog jezera i Šibenskog mosta.
su vrijeme udvostručivanja populacije heterotrofnih nanoflagelata (0,1 do 5,6 dana) i brzinu rasta njihovih populacija, prosječno 46 stanica mL-1 sat-1 (Krstulović et al. 1991., Šestanović et al. 2004., Šolić i Krstulović 1994.). Slabija korelacija između abundancije bakterija i klorofila, te jača korelacija između abundancije bakterija i brzine bakterijske produkcije pokazuje da u Kaštelanskom zaljevu aktivnost bakterija do 2004. godine ovisi velikim dijelom o organskoj tvari koja nije fitoplanktonskog porijekla (Šestanović et al. 2004.).
Nakon što su otpadne vode preusmjerene u Brački kanal 2004. godine, utvrđen je trend poboljšanja stanja u Kaštelanskom zaljevu, na što ukazuju kvantitativne i funkcionalne promjene mikrobnih zajednica (Krstulović et al. 2011.). U području Bračkog kanala, nakon puštanja podmorskog ispusta u rad, ekološko stanje je uglavnom nepromijenjeno u odnosu na ranija istraživanja.
4. ObjEKTI U POTOPLjENOM KRŠU Rogozničko jezero je malo slano jezero povezano
s obližnjim morem kroz pukotine u kršu, a nalazi se uz naselje Rogoznicu u srednjoj Dalmaciji. Većim dijelom godine to je stratificirani ekosustav s dobro izraženom termoklinom, haloklinom i kemoklinom, u kojem se vrlo često pojavljuje pridnena anoksija, uz stvaranje sumporovodika (Ciglenečki et al. 1998.). Povremeno se anoksično stanje razvije u cijelom vodenom stupcu pa dolazi do pomora bentoskih organizama (Barić et al. 2003.). U takvim je uvjetima utvrđen siromašan sastav planktona, te nakupljanje fitoplanktona uz haloklinu i termoklinu (Viličić et al. 1997.). U jezeru su povremeno brojne cijanobakterije. Pravilno se sezonski pojavljuju dvije rijetke vrste; autotrofni dinoflagelat Prorocentrum arcuatum i heterotrofni mikroflagelat Hermesinum adriaticum. U jezeru su prisutne vrste planktonskih olihotrihnih cilijata, a od metazoa samo kalanoidni kopepod Acartia italica (Kršinić et al. 2000.).
Anhialini objekti od Cavtata do Rovinja su geomorfološki oblici u potopljenom kršu i nalaze se u neposrednoj blizini morske obale (Riedl 1966., Sket 1994.). Obično nisu direktno povezani s morem, a slaba komunikacija postoji kroz šupljine u stijenama. Do razine mora su ispunjeni vodom, dublje je tanji sloj boćate vode, koji dalje graniči s morem visokog saliniteta. Ekološke prilike u anhialinim izoliranim sustavima su vrlo slične onima u velikim dubinama, gdje je nedostatak svjetlosti, mala godišnja varijabilnost temperature i saliniteta i smanjena koncentracija kisika. Zbog nedostatka svjetlosti u prehrambenim odnosima ne sudjeluje fitoplankton.Prisutne su mnoge životinje iz dubokog mora koje su dugo izolirane u ovim objektima. Dosadašnja istraživanja u 25 anhialinih objekata od Cavtata do Rovinja potvrđuju da su ovi jedinstveni biotopi staništa živih fosila – endemskih vrsta kopepodnih račića (Kršinić 2005.a, b; 2008.).
5. gOSPODARENjE I ZAŠTITAMjere za dugoročno održivo gospodarenje i
zaštitu pridnenih bioloških resursa su sastavni dio Planova upravljanja koje je Hrvatska obvezna provoditi nakon ulaska u EU za sve glavne tipove ribolova (Vrgoč 2012.). Oligotrofni karakter hrvatskog dijela Jadrana je komparativna prednost koju Hrvatska treba koristiti za razvoj održivih privrednih aktivnosti, a to su:
1. Razvoj kaveznog uzgoja ribe u kanalima između otoka koji se pružaju paralelno s obalom; zbog oligotrofnih svojstava istočnojadranske struje i
308 Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Slika 20: Dotok vode rijekom Neretvom i abundancija mikrofitoplanktona (MIKRO) u Malostonskom zaljevu.
slabog donosa hranjivih tvari krškim rijekama i vruljama.
2. Razvoj školjkarstva (organizama koji se hrane filtriranjem planktona) treba promovirati u estuarijima naših rijeka gdje je količina planktona veća. Stoga estuarije treba maksimalno zaštititi i kontrolirati ne samo sanitarne, nego i prirodne procese u njima.
3. Razvoj lučkih kapaciteta u Riječkom zaljevu može se preporučiti zbog dubine zaljeva, postojeće kontrole prometa radarskim sustavom i ekoloških pretpostavki. Oligotrofna svojstva Riječkog zaljeva ne dopuštaju razvoj toksičnih dinoflagelata, ukoliko se oni oslobode balastnim vodama. To su miksotrofni mikroorganizmi koji nemaju uvjete za razvoj u oligotrofnom moru.
