Embed Size (px)
Citation preview

Naunyn-Schmiedeberg's Arch. Pharmacol. 282, 33--44 (1974) �9 by Springer-Verlag 1974
Stimulation of Cyclic Adenosine 3',5'-Monophosphate Accumulation and Lipolysis in Fat Cells
by Adenosine Deaminase
U. Sehwabe and R. Ebe r t
Institut ffir Pharmakologie, Medizinisehe Hochschule Hannover, Hannover-Kleefeld Germany
Received October 26, 1973
Summary. The basal lipolytic activity of isolated fat cells of the rat was greatly enhanced in the presence of 0.01 to 30 t~g adenosine deaminase (ADA) per ml. This effect was more pronounced in dilute (20000 cells/ml) than in concentrated cell suspensions (100000 cells/ml); this is possibly due to the presence, in the incubation medium, of a high concentration of inosine which is formed by the deamination of the large amounts of adenosine released from high concentrations of fat cells. Inosine, although less potent than adenosine as an antilipolytic agent, markedly inhibited ADA-induced lipolysis at concentrations between 10 to 100 ~M. The lipolytic effect of ADA was identical with the stimulation of lipolysis by 1 ~M noradrenaline or 1 mM theophylline, while 1 mM dibutyryl cyclic AMP yielded two-fold higher values. The effects of ADA and lipolytic agents at maximally stimulating concentrations were not additive.
After 5 rain of incubation maximally effective concentrations of ADA which were also maximal with respect to lipolysis caused a 3- to 6-fold elevation of cyclic AMP levels in fat cells. A similar increase was observed with maximally effective concentrations of theophylline, whereas noradrenaline produced a 100- to 200-fold elevation. This indicates that a small accumulation of cyclic AMP may be sufficient to trigger the full lipolytic response. Furthermore, ADA, like theophylline, acted synergistically with norach'enaline and prevented the fall of cyclic AMP levels during 30 min of incubation.
Insulin (100 tLU/ml) and nicotinic acid (0.1 ~M) decreased cyclic AMP accumula- tion and glycerol production induced by ADA.
The results support the hypothesis that adenosine is released from isolated fat cells and that this nucleoside may serve as an inhibitor of adenyl cyclase activity, thus regulating cyclic AMP-dependent processes in adipose tissue.
Key words: Cyclic AMP -- Adenosine -- Lipolysis -- Fat Cell -- Adenosine Deaminase.
Adenosine is cont inuous ly released by fat cells in to the incuba t ion med ium in amoun t s which inh ib i t cyclic AMP accumula t ion and lipolysis due to noradrenal ine (Schwabe et al., 1973). The inh ib i to ry effect of adenosine was diminished when dilute cell suspensions, conta in ing only 20000 fat cells/ml, were used (Schwabe and Eber t , 1972). Under these condit ions the effect of hormones on cyclic AMP levels and lipolysis was
3 Naunyn-Schmiedeberg's Arch. Pharmacol., Vol. 282

34 U. Schwabe and R. Ebert
great ly increased. Adenosine has been shown to inhibt basal and hor- mone-st imulated adenyl cyclase in fat cell ghosts (Fain et al., 1972; MeKenzie and B~r, 1973). Therefore, it has been suggested tha t adenosine might serve as a physiological feedback regulator of cyclic AMP metab- olism in fat cells (Fain et al., 1972; Fain, 1973a; Schwabe et al., 1973).
One line of evidence for the adenosine release was the finding that , after the addit ion of adenosine deaminase (ADA) to the incubat ion medium, the inhibi tory ac t iv i ty of the medium was destroyed (Schwabe et al., 1973). Therefore, this enzyme appeared to be a useful tool to remove adenosine from the medium during the incubat ion and to obtain fur ther insight into the role of this nucleoside in cyclic AMP-dependent reactions in fat cells. We were encouraged to s tudy the effect of this enzyme on fat cells, since Fain (1973b) recently showed t h a t upon ad- dition of A D A to incubated fat cells a marked potent ia t ion occurred of the cyclic AMP accumulat ion and lipolysis caused by low concentrat ions of noradrenaline. I n this paper we present studies on the effect of A D A on cyclic AMP accumulat ion in the absence of hormonal st imulat ion and in the presence of antilipolytic agents.
Methods Fat Cell Preparation. Isolated fat cells were prepared according to Rodbell
(1964) from epididymal adipose tissue of male rats (Sprague Dawley NIH/Han; Zentralinstitut fiir Versuchstierzucht, Hannover-Linden) weighing 140--160g. Adipose tissue was treated with 1 mg collagenase per ml. Distribution and incuba- tion of the fat cell suspension were performed as previously described (Sehwabe et al., 1973).