Zaštita oligotrofnih svojstava mora treba biti pretpostavka za održivo privređivanje.
4. Potrebno je graditi kanalizacijski sustav uz najveći dio naše obale i adekvatne uređaje za pročišćavanje otpadnih voda. Treba nastaviti graditi ispuste na najpovoljnije lokacije dalje od obale i provjeravati kvalitetu septičkih jama u naseljima bez kanalizacije, kako bi spriječili porast trofičkog stupnja priobalnog mora.
5. Potrebno je smanjiti onečišćavanje dna krutim otpadom.
6. Treba spriječiti štetni antropogeni utjecaj na marikulturu, kao potencijalni izvor hrane u budućnosti. Uzgajališta školjkaša ne smiju biti izložena otpadnim tvarima i balastnim vodama, a time i toksičnim i potencijalno toksičnim dinoflagelatima koji mogu ugroziti zdravstveno stanje školjkaša i ljudi.
7. Zaštita od globalnih promjena moguća je prihvaćanjem međunarodnih konvencija. Globalne promjene bi mogle imati značajan učinak na gospodarstvo (porast razine mora i učinak na infrastrukturu uz obalu, zaslanjivanje poljoprivrednih površina) te na ponašanje i navike stanovništva u Primorju (migracije).
ZAhVALANajljepše zahvaljujem jednom od anonimnih
recenzenata na korisnim primjedbama te Renati Horvat koja je izradila karte s položajem postaja.
LITERATURAAlemany, D.; Acha, E. M.: Iribarne, O. O. (2013.): Marine
fronts are important fishing areas for demersal species at the Argentine Sea (Southwest Atlantic Ocean). Journal of sea research, 87, 56–67.
Artegiani, A.; Bregant, D.; Paschini, E.; Pinardi, N.; Raicich, F.; Russo, A.(1997.a.): The Adriatic Sea general circulation, 1: Air-Sea Interactions and Water Mass Structure. Journal of physical oceanography, 27, 1492-1514.
Artegiani, A.; Bregant, D.; Paschini, E.; Pinardi, N.; Raicich, F.; Russo, A.(1997.b.): The Adriatic Sea general
circulation, 2: Baroclinic circulation structure. Journal of physical oceanography, 27, 1515-1532.
Azam, F.; Smith, D. C.; Stewart, G. F.; Hagström, A., (1993.): Bacteria – organic matter coupling and its significance for oceanic carbon cycling. Microbial ecology, 28, 167 – 179.
Barić, A.; Grbec, B.; Kuspilić, G.; Marasović, I.; Nincević, Z.; Grubelić, I.(2003.): Mass mortality event in a small saline lake (Lake Rogoznica) caused by unusual holomictic conditions. Scientia marina, 67, 129 – 141.

309Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Batistić, M.; Kršinić, F.; Jasprica, N.; Carić, M.; Viličić, D.; Lučić, D.(2004.): Gelatinous invertebrate zooplankton of the South Adriatic: species composition and vertical distribution. Journal of plankton research, 26, 459-474.
Batistić, M.; Jasprica, N.; Carić, M.; Čalić, M.; Kovačević, V.; Garić R.; Njire J.; Mikuš J.; Bobanović-Ćolić, S. (2012.): Biological evidence of a winter convection event in the South Adriatic: A phytoplankton maximum in the aphotic zone. Continental shelf research, 44, 57-71.
Benović, A.(1973.): Diurnal vertical migration of Solmissum albescens (Hydromedusae) in the Southern Adriatic. Marine biology, 18, 298-301.
Borzelli, G.; Manzella, G.; Marullo, S.; Santoleri, R. (1999.): Observations of coastal filaments in the Adriatic Sea. Journal of marine system 20, 187 – 203.
Cauwet, G. (1991.): Carbon inputs and biogeochemical processes at the halocline in a stratified estuary: Krka River. Marine chemistry, 32, 269 – 283.
Cetinić, I.; Viličić, D.; Burić, Z.(2004.): Microphytoplankton composition across the frontal syste in the northern Adriatic (February, 2003). Rapport de la Commission internationale pour l’exploration scientifique de la Mer Méditerranée, 37, 270.
Cetinić, I.; Viličić, D.; Burić, Z.; Olujić, G.(2006.): Phytoplankton seasonality in a highly stratified karstic estuary (Krka, Adriatic Sea). Hydrobiologia, 555, 31 – 40.
Chiaudani, G.; Marchetti, R.; Vighi, M. (1980.): Eutrophication in Emilia – Romagna coastal waters (North Adriatic Sea, Italy): A case hystory. Progress in water technology, 12, 185 – 192.
Ciglenečki, I.; Kodba, Z.; Viličić, D.; Ćosović, B. (1998.): Seasonal variation of anoxic conditions in the Rogoznica Lake. Croatica chemica acta, 71, 217-232.
Civitarese, G. i Gačić, M. (2001.): Had the eastern mediterranean transient an impact on the new production in the southern Adriatic. Geophysical research letters, 28, 1627 – 1630.
Civitarese, G.; Gačić, M.; Lipizer, M.; Eusebi Borzelli, G. L. (2010.): On the impact of the Bimcicodal Oscillating System (BiOS) on the biogeochemistry and biology of the Adriatic and Ionian Seas (Eastern mediterranean). Biogeosciences, 7, 3987 – 3997.