Measurement o] Cyclic AMP. Incubations (volume: 1 ml) were terminated by addition of 1 ml 10% trichloroacetic acid and cyclic AMP was extracted as pre- viously described (Schwabe and Ebert, 1972). Cyclic AM/) was determined by the protein binding method of Gilman (1970). Dilute fat cell suspensions (20000 cells/ml) contained 0.6 to 1 pmoles cyclic AMP per ml. For reasons discussed earlier (Schwabe and Ebert, 1972) 0.015 to 0.025 pmoles cyclic AMP had to be determined with adequate accuracy, when 25 ~l of the incubation mixture were analysed. Therefore, the calibration curve was determined with 0.2 pmoles cyclic 3H-cAMP and 0.02 to 2 pmoles unlabelled cyclic AMP in a total volume of 200 ~1: In order to minimize reagent blanks, the calibration standards were dissolved in the incubation medium for the fat cells (Krebs-Ringer bicarbonate buffer, pH 7.4 containing 20/0 bovine serum albumin), treated with trichloroacetic acid and carried through the same extraction procedure as the fat cell suspensions. 40 pmoles of unlabelled cyclic AMP were added to 1 ml of acid-treated fat cell suspension as recovery standards. The average recovery rate was 98.5 ={= 1.3~ (mean 4- S.E., n = 21) and, therefore, no corrections for recovery were made.
.Measurement of Glycerol. Incubations (volume: 1 ml) were terminated after 60 rain (unless stated otherwise) by the addition of i ml 10~ trichloroaeetic acid. Lipolysis was determined by measurement of glycerol according to Lambert and Neish (1950), using the colorimetric determination of formaldehyde as described by ~ash (1953).
Effect of Adenosine Deaminase on Fat Cells 35
Materials. Bovine serum albumin (fraction V, Serva Feinbiochemica) was purified by charcoal treatment according to Chen (1967) ; crude bacterial collagenase (lot 1 JD, Worthington Biochemical Corporation) ; adenosine deaminase (Boehringer Mannheim GmbH) from calf intestine (specific activity of 200 U/rag) and nucleoside phosphorylase (Boehringer Mannheim GmbH) from calf spleen (25 U/rag) were diluted 100-fold with Krebs-Ringer bicarbonate buffer, pH 7.4 containing 2~ albumin the day before use and dialyzed overnight against 100 volumes of Krebs- Ringer bicarbonate buffer in order to remove the high content of ammonium sul- phate; adenosine-SH(G) 3',5'-cyclic phosphate (24.1 Ci/mmole, NEN Chemicals Gmbtt) was checked for radioehemieal purity by thin layer chromatography on cellulose with n-butanol/methanol/ethylacetate/25~ ammonium hydroxide, 7 :3:4:6 (v/v) ; cyclic adenosine 3',5'-monophosphate, cyclic N~,O2-dibutyryl adenosine 3',5'-monophosphate and inosine (Boehringer Mannheim GmbH); (-)-noradrenaline hydrochloride (Schuehardt GmbIt) ; nicotinic acid (E. Merck AG) ; crystalline zinc insulin (Farbwerke I-Ioechst AG, Op. Nr. C-033, 28 units/rag); theophylline (E. Merck AG) was dissolved in water without any additive. All other chemicals were reagent grade or best commercially available.
Results
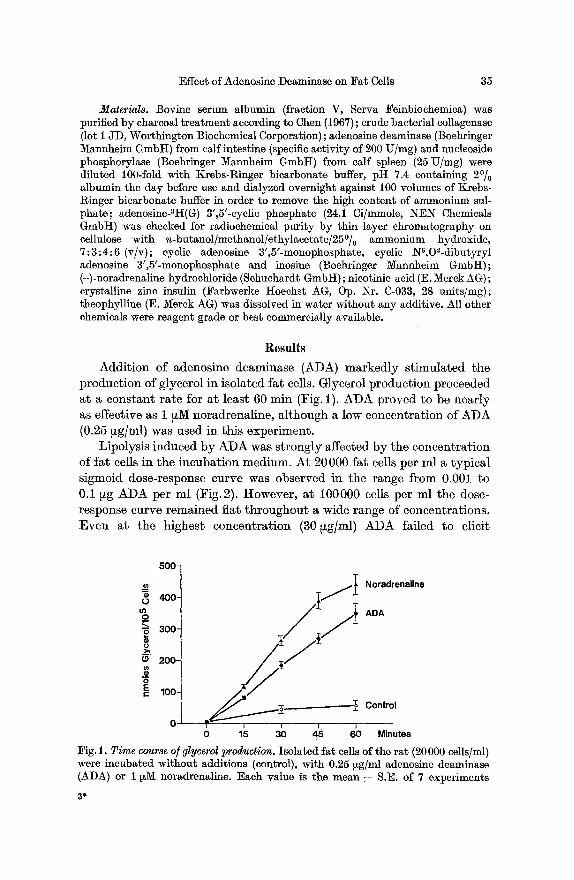
Addition of adenosine deaminase (ADA) markedly stimulated the production of glycerol in isolated fat cells. Glycerol production proceeded at a constant rate for at least 60 rain (Fig. 1). ADA proved to be nearly as effective as 1 ~M noradrenaline, although a low concentration of ADA (0.25 ~g/ml) was used in this experiment.
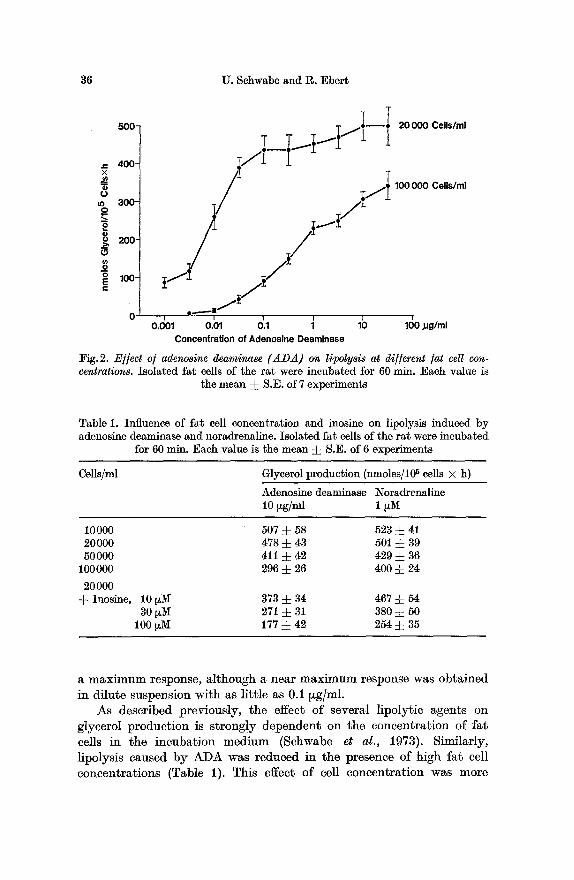
Lipolysis induced by ADA was strongly affected by the concentration of fat cells in the incubation medium. At 20000 fat cells per ml a typical sigmoid dose-response curve was observed in the range from 0.001 to 0.1 ~g ADA per ml (Fig.2). However, at 100000 cells per ml the dose- response curve remained flat throughout a wide range of concentrations. Even at the highest concentration (30 ~g/ml) ADA failed to elicit
500 - r
4 0 o -
LO 0
300-
200-
o E , - 1 0 0 -
t j l Noradrenaline
J ~ i ADA
Control
0 15 30 45 60 Minutes ~ig. 1. Time course o] glycerol production. Isolated fat cells of the rat (20 000 cells/ml) were incubated without additions (control), with 0.25 ~g/ml adenosine deaminase (ADA) or 1 ~ noradrenaline. Each value is the mean • S.E. of 7 experiments
8*
36 U. Schwabe and R. Ebert
oo. t ooo ~
,~ 400-
100 000 Cells/ml o
m ~ 300-
- / ~ "100- C
/ I
0,001 0.01 0.1 1 10 100 .ug/ml Concentration of Adenosine Deaminase
Fig.2. E//ect o/ adenosine dsaminase (ADA) on l@olysis at di//erent /at cell con- centrations. Isolated fat cells of the rat were incubated for 60 rain. Each value is
the mean ! S.E. of 7 experiments
Table 1. Influence of fat cell concentration and inosine on lipolysis induced by adenosine deaminase and noradrenaline. Isolated fat cells of the rat were incubated
for 60 rain. Each value is the mean • S.E. of 6 experiments
Cells/ml Glycerol production (nmoles/105 cells • h)
Adenosine deaminase Noradrenaline 10 ~g/ml 1 ~M
10000 507 • 58 523 ~ 41 20000 478 ~ 43 501 ~ 39 50000 411 ~ 42 429 ~ 36
100000 296 -4- 26 400 ~ 24
20000 Inosine, 10 tz]YI 373 • 34 467 ~ 54
30 ~ 271 • 31 380 ~ 50 100 ~ 177 ~ 42 254 i 35
a m a x i m u m response, a l though a near m a x i m u m response was ob ta ined in di lute suspension with as l i t t le as 0.1 ~g/ml.
As described previously, the effect of several lipoly~ic agents on glycerol product ion is s t rongly dependent on the concent ra t ion of fat cells in the incuba t ion med ium (Schwabe et al., 1973). Similarly, lipolysis caused by ADA was reduced in the presence of high fat cell concentra t ions (Table 1). This effect of cell concentra t ion was more
Effect of Adenosine Deaminase on Fat Cells 37
100 000 Cetls/ml E
= I •
0
E
0 ' 1 Control 0.(}01 0.01 0'.1 1 ' 10 jug/ml
Concentration of Adenosine Deaminase
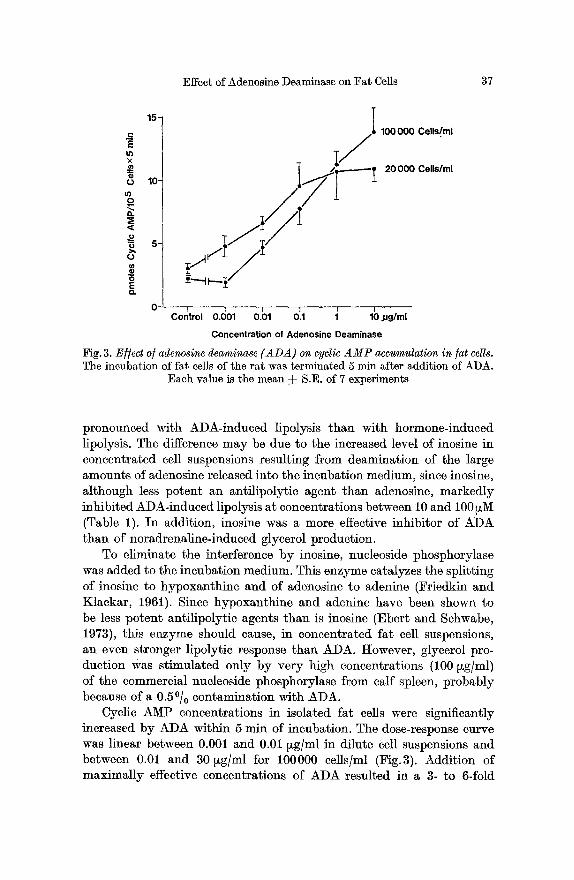
Fig. 3. E//ect o/adenosine deaminase (ADA) on cyclic AMP accumulation in fat cells. The incubation of fat cells of the rat was terminated 5 min after addition of ADA.