Crisciani, F.; Ferraro, S.; Raicich, F. (1991.): Climatological remarks on the algal bloom episodes of 1729 and 1991 in the northern Adriatic Sea. Bollettino di oceanologia teorica ed applicata, 9, 367 – 370.
Cushman-Roisin, B.; Gačić, M.; Poulain, P.-M.; Artegiani, A. (2001.): Physical oceanography of the Adriatic Sea. Kluwer Academic Publishers, Dordrecht, Neederlands.
Degobbis, D.; Gilmartin, M. (1990.): Nitrogen, phosphorus and biogenic silicon budgets for the northern Adriatic Sea. Oceanologica acta, 13, 31 – 45.
Degobbis, D.; Malej, A.; Fonda–Umani, S.(1999.): The mucilage phenomenon in the northern Adriatic Sea. A critical review of the present scientific hypotheses. Annali dell Istituto superiore di sanita,35, 373–381.
Degobbis, D.; Precali, R.; Ivančić, I.; Smodlaka, N.; Fuks, D.; Kveder, S. (2000.): Long–term changes in the northern Adriatic ecosystem related to anthropogenic eutrophication. International journal of environment and pollution, 13, 495–533.
Denant, V.; Saliot, A.; Mantoura, R.F.C. (1991.): Distribution of algal chlorophyll and carotenoid pigments in a stratified estuary: the Krka river, Adriatic Sea. Marine chemistry, 32, 285 – 297.
Dorman, C. E.; Carniel, S.; Cavaleri, L.; Sclavo, M.; Chiggiato, J.; Doyle, J.; Haack, T.; Pullen, J.; Grbec, B.; Vilibić, I.; Janeković, I.; Lee, C.; Malačić, V.; Orlić, M.; Paschini, E.; Russo, A.; Signell, R. P. (2006.): February 2003. marine atmospheric conditions and the bora over the northern Adriatic. Journal of geophysical research: oceans, 111, C03S03.
Dulčić, J. (1997.): Effects of environmental changes on early stages and reproduction of anchovy (Engraulis encsicolus L.) in the Adriatic Sea. Annales: anali za istarske in mediteranske študije. Series historia naturalis,11, 259 – 270.
Đakovac, T.; Degobbis, D.; Supić, N.; Precali, R. (2012.): Marked reduction of eutrophication pressure in the northeastern Adriatic in the period 2000.-2009. Estuarine coastal and shelf science, 115, 25-32.
Faganeli, J.; Mohar, B.; Kofol, R.; Pavlica, V.; Marinšek, T.; Rozman, A.; Kovač, N.; Šurca Vuk, A. (2010.): Nature and lability of northern Adriatic macroaggregates. Marine Drugs, 8, 2480–2492.
France, J.; Mozetič, P. (2006.): Ecological characterization of toxic phytoplankton species (Dinophysis spp., Dinophyceae) in Slovenian mariculture areas (Gulf of Trieste, Adriatic Sea) and the implications for monitoring. Marine pollution bulletin, 52, 1504 – 1516.
Fuks, D.; Devescovi, M.; Precali, R.; Krstulović, N; Šolić, M. (1991.): Bacterial abundance and activity in the highly stratified estuary of the Krka river. Marine chemistry, 32, 333 – 346.
Gačić, M.; Civitarese, G.; Ursella, L. (1999.): Spatial and seasonal variability of water and biochemical fluxes in the Adriatic Sea. In: Malanotte–Rizoli, P, Eremeev, V.N. (eds.), The eastern Mediterranean as a laboratory basin for the assessment of contrasting ecosystems, 335–357. Kluwer Academic Publ., Amsterdam.
Gačić, M.; Civitarese, G.; Miserocchi, S.; Cardin, V.; Crise, A.; Mauri, E. (2002.): The open ocean convection in the southern Adriatic: a controlling mechanism of the spring phytoplankton bloom. Continental shelf research, 22, 1897 – 1908.
Gajić-Čapka, M.; Perčec-Tadić, M.; Patarčić, M. (2003.): Digitalna godišnja oborinska karta Hrvatske. Hrvatski meteorološki časopis, 38, 21 – 33.
Gašparović, B.; Ćosović, B. (2001.): Distribution of surface – active substances in the northern Adriatic Sea. Marine chemistry, 75, 301 – 313.
Grbec, B.; Morović, M.; Dulčić, J.; Marasović, I.; Ninčević, Ž. (2008.): Impact of the climatic change on the Adriatic

310 Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Sea Ecosystem. Fresenius environmental bulletin, 17, 10a Special Issue, 1615-1620.
Gržetić, Z.; Precali, R.; Degobbis, D.; Škrivanić, A. (1991.): Nutrient enrichment and phytoplankton response in an Adriatic karstic estuary. Marine chemistry, 32, 313 – 331.
Heissenberger, A.; Leppard, G. G.; Herndl, G. J. (1996.): Ultrastructure of marine snow. II. Microbiological considerations. Marine ecology progress series, 135, 299 – 308.
Hure, J.; Scotto di Carlo, B. (1969.): Copepodi pelagici dell’Adriatico settentrionale nel periodo Gennaio-dicembre 1965. Pubblicazioni della Stazione zoological di Napoli, 37, 173-195.
Hure, J.; Kršinić, F.(1998.): Planktonic copepods of the Adriatic Sea. Natura croatica, 7, Suppl. 2, 1 - 135.
Hurrell J. W.(1995.): Decadal trends in the North Atlantic Oscillation index and relationship to regional temperature and precipitation. Science, 269, 676 – 679.
Ivančić, I.; Godrijan, J.; Pfannkuchen, M.; Marić, D.; Gašparović, B.; Djakovac, T.; Najdek, M. (2012.): Survival mechanisms of phytoplankton in conditions of stratification-induced deprivation of orthophosphate: Northern Adriatic case study. Limnology and oceanography, 57, 1721 – 1731.
Jasprica, N.; Carić, M. (1997.): A comparison of phytoplankton biomass estimators and their environmental correlates in the Mali Ston Bay (southern Adriatic). Marine ecology, 18, 35 – 50.
Jasprica, N.; Carić, M.; Viličić, N. (2001.): Relationships of subsurface chlorophyll maximum to diatoms and other microphytoplankton in the southern Adriatic Sea. Proceedings of the 16. International diatom symposyum, Athens, 365 – 379.
Jeffries, M. A.; Lee, C. M. (2007.): A climatology of the northern Adriatic Sea’s response to bora and river forcing, Journal of geophysical research, 112, C03S02, 1-18.
Kinne, O. (1971.): Marine ecology 1 (2). Inter – Research, Lüneburg, Germany.
Kniewald, G.; Marguš, D.; Mihelčić, G. (2006.): Formation of the Krka river estuary in Croatia and the travertine barrier phenomenon. In: Briand, F. (ed.) Fluxes of small and medium-size Meediterranean rivers: impact on coastal areas. CIESM workshop monographs, 30, 93 – 96.
Kraus, R.; Supić, N. (2011.): Impact of circulation on high phytoplankton blooms and fish catch in the northern Adriatic (1990.-2004.). Estuarine coastal and shelf science, 91, 198-210.
Krstulović, N.; Šolić, M.; Marasović, I.; Odžak, N. (1991.): Bacteria and their distribution under red – tide conditions. Kieler Meeresforschung, 8, 284 – 289.
Krstulović, N.; Šolić, M.; Šegvi, D.; Šestanović, S; Kušpilić, G. (2011.): Praćenje utjecaja podmorskog ispusta Stobreć na okoliš. Hrvatske vode 76, 127 - 132.
Kršinić, F. (1987.a.): Tintinnines (Ciliophora, Oligotrichida, Tintinnina) in eastern Adriatic bays. Estuarine coastaland shelf science, 24, 527 - 538.
Kršinić, F. (1987.b.): On the ecology of tintinnines in the Bay of Mali Ston (Eastern Adriatic). Estuarine coastal and shelf science, 24, 401 - 418.
Kršinić, F. (1998.): Vertical distribution of protozoan and microcopepod communities in the South Adriatic Pit. Journl of plankton research, 20, 1033 – 1060.
Kršinić, F. (2005.a.): Speleohvarella gamulini gen. et sp. nov., new copepod (Calanoida, Stephidae) from an anchialine cave in the Adriatic Sea. Journal of plankton research, 27, 607-615.
Kršinić, F. (2005.b.): Badijella jalzici – a new genus and species of calanoid copepod (Calanoida, Ridgewayiidae) from an anchialine cave on the Croatian Adriatic coast. Marine biological research, 1, 281-289.
Kršinić, F. (2008.): Description of Speleophria mestrovi sp. nov., a new copepod (Misophrioida) from an anchialine cave in the Adriatic Sea. Marine biological research, 4, 304 – 312.
Kršinić, F.; Carić, M.; Viličić, D.; Ciglenečki – Jušić, I.(2000.): Calanoid copepod Acartia italica Steuer phenomenon in the small saline Rogoznica lake (eastern Adriatic coast, Croatia). Journal of plankton research, 22, 1441 – 1464.
Kršinić, F.; Bojanić, D.; Precali, R.; Kraus, R. (2007.): Quantitative variability of the copepod assemblages in the northern Adriatic Sea from 1993. to 1997. Estuarine coastal and shelf science, 74, 528 – 538.
Kuzmić, M. (1991.): Exploring the effects of bura over the northern Adriatic: CZCS imagery and a mathematical model prediction. International journal of remote sensing, 12, 207 – 214.
Kuzmić, M.; Janeković, I.; Book, J. W.; Martin, P. J.; Doyle, J. D. (2006.): Modeling the northern Adriatic double-gyre response to intense bora wind: A revisit. Journal of geophysical research oceans, 111, S313.
Leder, N.; Smirčić, A.; Gržetić, Z.; Grbec, B.; Vilibić, I. (1996.): Dinamička svojstva akvatorija Palagruškog praga. Zbornik Palagruža (ur. Hodžić, M.), 339 – 343, Matica hrvatska, Split.