Each value is the mean =]= S.E. of 7 experiments
pronounced with ADA-induced lipolysis than with hormone-induced lipolysis. The difference may be due to the increased level of inosine in concentrated cell suspensions resulting from deamination of the large amounts of adenosine released into the incubation medium, since inosine, although less potent an antilipolytic agent than adenosine, markedly inhibited ADA-indnced lipolysis at concentrations between 10 and 100 ~ (Table 1). In addition, inosine was a more effective inhibitor of ADA than of noradrenaline-induced glycerol production.
To eliminate the interference by inosine, nueleoside phosphorylase was added to the incubation medium. This enzyme catalyzes the splitting of inosine to hypoxanthine and of adenosine to adenine (Friedkin and Klaekar, 1961). Since hypoxanthine and adenine have been shown to be less potent antilipolytic agents than is inosine (Ebert and Schwabe, 1973), this enzyme should cause, in concentrated fat cell suspensions, an even stronger lipolytic response than ADA. l~owever, glycerol pro- duction was stimulated only by very high concentrations (100 ~g/ml) of the commercial nucleoside phosphorylase from calf spleen, probably because of a 0.50/0 contamination with ADA.
Cyclic AMP concentrations in isolated fat cells were significantly increased by ADA within 5 rain of incubation. The dose-response curve was linear between 0.001 and 0.01 ~g/ml in dilute cell suspensions and between 0.01 and 30 ~g/ml for 100000 cells/ml (Fig.3). Addition of maximally effective concentrations of ADA resulted in a 3- to 6-fold
38 U. Sehwabe and R. Ebert
J= • r 0 o
tO 0
2 o~
r _r e E t-
500"
400"
300 -
200 -
100-
0
j ::tJlTl 0 & 0'.1 0'.3 4
.E E
1200-
o
IO o 800- O.
,<
.o. 400- o
"6 C E i i
0.03 o.1 0'.3 j,M Concentration of Noradranaline
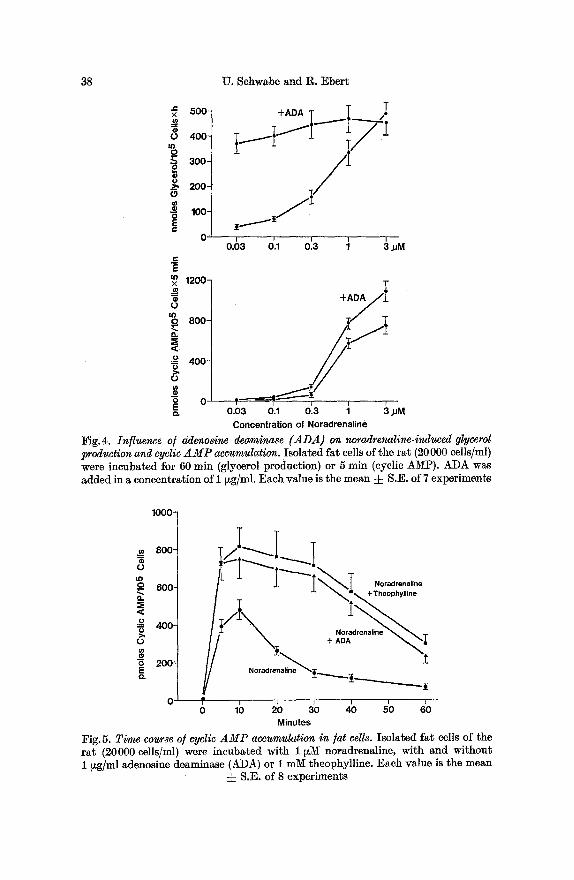
Fig.4. Influence of adenosine deaminase (ADA) on noradrenaline-indueed glycerol production and cyclic A M P accumulation. Isolated fat cells of the rat (20000 cells/ml) were incubated for 60 rain (glycerol production) or 5 rain (cyclic AMP). ADA was added in a concentration of 1 Izg/ml. Each value is the mean • S.E. of 7 experiments
(J to 0
a .