Lee, C. M.; Askari, F.; Book, J.; Carniel, S.; Cushman-Roisin, B.; Dorman, C.; Doyle, J.; Flament, P.; Harris, C.K.; Jones, B.H.; Kuzmic, M.; Martin, P.; Ogston, A.; Orlic, M.; Perkins, H.; Poulain, P.-M.; Pullen, J.; Russo, A.; Sherwood, C.; Signell, R. P.; Thaler D. (2005.): Northern Adriatic response to a wintertime bora wind event. Eos, Transactions american geophysical union, 86, 157 – 165.
Legović, T. (1991.): Exchange of water in a stratified estuary with an application to Krka (Adriatic Sea). Marine chemistry, 32, 121 – 135.
Legović, T.; Viličić, D.; Petricioli, D.; Žutić, V. (1991.): Subsurface Gonyaulax polyedra bloom in a stratified estuary. Marine chemistry, 32, 361 – 374
Legović, T.; Žutić, V.; Gržetić, Z.; Cauwet, G.; Precali, R.; Viličić, D. (1994.): Eutrophication in the Krka estuary. Marine chemistry, 46, 203 – 215.
Legović, T.; Žutić, V.; Viličić, D.; Gržetić, Z. (1996.): Transport of silica in a stratified estuary. Marine chemistry, 53, 69 – 80.

311Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Legović, T.; Justić, D. (1997.): When do phytoplankton blooms cause the most intense hypoxia in the northern Adriatic sea? Oceanologica acta, 20, 91 – 99.
Lučić, D.; Kršinić, F. (1998.): Annual variability of mesozooplankton assemblages in Mali Ston Bay (Southern Adriatic). Periodicum biologorum, 100, 43 - 52.
Lučić, D.; Benović, A.; Batistić, M.; Njire, J.; Onofri, V. (2005.): Calycophorae (Siphonophora) in the open waters of the central and southern Adriatic Sea during spring 2002. Journal of the marine biological association of the United Kingdom, 85, 495-501.
Lyons, D. M.; Supic, N.; Smodlaka, N. (2007.): Geostrophic circulation patterns in the northeastern Adriatic Sea and the effects of air-sea coupling: May-September 2003. Journal of geophysical research, 112, C03S08.
Malanotte – Rizzoli, P.; Bergamasco, A. (1989.): The circulation of the eastern Mediterranean. Oceanologica acta, 12, 335 – 352.
Manca, B.; Giorgetti, A. (1999.): Flow patterns of the main water masses across transversal areas in the southern Adriatic Sea: Seasonal variability. In: Malanotte – Rizoli, P., Eremeev, V.N. (eds.), The eastern Mediterranean as a laboratory basin for the assessment of contrasting ecosystems, 495 – 506. Kluwer Academic Publ., Amsterdam.
Marasović, I.; Pucher – Petković, T.; Alegria Hernandez, V. (1988.): Phytoplankton productivity of the Adriatic sea in relation to pelagic fisheries. Bilješke – Notes of the Institute of oceanography and fisheries Split, 72, 1 – 8.
Marasović, I.; Pucher – Petković, T.; Petrova – Karadjova, V. (1990.): Prorocentrum minimum in the Adriatic and Black Sea. Journal of the Marine biological association of the United Kingdom, 70, 473 – 476.
Marasović, I.; Gačić, M.; Kovačević, V.; Krstulović, N.; Kušpilić, G.; Pucher – Petković, T.; Odžak, N.; Šolić, M. (1991.): Development of the red tide in the Kastela Bay (Adriatic Sea). Marine chemistry, 32, 375 – 385.
Marchetti, R.; Provini, A.; Crosa, G. (1989.): Nutrient load carried by the river Po into the Adriatic sea, 1968 – 87. Marine pollution bulletin, 20, 168 – 172.
Marić, D.; Kraus, R.; Godrijan, J.; Supić, N.; Đakovac, T.; Precali, R. (2012.): Phytoplankton response to climatic and anthropogenic influences in the north-eastern Adriatic during the last four decades. Estuarine coastal and shelf science, 1, 1-12.
Mihanović, H.; Vilibić, I.; Carniel, S.; Tudor, M.; Russo, A.; Bergamasco, A.; Bubić, N.; Ljubešić, Z.; Viličić, D.; Boldrin, A.; Malačić, V.; Celio, M.; Comici, C.; Raicich, F. (2013.): Exceptional dense water formation in the Adriatic shelf in the winter of 2012. Ocean science, 9, 561-572.
Moreira – Turcq, P.; Martin, J. M.; Fleury, A. (1993.): Chemical and biological characterization of particles by flow cytometry in the Krka estuary, Croatia. Marine chemistry, 43, 115 – 126.
Mozetič, P.; Solidoro, C.; Cossarini, G.; Socal, G.; Precali, R.; France, J.; Bianchi, F.; De Vittor, C.; Smodlaka, N.; Fonda Umani, S. (2009.): Recent trends towards
oligotrophication of the northern Adriatic: evidence from chlorophyll a time series. Estuaries and coasts, 33, 362 – 375
MZOP (2012.): Početna procjena stanja i opterećenja morskog okoliša hrvatskog dijela Jadrana. Ministarstvo zaštite okoliša i prirode RH i Institut za oceanografiju i ribarstvo, Split. http://www.mzoip.hr/doc/More/Pocetna_procjena_morski_okolis.pdf
Najdek, M.; Blažina, M.; Djakovac, T.; Kraus, R. (2005.): The role of the diatom Cylindrotheca closterium in a mucilage event in the northern Adriatic sea: coupling with high salinity water intrusions. Journal of plankton research, 27, 851 – 862.
Orlić, M. (2001.): Croatian coastal waters. In: Cushman-Roisin, B., Gačić, M., Poulain, P.-M., Artegiani, A. (eds.), Physical oceanography of the Adriatic Sea: past, present and future, 189-214. Kluwer Academic Publishers, Dordrecht, Neederlands.
Orlić, M.; Ferenčak, M.; Gržetić, Z.; Limić, N.; Pasarić, Z.; Smirčić, A. (1991.): High – frequency oscillations observed in the Krka Estuary. Marine chemistry, 32, 137 – 151.
Orlić, M.; Gačić, M.; Laviolette, P. E. (1992.): The currents and circulation of the Adriatic Sea. Oceanologica acta, 15, 109 – 124.
Orlić, M.; Kuzmić, M; Pasarić, Z. (1994.): Response of the Adriatic Sea to bora and sirocco forcing. Continental shelf research, 14, 91 – 116.
Penzar, B.; Penzar, I.; Orlić, M. (2001.): Vrijeme i klima na hrvatskom Jadranu, Feletar, Zagreb.
Peters, H.; Lee, C; M., Orlić; M., Dorman; C. E. (2007.): Turbulence in the wintertime northern Adriatic Sea under strong atmospheric forcing. Journal of geophysical research oceans, 112, C03S09.
Polimene, L.; Pinardi, N.; Zavatarelli, M.; Colella, S. (2006.): The Adriatic Sea ecosystem seasonal cycle: Validation of a three-dimensional numerical model. Journal of geophysical research oceans, 111, C03S19.
Poulain, P.–M.; Raicich, F.(2001.): Forcings. In: Cushman–Roisin, B., Gačić, M., Poulain, P.–M., Artegiani, A. (eds.), Physical oceanography of the Adriatic Sea, 45–65. Kluwer Academic Publishers, Dordrecht, Neederlands.
Precali, R.; Šolić, M.; Šurmanović, D.; Ćosić-Flajsig, G. (2003.): Stanje kakvoće priobalnog mora Hrvatske. Zbornik radova 3. Hrvatske konferencije o vodama (ur. D. Gereš), 435 – 442, MTG-topograf, Velika Gorica.
Pucher–Petković, T.; Zore – Armanda (1973.): Essai d’évaluation et pronostic de la production en fonction des facteurs du milieu dans l’Adriatique. Acta Adriatica, 15, 1 – 37.
Regner, S. (1996.): Effects of environmental changes on early stages and reproduction of anchovy in the Adriatic Sea. Scientia marina, 60, 167 – 177
Revelante, N.; Gilmartin, M. (1977.): The effects of northern italian rivers and eastern mediterranean ingressions on the phytoplankton of the Adriatic Sea. Hydrobiologia, 56, 229 – 240.

312 Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Riedl, R. (1966.): Biologie der Meereshöhlen. Paul Parey, Berlin, Deutschland.
Samuel, S.; Haines, K.; Josey, S.; Myers, P. G. (1999.): Response of the Mediterranean Sea thermohaline circulation to observed changes in the winter wind stress field in the period 1980 – 1993. Journal of geophysical research Oceans, 104, 7771 – 7784.
Santojanni, A.; Arneri, E.; Bernardini, V.; Cingolani, N.; Di Marco; M., Russo, A. (2006.): Effects of environmental variables on recruitment of anchovy in the Adriatic Sea. Climate research, 31, 181 – 193.
Sinovčić, G. (2004.): Growth and length-weight relationship of the juvenile anchovy, Engraulis encrasicolus, in the nursery ground (Zrmanja River estuary – eastern Adriatic Sea). Journal of applied ichthyology, 20, 79 – 80.
Sket, B. (1994.): Distribution patterns of some subterranean crustacea in the territory of the former Yugoslavia. Hydrobiologia,287, 65-75.
Smayda, T. J. (1997.): What is a bloom? A commentary.Limnology and ocenaography 42, 1132 – 1136.
Smodlaka, N. (1986.): Primary production of the organic matter as an indicator of the eutrophication in the northern Adriatic Sea. Science of the total environment, 56, 211 – 220.
Supić, N.; Orlić, M.; Degobbis, D. (2000.): Istrian coastal counter-current and its year-to-year variability. Estuarine coastal and shelf science, 50, 385 – 397.
Supić, N.; Kraus, R.; Kuzmić, M.; Paschini, E.; Precali, R.; Russo, A.; Vilibić, I. (2012.): Predictability of northern Adriatic winter conditions. Journal of marine systems, 90, 42-57.
Svensen, C.; Viličić, D.; Wassmann, P.; Arashkevich, E.; Ratkova, T. (2007.): Plankton distribution and vertical flux of biogenic matter during high summer stratification in the Krka Estuary (Eastern Adriatic). Estuarine coastal and shelf science, 71, 381 – 390.