,r o
0
"6 E e~
1000-
800-
600-
400-
200 ~ Noradrenal ine x ~ ~ ~[
0 1; 20 3JO 40 5? dO Minutes
Fig.5. Time course o/cyclic A M P accumulation in ]at cells. Isolated fat cells of the rat (20000 cells/ml) were incubated with 1 ~M noradrenaline, with and without 1 ~g/ml adenosine deaminase (ADA) or 1 mM theophylline. Each value is the mean
=j= S.E. of 8 experiments
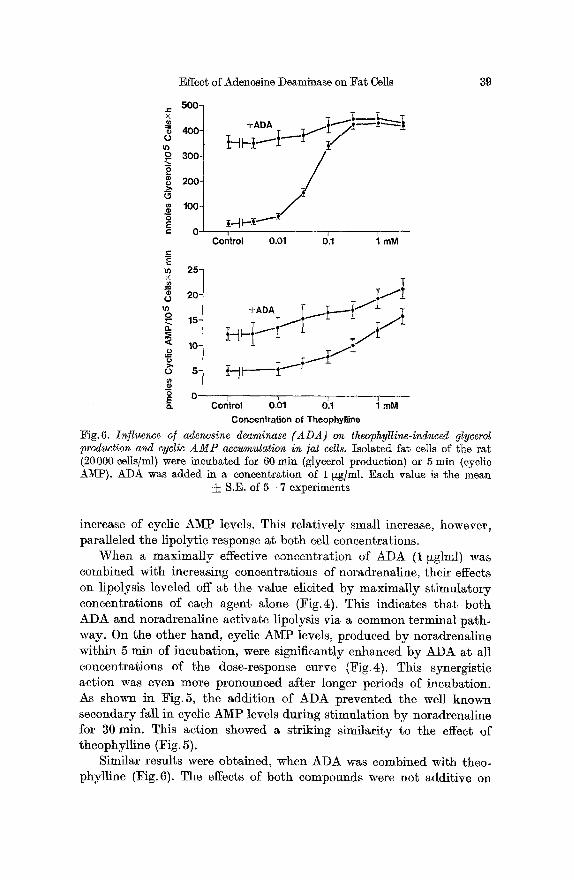
Effect of Adenosine Deaminase on Fat Cells 39
0
o =o ~D
C
500-
400-
300-
200-
100-
0
E 25-
•
=~ 20- to o 15- t l .
<~ 10- o
0 5-
o E
Control 0.01 0.1 1 mM
*AOA
jt/S 0 I J
Controt 0.01 OJ.1 1 mM Concentration of "Theophylline
Fig.6. InJtuence o] adenosine deaminase (ADA) on theophylline-induced glycerol production and cyclic A MP accumulation in ]at cells. Isolated fa t cells of the r a t (20000 eells/ml) were incubated for 60 rain (glycerol production) or 5 rain (cyclic AMP). ADA was added in a concentrat ion of 1 ~g/ml. Each value is the mean
4- S.E. of 5 - -7 experiments
increase of cyclic AMP levels. This relatively small increase, however, paralleled the lipolytic response at both cell concentrations.
When a maximally effective concentration of ADA (1 ~glml) was combined with increasing concentrations of noradrenaline, their effects on lipolysis leveled off at the value elicited by maximally stimulatory concentrations of each agent alone (Fig.4). This indicates tha t both ADA and noradrenaline activate lipolysis via a common terminal path- way. On the other hand, cyclic AMP levels, produced by noradrenaline within 5 rain of incubation, were significantly enhanced by ADA at all concentrations of the dose-response curve (Fig.4). This synergistic action was even more pronounced after longer periods of incubation. As shown in Fig. 5, the addition of ADA prevented the well known secondary fall in cyclic AMP levels during stimulation by noradrenaline for 30 rain. This action showed a striking similarity to the effect of theophylline (Fig. 5).
Similar results were obtained, when ADA was combined with theo- phylline (Fig. 6). The effects of both compounds were not additive on
40 U. Schwabe and R. Ebert
1000 -
= oo-
o 2 600
~ , t / ~ 1 - - ~
o 200'- E
!--4 ~-]~ ~-- ' ']P""'} 0 , j
Control 0101 0.1 1 mM Concentration of Dibutyryl Cyclic AMP
Fig. 7. E//ect o/ adenosine deaminase (ADA) on lipolysis stimulated by dibutyryl cyclic AMP. Isolated fat cells of the rat were incubated at a concentration of 20000 cells/ml for 60 rain, ADA was added in a maximally effective concentration
of i Ezg/ml. Each value is the mean q- S.E. of 6 experiments
Table 2. Effect of antilipolytic agents on lipolysis and cyclic AMP accumulation induced by adenosine deaminase in isolated fat cells of the rat (20000 cells/ml).
Each value is the mean • S.E., number of experiments in parentheses
Addition Glycerol production nmoles/10 ~ cells • h
Cyclic AMP level pmoles/105 cells • 5min
Control 31.8 4- 8.0 (7) 5.9 • 1.0 (11) Adenosine deaminase, 1 ~zg/ml 356.2 • 18.1 (11) 12.3 -t- 1.6 (11)
q- Insulin, 100 ~zU/ml 37.7 • 3.3 (11) 7.0 :k 1.0 (11) -k Nicotinic acid, 0.1 ~zl~ 11.9 • 5.2 (5) 6.7 :k 0.9 (11)
l ipolysis, b u t t he rise of cyclic A M P levels induced b y theophyl l ine was increased b y a cons tan t va lue of 6 to 8 pmoles pe r 10 5 cells t h roughou t the whole dose-response curve for theophyl l ine .
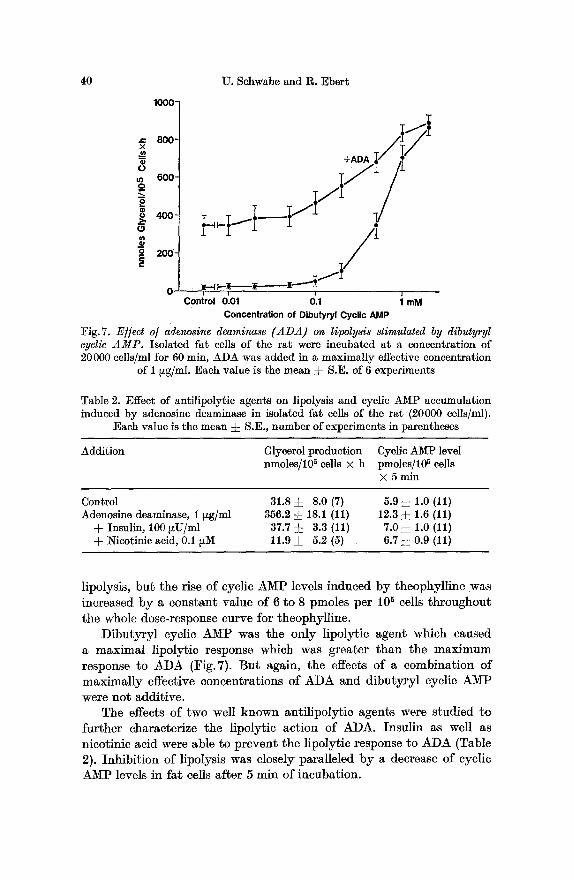
D i b u t y r y l cyclic A M P was the only l ipoly t ie agen t which caused a m a x i m a l lipoly~ie response which was grea te r t h a n the m a x i m u m response to A D A (Fig. 7). B u t again, t he effects of a combina t ion of m a x i m a l l y effective concent ra t ions of A D A and d i b u t y r y l cyclic A M P were no t addi t ive .
The effects of two well k n o w n an t i l ipo ly t ic agents were s tud ied to fur ther character ize the l ipolyt ic ac t ion of ADA. Insu l in as well as nicot inic acid were able to p r even t the l ipoly t ie response to A D A (Table 2). Inh ib i t i on of l ipolysis was closely para l le led b y a decrease of cyclic A M P levels in fa t cells a f te r 5 rain of incubat ion .

Effect of Adenosine Deamin~se on Fat Cells 41
Discussion
The results presented here indicate tha t the addition of ADA to incubation media of fat cells produces a strong lipolytic response ac- companied by a rise of cyclic AMP levels. Both effects were dose-depen- dent and already significant at a concentration of 0.01 ~g ADA per ml, thereby excluding non-specific effects of contaminants of the enzyme preparation. ADA catalyzes the deamination of adenosine to inosine. The increased lipolytie rate may result from a decrease of the con- centration of adenosine or from the formation of inosine and ammonia in the incubation medium. Inosine, however, has an inhibitory rather than a stimulant effect on lipolysis at concentrations of 10 to 100 ~M. Ammonium ions have been shown to be effective in replacing potassium ions at very high concentrations, but did not affect lipolysis at the con- centrations likely to have been present in our incubation media (Mosinger and Vaughan, 1967). However, in our study inosine may be responsible for the delay in the lipolytie response observed when concentrated cell suspensions were used. Therefore, our results suggest that ADA acts by decreasing adenosine in the incubation medium. The removal of adenosine from the incubation medium appears to release the fat cell from a preexisting inhibition. Thus, the present results with ADA support our previous conclusion that adenosine is released from isolated fat cells into the incubation medium (Schwabe et al., 1973).
Furthermore, our results indicate tha t adenosine acts on the outer surface of the fat cell. ADA, a large protein molecule, is not likely to penetrate the cell membrane. Therefore, ADA affects the adenosine concentration in the extracellular fluid only. This mode of action of ADA is in agreement with recent findings obtained with dipyridamole. The antflipolytic effect of adenosine was not antagonized by this com- pound, even though dipyridamole strongly inhibited the uptake of adenosine into fat cells (Ebert and Sehwabe, 1973; Fain, 1973b).