Šestanović, S.; Šolić, M.; Krstulović, N.; Ninčević, Ž. (2004.): Seasonal and vertical distribution of planktonic bacteria and heterotrophic nanoflagellates in the middle Adriatic Sea. Helgoland marine research, 58, 83 – 92.
Šolić, M.; Krstulović, N. (1994.): Role of predation in controlling bacterial and heterotrophic nanoflagellate standing stocks in the coastal Adriatic Sea: seasonal patterns. Marine ecology progress series, 114, 219 – 235.
Šolić, M.; Krstulović, N.; Marasović, I.; Baranović, A.; Pucher – Petković, T.; Vučetić, T. (1997.): Analysis of time series of planktonic communities in the Adriatic Sea: distinguishing between natural and man – induced changes. Oceanologica acta, 20, 131 – 143.
Ursella, L.; Gačić, M. (2001.): Use of the Acoustic doppler current profiler (ADCP) in the study of the circulation of the Adriatic Sea. Annals of geophysics, 19, 1183 – 1193.
Vilibić, I.; Orlić, M. (2002.): Adriatic water masses, their rates of formation and transport through the Otranto Strait. Deep-sea research, I 49, 1321 – 1340.
Vilibić, I.; Grbec, B.; Supić, N. (2004.): Dense water generation in the north Adriatic in 1999 and its
recirculation along the Jabuka Pit. Deep-sea research, I 51, 1457-1474.
Vilibić, I.; Matijević, S.; Šepić, J.; Kušpilić, G. (2012.): Changes in the Adriatic oceanographic properties induced by the Eastern Mediterranean Transient. Biogeosciences, 9, 2085-2097.
Vilibić, I.; Supić, N. (2005.): Dense water generation on a shelf: the case of the Adriatic Sea. Ocean Dynamics 55, 403–415.
Viličić, D. (1989.): Phytoplankton population density and volume as indicators of eutrophication in the eastern part of the Adriatic Sea. Hydrobiologia, 174, 117 – 132.
Viličić, D. (1991.): A study of phytoplankton in the Adriatic Sea after the July 1984 bloom. Internationale revue der gesamten hydrobiologie, 76, 197-211.
Viličić, D. (2011.): Estuarij Zrmanje i njegova ekološka svojstva. Hrvatske vode, 77, 201-214.
Viličić, D.; Vučak, Z.; Škrivanić; A., Gržetić, Z. (1989.a.): Phytoplankton blooms in the oligotrophic open south Adriatic waters. Marine chemistry, 28, 89 – 107.
Viličić, D.; Legović, T.; Žutić, V.(1989b.): Vertical distribution of phytoplankton in a stratified estuary. Aquatic sciences, 51, 31 – 46.
Viličić, D.; Mušin, D.; Jasprica, N.(1994.): Interrelations between hydrographic conditions, nanoplankton and bivalve larvae in the Mali Ston Bay (southern Adriatic). Acta Adriatica, 34, 55 – 64.
Viličić, D.; Marasović, I.; Kušpilić, G. (1997.): The heterotrophic ebridian microflagellate Hermesinum adriaticumZach. in the Adriatic sea. Archieve fuer protistenkunde, 147, 373 – 379.
Viličić, D.; Jasprica, N.; Carić, M.; Burić, Z.(1998.): Taxonomic composition and seasonal distribution of microphytoplankton in Mali Ston Bay (eastern Adriatic). Acta botanica Croatica, 57, 29 – 48.
Viličić, D.; Bosak, S.; Burić, Z.; Caput-Mihalić, K. (2007.): Phytoplankton seasonality and composition along the coastal NE Adriatic Sea during the extremely low Po River discharge in 2006. Acta botanica Croatica, 66, 101 - 115.
Viličić, D.; Terzić, S.; Ahel, M.; Burić, Z.; Jasprica, N.; Carić, M.; Caput-Mihalić K.; Olujić, G. (2008.a.):Phytoplankton abundance and pigment biomarkers in the oligotrophic, eastern Adriatic estuary. Environmental monitoring and assessment,142, 199 – 218.
Viličić, D.; Orlić, M.; Jasprica, N. (2008.b): Deep chlorophyll maximum in the coastal northeastern Adriatic Sea, July 2007. Acta botanica Croatica, 67, 33 – 43.
Viličić, D.; Đakovac, T.; Burić, Z.; Bosak, S. (2009.): Composition and annual cycle of phytoplankton assemblages in the northeastern Adriatic Sea. Botanica marina, 52, 291 – 305.
Viličić, D.; Kuzmić, M.; Tomažić, I.; Ljubešić, Z.; Bosak, S.; Precali, R.; Djakovac, T.; Marić, D.; Godrijan, J. (2013.): Northern Adriatic phytoplankton response to short Po River discharge pulses during summer stratified conditions. Marine ecology, 34, 389 – 405.

313Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
Vrgoč, N.; Arneri, E.; Jukić-Peladić, S.; Krstulović Šifner, S.; Mannini, P.; Marčeta, B.; Osmani, K.; Piccinetti C.; Ungaro, N. (2004.): Review of current knowledge on shared demersal stocks of the Adriatic Sea. AdriaMed technical document 12, 1-91.