Adenosine deficiency produced by ADA unexpectedly elicited a lipolytie response which was of the same order of magnitude as tha t obtained with noradrenaline and theophylline. In previous studies we at tempted to eliminate the influence of the inhibitory factor in the incubation medium by dilution of the cell suspension (Schwabe and Ebert, 1972 ; Schwabe et al., 1973). Glycerol production by the unstimu- lated controls was enhanced by this procedure, but never reached the levels produced by lipo]ytic stimulants. Similarly effective means of reducing adenosine levels in the incubation medium are offered by the use of perifused fat cells. In this method the medium is continuously replaced by perifusion, thus allowing a rapid monitoring of changes in rates of lipolysis (Katocs et al., 1973). However, the basal rates of

42 U. Schwabe and R. Ebert
lipolysis obtained with this new model are low. This may be due to relatively large amounts of fat cells (2 ml packed cells) and small volumes of the perifusion medium used (10 ml/min), resulting in an incomplete removal of adenosine. These findings and our dilution experiment in- dicate that small residual amounts of adenosine are sufficient to keep lipolysis at low rates in unstimulated cells. On the other hand, high concentrations of adenosine only partially antagonized the lipolytic effects of noradrenaline (Fain et al., 1972; Sehwabe et al., 1973; Ebert and Schwabe, 1973).
Our findings further indicate that lipoly$ie stimulants as well as adenosine deficiency influence a common terminal process governing the lipolytic response. Maximally effective concentrations of several lipoly~ic compounds and ADA were not additive in their effects on glycerol production. Both types of lipolysis were accompanied by a rise in cyclic AMP even though the magnitude of cyclic AMP formation was different for different agents. This increase in cyclic AMP levels appears to be mediated by an activation of adenyl cyclase, since inhibition of phosphodiesterase by ADA can be excluded, as this enzyme is unlikely to penetrate cell membranes. I t has recently been demonstrated that adenyl cyelase of fat cell ghosts is inhibited by adenosine (Fain et al., 1972; McKenzie and B~r, 1973). The inhibitory effect of adenosine was reversed by the addition of ADA (MeKenzie and B~r, 1973). Therefore, all present data on the effects of adenosine support the hypothesis that the adenyl cyclase in intact fat cells is permanently kept in a restrained state by adenosine. Consequently, the withdrawal of adenosine is one of several possibilities to activate this enzyme. Whether this mechanism is of any importance for the physiological regulation of cyclic AMP levels is beyond the scope of the present discussion.
Another point deserves attention. ADA caused only a small increase of cyclic AMP levels as compared to other lipolytic agents. I t has been argued that only discrete alterations of cyclic AMP levels are necessary to induce a maximal lipolytic response. In 1968, when Butcher et al. did not find any detectable increase of cyclic AMP levels in fat cells after incubation with adrenaline, a discussion star~ed concerning the levels of cyclic :AMP required for triggering a hormonal response (for review see Fain, 1973a). In the presence of methylxanthines (Butcher et al., 1968) or at very low concentrations of fat cells (Schwabe and Ebert, 1972) a pronounced accumulation of cyclic AMP has been observ- ed in response to hormones. Although such high values may be rather impressive, it becomes more and more questionable whether they are relevant for the induction of the lipolytic process. Increasing concentra- tions of adenosine decreased cyclic AMP levels before affecting glycerol production (Fain et al., 1972 ; Schwabe et al., 1973). Lipolysis in response

Effect of Adenosine Deaminase on Fat Cells 43
to hormones was reduced only at considerably higher concentrations of the nucleoside. Similar results have recently been obtained with other potent antilipolytic agents, phenylisopropyl adenosine and prosta- glandin E 1 and E~ (Fain, 1973b; Stock and Prilop, 1974). Our findings, therefore, support the hypothesis tha t excessive cyclic AMP levels observed in the presence of theophylline or in diluted fat cell suspen- sions represent an overproduction of cyclic AMP because of the removal of the physiological inhibition (Fain et al., 1972; Fain, 1973a). Similar conclusions were drawn by Ho and Sutherland (1971) in their investiga- tion of the formation of a hormone antagonist in fat cells. They inter- preted a rapid fall in cyclic AMP level as an economic measure preventing excessive energy expenditure.
Finally, we should like to point out the striking similarity in the effects of ADA and theophylline. Both agents produced a relatively small increase in cyclic AMP levels, but a maximal lipolytic response in fat cells. In agreement with Fain (1973b) we found that , like theophyl- line, ADA acted synergistically with noradrenaline on cyclic AMP ac- cumulation. This was most evident after longer periods of incubation, when both agents almost identically prevented the fall of cyclic AMP levels. The well known potentiation of hormonal effects by methylxan- thines has been generally attr ibuted to their inhibitory effect on cyclic nucleotide phosphodiesterase (Butcher et al., 1968). However, theophyl- line failed to enhance the isoprenaline-induced rise of cyclic AMP levels in diluted fat cell suspensions (Schwabe and Ebert, 1972). A similar lack of synergism has been described recently for caffeine and noradrenaline in hamster fat cells (Hittelman et aI., 1973). For several other reasons we have previously proposed that, apart from inhibition of phosphodies- terase, theophylline may exert its lipolytic effect as an adenosine antago- nist on the cell membrane (Ebert and Sehwabe, 1973). The striking similarity of theophylline and ADA in their effects on cyclic AMP levels and lipolysis does not prove this hypothesis; however, these findings add a further stimulus to the search for a possible common site of action of theophylline and adenosine.
Acknowledgements. This study was supported by the Deutsche Forschungs- gemeinschaft (Sch 83/6). We wish to thank Miss Helga Hannemarm and Mrs. Anna-Carmen Bauer for their excellent technical assistance. We are indebted to Drs. J. N. Fain and K. Stock for communicating their findings prior to publication.
References Butcher, R. W., Baird, C. E., Sutherland, E. W. : Effects of lipolytie and antilipoly-
tic substances on adenosine 3',5'-monophosphate levels in isolated fat cells. J . biol. Chem. 243, 1705--1712 (1968)
Chen, R. F. : Removal of fa t ty acids from serum albumin by charcoal treatment. J. biol. Chem. 24~, 173--181 (1967)

44 U. Schwabe and R. Ebert
Ebert, R., Schwabe, U. : Studies on the antilipolytic effect of adenosine and related compounds in isolated fag cells. Naunyn-Schmiedeberg's Arch. Pharmacol. 278, 247--259 (1973)
Fain, d. N. : Biochemical aspects of drug and hormone action on adipose tissue. Pharmacol. Rev. 25, 67--118 (1973a)
Fain, d. N.: Inhibition of adenosine cyclic 3',5'-monophosphate accumulation in fat cells by adenosine, N6-(phenylisopropyl) adenosine and related compounds. ~Iolec. Pharmacol. 9, 595--604 (1973b)
Fain, d.N., Pointer, R. H., Ward, W.F. : Effects of adenosine nucleosides on adenylate cyclase, phosphodiesterase, cyclic adenosine monophosphate accumu- lation and lipolysis in fat cells, d. biol. Chem. 247, 6866--6872 (1972)
Friedkin, M., Kalckar, H.: Nucleoside phosphorylases. In: P.D. Boyer, H.A. Lardy and K. 5Iyrb~ek (eds.): The enzymes, vol. 5, pp. 237--255. New York: Academic Press 1961
Gilman, A. G. : A protein binding assay for adenosine 3',5'-cyclic monophosphate. Prec. nat. Acad. Sci. (Wash.) 67, 305--312 (1970)
Hittelman, K.J . , Wu, C. F., Butcher, R.W.: Control of cyclic AMP levels in isolated fat cells from hamsters. Biochim. biophy. Acta (Amst.) 804, 188--196 (1973)
tic, R. J., Sutherland, E. W. : Formation and release of a hormone antagonist by rat adipocytes. J. biol. Chem. 246, 6822--6827 (1971)
Katocs, A. S., Largis, E. E., Allen, D. 0., Ashmore, J.: Perifused fat cells. Effect of lipolytic agents. J. biol. Chem. 248, 5089--5094 (1973)
Lambert, M., Neish, A. C. : Rapid method for estimation of glycerol in fermentation solutions. Canad. J. Res., Sect. B 28, 83--89 (1950)
McKenzie, S. G., Bit, H. P.: On the mechanism of adenyl cyclase inhibition by adenosine. Canad. d. Physiol. Pharmacol. 51, 190--196 (1973)
Mosinger, B., Vaughan, M.: Effects of electrolytes on epinephrine stimulated lipolysis in adipose tissue in vitro. Biochim. biophys. Acta (Amst.) 144, 556--568 (1967)
Nash, T. : The colorimetric estimation of formaldehyde by means of the Iiantzsch reaction. Biochem. J. 55, 416--421 (1953)
Rodbell, M.: Metabolism of isolated fat cells. I. Effect of hormones on glucose metabolism and lipolysis. J. biol. Chem. 289, 375--380 (1964)
Schwabe, U., Ebert, R. : Different effects of lipolytic hormones and phosphodies- terase inhibitors on cyclic 3',5'-AMP levels in isolated fat cells. Naunyn- Schmicdeberg's Arch. Pharmacol. 274, 287--298 (1972)
Schwabe, U., Ebert, R., Erbler, H. C. : Adenosine release from isolated fat cells and its significance for the effects of hormones on cyclic 3',5'-AMP levels and lipoly- sis. Naunyn-Schmiedeberg's Arch. Pharmaeol. 276, 133--148 (1973)
Stock, K., Prilop, M. : Dissociation of eatecholamine-induced formation of adeno- sine 3',5'-monophosphate and release of glycerol in fat cells by prostaglandin E i, E~ and iV~-phenylisopropyladenosine. Naunyn-Sehmiedeberg's Arch. Pharmaeol. 282, 15--31 (1974)
U. Schwabe Medizinische I-Ioohschule ttannover Institut fiir Pharmakologie D-3000 Hannover-Kleefeld Karl Wiechert-Allee 9 Federal Republic of Germany







![KB Id - UNT Digital Library/67531/metadc332161/... · 1-[bis(hydroxymethyl)amino]-3-tris(hydroxymethyl)propane adenosine 3',5'-monophosphate adenosine 31,5'-monophosphate dependent](https://img.pdfslide.net/doc/110x75/60bf6195247f5a484a422257/kb-id-unt-digital-library-67531metadc332161-1-bishydroxymethylamino-3-trishydroxymethylpropane.jpg)





