Vrgoč, N. (2012.): Hrvatsko morsko ribarstvo - stanje i perspektive na pragu EU-a. Elaborat za projekt COAST Očuvanje i održivo korištenje biološke raznolikosti na dalmatinskoj obali. GEF/UNDP, Ministarstvo poljoprivrede RH, Uprava ribarstva, Zagreb.
Vukić Lušić, D.; Peršić, V.; Horvatić, J.; Viličić, D.; Traven, L.; Djakovac T.; Mićović, V. (2008.):Assessment of nutrient limitation in Rijeka Bay, NE Adriatic Sea,using miniaturized bioassay. Journal of experimental marine biology and ecology, 358, 46 - 56.
Zavatarelli, M.; N. Pinardi; V. H. Kourafalou; Maggiore, A. (2002.): Diagnostic and prognostic model studies of the Adriatic Sea circulation: Seasonal variability. Journal of geophysical research Oceans, 107, C1 3004.
Zore – Armanda, M. (1969.): Water exchange between the Adriatic and the eastern Mediterranean. Deep-sea research, 16, 171 – 178.
Zore – Armanda, M. (1984.): Hydrographic and productivity conditions of the Palagruža region in the middle Adriatic. Acta Adriatica, 25, 119 – 138.
Žutić, V.; Legović, T. (1987.): A film of organic matter at the freshwater/seawater interface of an estuary. Nature, 328, 612 – 614.
Žutić, V.; Svetličić, V. (2000.): Interfacial processes. In: Hutzinger, O. (ed.), The handbook of environmental chemistry 5, 149–165. Springer Verlag, Berlin, Germany.

314 Hrvatske vode 22(2014) 90 297-314
D. viličić SpecifičnA OceAnOlOškA SvOJStvA hRvAtSkOg DiJelA JADRAnA
SPECIfIC OCEANOLOgICAL ChARACTERISTICS Of ThE CROATIAN PART Of ThE AdRIATIC
Abstract. Important characteristics of the Adriatic Sea are the following: a) position of the Po River which is the largest source of fresh water and nutrients in the Mediterranean; b) winter bora winds which participate in the creation of the Northern Adriatic dense water and initiate circulation in the eastern part of the Mediterranean; c) East Adriatic current which brings oligotrophic water from the Eastern Mediterranean along the Croatian coast; d) Istrian front; e) longitudinal and transverse gradient of distribution of physico-chemical and biological characteristics; f) phytoplankton bloom with appearance of matter in organic macro-aggregates in the Northern Adriatic; g) small difference between ebb and tide; h) karst hydrogeology and existence of highly stratified estuaries along the Croatian coast; i) larger share of small-size fractions of phytoplankton in the total primary biomass, and a larger share of bacterial biomass in the total microbial biomass in the Southern Adriatic than in the Northern Adriatic; j) isolated habitats originating from karst submersion. The most important characteristic of the Eastern Adriatic (which differentiates it from the Western Adriatic) is its oligotrophy, which is a comparative advantage which Croatia needs to use for development of appropriate economic activities.
Key words: Adriatic Sea, oceanographic characteristics, environmental characteristics, the pelagic, management, protection
SPEZIfISChE OZEANOLOgISChE EIgENSChAfTEN dER KROATISChEN AdRIA
Zusammenfassung. Die wichtigen Eigenschaften des Adriatischen Meeres sind: a) die Lage des Flusses Po, der die größte Quelle für Trinkwasser und Nährstoffe im Mittelmeer ist; b) Winterbora, die an der Entstehung von dickem Wasser in der Nordadria teilnimmt und die Zirkulation im östlichen Mittelmeer fördert; c) die ostadriatische Strömung, mit der oligothropisches Wasser aus dem östlichen Mittelmeers entlang der kroatischen Küste fließt; d) die istrianische Front; e) der Längen- und Quergradient der Verteilung von physikalisch-chemischen und biologischen Merkmalen; f) Phytoplanktonblüte mit dem Vorkommen von Makroaggregaten von organischen Stoffen in der Nordadria; g) kleiner Unterschied zwischen Flut und Ebbe; h) Karsthydrogeologie und hoch stratifizierte Ästuarien entlang der kroatischen Küste; i) ein höher Anteil kleinerer Phytoplanktonfraktionen an der gesamten Primärbiomasse sowie ein höher Anteil der Biomasse von Bakterien an der insgesamt vorhandenen mikrobiellen Biomasse in der Südadria als in der Nordadria; j) unterirdische, isolierte Habitate im Karst. Das wichtigste Merkmal der Ostadria, wodurch sich die Ostadria von der Westadria unterscheidet, ist die Oligotrophie, was ein Wettbewerbsvorteil ist, den Kroatien zur Entwicklung entsprechender Wirtschaftsaktivitäten nutzen sollte.
Schlüsselwörter: die Adria, ozeanologische Eigenschaften, ökologische Merkmale, Pelagial, Bewirtschaftung, Schutz





























